Lactobacillus plantarum CQPC11 Isolated from Sichuan Pickled Cabbages Antagonizes d-galactose-Induced Oxidation and Aging in Mice

 and
and
Abstract
:1. Introduction
2. Results
2.1. Isolation and Identification of LP-CQPC11
2.2. Mouse Organ Indices
2.3. Levels of NO, SOD, GSH-Px, GSH, and MDA
2.4. Pathological Observation of Mouse Liver and Spleen
2.5. Gene Expression Analysis of Mouse Liver
2.6. Gene Expression Analysis of Mouse Spleen
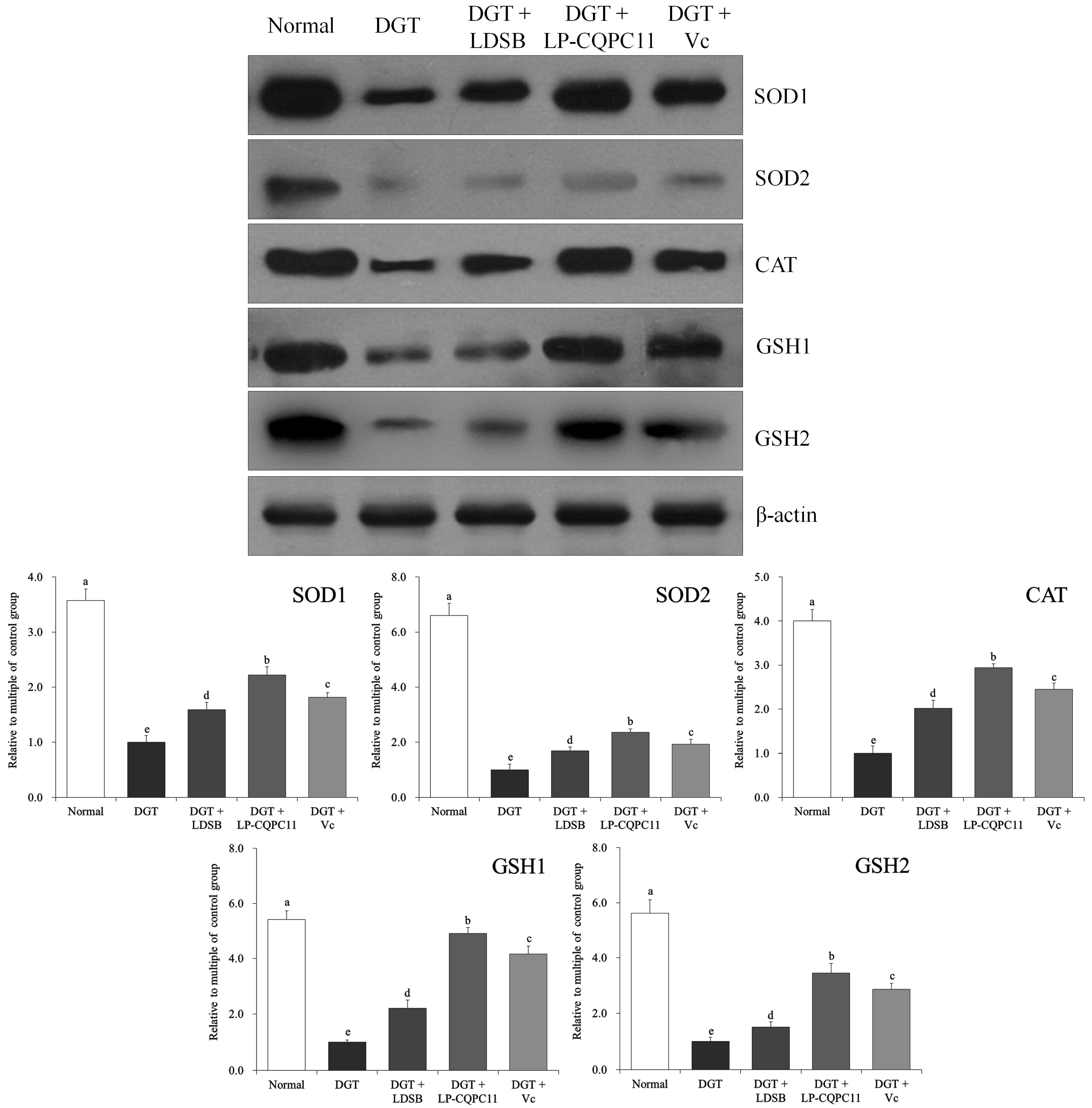
2.7. Mouse Liver Protein Level Analysis
2.8. Mouse Spleen Protein Level Analysis
3. Discussion
4. Materials and Methods
4.1. Isolation and Identification of Lactic Acid Bacteria
4.2. Animal Models of Oxidation-Induced Aging
4.3. Measurement of NO, SOD, GSH-Px, and MDA Levels in the Serum and Liver
4.4. Pathological Observation of Liver and Spleen Tissues
4.5. Quantitative PCR (qPCR) Assay
4.6. Western Blot Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chen, G.; Yu, W.H.; Zhang, Q.S.; Song, P.; Zhang, B.B.; Liu, Z.; You, J.G.; Li, H. Research of Sichuan Paocai and Lactic Acid Bacteria. Adv. J. Food Sci. Technol. 2014, 6, 1–5. [Google Scholar]
- Yang, J.X.; Zhang, L.L.; Jiang, H.Y.; He, Z.F. Isolation and identification of lactic bacteria in pickled vegetables from Meishan city. Food Sci. 2015, 36, 158–163. [Google Scholar]
- Huang, Y.; Luo, Y.B.; Zhai, Z.Y.; Zhang, H.X.; Yang, C.X.; Tian, H.T.; Li, Z.; Feng, J.N.; Liu, H.; Hao, Y.L. Characterization and application of an anti-Listeria bacteriocin produced by Pediococcus pentosaceus 05-10 isolated from Sichuan Pickle, a traditionally fermented vegetable product from China. Food Control. 2009, 20, 1030–1035. [Google Scholar] [CrossRef]
- Liu, A.; Li, X.; Pu, B.; Ao, X.; Zhou, K.; He, L.; Chen, S.; Liu, S. Use of psychrotolerant lactic acid bacteria (Lactobacillus spp. and Leuconostoc spp.) Isolated from Chinese Traditional Paocai for the Quality Improvement of Paocai Products. J. Agric. Food Chem. 2017, 65, 2580–2587. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.L.; Zhang, L.W.; Shi, J.; Yi, H.X. Effect of complex food environment on production of enteriocin IN 3531 with Enterococcus faecium IN3531 as a starter in Chinese fermentation Paocai making. Adv. Mater. Res. 2014, 884–885, 429–432. [Google Scholar] [CrossRef]
- Wang, J.J.; Cui, B.N.; Zhang, Z.Z. Flavor generation mechanism of pickle. Food Res. Dev. 2008, 29, 163–166. [Google Scholar]
- Liang, H.; Yin, L.; Zhang, Y.; Chang, C.; Zhang, W. Dynamics and diversity of a microbial community during the fermentation of industrialized Qingcai paocai, a traditional Chinese fermented vegetable food, as assessed by Illumina MiSeq sequencing, DGGE and qPCR assay. Ann. Microbiol. 2018, 68, 111–122. [Google Scholar] [CrossRef]
- Qian, Y.; Song, J.-L.; Yi, R.; Li, G.; Sun, P.; Zhao, X.; Huo, G. Preventive effects of Lactobacillus plantarum YS4 on constipation induced by activated carbon in mice. Appl. Sci. 2018, 8, 363. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, X.; Wang, H.; Yang, Z.; Li, J.; Suo, H. Prevent effects of Lactobacillus fermentum HY01 on dextran sulfate sodium-induced colitis in mice. Nutrients 2017, 9, 545. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, J.; Yi, R.; Mu, J.; Zhao, X.; Yang, Z. Hepatoprotective effects of Lactobacillus on carbon tetrachloride-induced acute liver injury in mice. Int. J. Mol. Sci. 2018, 19, 2212. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Tan, F.; Yi, R.; Mu, J.; Zhao, X.; Yang, Z. Effects of Lactobacillus on mice with diabetes induced by high-fat diet with streptozotocin (STZ). Appl. Sci. 2018, 8, 1249. [Google Scholar] [CrossRef]
- Buford, T.W. Hypertension and aging. Ageing Res. Rev. 2016, 26, 96–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chard, S.; Harris-Wallace, B.; Roth, E.G.; Girling, L.M.; Rubinstein, R.; Reese, A.M.; Quinn, C.C.; Eckert, J.K. Successful aging among African American older adults with type 2 diabetes. J. Gerontol. B Psychol. Sci. Soc. Sci. 2017, 72, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Kitada, M.; Ogura, Y.; Koya, D. The protective role of Sirt1 in vascular tissue: Its relationship to vascular aging and atherosclerosis. Aging 2016, 8, 2290–2307. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xu, M.; Shen, B.; Li, M.; Gao, Q.; Wei, S.G. Moderate exercise prevents neurodegeneration in d-galactose-induced aging mice. Neural. Regen. Res. 2016, 11, 807–815. [Google Scholar] [PubMed]
- Tang, T.; He, B. Treatment of d-galactose induced mouse aging with Lycium barbarum polysaccharides and its mechanism study. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.S.; Singer, B.D.; Vaughan, D.E. Molecular and physiological manifestations and measurement of aging in humans. Aging Cell 2017, 16, 624–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manini, T.M. Energy expenditure and aging. Ageing Res. Rev. 2010, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Tessari, P. Nitric oxide in the normal kidney and in patients with diabetic nephropathy. J. Nephrol. 2015, 28, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Eroglu, E.; Hallström, S.; Bischof, H.; Opelt, M.; Schmidt, K.; Mayer, B.; Waldeck-Weiermair, M.; Graier, W.F.; Malli, R. Real-time visualization of distinct nitric oxide generation of nitric oxide synthase isoforms in single cells. Nitric Oxide 2017, 70, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Hyun, D.H.; Jenner, P.; Halliwell, B. Effect of proteasome inhibition on cellular oxidative damage, antioxidant defences and nitric oxide production. J. Neurochem. 2001, 78, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Bonthius, D.J.J.; Winters, Z.; Karacay, B.; Bousquet, S.L.; Bonthius, D.J. Importance of genetics in fetal alcohol effects: Null mutation of the nNOS gene worsens alcohol-induced cerebellar neuronal losses and behavioral deficits. Neurotoxicology 2015, 46, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Fukai, T.; Siegfried, M.R.; Ushio-Fukai, M.; Cheng, Y.; Kojda, G.; Harrison, D.G. Regulation of the vascular extracellular superoxide dismutase by nitric oxide and exercise training. J. Clin. Invest. 2000, 105, 1631–1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, S.M.; Holian, A. Asymmetric dimethylarginine induces oxidative and nitrosative stress in murine lung epithelial cells. Am. J. Respir. Cell Mol. Biol. 2007, 36, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Bakır, M.; Geyikoglu, F.; Colak, S.; Turkez, H.; Bakır, T.O.; Hosseinigouzdagani, M. The carvacrol ameliorates acute pancreatitis-induced liver injury via antioxidant response. Cytotechnology 2016, 68, 1131–1146. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.T.; Li, H.J.; Zhou, L.S. Protective effects of prostaglandin E1 on human umbilical vein endothelial cell injury induced by hydrogen peroxide. Acta Pharmacol. Sin. 2010, 31, 485–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosen, M.B.; Islam, M.R.; Begum, F.; Kabir, Y.; Howlader, M.Z. Oxidative stress induced sperm DNA damage, a possible reason for male infertility. Iran J. Reprod. Med. 2015, 13, 525–532. [Google Scholar] [PubMed]
- Berndt, C.; Lillig, C.H. Glutathione, glutaredoxins, and iron. Antioxid. Redox. Signal. 2017, 27, 1235–1251. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, M.; Zhang, Y.; Huo, T.; Fang, Y.; Jiao, X.; Yuan, M.; Jiang, H. Effects of realgar on GSH synthesis in the mouse hippocampus: Involvement of system XAG(-), system XC(-), MRP-1 and Nrf2. Toxicol. Appl. Pharmacol. 2016, 308, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Iwayama, K.; Kusakabe, A.; Ohtsu, K.; Nawano, T.; Tatsunami, R.; Ohtaki, K.I.; Tampo, Y.; Hayase, N. Long-term treatment of clarithromycin at a low concentration improves hydrogen peroxide-induced oxidant/antioxidant imbalance in human small airway epithelial cells by increasing Nrf2 mRNA expression. BMC Pharmacol. Toxicol. 2017, 18, 15. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.P.; Tang, J.Y.; Xu, Z.; Han, P.; Qin, Z.Q.; Yang, C.D.; Wang, S.Q.; Tang, M.; Wang, W.; Qin, C.; et al. Sulforaphane attenuates di-N-butylphthalate-induced reproductive damage in pubertal mice: Involvement of the Nrf2-antioxidant system. Environ. Toxicol. 2017, 32, 1908–1917. [Google Scholar] [CrossRef] [PubMed]
- Warner, H.R. Superoxide dismutase, aging, and degenerative disease. Free Radic. Biol. Med. 1994, 17, 249–258. [Google Scholar] [CrossRef]
- Wang, F.; Liu, J.; Zhou, L.; Pan, G.; Li, Z.; Zaidi, S.H.; Cheng, F. Senescence-specific change in ROS scavenging enzyme activities and regulation of various SOD isozymes to ROS levels in psf mutant rice leaves. Plant Physiol. Biochem. 2016, 109, 248–261. [Google Scholar] [CrossRef] [PubMed]
- Kosenko, E.A.; Tikhonova, L.A.; Alilova, G.A.; Montoliu, C.; Barreto, G.E.; Aliev, G.; Kaminsky, Y.G. Portacaval shunting causes differential mitochondrial superoxide production in brain regions. Free Radic. Biol. Med. 2017, 113, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Folbergrová, J.; Ješina, P.; Nůsková, H.; Houštěk, J. Antioxidant enzymes in cerebral cortex of immature rats following experimentally-induced seizures: Upregulation of mitochondrial MnSOD (SOD2). Int. J. Dev. Neurosci. 2013, 31, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Selvaratnam, J.S.; Robaire, B. Effects of aging and oxidative stress on spermatozoa of superoxide-dismutase 1- and catalase-null mice. Biol. Reprod. 2016, 95, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Chen, Y.; Ou, L.; Xu, Y.; Yu, X. Aqueous root extract of Asparagus cochinchinensis (Lour.) Merr. Has antioxidant activity in d-galactose-induced aging mice. BMC Complement. Altern. Med. 2017, 17, 469. [Google Scholar] [CrossRef] [PubMed]
- Sue, Y.M.; Cheng, C.F.; Chang, C.C.; Chou, Y.; Chen, C.H.; Juan, S.H. Antioxidation and anti-inflammation by haem oxygenase-1 contribute to protection by tetramethylpyrazine against gentamicin-induced apoptosis in murine renal tubular cells. Nephrol. Dial. Transplant. 2009, 24, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.; Cao, J.; Wu, C.F.; Kadioglu, O.; Schüffler, A.; Kauhl, U.; Klauck, S.M.; Opatz, T.; Thines, E.; Paul, N.W.; Efferth, T. The Chinese herbal formula Free and Easy Wanderer ameliorates oxidative stress through KEAP1-NRF2/HO-1 pathway. Sci. Rep. 2017, 7, 11551. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Lei, L.; Li, B.; Lin, Y.; Xiao, X.; Tang, S. Involvement of the activation of Nrf2/HO-1, p38 MAPK signaling pathways and endoplasmic reticulum stress in furazolidone induced cytotoxicity and S phase arrest in human hepatocyte L02 cells: Modulation of curcumin. Toxicol. Mech. Method 2017, 27, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Y.; Xiao, J.; Suo, H.Y. Screening and identification of resistant lactobacillus in traditional pickles. Food Mach. 2017, 7, 26–30. [Google Scholar]
- Zhao, X.; Song, J.L.; Yi, R.; Li, G.; Sun, P.; Park, K.Y.; Suo, H. Comparison of antioxidative effects of Insect tea and its raw tea (Kuding tea) polyphenols in Kunming mice. Molecules 2018, 23, 204. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhou, X.; Chen, B.; Long, X.; Mu, J.; Pan, Y.; Song, J.L.; Zhao, X.; Yang, Z. Preventive effect of Lactobacillus plantarum CQPC10 on activated carbon induced constipation in Institute of Cancer Research (ICR) mice. Appl. Sci. 2018, 8, 1498. [Google Scholar] [CrossRef]
- Li, G.J.; Wang, J.; Cheng, Y.J.; Tan, X.; Zhai, Y.L.; Wang, Q.; Gao, F.J.; Liu, G.L.; Zhao, X.; Wang, H. Prophylactic effects of polymethoxyflavone-rich orange peel oil on Nω-Nitro-L-arginine-induced hypertensive rats. Appl. Sci. 2018, 8, 752. [Google Scholar] [CrossRef]
- Vinusha, K.S.; Deepika, K.; Johnson, T.S.; Agrawal, G.K.; Rakwal, R. Proteomic studies on lactic acid bacteria: A review. Biochem. Biophys. Rep. 2018, 14, 140–148. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the microbial strains are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Thymus Index | Brain Index | Cardiac Index | Liver Index | Spleen Index | Kidney Index |
|---|---|---|---|---|---|---|
| Normal | 0.25 ± 0.02 a | 1.91 ± 0.04 a | 5.02 ± 0.06 a | 38.21 ± 0.52 a | 4.12 ± 0.13 a | 12.89 ± 0.39 a |
| DGT | 0.14 ± 0.02 d | 1.17 ± 0.07 d | 4.12 ± 0.05 e | 30.32 ± 1.32 e | 2.67 ± 0.28 e | 9.22 ± 0.27 e |
| DGT + LDSB | 0.18 ± 0.02 c | 1.48 ± 0.06 c | 4.35 ± 0.08 d | 33.15 ± 0.46 d | 3.28 ± 0.12 d | 10.03 ± 0.26 d |
| DGT + LP-CQPC11 | 0.22 ± 0.01 b | 1.79 ± 0.06 b | 4.81 ± 0.07 b | 35.73 ± 0.67 b | 3.77 ± 0.15 b | 12.11 ± 0.20 b |
| DGT + Vc | 0.19 ± 0.02 c | 1.54 ± 0.07 c | 4.62 ± 0.07 c | 34.39 ± 0.47 c | 3.49 ± 0.11 c | 11.46 ± 0.25 c |
| Group | NO (μmol/L) | SOD (U/mL) | GSH-Px (U/mL) | GSH (mg/L) | MDA (nmol/mL) |
|---|---|---|---|---|---|
| Normal | 18.79 ± 0.45 e | 275.36 ± 8.71 a | 182.36 ± 7.87 a | 37.33 ± 0.52 a | 5.57 ± 0.35 e |
| DGT | 63.69 ± 1.12 a | 97.63 ± 8.16 e | 103.55 ± 6.89 e | 14.02 ± 1.13 e | 29.35 ± 1.03 a |
| DGT + LDSB | 42.72 ± 1.02 b | 203.56 ± 9.71 d | 119.71 ± 4.38 d | 18.33 ± 0.55 d | 20.45 ± 0.59 b |
| DGT + LP-CQPC11 | 30.55 ± 0.67 d | 147.83 ± 7.89 b | 152.03 ± 5.88 b | 29.42 ± 0.63 b | 10.32 ± 0.38 d |
| DGT + Vc | 38.32 ± 0.66 c | 178.36 ± 6.25 c | 133.20 ± 6.34 c | 23.57 ± 0.61 c | 16.74 ± 0.68 c |
| Group | NO (μmol/gprot) | SOD (U/mgprot) | GSH-Px (U/mgprot) | GSH (mg/gprot) | MDA (nmol/mgprot) |
|---|---|---|---|---|---|
| Normal | 3.21 ± 0.15 e | 95.18 ± 5.32 a | 163.21 ± 7.91 a | 8.32 ± 0.28 a | 2.11 ± 0.19 e |
| DGT | 9.45 ± 0.41 a | 31.02 ± 2.65 e | 91.03 ± 4.53 e | 2.65 ± 0.29 e | 9.39 ± 0.42 a |
| DGT + LDSB | 7.16 ± 0.33 b | 48.63 ± 4.61 d | 113.25 ± 5.33 d | 4.32 ± 0.25 d | 6.41 ± 0.33 b |
| DGT + LP-CQPC11 | 4.85 ± 0.12 d | 74.36 ± 5.01 b | 140.73 ± 3.25 b | 6.12 ± 0.22 b | 3.89 ± 0.21 d |
| DGT + Vc | 5.74 ± 0.20 c | 60.87 ± 4.33 c | 127.18 ± 4.25 c | 5.39 ± 0.28 c | 4.77 ± 0.18 c |
| Group | NO (μmol/gprot) | SOD (U/mgprot) | GSH-Px (U/mgprot) | GSH (mg/gprot) | MDA (nmol/mgprot) |
|---|---|---|---|---|---|
| Normal | 1.49 ± 0.15 e | 70.87 ± 3.26 a | 115.36 ± 4.22 a | 5.89 ± 0.18 a | 1.03 ± 0.06 e |
| DGT | 7.41 ± 0.28 a | 22.36 ± 4.12 e | 44.31 ± 3.55 e | 1.52 ± 0.23 e | 4.24 ± 0.16 a |
| DGT + LDSB | 5.88 ± 0.24 c | 45.97 ± 3.87 d | 60.12 ± 2.88 d | 2.74 ± 0.21 d | 3.11 ± 0.15 b |
| DGT + LP-CQPC11 | 2.28 ± 0.14 d | 61.25 ± 2.02 b | 87.71 ± 3.91 b | 4.12 ± 0.24 b | 1.62 ± 0.10 d |
| DGT + Vc | 3.31 ± 0.20 c | 51.88 ± 2.31 c | 69.78 ± 3.02 c | 3.65 ± 0.22 c | 2.41 ± 0.11 c |
| Gene Name | Sequence |
|---|---|
| nNOS | Forward: 5′-ACGGCAAACTGCACAAAGC-3′ |
| Reverse: 5′-CGTTCTCTGAATACGGGTTGTTG-3′ | |
| eNOS | Forward: 5′-TCAGCCATCACAGTGTTCCC-3′ |
| Reverse: 5′-ATAGCCCGCATAGCGTATCAG-3′ | |
| iNOS | Forward: 5′-GTTCTCAGCCCAACAATACAAGA-3′ |
| Reverse: 5′-GTGGACGGGTCGATGTCAC-3′ | |
| Cu/Zn-SOD | Forward: 5′-AACCAGTTGTGTTGTCAGGAC-3′ |
| Reverse: 5′-CCACCATGTTTCTTAGAGTGAGG-3′ | |
| Mn-SOD | Forward: 5′-CAGACCTGCCTTACGACTATGG-3′ |
| Reverse: 5′-CTCGGTGGCGTTGAGATTGTT-3′ | |
| CAT | Forward: 5′- GGAGGCGGGAACCCAATAG -3′ |
| Reverse: 5′-GTGTGCCATCTCGTCAGTGAA-3′ | |
| HO-1 | Forward: 5′-ACAGATGGCGTCACTTCG-3′ |
| Reverse: 5′-TGAGGACCCACTGGAGGA-3′ | |
| Nrf2 | Forward: 5′-CAGTGCTCCTATGCGTGAA-3′ |
| Reverse: 5′-GCGGCTTGAATGTTTGTC-3′ | |
| γ-GCS | Forward: 5′-GCACATCTACCACGCAGTCA-3′ |
| Reverse: 5′-CAGAGTCTCAAGAACATCGCC-3′ | |
| NQO1 | Forward: 5′-CTTTAGGGTCGTCTTGGC-3′ |
| Reverse: 5′-CAATCAGGGCTCTTCTCG-3′ | |
| GAPDH | Forward: 5′-AGGTCGGTGTGAACGGATTTG-3′ |
| Reverse: 5′-GGGGTCGTTGATGGCAACA-3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qian, Y.; Zhang, J.; Zhou, X.; Yi, R.; Mu, J.; Long, X.; Pan, Y.; Zhao, X.; Liu, W. Lactobacillus plantarum CQPC11 Isolated from Sichuan Pickled Cabbages Antagonizes d-galactose-Induced Oxidation and Aging in Mice. Molecules 2018, 23, 3026. https://doi.org/10.3390/molecules23113026
Qian Y, Zhang J, Zhou X, Yi R, Mu J, Long X, Pan Y, Zhao X, Liu W. Lactobacillus plantarum CQPC11 Isolated from Sichuan Pickled Cabbages Antagonizes d-galactose-Induced Oxidation and Aging in Mice. Molecules. 2018; 23(11):3026. https://doi.org/10.3390/molecules23113026
Chicago/Turabian StyleQian, Yu, Jing Zhang, Xianrong Zhou, Ruokun Yi, Jianfei Mu, Xingyao Long, Yanni Pan, Xin Zhao, and Weiwei Liu. 2018. "Lactobacillus plantarum CQPC11 Isolated from Sichuan Pickled Cabbages Antagonizes d-galactose-Induced Oxidation and Aging in Mice" Molecules 23, no. 11: 3026. https://doi.org/10.3390/molecules23113026