Identification of Potential Auxin-Responsive Small Signaling Peptides through a Peptidomics Approach in Arabidopsis thaliana


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Optimization of Peptide Extraction
2.1.1. Peptides Extracted by Different Lysate Buffer
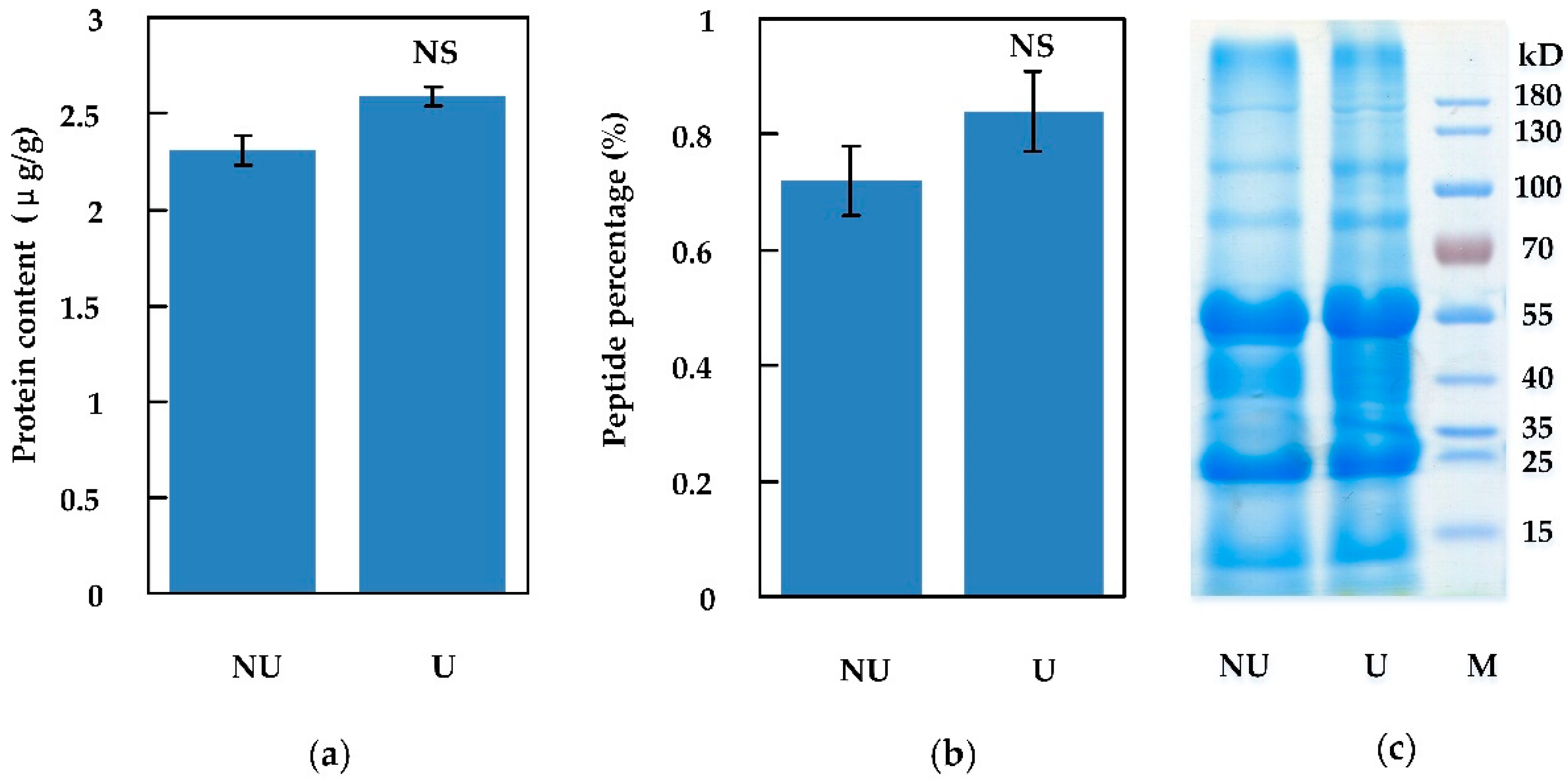
2.1.2. Peptides Extracted by Ultrasonic and Non-Ultrasonic Methods
2.1.3. Peptide Extraction from Protoplasts
2.2. Peptide Identification in Protoplasts
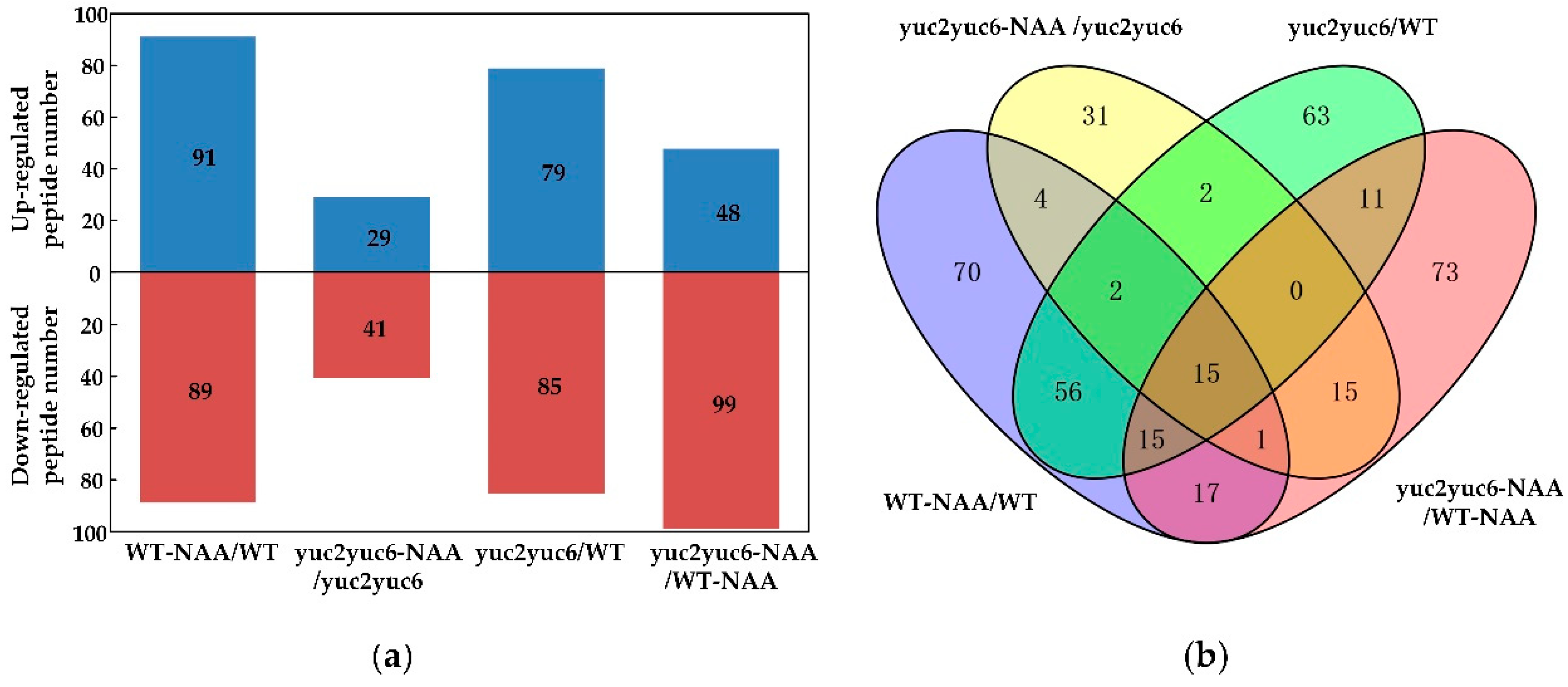
2.3. Effect of Auxin on Peptidomics
3. Materials and Methods
3.1. Reagents/Chemicals and Instrumentation
3.2. Plant Materials and Growth Conditions
3.3. Total-Protein Extraction in Plant Leaves
3.4. Protoplast Preparation and Total-Protein Extraction
3.5. Peptide Sample Preparation
3.6. Bradford Assay and SDS-PAGE
3.7. Peptide Analysis through TOF-MS/MS
3.8. Data Analysis
3.9. Bioinformatics and Annotations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Olsson, V.; Joos, L.; Zhu, S.; Gevaert, K.; Butenko, M.A.; De Smet, I. Look closely, the beautiful may be small: Precursor-derived peptides in plants. Annu. Rev. Plant. Biol. 2019, 70, 153–186. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Han, Z.; Song, W.; Chai, J. Structural insight into recognition of plant peptide hormones by receptors. Mol. Plant. 2016, 9, 1454–1463. [Google Scholar] [CrossRef] [PubMed]
- Farrokhi, N.; Whitelegge, J.P.; Brusslan, J.A. Plant peptides and peptidomics. Plant. Biotechnol. J. 2008, 6, 105–134. [Google Scholar] [CrossRef] [PubMed]
- Matsubayashi, Y. Posttranslationally modified small-peptide signals in plants. Annu. Rev. Plant. Biol. 2014, 65, 385–413. [Google Scholar] [CrossRef] [PubMed]
- Huffaker, A.; Pearce, G.; Ryan, C.A. An endogenous peptide signal in Arabidopsis activates components of the innate immune response. Proc. Natl. Acad. Sci. USA 2006, 103, 10098–10103. [Google Scholar] [CrossRef] [PubMed]
- Nakaminami, K.; Okamoto, M.; Higuchi-Takeuchi, M.; Yoshizumi, T.; Yamaguchi, Y.; Fukao, Y.; Shimizu, M.; Ohashi, C.; Tanaka, M.; Matsui, M. AtPep3 is a hormone-like peptide that plays a role in the salinity stress tolerance of plants. Proc. Natl. Acad. Sci. USA 2018, 115, 5810–5815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minami, E.; Kouchi, H.; Cohn, J.R.; Ogawa, T.; Stacey, G. Expression of the early nodulin, ENOD40, in soybean roots in response to various lipo-chitin signal molecules. Plant J. 1996, 10, 23–32. [Google Scholar] [CrossRef]
- Wen, J.; Lease, K.A.; Walker, J.C. DVL, a novel class of small polypeptides: Overexpression alters Arabidopsis development. Plant J. 2004, 37, 668–677. [Google Scholar] [CrossRef]
- Narita, N.N.; Moore, S.; Horiguchi, G.; Kubo, M.; Demura, T.; Fukuda, H.; Goodrich, J.; Tsukaya, H. Overexpression of a novel small peptide ROTUNDIFOLIA4 decreases cell proliferation and alters leaf shape in Arabidopsis thaliana. Plant J. 2004, 38, 699–713. [Google Scholar] [CrossRef]
- Chilley, P.M.; Casson, S.A.; Tarkowski, P.; Hawkins, N.; Wang, K.L.C.; Hussey, P.J.; Beale, M.; Ecker, J.R.; Sandberg, G.K.; Lindsey, K. The POLARIS peptide of Arabidopsis regulates auxin transport and root growth via effects on ethylene signaling. Plant Cell 2006, 18, 3058–3072. [Google Scholar] [CrossRef]
- Pearce, G. A polypeptide from tomato leaves activates the expression of proteinase inhibitor genes. Science 1991, 253, 895–898. [Google Scholar] [CrossRef] [PubMed]
- Lease, K.A.; Walker, J.C. The Arabidopsis unannotated secreted peptide database, a resource for plant peptidomics. Plant. Physiol. 2006, 142, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Motomitsu, A.; Sawa, S.; Ishida, T. Plant peptide hormone signalling. Essays Biochem. 2015, 58, 115–131. [Google Scholar] [CrossRef] [PubMed]
- Marmiroli, N.; Maestri, E. Plant peptides in defense and signaling. Peptides 2014, 56, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Haruta, M.; Sabat, G.; Stecker, K.; Minkoff, B.B.; Sussman, M.R. A peptide hormone and its receptor protein kinase regulate plant cell expansion. Science 2014, 343, 408–411. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, H.; Moriyama, Y.; Ohyama, K.; Matsubayashi, Y. Biochemical mapping of a ligand-binding domain within Arabidopsis BAM1 reveals diversified ligand recognition mechanisms of plant LRR-RKs. Plant J. 2012, 70, 845–854. [Google Scholar] [CrossRef]
- Qu, L.J.; Li, L.; Lan, Z.; Dresselhaus, T. Peptide signalling during the pollen tube journey and double fertilization. J. Exp. Bot. 2015, 66, 5139–5150. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Liang, L.; Xue, Y.; Jia, P.-F.; Chen, W.; Zhang, M.X.; Wang, Y.C.; Li, H.J.; Yang, W.C. A receptor heteromer mediates the male perception of female attractants in plants. Nature 2016, 531, 241. [Google Scholar] [CrossRef]
- Zhong, S.; Liu, M.; Wang, Z.; Huang, Q.; Hou, S.; Xu, Y.C.; Ge, Z.; Song, Z.; Huang, J.; Qiu, X. Cysteine-rich peptides promote interspecific genetic isolation in Arabidopsis. Science 2019, 364, eaau9564. [Google Scholar] [CrossRef]
- de Bang, T.C.; Lay, K.S.; Scheible, W.R.; Takahashi, H. Small peptide signaling pathways modulating macronutrient utilization in plants. Curr. Opin. Plant. Biol. 2017, 39, 31–39. [Google Scholar] [CrossRef]
- Oh, E.; Seo, P.J.; Kim, J. Signaling peptides and receptors coordinating plant root development. Trends Plant. Sci. 2018, 23, 337–351. [Google Scholar] [CrossRef] [PubMed]
- Kondo, Y.; Hirakawa, Y.; Kieber, J.J.; Fukuda, H. CLE peptides can negatively regulate protoxylem vessel formation via cytokinin signaling. Plant Cell Physiol. 2011, 52, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Whitford, R.; Fernandez, A.; De Groodt, R.; Ortega, E.; Hilson, P. Plant CLE peptides from two distinct functional classes synergistically induce division of vascular cells. Proc. Natl. Acad. Sci. USA 2008, 105, 18625–18630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.S.; Hnilova, M.; Maes, M.; Lin, Y.C.L.; Putarjunan, A.; Han, S.K.; Avila, J.; Torii, K.U. Competitive binding of antagonistic peptides fine-tunes stomatal patterning. Nature 2015, 522, 439. [Google Scholar] [CrossRef] [PubMed]
- Stenvik, G.E.; Tandstad, N.M.; Guo, Y.; Shi, C.L.; Kristiansen, W.; Holmgren, A.; Clark, S.E.; Aalen, R.B.; Butenko, M.A. The EPIP peptide of INFLORESCENCE DEFICIENT IN ABSCISSION is sufficient to induce abscission in Arabidopsis through the receptor-like kinases HAESA and HAESA-LIKE2. Plant Cell 2008, 20, 1805–1817. [Google Scholar] [CrossRef]
- Murphy, E.; Smith, S.; De Smet, I. Small signaling peptides in Arabidopsis development: How cells communicate over a short distance. Plant Cell 2012, 24, 3198–3217. [Google Scholar] [CrossRef] [PubMed]
- Matsubayashi, Y.; Sakagami, Y. Phytosulfokine, sulfated peptides that induce the proliferation of single mesophyll cells of Asparagus officinalis L. Proc. Natl. Acad. Sci. USA 1996, 93, 7623–7627. [Google Scholar] [CrossRef]
- Fletcher, J.C.; Brand, U.; Running, M.P.; Simon, R.; Meyerowitz, E.M. Signaling of cell fate decisions by CLAVATA3 in Arabidopsis shoot meristems. Science 1999, 283, 1911–1914. [Google Scholar] [CrossRef]
- Clark, S.E.; Running, M.P.; Meyerowitz, E.M. CLAVATA3 is a specific regulator of shoot and floral meristem development affecting the same processes as CLAVATA1. Development 1995, 121, 2057–2067. [Google Scholar]
- Mallick, P.; Kuster, B. Proteomics: A pragmatic perspective. Nat. Biotechnol. 2010, 28, 695. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Huffaker, A. Endogenous peptide elicitors in higher plants. Curr. Opin. Plant. Biol. 2011, 14, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Silverstein, K.A.; Gao, L.; Walton, J.D.; Nallu, S.; Guhlin, J.; Young, N.D. Detecting small plant peptides using SPADA (small peptide alignment discovery application). BMC Bioinform. 2013, 14, 335. [Google Scholar] [CrossRef] [PubMed]
- Porto, W.; Souza, V.; Nolasco, D.; Franco, O. In silico identification of novel hevein-like peptide precursors. Peptides 2012, 38, 127. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, K.; Ogawa, M.; Matsubayashi, Y. Identification of a biologically active, small, secreted peptide in Arabidopsis by in silico gene screening, followed by LC-MS-based structure analysis. Plant J. 2008, 55, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Butenko, M.A.; Vie, A.K.; Brembu, T.; Aalen, R.B.; Bones, A.M. Plant peptides in signalling: Looking for new partners. Trends Plant. Sci. 2009, 14, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L.; Lee, C.Y.; Cheng, K.T.; Chang, W.; Huang, R.; Nam, H.G.; Chen, Y. Quantitative peptidomics study reveals that a wound-induced peptide from PR-1 regulates immune signaling in tomato. Plant Cell 2014, 26, 4135–4148. [Google Scholar] [CrossRef] [PubMed]
- Holton, N.; Caño-Delgado, A.; Harrison, K.; Montoya, T.; Chory, J.; Bishop, G.J. Tomato BRASSINOSTEROID INSENSITIVE1 is required for systemin-induced root elongation in Solanum pimpinellifolium but is not essential for wound signaling. Plant Cell 2007, 19, 1709–1717. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, F.; Suzuki, T.; Osakabe, Y.; Betsuyaku, S.; Kondo, Y.; Dohmae, N.; Fukuda, H.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A small peptide modulates stomatal control via abscisic acid in long-distance signalling. Nature 2018, 556, 235. [Google Scholar] [CrossRef]
- Cammarata, J.; Roeder, A.H.; Scanlon, M.J. Cytokinin and CLE signaling are highly intertwined developmental regulators across tissues and species. Curr. Opin. Plant. Biol. 2019, 51, 96–104. [Google Scholar] [CrossRef]
- Tameshige, T.; Okamoto, S.; Lee, J.S.; Aida, M.; Tasaka, M.; Torii, K.U.; Uchida, N. A secreted peptide and its receptors shape the auxin response pattern and leaf margin morphogenesis. Curr. Biol. 2016, 26, 2478–2485. [Google Scholar] [CrossRef]
- Meng, L.; Buchanan, B.B.; Feldman, L.J.; Luan, S. CLE-like (CLEL) peptides control the pattern of root growth and lateral root development in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 1760–1765. [Google Scholar] [CrossRef] [PubMed]
- Whitford, R.; Fernandez, A.; Tejos, R.; Pérez, A.C.; Kleine-Vehn, J.; Vanneste, S.; Drozdzecki, A.; Leitner, J.; Abas, L.; Aerts, M. GOLVEN secretory peptides regulate auxin carrier turnover during plant gravitropic responses. Dev. Cell 2012, 22, 678–685. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Hu, Z.; Lei, C.; Zheng, C.; Wang, J.; Shao, S.; Li, X.; Xia, X.; Cai, X.; Zhou, J. A plant phytosulfokine peptide initiates auxin-dependent immunity through cytosolic Ca2+ signaling in tomato. Plant Cell 2018, 30, 652–667. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Tian, L.; Yang, J.; Zhao, Y.-N.; Zhu, Y.-X.; Dai, X.; Zhao, Y.; Yang, Z.-N. Auxin production in diploid microsporocytes is necessary and sufficient for early stages of pollen development. PLoS Genet. 2018, 14, e1007397. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Dai, X.; Zhao, Y. Auxin biosynthesis by the YUCCA flavin monooxygenases controls the formation of floral organs and vascular tissues in Arabidopsis. Genes Dev. 2006, 20, 1790–1799. [Google Scholar] [CrossRef] [PubMed]
- Skripnikov, A.Y.; Anikanov, N.; Kazakov, V.; Dolgov, S.; Ziganshin, R.K.; Govorun, V.; Ivanov, V. The search for and identification of peptides from the moss Physcomitrella patens. Russ. J. Bioorg. Chem. 2011, 37, 95–104. [Google Scholar] [CrossRef]
- Potocka, I.; Baldwin, T.C.; Kurczynska, E.U. Distribution of lipid transfer protein 1 (LTP1) epitopes associated with morphogenic events during somatic embryogenesis of Arabidopsis thaliana. Plant Cell Rep. 2012, 31, 2031–2045. [Google Scholar] [CrossRef]
- Wang, H.; Sun, Y.; Chang, J.; Zheng, F.; Pei, H.; Yi, Y.; Chang, C.; Dong, C.-H. Regulatory function of Arabidopsis lipid transfer protein 1 (LTP1) in ethylene response and signaling. Plant. Mol. Biol. 2016, 91, 471–484. [Google Scholar] [CrossRef]
- Goda, H.; Sawa, S.; Asami, T.; Fujioka, S.; Shimada, Y.; Yoshida, S. Comprehensive comparison of auxin-regulated and brassinosteroid-regulated genes in Arabidopsis. Plant. Physiol. 2004, 134, 1555–1573. [Google Scholar] [CrossRef]
Sample Availability: Samples of the used plant materials are available from the authors. |






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, W.; Xiao, Y.; Liang, Q.; Su, Y.; Xiao, L. Identification of Potential Auxin-Responsive Small Signaling Peptides through a Peptidomics Approach in Arabidopsis thaliana. Molecules 2019, 24, 3146. https://doi.org/10.3390/molecules24173146
Luo W, Xiao Y, Liang Q, Su Y, Xiao L. Identification of Potential Auxin-Responsive Small Signaling Peptides through a Peptidomics Approach in Arabidopsis thaliana. Molecules. 2019; 24(17):3146. https://doi.org/10.3390/molecules24173146
Chicago/Turabian StyleLuo, Weigui, Yuan Xiao, Qiwen Liang, Yi Su, and Langtao Xiao. 2019. "Identification of Potential Auxin-Responsive Small Signaling Peptides through a Peptidomics Approach in Arabidopsis thaliana" Molecules 24, no. 17: 3146. https://doi.org/10.3390/molecules24173146