
Ivalin Induces Mitochondria-Mediated Apoptosis Associated with the NF-κB Activation in Human Hepatocellular Carcinoma SMMC-7721 Cells

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
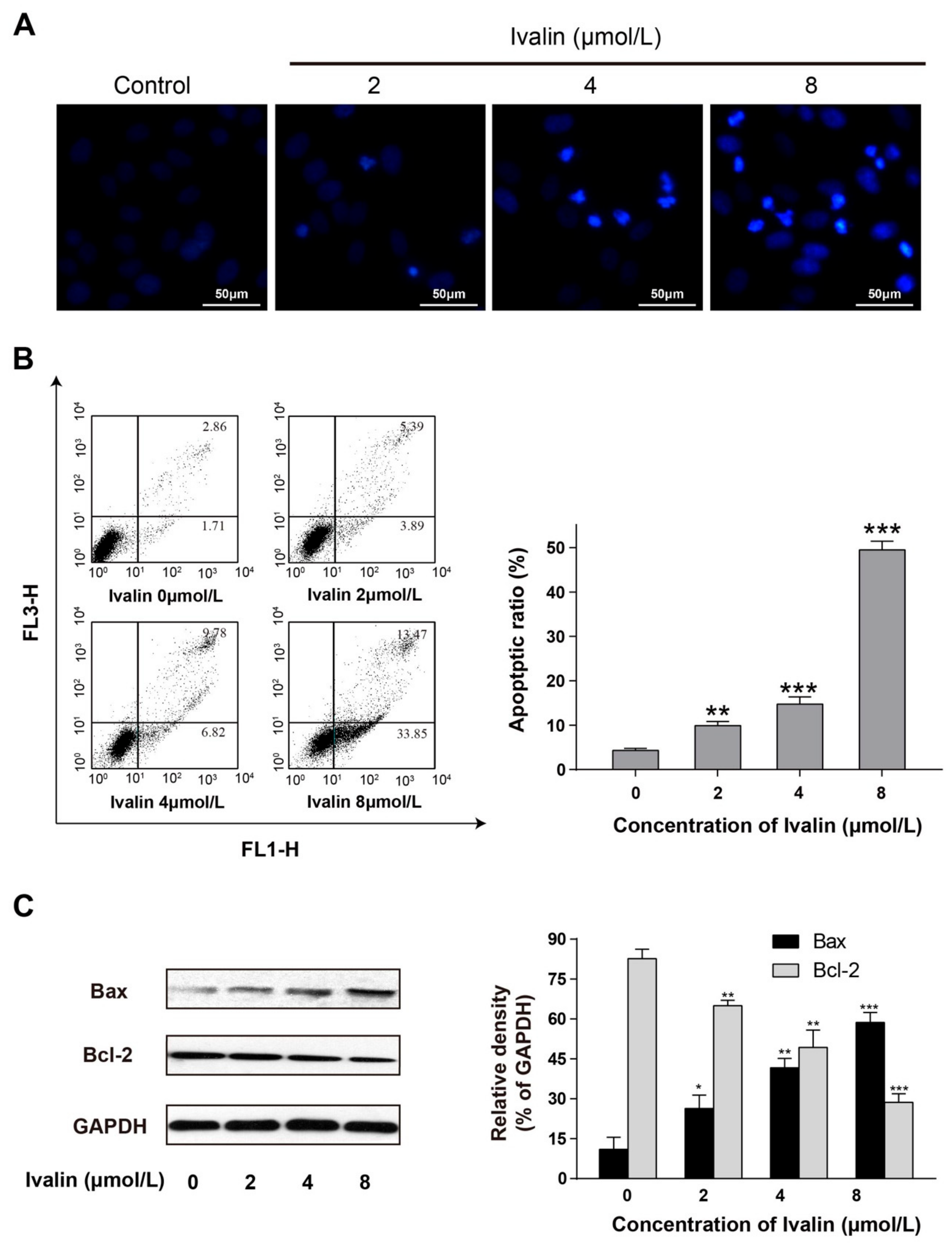
2.1. Apoptotic Effect of Ivalin
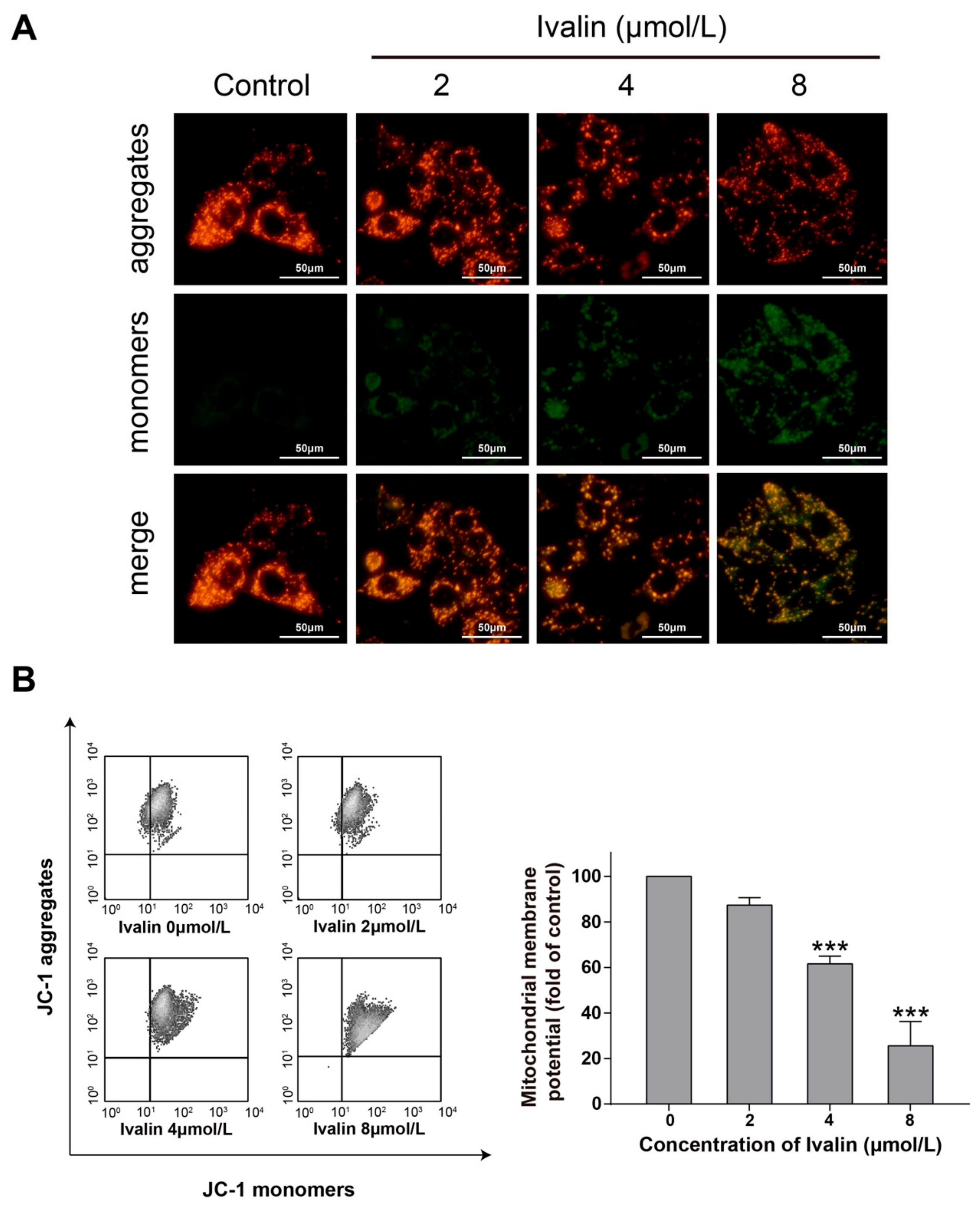
2.2. Ivalin Triggered the Loss of Mitochondrial Membrane Potential (MMP) in SMMC-7721 Cells
2.3. The Generation of ROS Appeared in Ivalin-Treated Cells
2.4. Ivalin Modulated the Protein and mRNA Levels of NF-κB, IκB, and p53
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Human Hepatocellular Carcinoma and Cell Culture
4.3. DAPI Staining
4.4. Mitochondrial Membrane Potential
4.5. Apoptosis Detection
4.6. Measurement of Intracellular ROS Levels
4.7. Western Blot Analysis
4.8. Real-Time PCR Analysis
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. Ca-A. Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, E.K.; Vauthey, J.N. Focus on treatment of large hepatocellular carcinoma. Ann. Surg. Oncol. 2004, 11, 1035–1036. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.-D.; Yao, C.-J.; Chow, J.-M.; Chang, C.-L.; Hwang, P.-A.; Chuang, S.-E.; Whang-Peng, J.; Lai, G.-M. Fucoidan Elevates MicroRNA-29b to Regulate DNMT3B-MTSS1 Axis and Inhibit EMT in Human Hepatocellular Carcinoma Cells. Mar. Drugs 2015, 13, 6099–6116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mor, E.; Kaspa, R.T.; Sheiner, P.; Schwartz, M. Treatment of hepatocellular carcinoma associated with cirrhosis in the era of liver transplantation. Ann. Intern. Med. 1998, 129, 643–653. [Google Scholar] [CrossRef]
- Li, X.; Zhao, Y.; Wu, W.K.K.; Liu, S.; Cui, M.; Lou, H. Solamargine induces apoptosis associated with p53 transcription-dependent and transcription-independent pathways in human osteosarcoma U2OS cells. Life Sci. 2011, 88, 314–321. [Google Scholar] [CrossRef]
- Wiseman, L.R.; Markham, A. Irinotecan—A review of its pharmacological properties and clinical efficacy in the management of advanced colorectal cancer. Drugs 1996, 52, 606–623. [Google Scholar] [CrossRef]
- Perez, E.A. Microtubule inhibitors: Differentiating tubulin-inhibiting agents based on mechanisms of action, clinical activity, and resistance. Mol. Cancer Ther. 2009, 8, 2086–2095. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.-X.; Shi, Y.-P.; Jia, Z.-J. Eudesmane sesquiterpenoids from the Asteraceae family. Nat. Prod. Rep. 2006, 23, 699–734. [Google Scholar] [CrossRef]
- Da Costa, F.B.; Terfloth, L.; Gasteiger, J. Sesquiterpene lactone-based classification of three Asteraceae tribes: A study based on self-organizing neural networks applied to chemo systematics. Phytochemistry 2005, 66, 345–353. [Google Scholar] [CrossRef]
- Zheng, B.; Wu, L.; Ma, L.; Liu, S.; Li, L.; Xie, W.; Li, X. Telekin Induces Apoptosis Associated with the Mitochondria-Mediated Pathway in Human Hepatocellular Carcinoma Cells. Biol. Pharm. Bull. 2013, 36, 1118–1125. [Google Scholar] [CrossRef] [Green Version]
- Cui, M.; Zhang, Y.; Liu, S.; Xie, W.; Ji, M.; Lou, H.; Li, X. 1-Oxoeudesnn-11(13)-ene-12,8 alpha-lactone-induced Apoptosis via ROS Generation and Mitochondria Activation in MCF-7 Cells. Arch. Pharmacal Res. 2011, 34, 1323–1329. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.-D.; Wang, X.-R.; Ma, L.-S.; Li, X.; Row, K.-H. Sesquiterpenoids from Carpesium divaricatum and their cytotoxic activity. Fitoterapia 2012, 83, 1351–1355. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Lin, S.; Zhang, C.; Ma, J.; Han, Z.; Jia, F.; Xie, W.; Li, X. The Novel Nature Microtubule Inhibitor Ivalin Induces G2/M Arrest and Apoptosis in Human Hepatocellular Carcinoma SMMC-7721 Cells In Vitro. Medicina 2019, 55, 470. [Google Scholar] [CrossRef] [PubMed]
- Bonvin, C.; Guillon, A.; Van Bemmelen, M.X.; Gerwins, P.; Johnson, G.L.; Widmann, C. Role of the amino-terminal domains of MEKKs in the activation of NF kappa B and MAPK pathways and in the regulation of cell proliferation and apoptosis. Cell. Signal. 2002, 14, 123–131. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, Y.; Wu, D.; Tashiro, S.-i.; Onodera, S.; Ikejima, T. NF-kappa b facilitates oridonin-induced apoptosis and autophagy in HT1080 cells through a p53-mediated pathway. Arch. Biochem. Biophys. 2009, 489, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.; Gao, B.; Gao, Y.; Yang, X.; Cheng, X.; Ou, Y. Phycocyanin Inhibits Tumorigenic Potential of Pancreatic Cancer Cells: Role of Apoptosis and Autophagy. Sci. Rep. 2016, 6, 34564. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, P.D.S.; Ornellas, A.A.; Campos, M.R.M.; Scheiner, M.A.M.; Fiedler, W.; Alves, G. Bax and Bcl-2 imbalance and HPB infection in penile tumors and adjacent tissues. Prog. En Urol. 2004, 14, 353–359. [Google Scholar]
- Cheng, E. Molecular Control of Mitochondrial Apoptosis by the BCL-2 Family. Blood 2009, 114, 1577–1578. [Google Scholar] [CrossRef]
- Ye, J.; Ma, J.; Liu, C.; Huang, J.; Wang, L.; Zhong, X. A novel iron(II) phenanthroline complex exhibits anticancer activity against TFR1-overexpressing esophageal squamous cell carcinoma cells through ROS accumulation and DNA damage. Biochem. Pharmacol. 2019, 166, 93–107. [Google Scholar] [CrossRef]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Thayyullathil, F. Reactive oxygen species and cancer paradox: To promote or to suppress? Free Radic. Biol. Med. 2017, 104, 144–164. [Google Scholar] [CrossRef]
- Fujioka, S.; Schmidt, C.; Sclabas, G.M.; Li, Z.K.; Pelicano, H.; Peng, B.; Yao, A.; Niu, J.G.; Zhang, W.; Evans, D.B.; et al. Stabilization of p53 is a novel mechanism for proapoptotic function of NF-kappa B. J. Biol. Chem. 2004, 279, 27549–27559. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G.; Galluzzi, L.; Brenner, C. Mitochondrial membrane permeabilization in cell death. Physiol. Rev. 2007, 87, 99–163. [Google Scholar] [CrossRef] [PubMed]
- Czabotar, P.E.; Lessene, G.; Strasser, A.; Adams, J.M. Control of apoptosis by the BCL-2 protein family: Implications for physiology and therapy. Nat. Rev. Mol. Cell Biol. 2014, 15, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.C.; Zong, W.X.; Cheng, E.H.Y.; Lindsten, T.; Panoutsakopoulou, V.; Ross, A.J.; Roth, K.A.; MacCregor, G.R.; Thompson, C.B.; Korsmeyer, S.J. Proapoptotic BAX and BAK: A requisite gateway to mitochondrial dysfunction and death. Science 2001, 292, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Sen, R.; Baltimore, D. Inducibility of kappa immunoglobulin enhancer-binding protein Nf-kappa B by a posttranslational mechanism. Cell 1986, 47, 921–928. [Google Scholar] [CrossRef]
- Lee, C.H.; Jeon, Y.-T.; Kim, S.-H.; Song, Y.-S. NF-kappa B as a potential molecular target for cancer therapy. Biofactors 2007, 29, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Bernard, D.; Monte, D.; Vandenbunder, B.; Abbadie, C. The c-Rel transcription factor can both induce and inhibit apoptosis in the same cells via the upregulation of MnSOD. Oncogene 2002, 21, 4392–4402. [Google Scholar] [CrossRef] [Green Version]
- Kaltschmidt, B.; Kaltschmidt, C.; Hofmann, T.G.; Hehner, S.P.; Droge, W.; Schmitz, M.L. The pro- or anti-apoptotic function of NF-kappa B is determined by the nature of the apoptotic stimulus. Eur. J. Biochem. 2000, 267, 3828–3835. [Google Scholar] [CrossRef]
- Sheehy, A.M.; Schlissel, M.S. Overexpression of RelA causes G1 arrest and apoptosis in a pro-B cell line. J. Biol. Chem. 1999, 274, 8708–8716. [Google Scholar] [CrossRef]
- Tarabin, V.; Schwaninger, M. The role of NF-kappa B in 6-hydroxydopamine- and TNF alpha-induced apoptosis of PC12 cells. Naunyn-Schmiedebergs Arch. Pharmacol. 2004, 369, 563–569. [Google Scholar] [CrossRef]
- Yeh, P.Y.; Chuang, S.E.; Yeh, K.H.; Song, Y.C.; Ea, C.K.; Cheng, A.L. Increase of the resistance of human cervical carcinoma cells to cisplatin by inhibition of the MEK to ERK signaling pathway partly via enhancement of anticancer drug-induced NF kappa B activation. Biochem. Pharmacol. 2002, 63, 1423–1430. [Google Scholar] [CrossRef]
- Liu, S.; Wu, D.; Li, L.; Sun, X.; Xie, W.; Li, X. NF-kappa B activation was involved in reactive oxygen species-mediated apoptosis and autophagy in 1-oxoeudesm-11(13)-eno-12,8 alpha-lactone-treated human lung cancer cells. Arch. Pharmacal Res. 2014, 37, 1039–1052. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhou, Y.; Shen, P. NF-kappa B and Its Regulation on the Immune System. Cell. Mol. Immunol. 2004, 1, 343–350. [Google Scholar] [PubMed]
- Perkins, N.D. Integrating cell-signalling pathways with NF-kappa B and IKK function. Nat. Rev. Mol. Cell Biol. 2007, 8, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.-L.; Guo, C.; Qi, J.; Ma, J.-H.; Liu, F.-Y.; Lin, S.-Q.; Zhang, C.-Y.; Xie, W.-D.; Zhuang, J.-J.; Li, X. Protective effects of 3-angeloyloxy-8, 10-dihydroxyeremophila-7(11)-en-12, 8-lactone on paraquat-induced oxidative injury in SH-SY5Y cells. J. Asian Nat. Prod. Res. 2019, 21, 364–376. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Zhang, C.; Liu, F.; Ma, J.; Jia, F.; Han, Z.; Xie, W.; Li, X. Actinomycin V Inhibits Migration and Invasion via Suppressing Snail/Slug-Mediated Epithelial-Mesenchymal Transition Progression in Human Breast Cancer MDA-MB-231 Cells In Vitro. Mar. Drugs 2019, 17, 305. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Z.; Liu, F.-y.; Lin, S.-q.; Zhang, C.-y.; Ma, J.-h.; Guo, C.; Jia, F.-j.; Zhang, Q.; Xie, W.-d.; Li, X. Ivalin Induces Mitochondria-Mediated Apoptosis Associated with the NF-κB Activation in Human Hepatocellular Carcinoma SMMC-7721 Cells. Molecules 2019, 24, 3809. https://doi.org/10.3390/molecules24203809
Han Z, Liu F-y, Lin S-q, Zhang C-y, Ma J-h, Guo C, Jia F-j, Zhang Q, Xie W-d, Li X. Ivalin Induces Mitochondria-Mediated Apoptosis Associated with the NF-κB Activation in Human Hepatocellular Carcinoma SMMC-7721 Cells. Molecules. 2019; 24(20):3809. https://doi.org/10.3390/molecules24203809
Chicago/Turabian StyleHan, Zhuo, Fang-yuan Liu, Shi-qi Lin, Cai-yun Zhang, Jia-hui Ma, Chao Guo, Fu-juan Jia, Qian Zhang, Wei-dong Xie, and Xia Li. 2019. "Ivalin Induces Mitochondria-Mediated Apoptosis Associated with the NF-κB Activation in Human Hepatocellular Carcinoma SMMC-7721 Cells" Molecules 24, no. 20: 3809. https://doi.org/10.3390/molecules24203809