Identification and Quantification of Flavanol Glycosides in Vitis vinifera Grape Seeds and Skins during Ripening


Abstract
:1. Introduction
2. Results and Discussion
2.1. Identification of Flavanol Glycosides in Grape Skins and Seeds
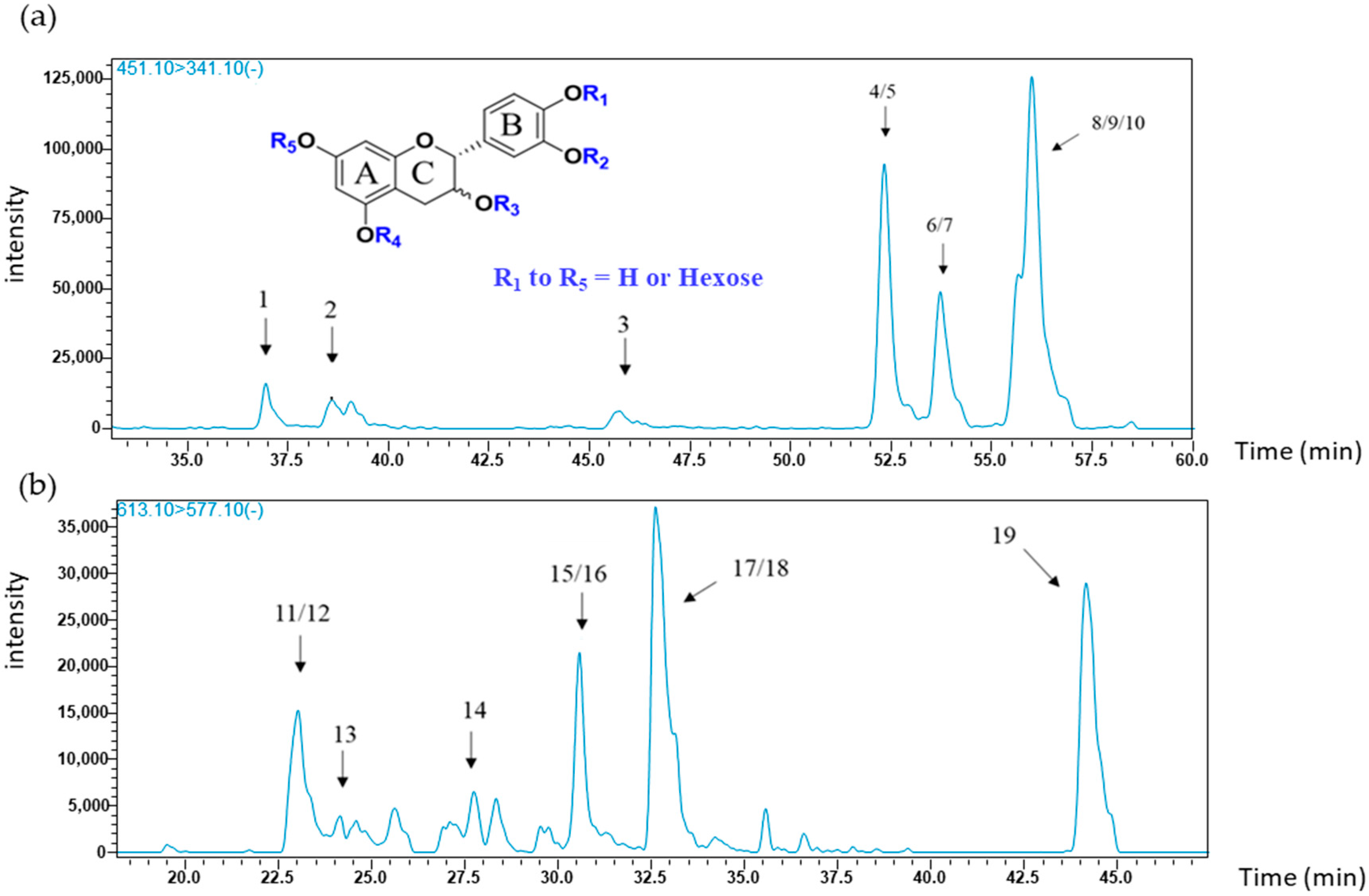
2.1.1. Flavanol Glycoside Monomers
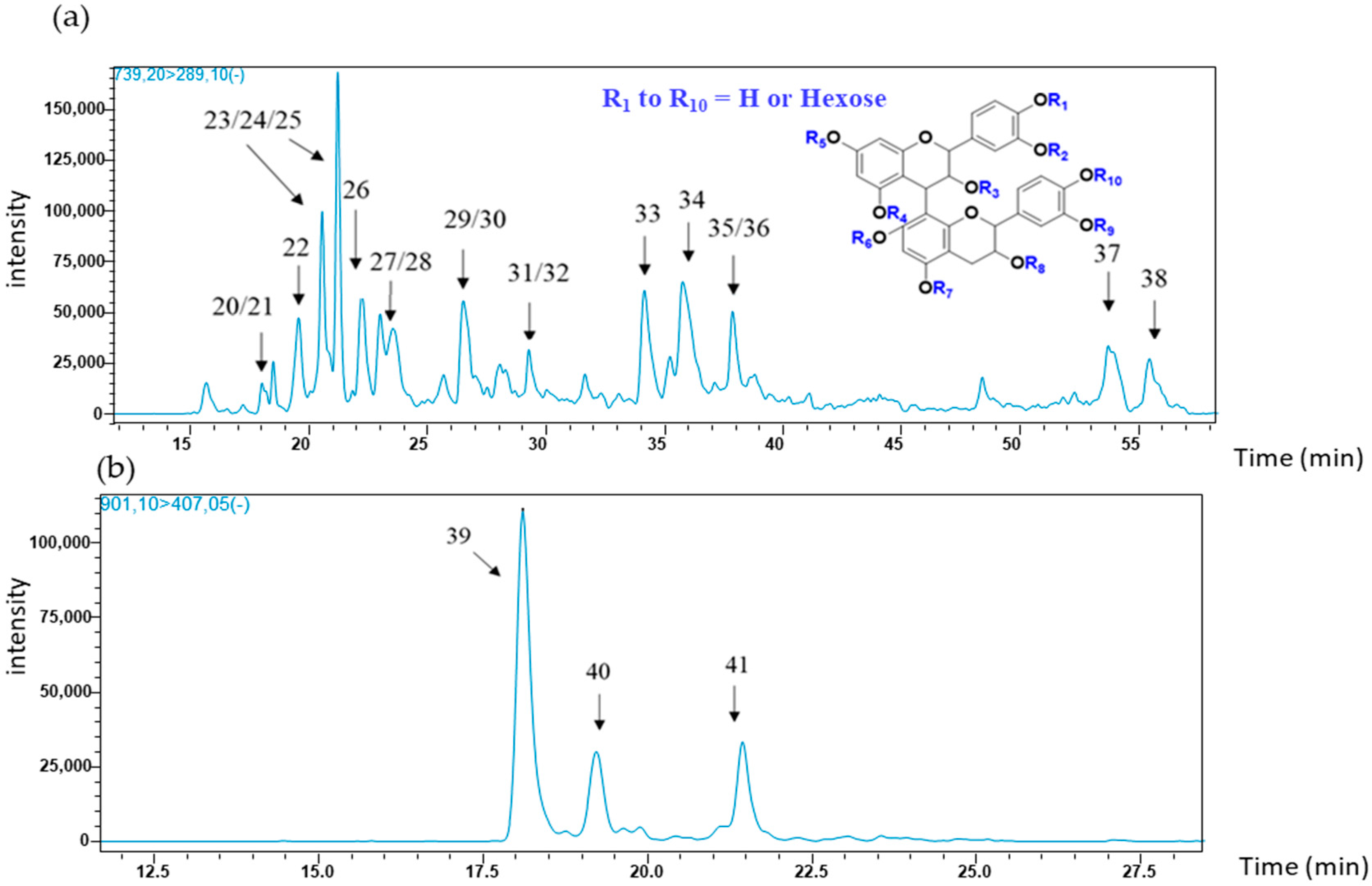
2.1.2. Flavanol Glycoside Dimers
2.2. Quantification of Flavanol Glycosides in Syrah, Tannat and Merlot Grape Skins and Seeds during Ripening
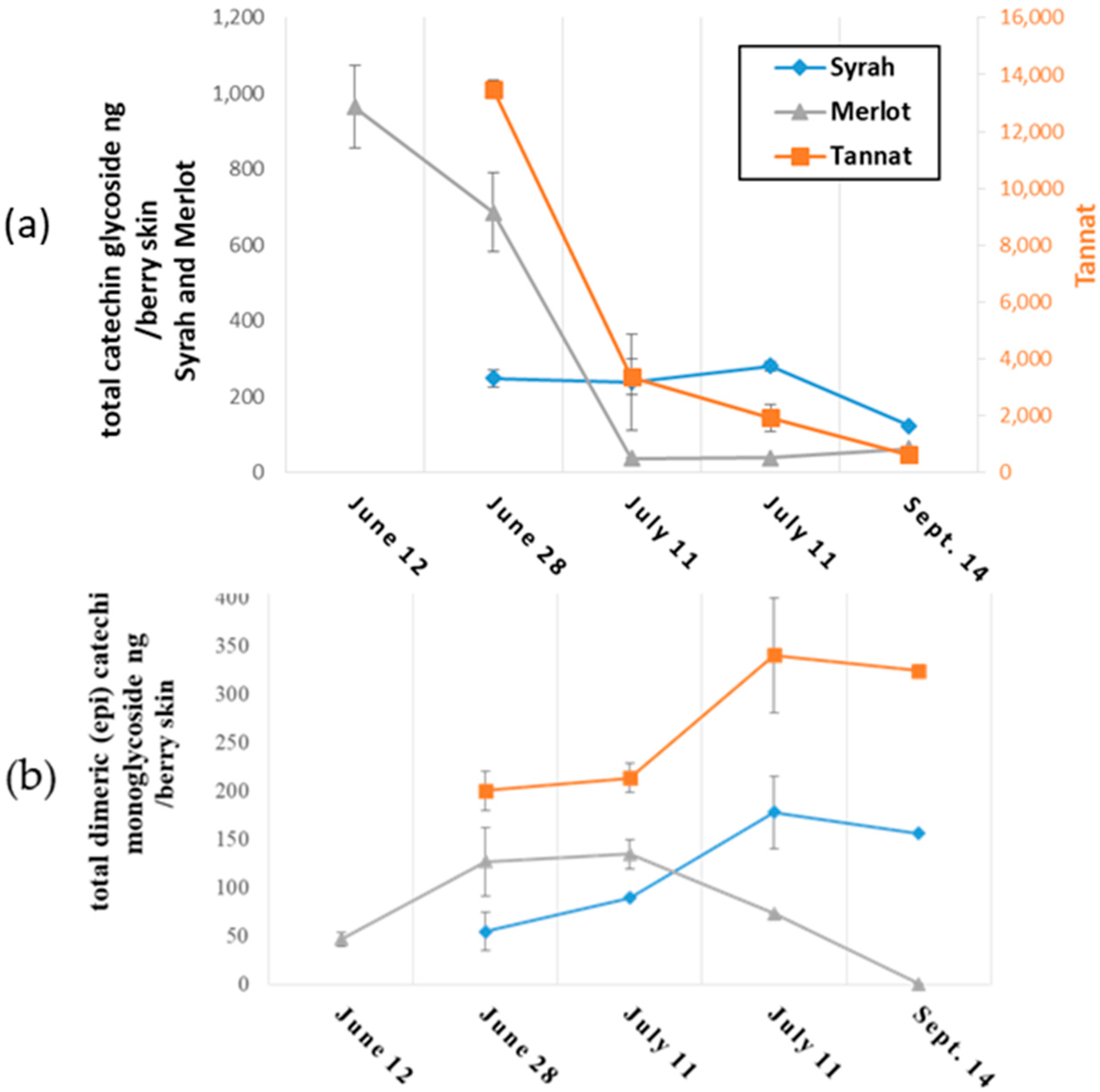
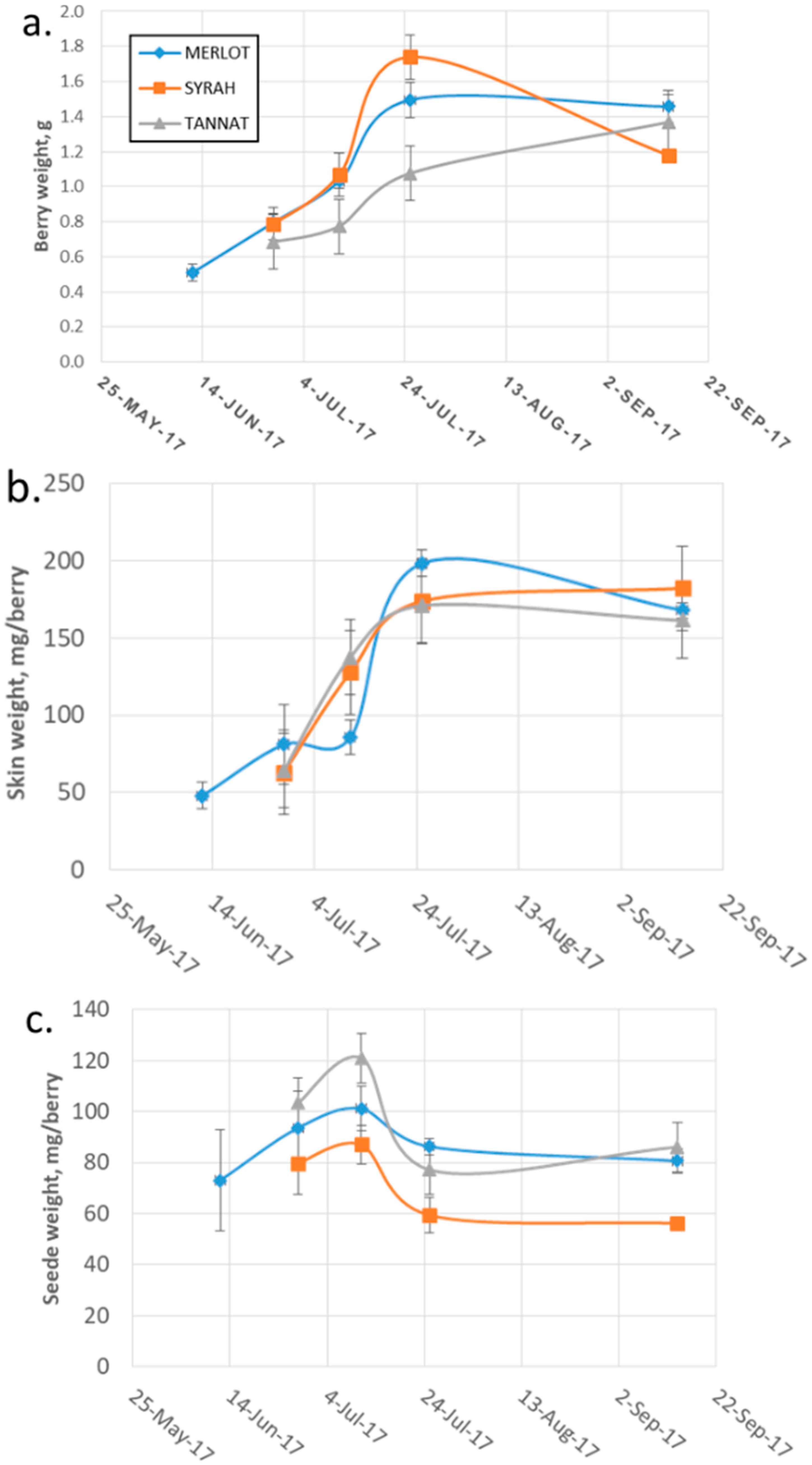
2.2.1. Changes in Monomeric and Oligomeric Flavanol Glycosides in Grape Skins during Ripening
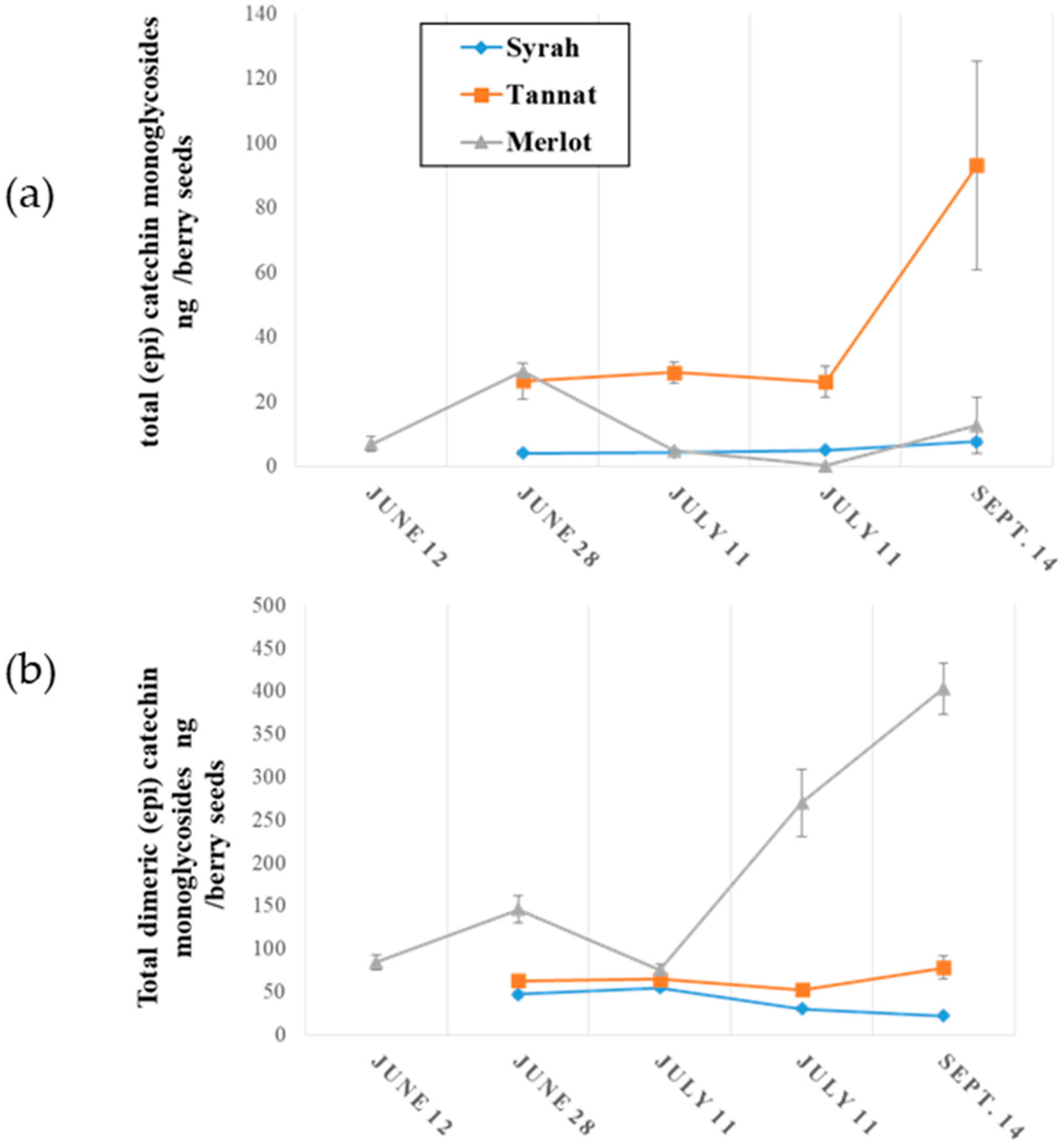
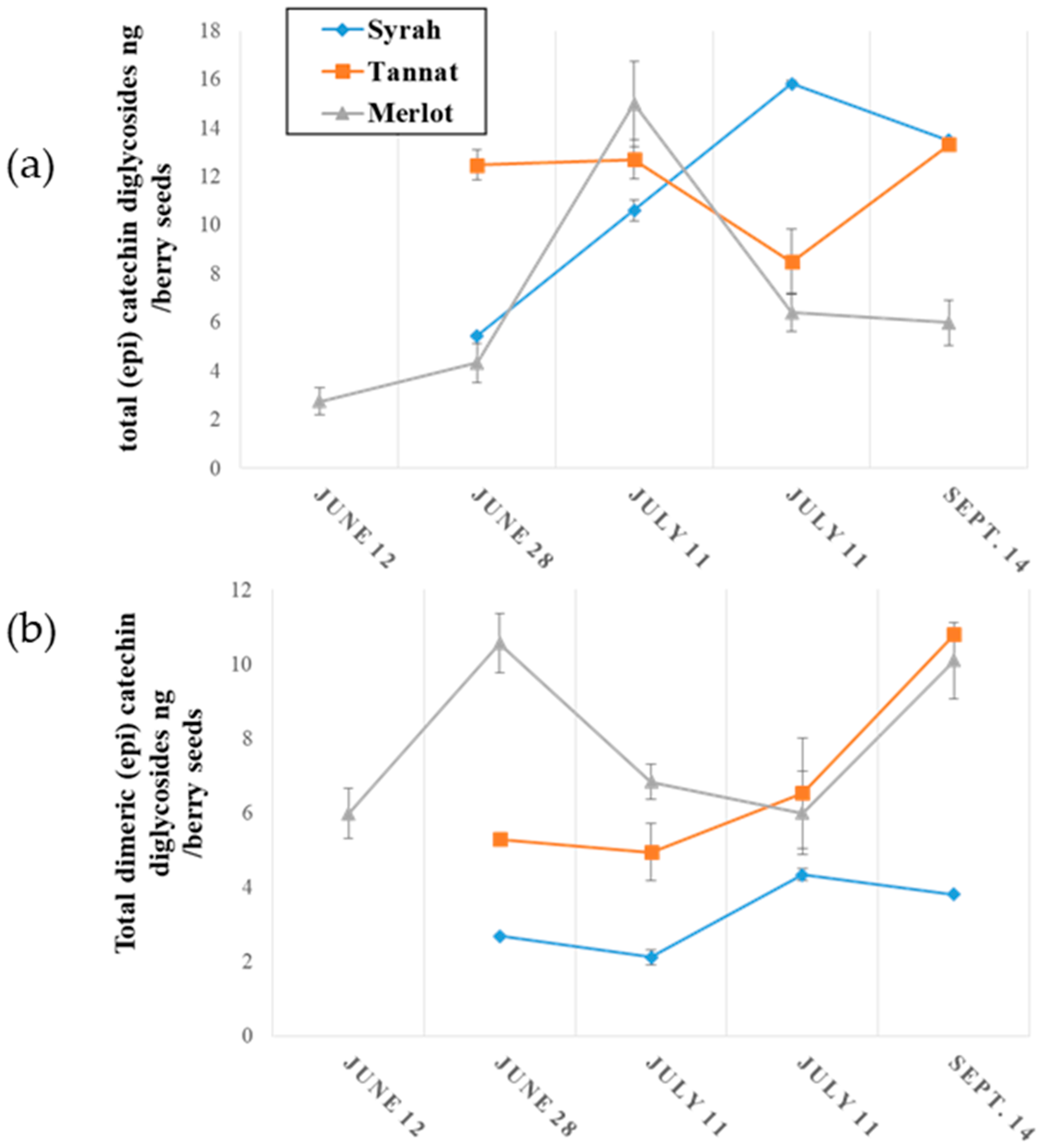
2.2.2. Evolution of Monomeric and Oligomeric Flavanol Glycosides in Grape Seeds during Ripening
3. Materials and Methods
3.1. Reagents, Standards and Calibration
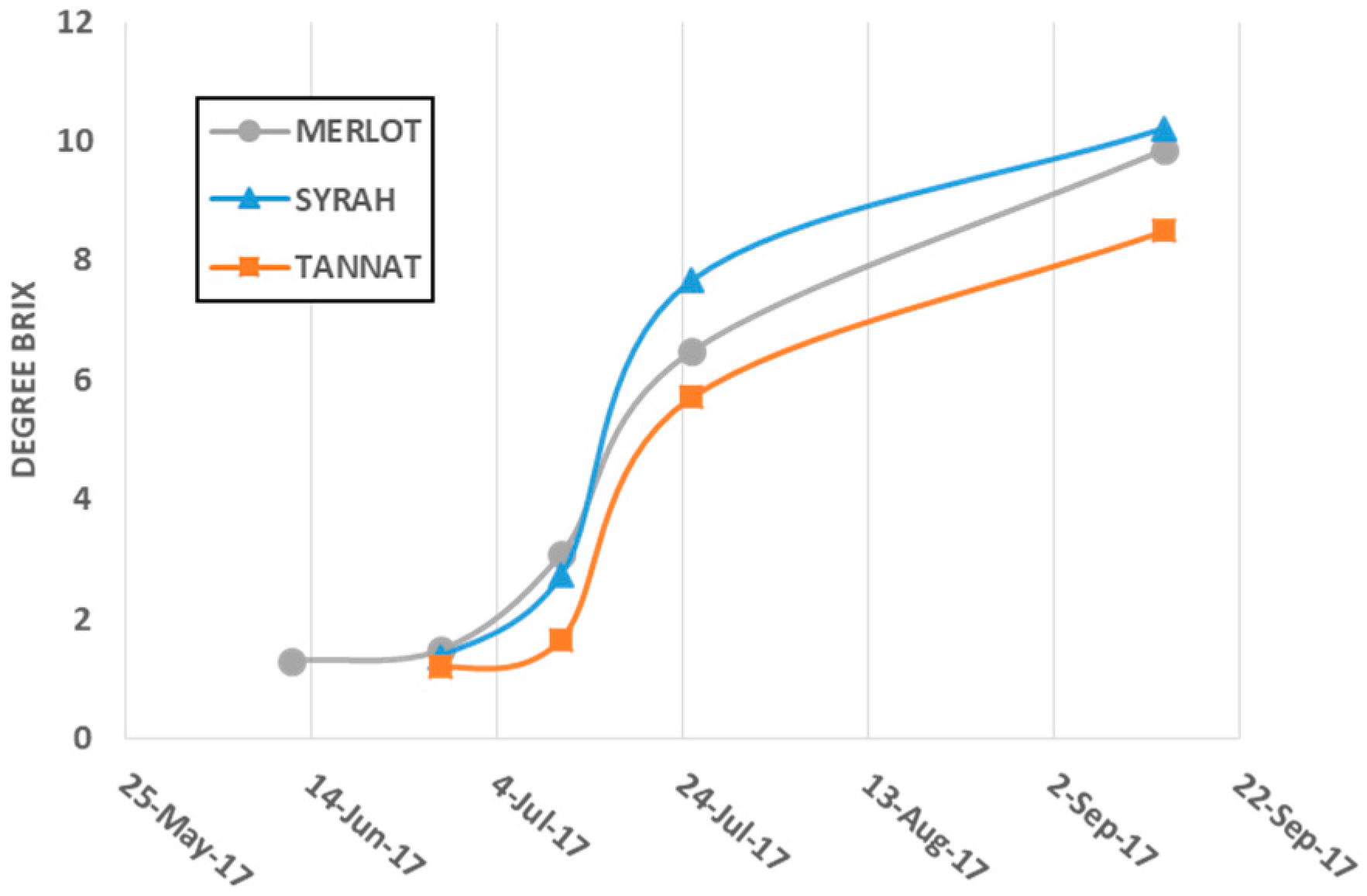
3.2. Grapes
3.2.1. Samples used for Quantification of Flavanol Glycosides in Berry Skin and Seeds
3.2.2. Extraction of Grape Polyphenols
3.3. Optimization of a UHPLC-ESI-QQQ-MS/MS in Multiple Reaction Monitoring (MRM) Method for Quantification of Flavanol Glycosides
3.3.1. UHPLC Analysis
3.3.2. MS/MS Detection
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouysegu, L. Plant polyphenols: Chemical properties, biological activities, and synthesis. Angew. Chem. Int. Ed. 2011, 50, 586–621. [Google Scholar] [CrossRef] [PubMed]
- Heiss, C.; Keen, C.L.; Kelm, M. Flavanols and cardiovascular disease prevention. Eur. Heart J. 2010, 31, 2583–2592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chira, K.; Suh, J.H.; Saucier, C.; Teissèdre, P.L. Les polyphénols du raisin. Phytothérapie 2008, 6, 75–82. [Google Scholar] [CrossRef]
- Di Lecce, G.; Arranz, S.; Jáureguic, O.; Tresserra-Rimbau, A.; Quifer-Rada, P.; Lamuela-Raventós, R.M. Phenolic profiling of the skin, pulp and seeds of Albariño grapes using hybrid quadrupole time-of-flight and triple-quadrupole mass spectrometry. Food Chem. 2014, 145, 874–882. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.A.; Matthews, M.A.; Waterhouse, A.L. Changes in grape seed polyphenols during fruit ripening. Phytochemistry 2000, 55, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Prieur, C.; Rigaud, J.; Cheynier, V.; Moutounet, M. Oligomeric and polymeric procyanidins from grape seeds. Phytochemistry 1994, 36, 781–784. [Google Scholar] [CrossRef]
- Downey, M.O.; Harvey, J.S.; Robinson, S.P. Analysis of tannins in seeds and skins of Shiraz grapes throughout berry development. Aust. J. Grape Wine R. 2003, 9, 15–27. [Google Scholar] [CrossRef]
- Peyrot des Gachons, C.; Kennedy, J.A. Direct Method for Determining Seed and Skin Proanthocyanidin Extraction into Red Wine. J. Am. Chem. Soc. 2003, 51, 5877–5881. [Google Scholar] [CrossRef] [PubMed]
- Monagas, M.; Gómez-Cordovés, C.; Bartolomé, B.; Laureano, O.; Ricardo da Silva, J.M. Monomeric, Oligomeric, and Polymeric Flavan-3-ol Composition of Wines and Grapes from Vitis vinifera L. Cv. Graciano, Tempranillo, and Cabernet Sauvignon. J. Agric. Food Chem. 2003, 51, 6475–6481. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.F.; Cheynier, V.; Terrier, N. Shedding Light on the Black Boxes of the Proanthocyanidin Pathway with Grapevine. Recent Adv. Polyphenol Res. 2012, 3, 161–190. [Google Scholar]
- Kuhnle, G.G.C. Nutrition epidemiology of flavan-3-ols: The known unknowns. Mol. Asp. Med. 2018, 61, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Aron, P.M.; Kennedy, J.A. Flavan-3-ols: Nature, occurrence and biological activity. Mol. Nutr. Food Res. 2008, 52, 79–104. [Google Scholar] [CrossRef] [PubMed]
- Corder, R.; Mullen, W.; Khan, N.Q.; Marks, S.C.; Wood, E.G.; Carrier, M.J.; Crozier, A. Oenology: Red wine procyanidins and vascular health. Nature 2006, 444, 566. [Google Scholar] [CrossRef] [PubMed]
- Manach, C.; Donovan, J.L. Pharmacokinetics and metabolism of dietary flavonoids in humans. Free Radic. Res. 2004, 38, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Kitao, S.; Ariga, T.; Matsudo, T.; Sekine, H. The Syntheses of Catechin-glucosides by Transglycosylation with Leuconostoc mesenteroides Sucrose Phosphorylase. Biosci. Biotechnol. Biochem. 1993, 57, 2010–2015. [Google Scholar] [CrossRef]
- Raab, T.; Barron, D.; Arce Vera, F.; Crespy, V.; Oliveira, M.; Williamson, G. Catechin Glucosides: Occurrence, Synthesis, and Stability. Food Chem. 2010, 58, 2138–2149. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.; Peel, G.J.; Sharma, S.B.; Tang, Y.; Dixon, R.A. A transcript profiling approach reveals an epicatechin-specific glucosyltransferase expressed in the seed coat of Medicago truncatula. Proc. Natl. Acad. Sci. USA 2008, 105, 14210–14215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Y.; Cheng, X.; Huhman, D.V.; Ma, J.; Peel, G.J.; Yonekura-Sakakibara, K.; Saito, K.; Shen, G.; Sumner, L.; Tang, Y.; et al. Medicago glucosyltransferase UGT72L1: Potential roles in proanthocyanidin biosynthesis. Planta 2013, 238, 139–154. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, W.; Galensa, R. Identification of a new flavanol glucoside from barley (Hordeum vulgar L.) and malt. Eur. Food Res. Technol. 2002, 214, 388–393. [Google Scholar] [CrossRef]
- Watanabe, M. Catechins as Antioxidants from Buckwheat (Fagopyrum esculentum Moench) Groats. J. Agric. Food Chem. 1998, 46, 839–845. [Google Scholar] [CrossRef]
- Dueñas, M.; Sun, B.; Hernández, T.; Estrella, I.; Spranger, M.I. Proanthocyanidin Composition in the Seed Coat of Lentils (Lens culinaris L.). J. Agric. Food Chem. 2003, 51, 7999–8004. [Google Scholar] [CrossRef] [PubMed]
- Cui, E.-J.; Song, N.-A.; Shrestha, S.; Chung, I.-S.; Kim, J.-Y.; Jeong, T.-S.; Baek, N.-I. Flavonoid glycosides from cowpea seeds (Vigna sinensis K.) inhibit LDL. Food Sci. Biotechnol. 2012, 21, 619–624. [Google Scholar] [CrossRef]
- Ojwang, L.O.; Yang, L.; Dykes, L.; Awika, J. Proanthocyanidin profile of cowpea (Vigna unguiculata) reveals catechin-O-glucoside as the dominant compound. Food Chem. 2013, 139, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Smaali, I.; Maugard, T.; Limam, F.; Legoy, M.D.; Marzouki, N. Efficient synthesis of gluco-oligosaccharides and alkyl-glucosides by. World J. Microb. Biotechnol. 2007, 23, 145–149. [Google Scholar] [CrossRef]
- Delcambre, A.; Saucier, C. Identification of new flavan-3-ol monoglycosides by UHPLC-ESI-Q-TOF in grapes and wine. J. Mass Spectrom. 2012, 47, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Bae, Y.S.; Burger, J.F.W.; Steynberg, J.P.; Ferreira, D.; Hemingway, R.W. Flavan and procyanidin glycosides from the bark of blackjack oak. Phytochemistry 1994, 35, 473–478. [Google Scholar] [CrossRef] [Green Version]
- Zerbib, M.; Mazauric, J.P.; Meudec, E.; Le Guernevé, C.; Lepak, A.; Nidetzky, B.; Cheynier, V.; Terrier, N.; Saucier, C. New flavanol O-glycosides in grape and wine. Food Chem. 2018, 266, 441–448. [Google Scholar] [CrossRef]
- Pinasseau, L.; Verbaere, A.; Roques, M.; Meudec, E.; Vallverdú-Queralt, A.; Terrier, N.; Boulet, J.C.; Cheynier, V.; Sommerer, N. A Fast and Robust UHPLC-MRM-MS Method to Characterize and Quantify Grape Skin Tannins after Chemical Depolymerization. Molecules 2016, 21, 1409. [Google Scholar] [CrossRef] [PubMed]
- Ehrhardt, C.; Arapitsas, P.; Stefanini, M.; Flick, G.; Mattivi, F. Analysis of the phenolic composition of fungus-resistant grape varieties cultivated in Italy and Germany using UHPLC-MS/MS-Ehrhardt-2014-Journal of Mass Spectrometry-Wiley Online Library. J. Mass Spectrom. 2014, 49, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Owczarek, A.; Magiera, A.; Matczak, M.; Piotrowska, D.G.; Olszewska, M.A.; Marchelak, A. Optimisation of preparative HPLC separation of four isomeric kaempferol diglycosides from Prunus spinosa L. by application of the response surface methodology. Phytochem. Lett. 2017, 20, 415–424. [Google Scholar] [CrossRef]
- Garrido, J.; Borges, F. Wine and grape polyphenols—A chemical perspective. Food Res. Int. 2013, 54, 1844–1858. [Google Scholar] [CrossRef]
- Portugal Gomes Rebello, L.; Lago-Vanzela, E.S.; Barcia Teixeira, M.; Ramos Mota, A.; Stringheta, P.C.; Da-Silva, R.; Castillo-Muñoz, N.; Gómez-Alonso, S.; Hermosín-Gutiérrez, I. Phenolic composition of the berry parts of hybrid grape cultivar BRS Violeta (BRS Rubea×IAC 1398-21) using HPLC–DAD–ESI-MS/MS. Food Res. Int. 2013, 54, 354–366. [Google Scholar] [CrossRef]
- Bishop, P.D.; Nagel, C.W. Characterization of the condensation product of malvidin diglucoside and catechin. J. Agric. Food Chem. 1984, 32, 1022–1026. [Google Scholar] [CrossRef]
- Guo, D.; Xu, Y.; Kang, Y.; Han, S.; Zheng, S. Synthesis of octyl-beta-d-glucopyranoside catalyzed by Thai rosewood beta-glucosidase-displaying Pichia pastoris in an aqueous/organic two-phase system. Enzym. Microb. Technol. 2016, 85, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, Y.; Toledo, R.T. Major Flavonoids in Grape Seeds and Skins: Antioxidant Capacity of Catechin, Epicatechin, and Gallic Acid. J. Agric. Food Chem. 2004, 52, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Chira, K. Structures moléculaire et perception tannique des raisins et des vins (Cabernet-Sauvignon, Merlot) du Bordelais. Ph.D. Thesis, Université de Bordeaux, Bordeaux, France, December 2009. [Google Scholar]
Sample Availability: Not available. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peaks | RT (min) | Tissues, Seeds (se), Skins (sk) | MS/MS Product Ions m/z | Compounds (Corresponding m/z) |
|---|---|---|---|---|
| 1 | 36.95 | se, sk | 177; 289; 341 | (epi) catechin monoglycoside (451) |
| 2 | 38.60 | sk | 177; 341 | |
| 3 | 45.72 | se, sk | 177; 341 | |
| 4/5 | 52.33 | se, sk | 177; 289; 341 | |
| 6/7 | 53.77 | se, sk | 177; 289; 341 | |
| 8/9/10 | 55.99 | se, sk | 177; 341 | |
| 11/12 | 23.33 | se | 577; 289 | (epi) catechin diglycoside (613) |
| 13 | 24.15 | se | ||
| 14 | 27.74 | se | ||
| 15/16 | 30.57 | se | ||
| 17/18 | 32.64 | se | ||
| 19 | 44.17 | se | ||
| 20/21 | 18.48 | se, sk | 177; 287; 289; 451; 577 | (epi) catechin dimer monoglycoside (739) |
| 22 | 19.54 | se | 177; 287; 289; 451; 577 | |
| 23/24/25 | 21.20 | se, sk | 177; 287; 289; 339; 451; 577 | |
| 26 | 22.21 | se, sk | 177; 287; 289; 339; 451; 577 | |
| 27/28 | 23.52 | se, sk | 177; 287; 289; 339; 451; 577 | |
| 29/30 | 26.17 | se | 177; 287; 289; 339; 451; 577 | |
| 31/32 | 28.04 | se | 177; 289; 451; 577 | |
| 33 | 34.13 | se, sk | 177; 287; 289; 339; 451; 577 | |
| 34 | 35.75 | se, sk | 177; 287; 289; 339; 451; 577 | |
| 35/36 | 37.85 | se, sk | 177; 289; 339; 451; 577 | |
| 37 | 48.39 | se | 287; 289; 339; 451 | |
| 38 | 53.73 | se | 287; 289; 339; 451; 577 | |
| 39 | 18.09 | se | 407 | (epi) catechin dimer diglycoside (901) |
| 40 | 19.25 | se | ||
| 41 | 21.43 | se |
| Compound | Precursor Ion | Product Ion | Q1 Prebias | Collision Energy (eV) | Q3 Prebias |
|---|---|---|---|---|---|
| 1 | 451 | 341 | 22 | 18 | 25 |
| 2 | 739 | 289 | 40 | 31 | 26 |
| 3 | 613 | 577 | 30 | 12 | 22 |
| 4 | 901 | 407 | 24 | 49 | 14 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zerbib, M.; Cazals, G.; Enjalbal, C.; Saucier, C. Identification and Quantification of Flavanol Glycosides in Vitis vinifera Grape Seeds and Skins during Ripening. Molecules 2018, 23, 2745. https://doi.org/10.3390/molecules23112745
Zerbib M, Cazals G, Enjalbal C, Saucier C. Identification and Quantification of Flavanol Glycosides in Vitis vinifera Grape Seeds and Skins during Ripening. Molecules. 2018; 23(11):2745. https://doi.org/10.3390/molecules23112745
Chicago/Turabian StyleZerbib, Marie, Guillaume Cazals, Christine Enjalbal, and Cédric Saucier. 2018. "Identification and Quantification of Flavanol Glycosides in Vitis vinifera Grape Seeds and Skins during Ripening" Molecules 23, no. 11: 2745. https://doi.org/10.3390/molecules23112745