Structural Analysis of Gluco-Oligosaccharides Produced by Leuconostoc lactis and Their Prebiotic Effect


Abstract
:1. Introduction
2. Results
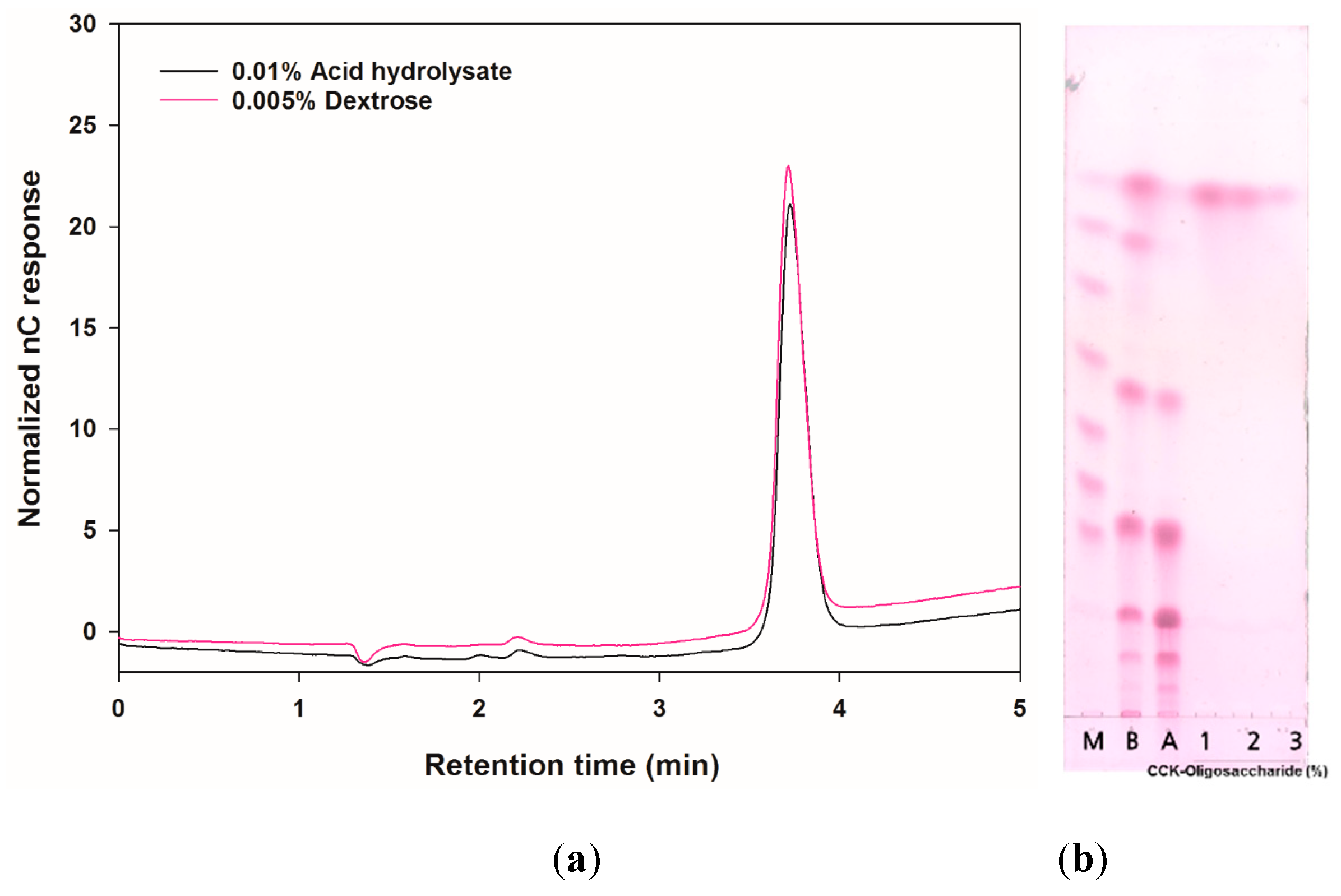
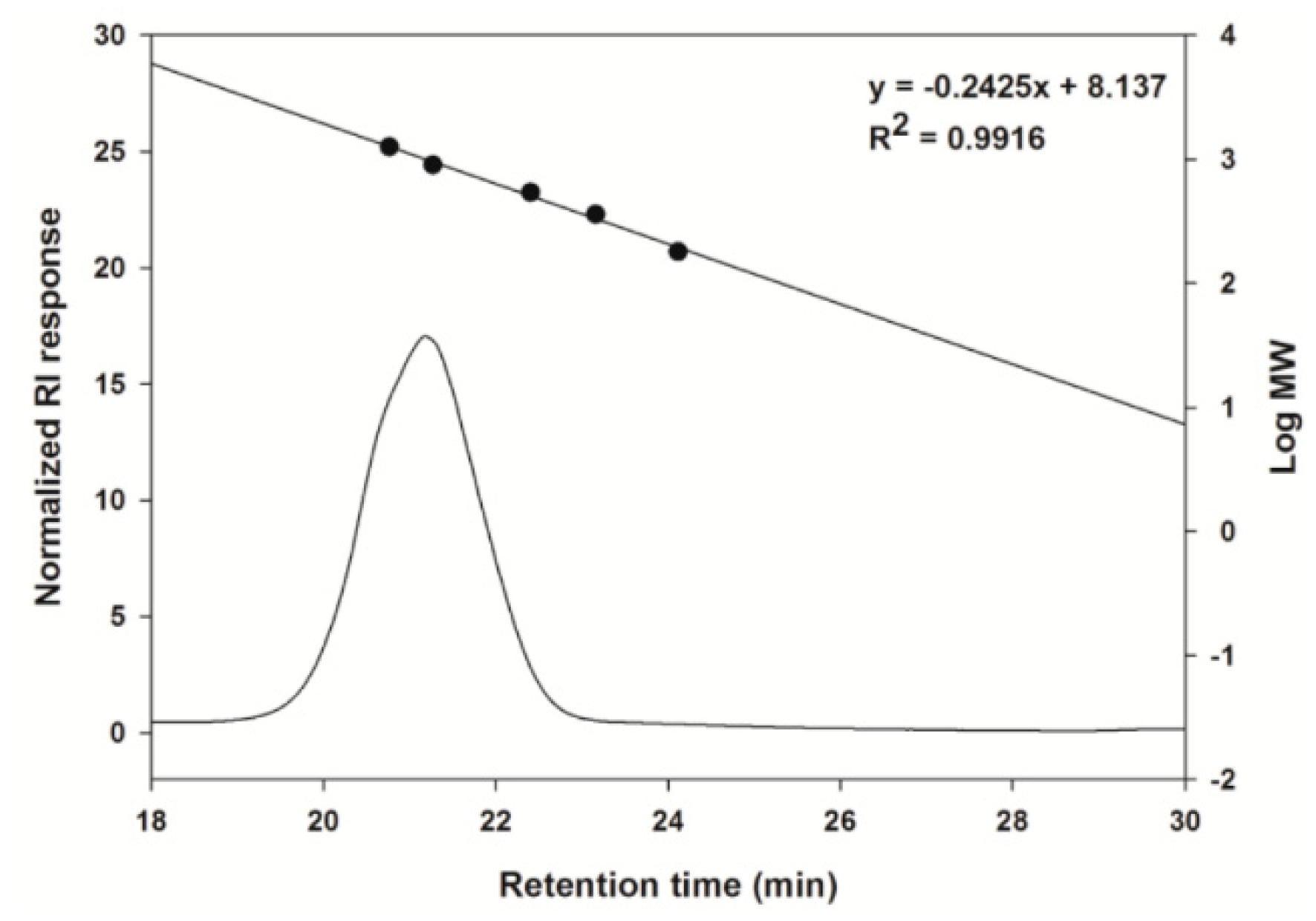
2.1. Analysis of the Carbohydrate Composition of the CCK-Oligosaccharides
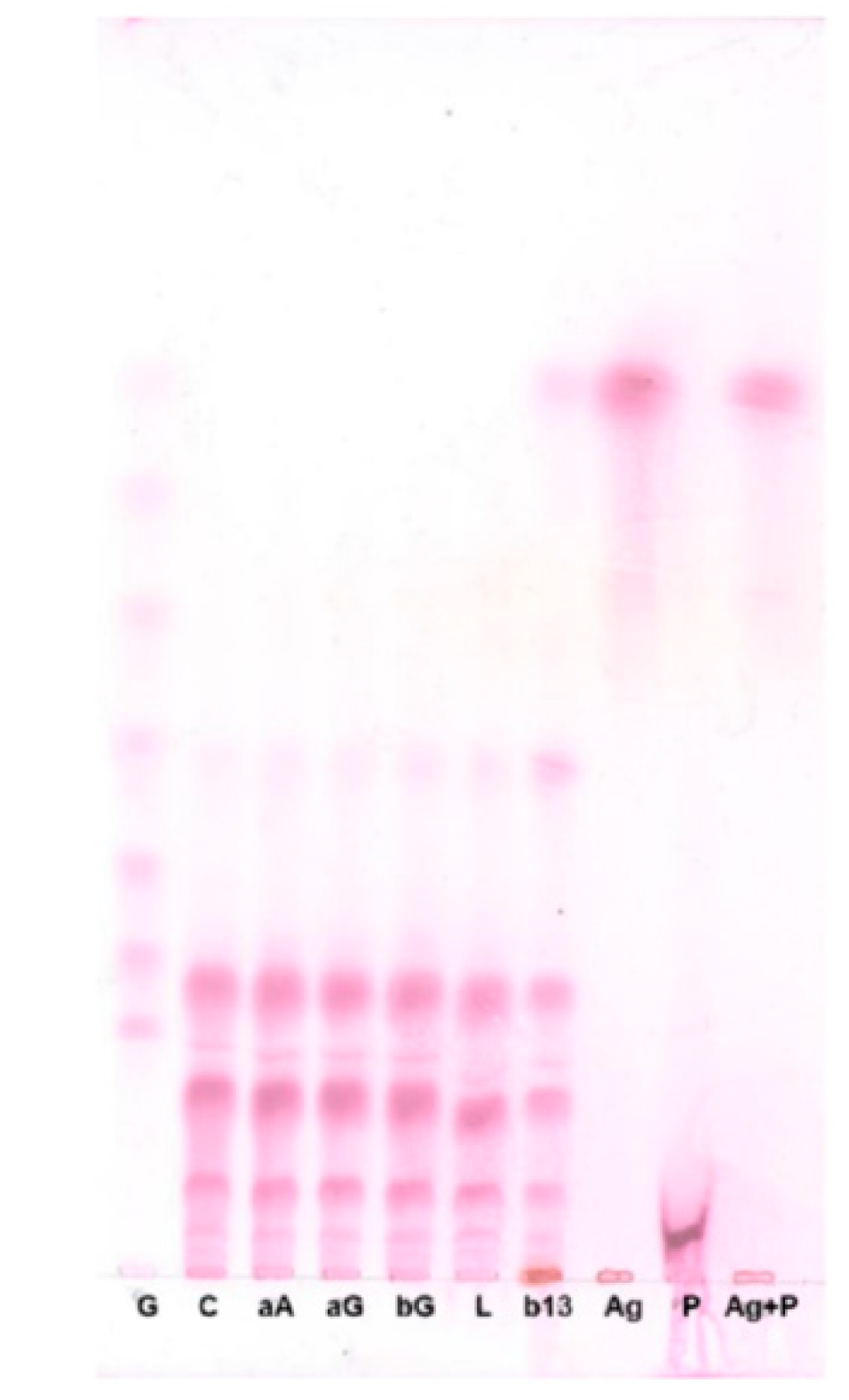
2.2. Determination of the Types of Glycosidic Bonds
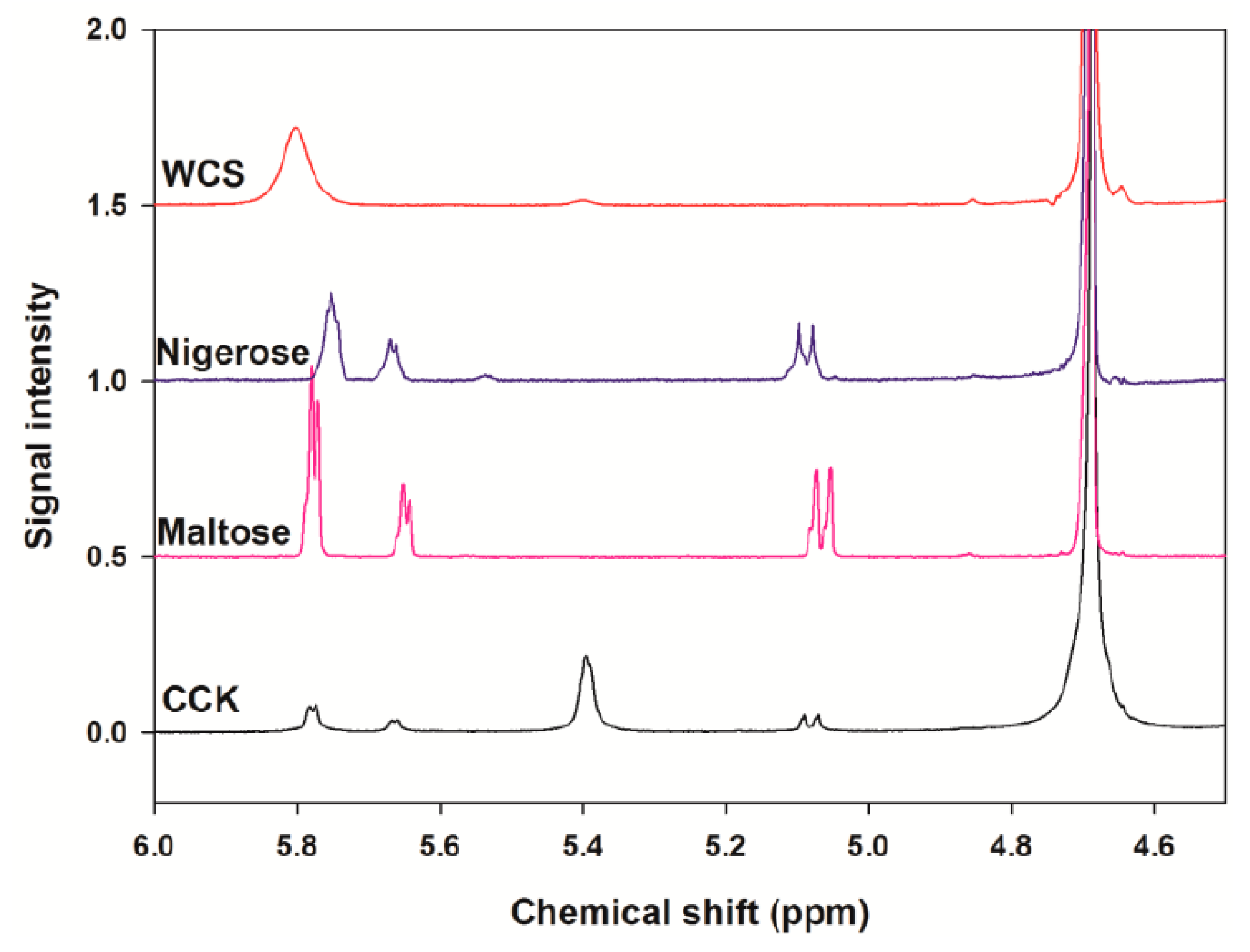
2.3. Analysis of the Linkage Ratio by 1H-NMR Spectroscopy
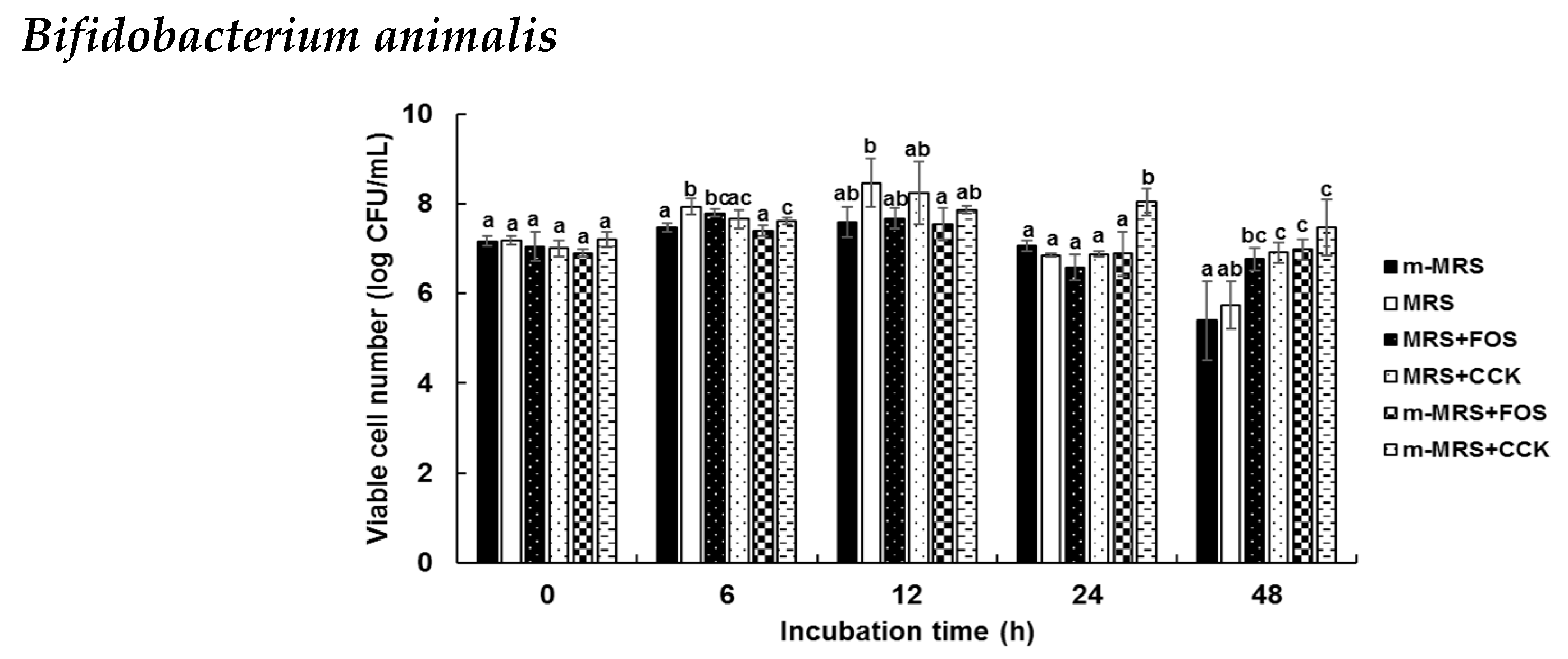
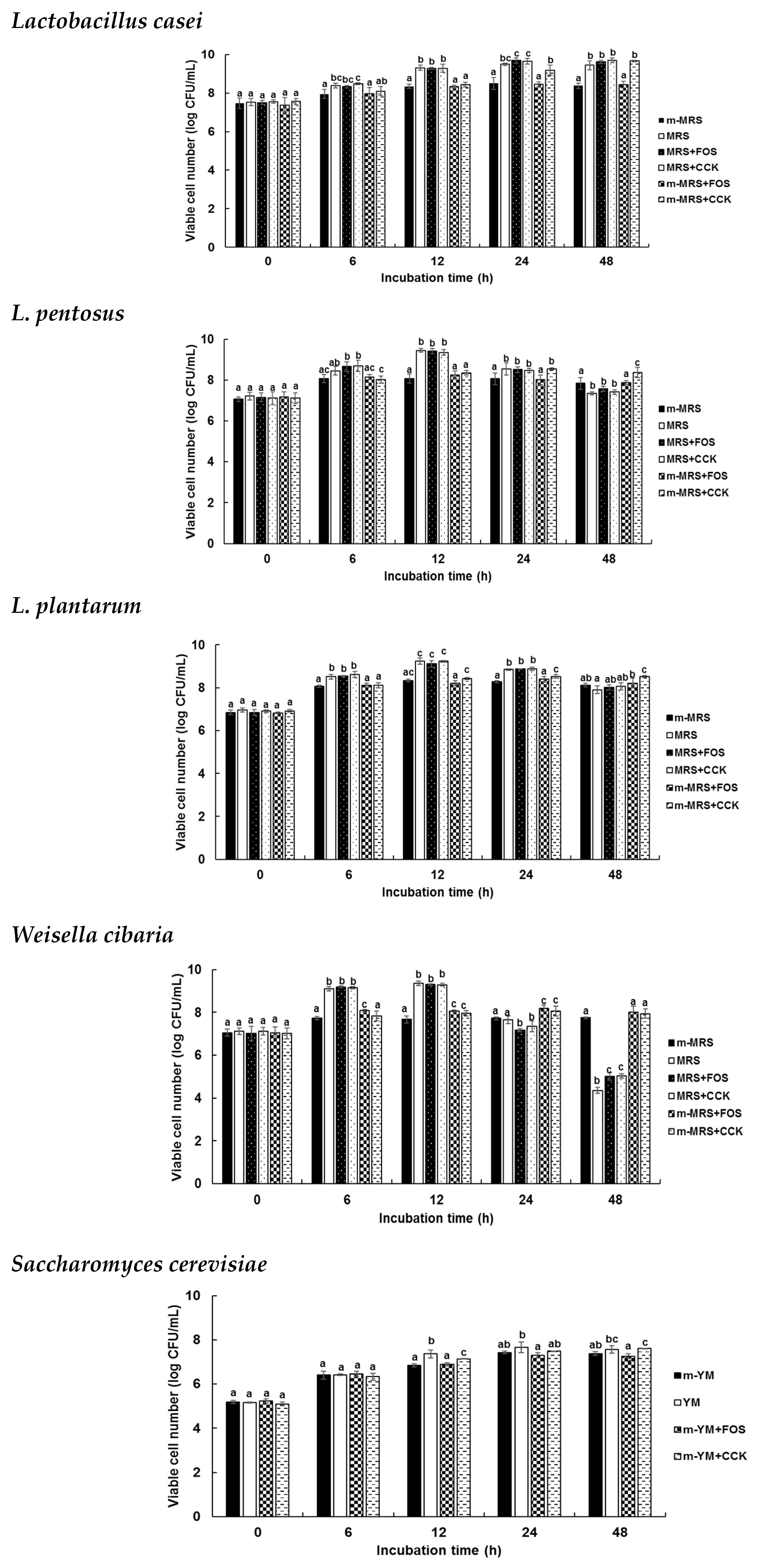
2.4. Prebiotic Effects of the CCK-Oligosaccharides
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Determination of Oligosaccharide Structure
4.2.1. Purification of Oligosaccharides
4.2.2. Thin-Layer Chromatography (TLC)
4.2.3. HPAEC-PAD Analysis
4.2.4. Size Exclusion HPLC Analysis
4.2.5. Analysis of Sugar Composition
4.2.6. Composition of Glycosidic Bonds by Enzymatic Analysis
4.2.7. Analysis of Linkage Ratio by Proton Nuclear Magnetic Resonance (1H-NMR) Spectroscopy
4.3. Prebiotic Effects
4.3.1. Selection of Probiotic Strains
4.3.2. Growth Conditions of Probiotic Strains
4.3.3. Microbiological Analysis
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Coutinho, P.M.; Deleury, E.; Davies, G.J.; Henrissat, B. An evolving hierarchical family classification for glycosyltransferase. J. Mol. Biol. 2003, 328, 307–317. [Google Scholar] [CrossRef]
- Lairson, L.L.; Henrissat, B.; Davies, G.J.; Withers, S.G. Glycosyltransferase: Structures, functions, and mechanisms. Ann. Rev. Biochem. 2008, 77, 521–555. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Hanh, N.T.T.; Cho, J.Y.; Kim, J.Y.; Moon, Y.H.; Yeom, S.C.; Kim, G.J.; Kim, D. Glucooligosaccharide production by Leuconostoc mesenteroides fermentation with efficient pH control, using a calcium hydroxide–sucrose solution. Biotechnol. Bioprocess Eng. 2016, 21, 39–45. [Google Scholar] [CrossRef]
- Monsan, P.F.; Ouarné, F. Chapter 10. Oligosaccharides derived from sucrose. In Prebiotics and Probiotics Science and Technology; Charalampopoulos, D., Rastall, R.A., Eds.; Springer Science+Business Media: New York, NY, USA, 2009; pp. 293–336. [Google Scholar]
- Naessens, M.; Cerdobbel, A.; Soetaert, W.; Vandamme, E.J. Leuconostoc dextransucrase and dextran: Production, properties and applications. J. Chem. Technol. Biotechnol. 2005, 80, 845–860. [Google Scholar] [CrossRef]
- Heincke, K.; Demuth, B.; Jördening, H.J.; Buchholz, K. Kinetics of the dextransucrase acceptor reaction with maltose—experimental results and modeling. Enzyme Microb. Technol. 1999, 24, 523–534. [Google Scholar] [CrossRef]
- Leemhuis, H.; Pijning, T.; Dobruchowska, J.M.; van Leeuwen, S.S.; Kralj, S.; Dijkstra, B.W.; Dijkhuizen, L. Glucansucrases: Three–dimensional structures, reactions, mechanism, α–glucan analysis and their implications in biotechnology and food applications. J. Biotechnol. 2013, 163, 250–272. [Google Scholar] [CrossRef]
- Monchois, V.; Willemot, R. –M.; Monsan, P. Glucansucrases: Mechanism of action and structure function relationships. FEMS Microbiol. Rev. 1999, 23, 131–151. [Google Scholar] [CrossRef]
- Bindels, L.B.; Delzenne, N.M.; Cani, P.D.; Walter, J. Towards a more comprehensive concept for prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 303–310. [Google Scholar] [CrossRef]
- de Vrese, M.; Schrezenmeir, J. Probiotics, prebiotics, and synbiotics. Adv. Biochem. Eng. Biotechnol. 2008, 111, 1–66. [Google Scholar] [CrossRef]
- Roberfroid, M. Prebiotics: The concept revisited. J. Nutr. 2007, 137, 830–837. [Google Scholar] [CrossRef]
- Costabile, A.; Fava, F.; Röytiö, H.; Forssten, S.D.; Olli, K.; Klievink, J.; Rowland, I.R.; Ouwehand, A.C.; Rastall, R.A.; Gibson, G.R.; et al. Impact of polydextrose on the faecal microbiota: A double–blind, crossover, placebo–controlled feeding study in healthy human subjects. Br. J. Nutr. 2012, 108, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Putaala, H.; Mäkivuokko, H.; Tiihonen, K.; Rautonen, N. Simulated colon fiber metabolome regulates genes involved in cell cycle, apoptosis, and energy metabolism in human colon cancer cells. Mol. Cell. Biochem. 2011, 357, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Park, G.-G.; Jang, J.-K.; Park, Y.-S. Optimization of oligosaccharide production from Leuconostoc lactis using a response surface methodology and the immunostimulating effects of these oligosaccharides on macrophage cells. Molecules 2018, 23, 2118. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.H. Production of glucooligosaccharides and mannitol from Leuconostoc mesenteroides B–742 fermentation and its separation from byproducts. J. Microbiol. Biotechnol. 2006, 16, 325–329. [Google Scholar]
- Finke, B.; Stahl, B.; Pfenninger, A.; Karas, M.; Daniel, H.; Sawatzki, G. Analysis of high-molecular-weight oligosaccharides from human milk by liquid chromatography and MALDI-MS. Anal. Chem. 1999, 71, 3755–3762. [Google Scholar] [CrossRef] [PubMed]
- Koepsell, H.J.; Tsuchiya, H.M.; Hellman, N.N.; Kazenko, A.; Hoffman, C.A.; Sharpe, E.S.; Jackson, R.W. Enzymatic synthesis of dextran, acceptor specificity and chain initiation. J. Biol. Chem. 1953, 200, 793–801. [Google Scholar]
- Gidley, M.J. Quantification of the structural features of starch polysaccharides by NMR spectroscopy. Carbohydr. Res. 1985, 139, 85–93. [Google Scholar] [CrossRef]
- Dobruchowska, J.M.; Gerwig, G.J.; Kralj, S.; Grijpstra, P.; Leemhuis, H.; Dijkhuizen, L.; Kamerling, J.P. Structural characterization of linear isomalto-/malto-oligomer products synthesized by the novel GTFB 4,6-α-glucanostransferase enzyme from Lactobacillus reuteri 121. Glycobiology 2012, 22, 517–528. [Google Scholar] [CrossRef]
- Côté, G.L. Alternan. In Biopolymers; Steinbüchel, A., Ed.; Wiley-VCH: Weinheim, Germany, 2002; Volume 5, pp. 323–350. [Google Scholar]
- Kothari, D.; Goyal, A. Gentio–oligosaccharides from Leuconostoc mesenteroides NRRL B–1426 dextransucrase as prebiotics and as a supplement for functional foods with anti–cancer properties. Food Funct. 2015, 6, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Sanz, M.; Côté, G.; Gibson, G.; Rastall, R. Prebiotic properties of alternansucrase maltose–acceptor oligosaccharides. J. Agric. Food Chem. 2005, 53, 5911–5916. [Google Scholar] [CrossRef]
- Hayek, S.A.; Ibrahim, S.A. Current limitations and challenges with lactic acid bacteria: A review. Food Nutr. Sci. 2013, 73–87. [Google Scholar] [CrossRef]
- Abelovska, L.; Bujdos, M.; Kubova, J.; Petrezselyove, S.; Nosek, J.; Tomaska, L. Comparison of element levels in minimal and complex yeast media. Can. J. Microbiol. 2007, 53, 533–535. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Park, Y.-S. Oligosaccharide production by Leuconostoc lactis CCK940 which has glucansucrase activity. Food Eng. Prog. 2017, 21, 383–390. [Google Scholar] [CrossRef]
- Astolfi, M.L.; Protano, C.; Schiavi, E.; Marconi, E.; Capobianco, D.; Massimi, L.; Ristorini, M.; Baldassarre, M.E.; Laforgia, N.; Vitali, M.; et al. A prophylactic multi-strain probiotic treatment to reduce the absorption of toxic elements: In-vitro study and biomonitoring of breast milk and infant stools. Environ. Int. 2019, 130, 104818. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| α-1,4 Linkages (%) 1 | α-1,6 Linkages (%) | Ratio of α-1,4 to α-1,6 | |
|---|---|---|---|
| WCS | 96.2 ± 0.0 | 3.8 ± 0.0 | 25.5 ± 0.3 |
| CCK-oligosaccharides | 22.4 ± 1.2 | 77.6 ± 1.2 | 0.3 ± 0.0 |
| Ingredient | Amount (g/L) |
|---|---|
| Proteose peptone No. 3 | 10.0 |
| Beef extract | 10.0 |
| Yeast extract | 5.0 |
| Polysorbate 80 | 1.0 |
| Ammonium citrate | 2.0 |
| Sodium acetate | 5.0 |
| Magnesium sulfate | 0.1 |
| Manganese sulfate | 0.05 |
| Dipotassium phosphate | 2.0 |
| Ingredient | Amount (g/L) |
|---|---|
| Yeast extract | 3.0 |
| Malt extract | 3.0 |
| Peptone | 5.0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Park, J.; Jang, J.-K.; Lee, B.-H.; Park, Y.-S. Structural Analysis of Gluco-Oligosaccharides Produced by Leuconostoc lactis and Their Prebiotic Effect. Molecules 2019, 24, 3998. https://doi.org/10.3390/molecules24213998
Lee S, Park J, Jang J-K, Lee B-H, Park Y-S. Structural Analysis of Gluco-Oligosaccharides Produced by Leuconostoc lactis and Their Prebiotic Effect. Molecules. 2019; 24(21):3998. https://doi.org/10.3390/molecules24213998
Chicago/Turabian StyleLee, Sulhee, Jisun Park, Jae-Kweon Jang, Byung-Hoo Lee, and Young-Seo Park. 2019. "Structural Analysis of Gluco-Oligosaccharides Produced by Leuconostoc lactis and Their Prebiotic Effect" Molecules 24, no. 21: 3998. https://doi.org/10.3390/molecules24213998