Melatonin-Mediated Development of Ovine Cumulus Cells, Perhaps by Regulation of DNA Methylation

Abstract
:1. Introduction
2. Results
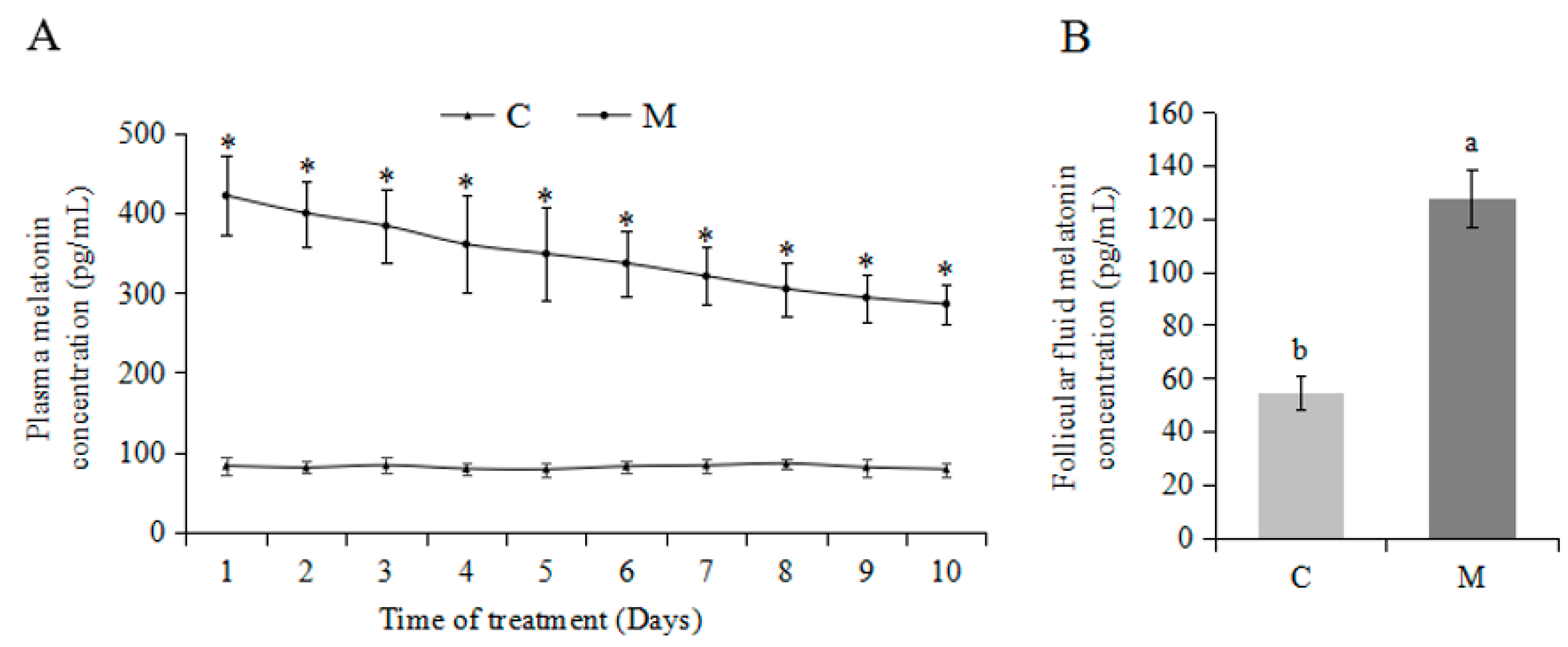
2.1. Effects of Exogenous Melatonin on Plasma and Follicular Fluid Melatonin Concentrations
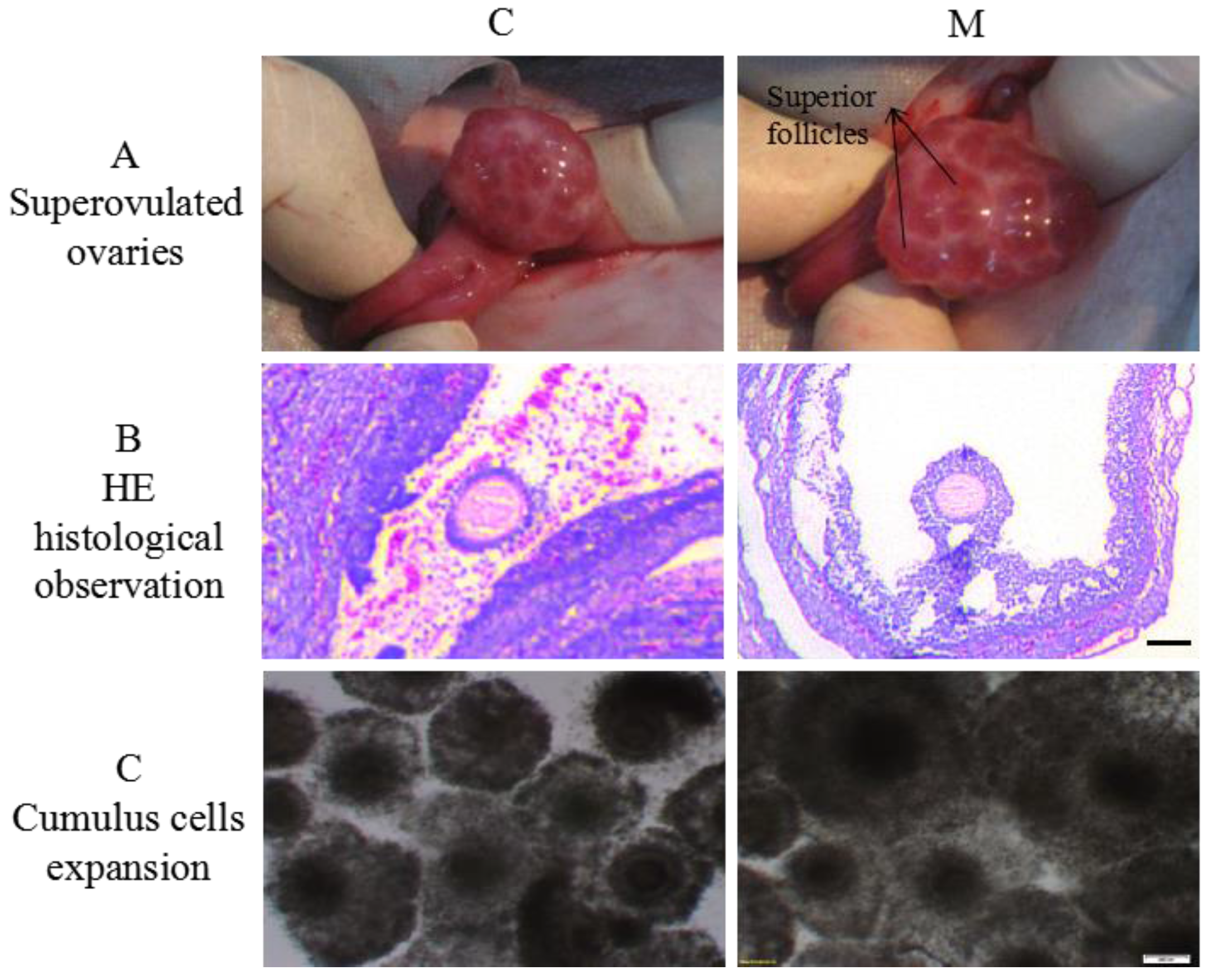
2.2. Effects of Melatonin on Superovulaton and Cumulus Cells Expansion
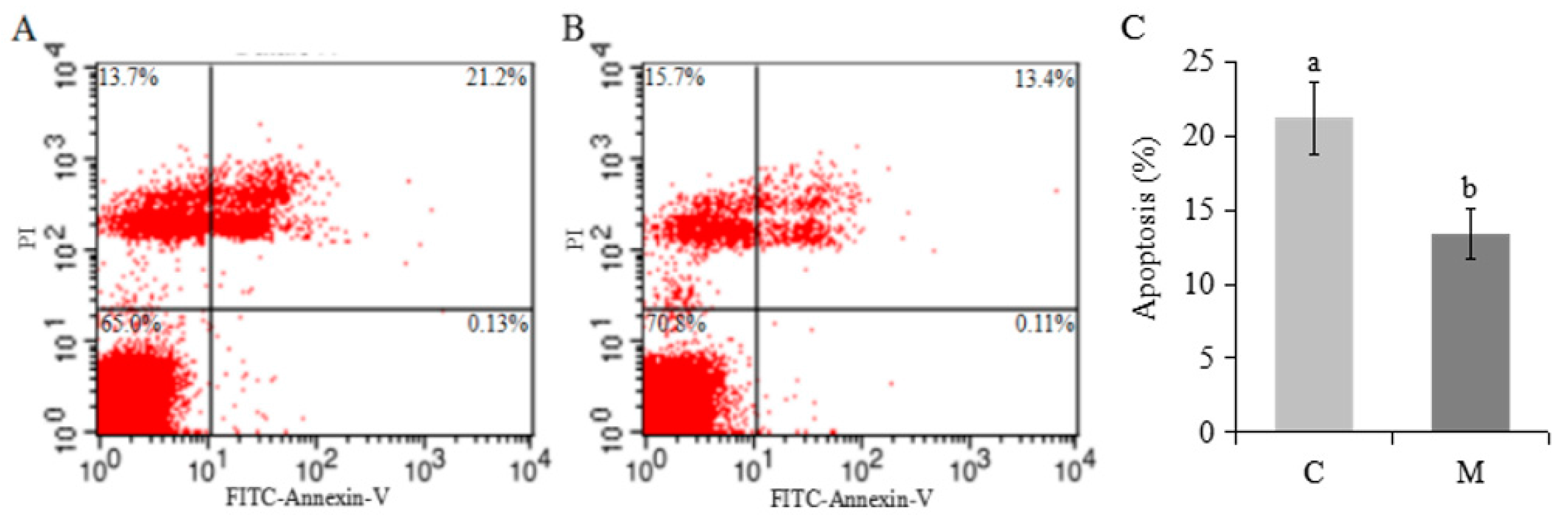
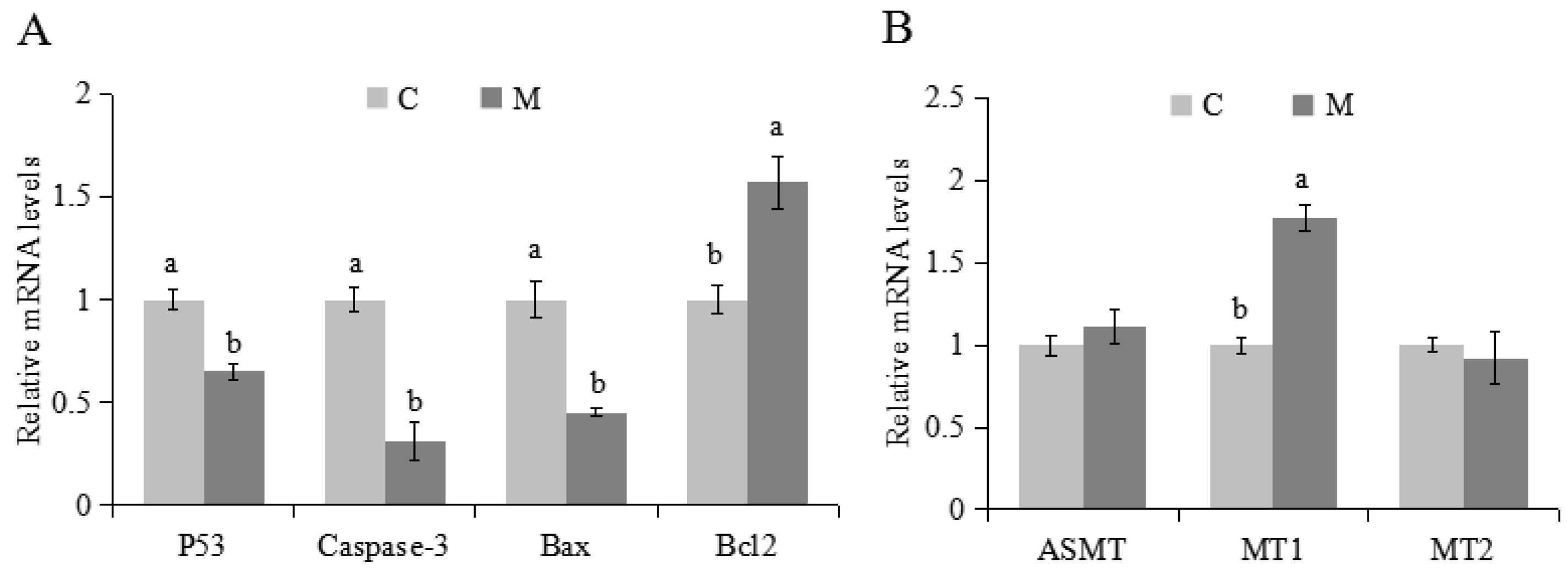
2.3. Effects of Melatonin on Apoptosis and Expression of Related Genes in Cumulus Cells
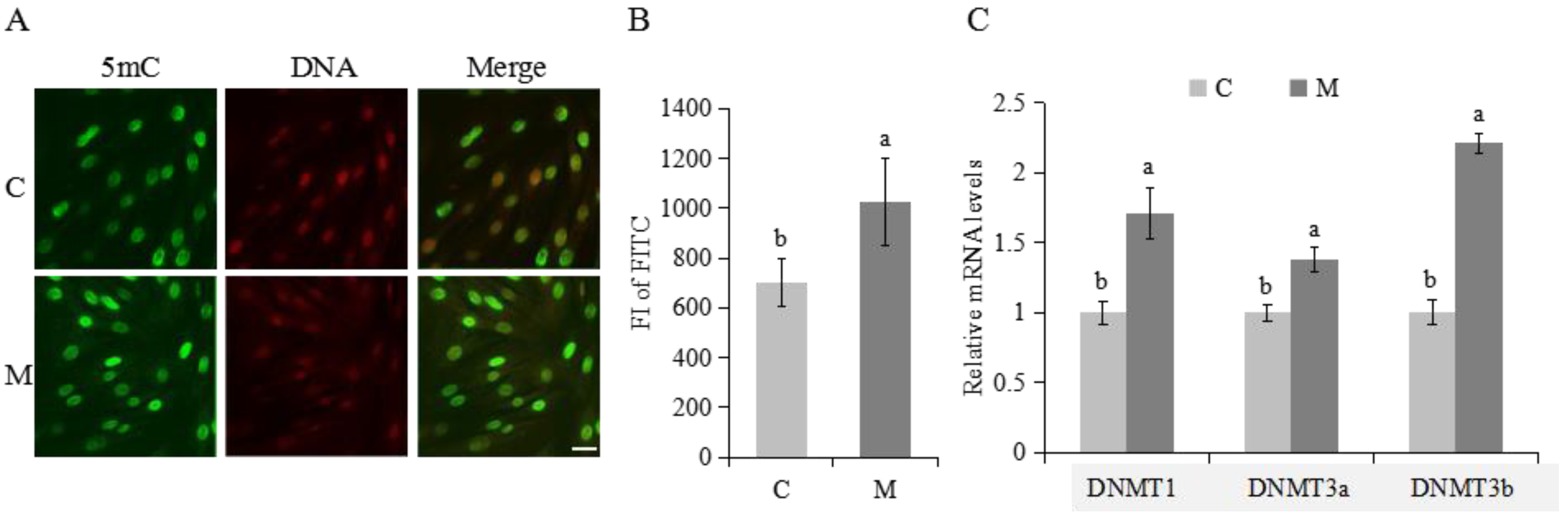
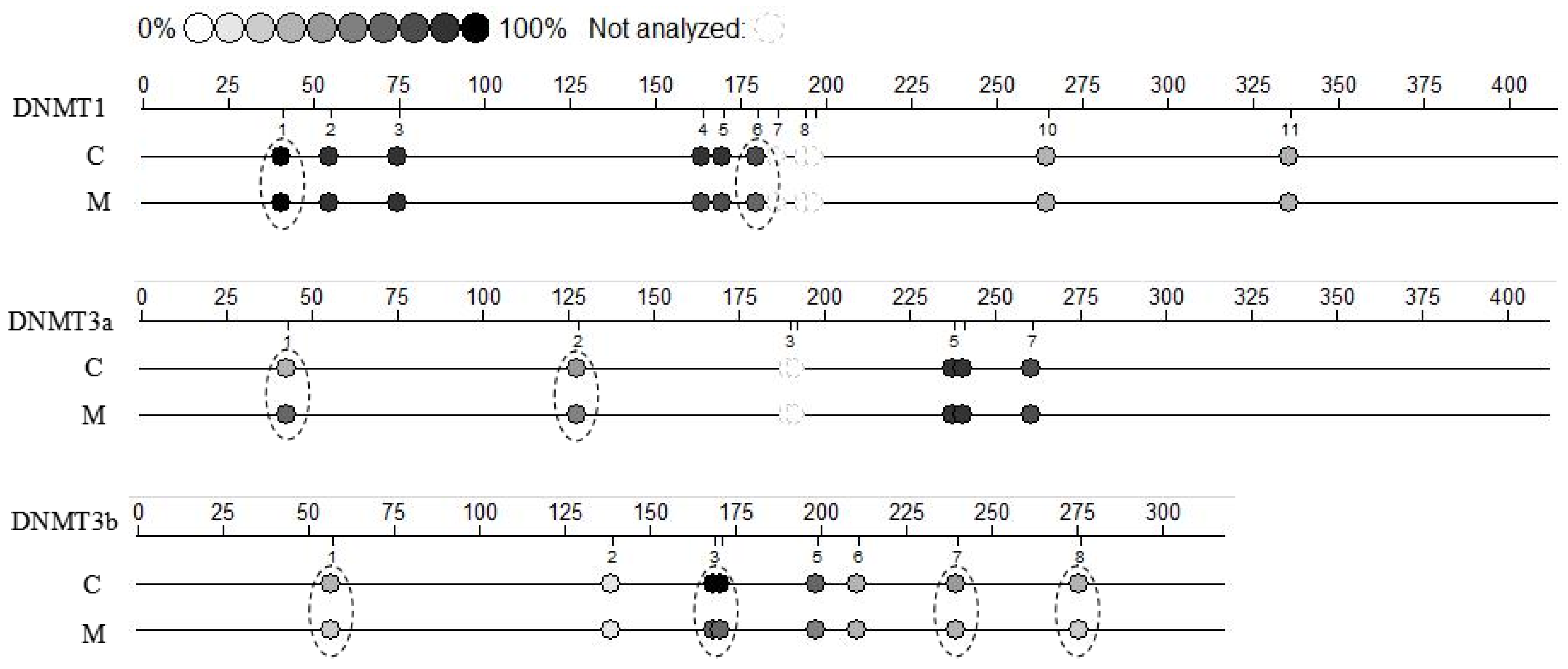
2.4. Methylation Modifications in Cumulus Cells
3. Discussion
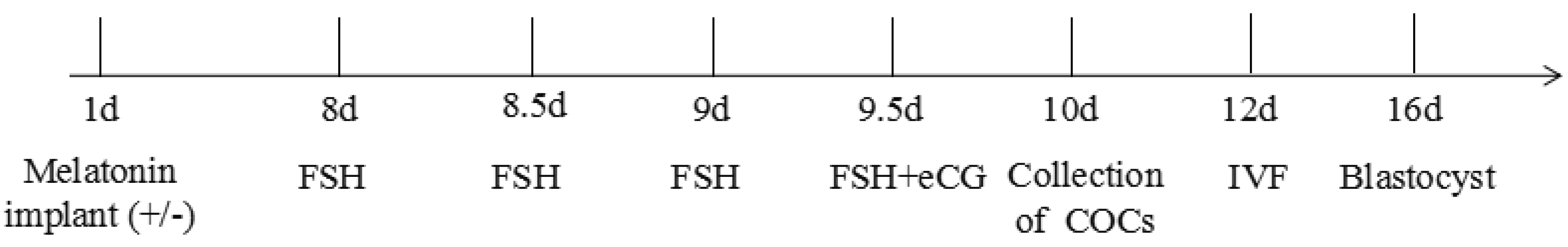
4. Materials and Methods
4.1. Animals
4.2. Donor Superovulation
4.3. Blood Sampling and Hormone Assay
4.4. The Cumulus Oocyte Complexes Collection
4.5. Histology Staining
4.6. In Vitro Maturation (IVM)
4.7. In Vitro Fertilization (IVF) and In Vitro Culture
4.8. Cumulus Cell Apoptosis
4.9. Global Methylation Analysis
4.10. RNA Purification and qRT-PCR
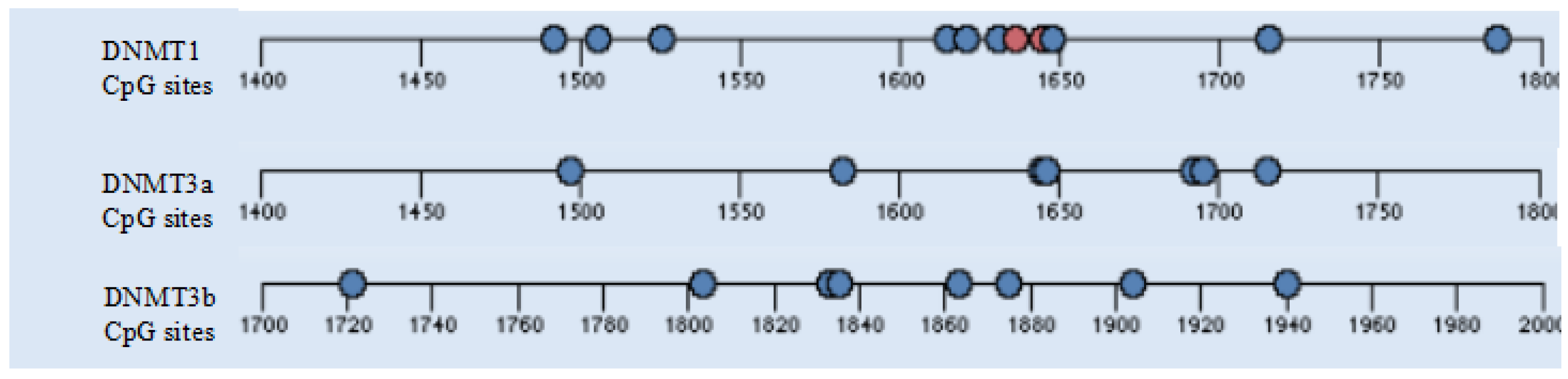
4.11. DNA Methylation Sequence
4.12. Statistical Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Sum No. COCs | No. COCs Examined | |||
|---|---|---|---|---|---|
| Expansion | Apoptosis | RT-PCR | Methylation | ||
| C | 497 | 216 | 69 | 81 | 131 |
| M | 777 | 366 | 92 | 88 | 231 |
| Group | CpG Position (%) | Sum (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 40 | 54 | 74 | 163:169 | 179 | 185 | 193 | 196 | 264 | 335 | ||
| C | 100 ± 5.55 a | 84.2 ± 9.13 | 88.5 ± 8.02 | 77.1 ± 5.73 | 80.2 ± 9.09 a | NA | NA | NA | 35.4 ± 4.24 | 35.1 ± 3.51 | 70.6 ± 8.71 |
| M | 79.7 ± 9.01 b | 96.6 ± 11.6 | 81.9 ± 6.33 | 80.6 ± 9.11 | 62.4 ± 5.83 b | NA | NA | NA | 35.9 ± 2.89 | 35.6 ± 3.77 | 67.6 ± 7.49 |
| Group | CpG Position (%) | Sum (%) | ||||
|---|---|---|---|---|---|---|
| 42 | 127 | 189:191 | 237:240 | 260 | ||
| C | 47.5 ± 4.02 b | 44.3 ± 5.05 b | NA | 85.4 ± 8.08 | 77.4 ± 6.48 | 63.3 ± 7.34 |
| M | 64.1 ± 7.17 a | 53.2 ± 4.13 a | NA | 81.9 ± 6.91 | 79.7 ± 7.05 | 69.3 ± 5.77 |
| Group | CpG Position (%) | Sum (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| 56 | 138 | 168:170 | 198 | 210 | 239 | 275 | ||
| C | 36.1 ± 4.55 a | 15.4 ± 2.13 | 92.9 ± 8.81 a | 66.6 ± 5.37 | 37.2 ± 4.33 | 50.6 ± 4.22 a | 31.4 ± 3.25 a | 46.7 ± 5.79 a |
| M | 27.6 ± 3.01 b | 14.7 ± 1.64 | 68.4 ± 5.19 b | 51.4 ± 4.42 | 34.8 ± 3.47 | 38.7 ± 4.09 b | 20.6 ± 1.68 b | 36.0 ± 4.82 b |

References
- Zhang, X.S.; Zhan, J.L.; Cui, M.S.; Li, Y.H.; Feng, X.Z. The study on the effect factors of superovulation on lamb. Chin. Herbiv. Sci. 2013, 33, 67–70. (In Chinese) [Google Scholar]
- Kelly, J.M.; Kleemann, D.O.; Walker, S.K. Enhanced efficiency in the production of offspring from 4- to 8-week-old lambs. Theriogenology 2005, 63, 1876–1890. [Google Scholar] [CrossRef] [PubMed]
- Kochhar, H.P.; Wu, B.; Morris, L.; Buckrell, B.C.; Pollard, J.W. Maturation status, protein synthesis and developmental competence of oocytes derived from lambs and ewes. Reprod. Domest. Anim. 2002, 37, 19–25. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.K.; Dwarte, D.; Ryan, J.P.; Maxwell, W.M.; Evans, G. Developmental capacity, energy metabolism and ultrastructure of mature oocytes from prepubertal and adult sheep. Reprod. Fertil. Dev. 1996, 8, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.J.; Wu, S.N.; Shen, J.P.; Wang, D.H.; Kong, X.W. The beneficial effects of cumulus cells and oocyte-cumulus cell gap junctions depends on oocyte maturation and fertilization methods in mice. PeerJ 2016, 4, 1761. [Google Scholar] [CrossRef] [PubMed]
- Ptak, P.L.; Dattena, M.; Tischner, M.; Cappai, P. Offspring from one-month-old lambs: studies on the developmental capability of prepubertal oocytes. Biol. Reprod. 1999, 61, 1568–1574. [Google Scholar] [CrossRef] [PubMed]
- Haenisch-Woehl, A.; Kölle, S.; Neumüller, C.; Sinowatz, F.; Braun, J. Morphology of canine cumulus-oocyte complexes in pre-pubertal bitches. Anat. Histol. Embryol. 2003, 32, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Anguita, B.; Paramio, M.T.; Morató, R.; Romaguera, R.; Jiménez-Macedo, A.R. Effect of the apoptosis rate observed in oocytes and cumulus cells on embryo development in prepubertal goats. Anim. Reprod. Sci. 2009, 116, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Zhang, X.S.; Zhang, J.L.; Zhong, R.Z.; Zhou, D.W. Global DNA methylation and related mRNA profiles in sheep oocytes and early embryos derived from pre-pubertal and adult donors. Anim. Reprod. Sci. 2016, 164, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Diederich, M.; Hansmann, T.; Heinzmann, J.; Bargkues, B.; Herrmann, D. DNA methylation and mRNA expression profiles in bovine oocytes derived from prepubertal and adult donors. Reproduction 2012, 144, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Ptak, G.; Matsukawa, K.; Palmieri, C.; Salda, L.D.; Scapolo, P.A.; Loi, P. Developmental and functional evidence of nuclear immaturity in prepubertal oocytes. Hum. Reprod. 2006, 21, 2228–2237. [Google Scholar] [CrossRef] [PubMed]
- Biian, E.L.; Karsch, F.J.; Hopkins, J.W. Role of the pineal gland in ovine photoperiodism:regulation of seasonal breeding and negative feedback effects of estradiol upon luteinizinghormone secretion. Endocrinology 1983, 113, 329–336. [Google Scholar]
- Claypool, L.E.; Wood, R.I.; Yellon, S.M.; Foster, D.L. The ontogeny of melatonin secretion in the lamb. Endocrinology 1989, 124, 2135–2143. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, A.; Topal, T.; Oter, S.; Tan, D.X.; Reiter, R.J. Hyperglycemia-related pathophysiologic mechanisms and potential beneficial actions of melatonin. Mini Rev. Med. Chem. 2008, 8, 1144–1153. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, A.; Rosales-Corral, S.; Reiter, R.J. Gene regulation by melatonin linked to epigenetic phenomena. Gene 2012, 503, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Menendez-Pelaez, A.; Poeggeler, B.; Reiter, R.J.; Barlow, W.L.; Pablos, M.I.; Tan, D.X. Nuclear localization ofmelatonin in different mammalian tissues: Immunocytochemical and radioimmunoassay evidence. J. Cell. Biochem. 1993, 53, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Tomas-Zapico, C.; Coto-Montes, A. A proposed mechanismto explain the stimulatory effect of melatonin on antioxidativeenzymes. J. Pineal Res. 2005, 39, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Karasek, M.; Pawlikowski, M. Antiproliferative effects of melatonin and CGP 52608. Biol. Signals Recept. 1999, 8, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Yie, S.M.; Niles, L.P.; Younglai, E.V. Melatonin receptors on human granulosa cell membranes. J. Clin. Endocrinol. Metab. 1995, 80, 1747–1749. [Google Scholar] [PubMed]
- Niles, L.P.; Wang, J.; Shen, L.; Lobb, D.K.; Younglai, E.V. Melatonin receptor mRNA expression in human granulosa cells. Mol. Cell. Endocrinol. 1999, 156, 107–110. [Google Scholar] [CrossRef]
- Kang, J.T.; Koo, O.J.; Kwon, D.K.; Park, H.J.; Jang, G.; Kang, S.K.; Lee, B.C. Effects of melatonin on in vitro maturation of porcine oocyte and expression of melatonin receptor RNA in cumulus and granulosa cells. J. Pineal Res. 2009, 46, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Peyrot, F.; Ducrocq, C. Potential role of tryptophan derivatives in stress responses characterized by the generation of reactive oxygen and nitrogen species. J. Pineal Res. 2008, 45, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Carcangiu, V.; Vacca, G.M.; Mura, M.C.; Dettori, M.L.; Pazzola, M.; Luridiana, S.; Bini, P.P. Relationship between MTNR1A melatonin receptor gene polymorphism and seasonal reproduction in different goat breeds. Anim. Reprod. Sci. 2009, 110, 71–78. [Google Scholar] [CrossRef] [PubMed]
- El-Raey, M.; Geshi, M.; Somfai, T.; Kaneda, M.; Hirako, M.; Abdel-Ghaffar, A.E.; Sosa, G.A.; El-Roos, M.E.; Nagai, T. Evidence of melatonin synthesis in the cumulus oocyte complexes and its role in enhancing oocyte maturation in vitro in cattle. Mol. Reprod. Dev. 2011, 78, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Weaver, D.R.; Liu, C.; Reppert, S.M. Nature’s knockout: the Mel1b receptor is not necessary for reproductive and circadian responses to melatonin in Siberian hamsters. Mol. Endocrinol. 1996, 10, 1478–1487. [Google Scholar] [PubMed]
- Takada, L.; Junior, A.M.; Mingoti, G.Z.; Balieiro, J.C.; Cipolla-Neto, J.; Coelho, L.A. Effect of melatonin on DNA damage of bovine cumulus cells during in vitro maturation (IVM) and on in vitro embryo development. Res. Vet. Sci. 2012, 92, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Brzezinski, A.; Seibel, M.M.; Lynch, H.J.; Deng, M.H.; Wurtman, R.J. Melatonin in human preovulatory follicular fluid. J. Clin. Endocrinol. Metab. 1987, 64, 865–867. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.M.; Tian, X.Z.; Zhou, G.B.; Wang, L.; Gao, C.; Zhu, S.E.; Zeng, S.M.; Tian, J.H.; Liu, G.S. Melatonin exists in porcine follicular fluid and improves in vitro maturation and parthenogenetic development of porcine oocytes. J. Pineal Res. 2009, 47, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Osorio, N.; Kim, I.J.; Wang, H.; Kaya, A.; Memili, E. Melatonin increases cleavage rate of porcine preimplantation embryos in vitro. J. Pineal Res. 2007, 43, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.M.; Hao, H.S.; Du, W.H.; Zhao, S.J.; Wang, H.Y.; Wang, N.; Wang, D.; Liu, Y.; Qin, T.; Zhu, H.B. Melatonin inhibits apoptosis and improves the developmental potential of vitrified bovine oocytes. J. Pineal Res. 2016, 60, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.W.; Sun, Y.Q.; Jiang, X.L.; Huang, Z.Q.; Zhao, S.J.; Du, W.H.; Hao, H.S.; Zhao, X.M.; Zhu, H.B. Protective effects of melatonin on bovine sperm characteristics and subsequent in vitro embryo development. Mol. Reprod. Dev. 2016, 83, 993–1002. [Google Scholar] [CrossRef] [PubMed]
- Coelho, L.A.; Peres, R.; Amaral, F.G.; Reiter, R.J.; Cipolla-Neto, J. Daily differential expression of melatonin-related genes and clock genes in rat cumulus-oocyte complex: Changes after pinealectomy. J. Pineal Res. 2015, 58, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Tamura, H.; Takasaki, A.; Miwa, I.; Taniguchi, K.; Maekawa, R.; Asada, H.; Taketani, T.; Matsuoka, A.; Yamagata, Y.; Shimamura, K.; et al. Oxidative stress impairs oocyte quality and melatonin protects oocytes from free radical damage and improves fertilization rate. J. Pineal Res. 2008, 44, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Tamura, H.; Takayama, H.; Kato, H. Increased endogenous level of melatonin in preovulatory human follicles does not directly influence progesterone production. Fertil. Steril. 2003, 80, 1012–1016. [Google Scholar] [CrossRef]
- Tamura, H.; Nakamura, Y.; Korkmaz, A.; Manchester, L.C.; Tan, D.X.; Sugino, N.; Reiter, R.J. Melatonin and the ovary: Physiological and pathophysiological implications. Fertil. Steril. 2009, 92, 328–343. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Manchester, L.C.; Paredes, S.D.; Mayo, J.C.; Sainz, R.M. Melatonin and reproduction revisited. Biol. Reprod. 2009, 81, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Woo, M.M.; Tai, C.J.; Kang, S.K.; Nathwani, P.S.; Pang, S.F.; Leung, P.C. Direct action of melatonin in human granulosa-luteal cells. J. Clin. Endocrinol. Metab. 2001, 86, 4789–4797. [Google Scholar] [CrossRef] [PubMed]
- Picinato, M.C.; Hirata, A.E.; Cipolla-Neto, J.; Curi, R.; Carvalho, C.R.; Anhe, G.F.; Carpinelli, A.R. Activation of insulin and IGF-1 signaling pathways by melatonin through MT1 receptor in isolated rat pancreatic islets. J. Pineal Res. 2008, 44, 88–94. [Google Scholar] [CrossRef] [PubMed]
- PTsiligianni, T.; Valasi, I.; Cseh, S.; Vainas, E.; Faigl, V. Effects of melatonin treatment on follicular development and oocyte quality in Chios ewes-short communication. Acta Vet. Hung. 2009, 57, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Haghighat, N.; Van Winkle, L.J. Developmental change in follicular cell-enhanced amino acid uptake into mouse oocytes that depends on intact gap junctions and transport system Gly. J. Exp. Zool. 1990, 253, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Buccione, R.; Schroeder, A.C.; Eppig, J.J. Interactions between somatic cells and germ cells throughout mammalian oogenesis. Biol. Reprod. 1990, 43, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Thibault, C.; Szollosi, D.; Gerard, M. Mammalian oocyte maturation. Reprod. Nutr. Dev. 1987, 27, 865–896. [Google Scholar] [CrossRef] [PubMed]
- Zuelke, K.A.; Brackett, B.G. Luteinizing hormone-enhanced in vitro maturation of bovine oocytes with and without protein supplementation. Biol. Reprod. 1990, 43, 784–787. [Google Scholar] [CrossRef] [PubMed]
- Gutnisky, C.; Dalvit, G.C.; Pintos, L.N.; Thompson, J.G.; Beconi, M.T.; Cetica, P.D. Influence of hyaluronic acid synthesis and cumulus mucification on bovine oocyte in vitro maturation, fertilisation and embryo development. Reprod. Fertil. Dev. 2007, 19, 488–497. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Park, S.M.; Lee, E.; Kim, J.H.; Jeong, Y.I.; Lee, J.Y.; Park, S.W.; Kim, H.S.; Hossein, M.S.; Jeong, Y.W.; et al. Anti-apoptotic effect of melatonin on preimplantation development of porcine parthenogenetic embryos. Mol. Reprod. Dev. 2008, 75, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Joo, S.S.; Yoo, Y.M. Melatonin induces apoptotic death in LNCaP cells via p38 and JNK pathways: Therapeutic implications for prostate cancer. J. Pineal Res. 2009, 47, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Leja-Szpak, A.; Jaworek, J.; Pierzchalski, P.; Reiter, R.J. Melatonin induces pro-apoptotic signaling pathway in human pancreatic carcinoma cells (PANC-1). J. Pineal Res. 2010, 49, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.H.; Hong, S.W.; Zheng, H.M.; Lee, H.S.; Lee, H.; Lee, D.H.; Lee, S.Y.; Hong, S.S. Melatonin ameliorates cerulein-induced pancreatitis by the modulation of nuclear erythroid 2-related factor 2 and nuclear factor-kappaB in rats. J. Pineal Res. 2010, 48, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.C.; Ryan, A.F. Mechanisms of sensorineural celldamage, death and survival in the Cochlea. Front. Aging Neurosci. 2015, 7, 58. [Google Scholar] [CrossRef] [PubMed]
- Juknat, A.A.; Mendez, M.V.; Quaglino, A.; Fameli, C.I.; Mena, M.; Kotler, M.L. Melatonin prevents hydrogen peroxide-induced Bax expression in cultured rat astrocytes. J. Pineal Res. 2005, 38, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Maity, P.; Bindu, S.; Dey, S.; Goyal, M.; Alam, A.; Pal, C.; Reiter, R.; Bandyopadhyay, U. Melatonin reduces indomethacin-induced gastric mucosal cell apoptosis by preventing mitochondrial oxidative stress and the activation of mitochondrial pathway of apoptosis. J. Pineal Res. 2009, 46, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Wang, F.; He, C.; Zhang, L.; Tan, D.; Reiter, R.J.; Xu, J.; Ji, P.; Liu, G. Beneficial effects of melatonin on bovine oocytes maturation: a mechanistic approach. J. Pineal Res. 2014, 57, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Talpur, H.S.; Worku, T.; Rehman, Z.U.; Dad, R.; Bhattarai, D.; Bano, I.; Farmanullah; Liang, A.; He, C.; Yang, L. Knockdown of melatonin receptor 1 and induction of follicle-stimulating hormone on the regulation of mouse granulosa cell function. Reprod. Biol. 2017, 17, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Ottenhof, T.; Rzeczkowska, P.A.; Niles, L.P. Epigenetic targets for melatonin: Induction of histone H3 hyperacetylation and gene expression in C17.2 neural stem cells. J. Pineal Res. 2008, 45, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.G.; Tang, S.T.; Tseng, H.P.; Wu, K.K. Melatonin suppresses macrophage cyclooxygenase-2 and inducible nitric oxide synthase expression by inhibiting p52 acetylation and binding. Blood 2006, 108, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Huang, L.T.; Chan, J.Y. Transcriptional regulation of programmed hypertension by melatonin: An epigenetic perspective. Int. J. Mol. Sci. 2014, 15, 18484–18495. [Google Scholar] [CrossRef] [PubMed]
- Adcock, I.M.; Cosio, B.; Tsaprouni, L.; Barnes, P.J.; Ito, K. Redox regulation of histone deacetylases and glucocorticoid-mediated inhibition of the inflammatory response. Antioxid. Redox Signal. 2005, 7, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Glass, C.K.; Ogawa, S. Combinatorial roles of nuclear receptors in inflammation and immunity. Nat. Rev. Immunol. 2006, 6, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, A.; Reiter, R.J. Epigenetic regulation: A new research area for melatonin? J. Pineal Res. 2008, 44, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Crosio, C.; Cermakian, N.; Allis, C.D.; Sassone-Corsi, P. Light induces chromatin modification in cells of the mammalian circadian clock. Nat. Neurosci. 2000, 3, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Marei, W.F.; Wathes, D.C.; Fouladi-Nashta, A.A. Impact of linoleic acid on bovine oocyte maturation and embryo development. Reproduction 2010, 139, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |







| Groups | Mean ± SEM No. Follicles ≥ 2 mm (n = 9) | Mean ± SEM No. Oocytes (n = 9) | IVF | ||
|---|---|---|---|---|---|
| Maturation Rate (%) (No. Mature/No. Total) | Cleavage Rate (%) (No. Cleavage/No. Mature) | Blastocyst Rate (%) (No. Blastocyst/No. Cleavage) | |||
| C | 79.4 ± 9.22 b | 55.2 ± 4.29 b | 71.8 (155/216) b | 52.3(81/155) b | 7.41(6/81) b |
| M | 106 ± 14.9 a | 86.3 ± 7.77 a | 79.5 (291/366) a | 68.4(199/291) a | 14.1(28/199) a |
| Groups | No. COCs Examined | No. (%) Oocytes with Cumulus Cells | ||
|---|---|---|---|---|
| Not Expanded | Partially Expanded | Fully Expanded | ||
| C | 216 | 39 (18.0) a | 67 (31.0) a | 110 (51.0) b |
| M | 366 | 29 (8.00) b | 59 (16.1) b | 278 (76.0) a |
| Gene | Primer Sequence | T annealing (°C) × No. Cycles | Fragment Size (bp) |
|---|---|---|---|
| β-Actin | 5′GTCATCACCATCGGCAATGA 3′CGTGAATGCCGCAGGATT | 60 × 35 | 159 |
| ASMT | 5′TCATTTTCCTGAGTGCGTTG 3′CTCCCAGGTTCTCTTTGCTG | 58 × 35 | 205 |
| MT1 | 5′GGAGGGTGAAACCTGACGAC 3′CCCAGCAAATGGCAAAGAGG | 57 × 35 | 99 |
| MT2 | 5′GGCTCCGTCTTCAACATCACC 3′GCAGAAGGACCAGCAGGGTG | 60 × 35 | 145 |
| P53 | 5′GAAGACCTACCCTGGCAATTAC 3′AGAACAGCTTGTTAAGGGAAGG | 60 × 35 | 103 |
| Bax | 5′GGCTGGACATTGGACTTCCTTC 3′TGGTCACTGTCTGCCATGTGG | 57 × 35 | 112 |
| Caspase 3 | 5′CAGACAGTGGTGCTGAGGATGA 3′GCTACCTTTCGGTTAACCCGA | 56 × 35 | 211 |
| Bcl2 | 5′GACTGACACTGAGTTTGGCTACG 3′GAGTCCTTTCCACTTCGTCCTG | 58 × 35 | 152 |
| DNMT1 | 5′CGCATGGGCTACCAGTGCACCTT 3′GGGCTCCCCGTTGTATGAAATCT | 58 × 35 | 158 |
| DNMT3a | 5′CAACGGAGAAGCCTAAGGTCAA 3′TTGAGGCTCCCACAAGAGATG | 60 × 35 | 244 |
| DNMT3b | 5′GACTCATTGGAGGACCAGCTGAAGC 3′CAGCACCTCCAGGCACTCCACACAG | 59 × 35 | 130 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, Y.; Deng, S.; Zhang, J.; Liu, H.; Li, Y.; Zhang, X.; Liu, Y. Melatonin-Mediated Development of Ovine Cumulus Cells, Perhaps by Regulation of DNA Methylation. Molecules 2018, 23, 494. https://doi.org/10.3390/molecules23020494
Fang Y, Deng S, Zhang J, Liu H, Li Y, Zhang X, Liu Y. Melatonin-Mediated Development of Ovine Cumulus Cells, Perhaps by Regulation of DNA Methylation. Molecules. 2018; 23(2):494. https://doi.org/10.3390/molecules23020494
Chicago/Turabian StyleFang, Yi, Shoulong Deng, Jinlong Zhang, Haijun Liu, Yihai Li, Xiaosheng Zhang, and Yixun Liu. 2018. "Melatonin-Mediated Development of Ovine Cumulus Cells, Perhaps by Regulation of DNA Methylation" Molecules 23, no. 2: 494. https://doi.org/10.3390/molecules23020494