
Sialic Acid-Binding Lectin from Bullfrog Eggs Exhibits an Anti-Tumor Effect Against Breast Cancer Cells Including Triple-Negative Phenotype Cells


Abstract
:1. Introduction
2. Results
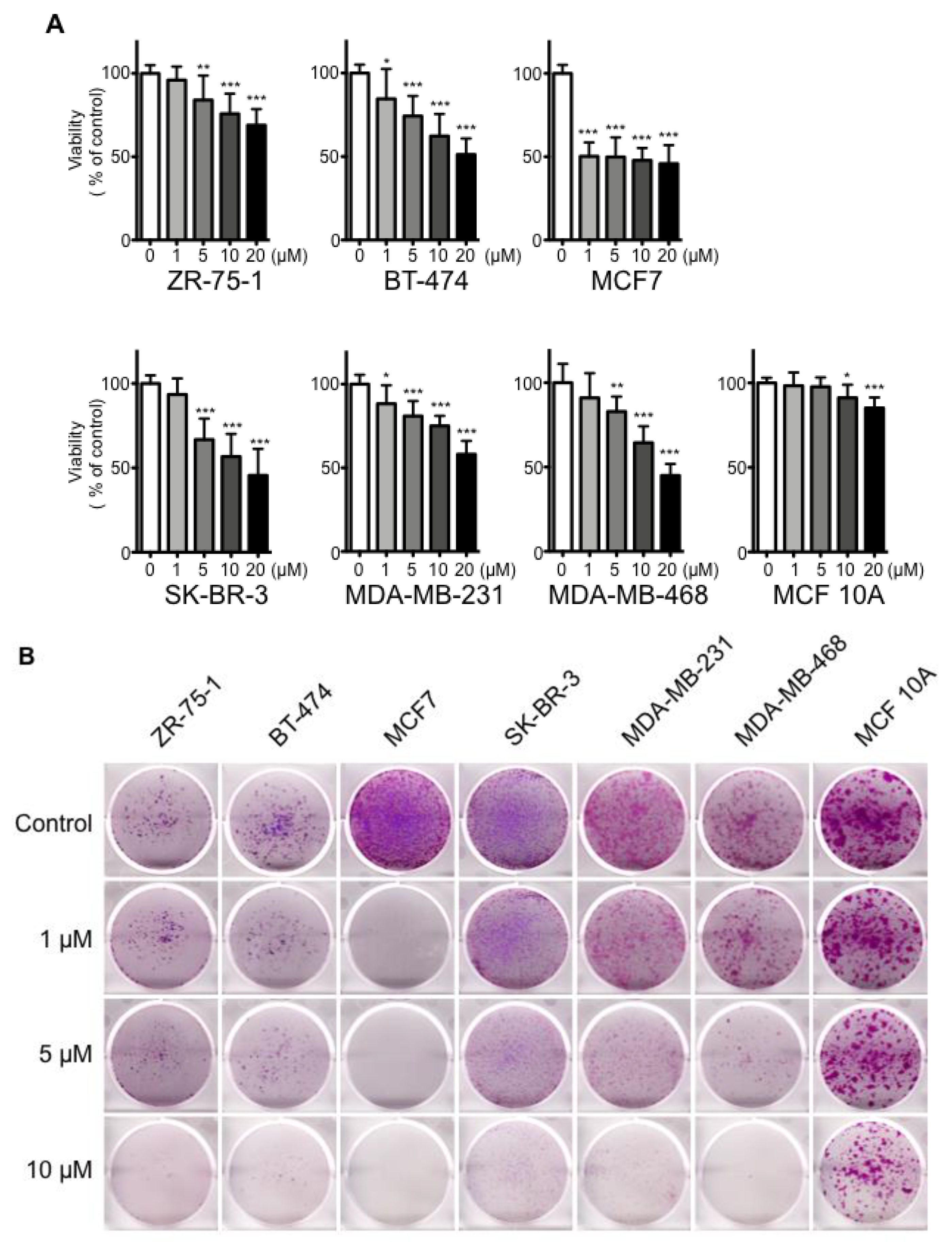
2.1. Effects of cSBL on Breast Cancer Cell Growth
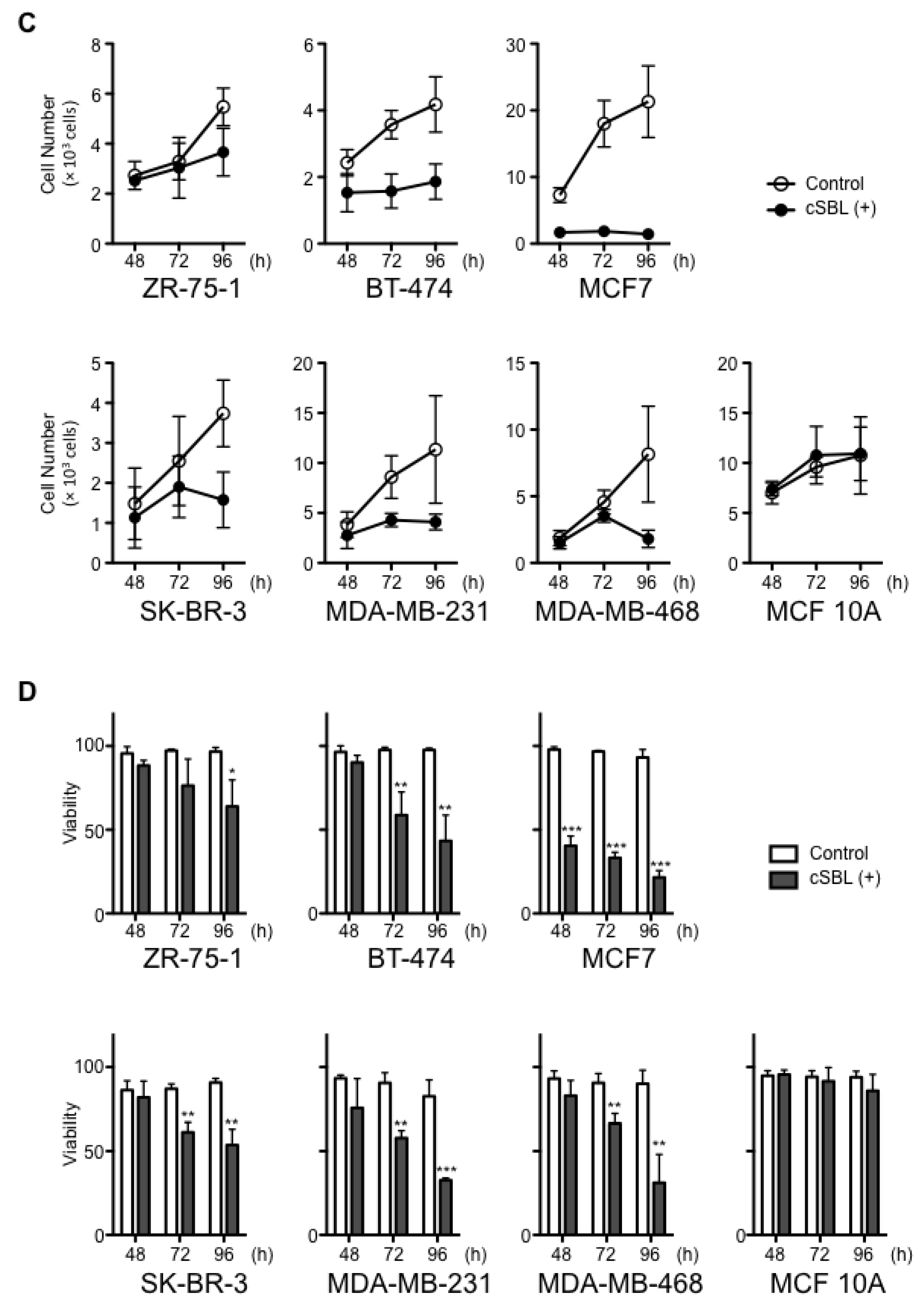
2.2. cSBL-Induced Apoptosis
2.3. Effects of cSBL on Cell Survival-Related Molecules
2.4. Expressions of ErbB Family Protein in Breast Cancer Cells Treated with cSBL
2.5. Expressions of EGFR and Its Signaling in Triple-Negative Breast Cancer Cells Treated with cSBL
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. WST Assays
4.4. Clonogenic Assay
4.5. Trypan Blue Dye Exclusion Assay
4.6. Observation of Nuclear Morphology
4.7. Western Blotting
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yau, T.; Dan, X.; Ng, C.C.W.; Ng, T.B. Lectins with potential for anti-cancer therapy. Molecules 2015, 20, 3791–3810. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Alvarez, J.A.; Krishnan, H.; Shen, Y.; Acharya, N.K.; Han, M.; McNulty, D.E.; Hasegawa, H.; Hyodo, T.; Senga, T.; Geng, J.-G.; et al. Plant Lectin Can Target Receptors Containing Sialic Acid, Exemplified by Podoplanin, to Inhibit Transformed Cell Growth and Migration. PLoS ONE 2012, 7, e41845. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhang, B.; Min, M.; Bian, H.; Chen, L.; Liu, Q.; Bao, J. Induction of apoptosis by Polygonatum odoratum lectin and its molecular mechanisms in murine fibrosarcoma L929 cells. Biochim. Biophys. Acta Gen. Subj. USA/PAN 2009, 1790, 840–844. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wu, L.; Duan, X.; Cui, L.; Luo, J.; Li, G. Adenovirus carrying gene encoding discus sialic acid binding lectin induces cancer cell apoptosis. Mar. Drugs 2014, 12, 3994–4004. [Google Scholar] [CrossRef] [PubMed]
- Miyagi, T.; Wada, T.; Yamaguchi, K.; Shiozaki, K.; Sato, I.; Kakugawa, Y.; Yamanami, H.; Fujiya, T. Human sialidase as a cancer marker. Proteomics 2008, 8, 3303–3311. [Google Scholar] [CrossRef] [PubMed]
- Hakomori, S. Glycosylation defining cancer malignancy: New wine in an old bottle. Proc. Natl. Acad. Sci. USA/PNAS 2002, 99, 10231–10233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dennis, J.W.; Granovsky, M.; Warren, C.E. Glycoprotein glycosylation and cancer progression. Biochim. Biophys. Acta 1999, 1473, 21–34. [Google Scholar] [CrossRef]
- Kawauchi, H.; Sakakibara, F.; Watanabe, K. Agglutinins of frog eggs: A new class of proteins causing preferential agglutination of tumor cells. Experientia 1975, 31, 364–365. [Google Scholar] [CrossRef] [PubMed]
- Nitta, K.; Takayanagi, G.; Kawauchi, H.; Hakomori, S. Isolation and characterization of Rana catesbeiana lectin and demonstration of the lectin-binding glycoprotein of rodent and human tumor cell membranes. Cancer Res. 1987, 47, 4877–4883. [Google Scholar] [PubMed]
- Okabe, Y.; Katayama, N.; Iwama, M.; Watanabe, H.; Ohgi, K.; Irie, M.; Nitta, K.; Kawauchi, H.; Takayanagi, Y.; Oyama, F. Comparative base specificity, stability, and lectin activity of two lectins from eggs of Rana catesbeiana and R. japonica and liver ribonuclease from R. catesbeiana. J. Biochem. 1991, 109, 786–790. [Google Scholar] [CrossRef] [PubMed]
- Nitta, K.; Ozaki, K.; Ishikawa, M.; Furusawa, S.; Hosono, M.; Kawauchi, H.; Sasaki, K.; Takayanagi, Y.; Tsuiki, S.; Hakomori, S. Inhibition of cell proliferation by Rana catesbeiana and Rana japonica lectins belonging to the ribonuclease superfamily. Cancer Res. 1994, 54, 920–927. [Google Scholar] [PubMed]
- Titani, K.; Takio, K.; Kuwada, M.; Nitta, K.; Sakakibara, F.; Kawauchi, H.; Takayanagi, G.; Hakomori, S. Amino acid sequence of sialic acid binding lectin from frog (Rana catesbeiana) eggs. Biochemistry 1987, 26, 2189–2194. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.-D. A pyrimidine-guanine sequence-specific ribonuclease from Rana catesbeiana (bullfrog) oocytes. Nucleic Acids Res. 1992, 20, 1371–1377. [Google Scholar] [CrossRef] [PubMed]
- Nitta, K.; Ozaki, K.; Tsukamoto, Y.; Hosono, M.; Ogawakonno, Y.; Kawauchi, H.; Takayanagi, Y.; Tsuiki, S.; Hakomori, S. Catalytic lectin (leczyme) from bullfrog (Rana catesbeiana) eggs. Int. J. Oncol. 1996, 9, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Sharon, N.; Lis, H. Lectins; Springer: Dordrecht, The Netherlands, 2007; ISBN 978-1-4020-6605-4. [Google Scholar]
- Neve, R.M.; Chin, K.; Fridlyand, J.; Yeh, J.; Baehner, F.L.; Fevr, T.; Clark, L.; Bayani, N.; Coppe, J.P.; Tong, F.; et al. A collection of breast cancer cell lines for the study of functionally distinct cancer subtypes. Cancer Cell 2006, 10, 515–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curigliano, G.; Burstein, H.J.; Winer, E.P.; Gnant, M.; Dubsky, P.; Loibl, S.; Colleoni, M.; Regan, M.M.; Piccart-Gebhart, M.; Senn, H.-J.; et al. De-escalating and escalating treatments for early-stage breast cancer: The St. Gallen International Expert Consensus Conference on the Primary Therapy of Early Breast Cancer 2017. Ann. Oncol. 2017, 28, 1700–1712. [Google Scholar] [CrossRef] [PubMed]
- Prat, A.; Perou, C.M. Deconstructing the molecular portraits of breast cancer. Mol. Oncol. 2011, 5, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Jitariu, A.-A.; Cîmpean, A.M.; Ribatti, D.; Raica, M. Triple negative breast cancer: The kiss of death. Oncotarget 2017, 8, 46652–46662. [Google Scholar] [CrossRef] [PubMed]
- Schlessinger, J. Cell signaling by receptor tyrosine kinases. Cell 2000, 103, 211–225. [Google Scholar] [CrossRef]
- Park, H.S.; Jang, M.H.; Kim, E.J.; Kim, H.J.; Lee, H.J.; Kim, Y.J.; Kim, J.H.; Kang, E.; Kim, S.-W.; Kim, I.A.; et al. High EGFR gene copy number predicts poor outcome in triple-negative breast cancer. Mod. Pathol. 2014, 27, 1212–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sassen, A.; Rochon, J.; Wild, P.; Hartmann, A.; Hofstaedter, F.; Schwarz, S.; Brockhoff, G. Cytogenetic analysis of HER1/EGFR, HER2, HER3 and HER4 in 278 breast cancer patients. Breast Cancer Res. 2008, 10, R2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, R.; Shah, A.N.; Santa-Maria, C.A.; Cruz, M.R.; Mahalingam, D.; Carneiro, B.A.; Chae, Y.K.; Cristofanilli, M.; Gradishar, W.J.; Giles, F.J. Targeting Epidermal Growth Factor Receptor in triple negative breast cancer: New discoveries and practical insights for drug development. Cancer Treat. Rev. 2017, 53, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Tseng, H.-H.; Yu, Y.-L.; Chen, Y.-L.S.; Chen, J.-H.; Chou, C.-L.; Kuo, T.-Y.; Wang, J.-J.; Lee, M.-C.; Huang, T.-H.; Chen, M.H.-C.; et al. RC-RNase-induced cell death in estrogen receptor positive breast tumors through down-regulation of Bcl-2 and estrogen receptor. Oncol. Rep. 2011, 25, 849–853. [Google Scholar] [CrossRef] [PubMed]
- Kariya, Y.; Tatsuta, T.; Sugawara, S.; Kariya, Y.; Nitta, K.; Hosono, M. RNase activity of sialic acid-binding lectin from bullfrog eggs drives antitumor effect via the activation of p38 MAPK to caspase-3/7 signaling pathway in human breast cancer cells. Int. J. Oncol. 2016, 49, 1334–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Lin, Y.C. Transformation of MCF-10A human breast epithelial cells by zeranol and estradiol-17β. Breast J. 2004, 10, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Subik, K.; Lee, J.F.; Baxter, L.; Strzepek, T.; Costello, D.; Crowley, P.; Xing, L.; Hung, M.C.; Bonfiglio, T.; Hicks, D.G.; et al. The expression patterns of ER, PR, HER2, CK5/6, EGFR, KI-67 and AR by immunohistochemical analysis in breast cancer cell lines. Breast Cancer Basic Clin. Res. 2010, 4, 35–41. [Google Scholar] [CrossRef]
- Jänicke, R.U. MCF-7 breast carcinoma cells do not express caspase-3. Breast Cancer Res. Treat. 2009, 117, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Tatsuta, T.; Sugawara, S.; Takahashi, K.; Ogawa, Y.; Hosono, M.; Nitta, K. Leczyme: A new candidate drug for cancer therapy. Biomed. Res. Int. 2014, 2014, 421415. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Bai, W. Regulation of Estrogen Receptor Nuclear Export by Ligand-Induced and p38-Mediated Receptor Phosphorylation. Mol. Cell. Biol. 2002, 22, 5835–5845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, J.A.; Amazit, L.; Bellance, C.; Guiochon-Mantel, A.; Lombès, M.; Loosfelt, H. p38 and p42/44 MAPKs Differentially Regulate Progesterone Receptor A and B Isoform Stabilization. Mol. Endocrinol. 2011, 25, 1710–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Driggers, P.H.; Segars, J.H. Estrogen action and cytoplasmic signaling pathways. Part II: The role of growth factors and phosphorylation in estrogen signaling. Trends Endocrinol. Metab. 2002, 13, 422–427. [Google Scholar] [CrossRef]
- Bunone, G.; Briand, P.A.; Miksicek, R.J.; Picard, D. Activation of the unliganded estrogen receptor by EGF involves the MAP kinase pathway and direct phosphorylation. EMBO J. 1996, 15, 2174–2183. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Endoh, H.; Masuhiro, Y.; Kitamoto, T.; Uchiyama, S.; Sasaki, H.; Masushige, S.; Gotoh, Y.; Nishida, E.; Kawashima, H.; et al. Activation of the estrogen receptor through phosphorylation by mitogen-activated protein kinase. Science 1995, 270, 1491–1494. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Horwitz, K.B.; Lange, C.A. Transcriptional hyperactivity of human progesterone receptors is coupled to their ligand-dependent down-regulation by mitogen-activated protein kinase-dependent phosphorylation of serine 294. Mol. Cell. Biol. 2001, 21, 6122–6131. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.A.; Shen, T.; Horwitz, K.B. Phosphorylation of human progesterone receptors at serine-294 by mitogen-activated protein kinase signals their degradation by the 26S proteasome. Proc. Natl. Acad. Sci. USA 2000, 97, 1032–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, G.; Hübner, M.R.; Métivier, R.; Brand, H.; Denger, S.; Manu, D.; Beaudouin, J.; Ellenberg, J.; Gannon, F. Cyclic, proteasome-mediated turnover of unliganded and liganded ERalpha on responsive promoters is an integral feature of estrogen signaling. Mol. Cell. 2003, 11, 695–707. [Google Scholar] [CrossRef]
- Laïos, I.; Journé, F.; Nonclercq, D.; Vidal, D.S.; Toillon, R.-A.; Laurent, G.; Leclercq, G. Role of the proteasome in the regulation of estrogen receptor alpha turnover and function in MCF-7 breast carcinoma cells. J. Steroid Biochem. Mol. Biol. 2005, 94, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Calvo, V.; Beato, M. BRCA1 counteracts progesterone action by ubiquitination leading to progesterone receptor degradation and epigenetic silencing of target promoters. Cancer Res. 2011, 71, 3422–3431. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Xiao, Z.; Meng, Z.; Katzenellenbogen, B.S. Phosphorylation by p38 mitogen-activated protein kinase promotes estrogen receptor α turnover and functional activity via the SCF(Skp2) proteasomal complex. Mol. Cell. Biol. 2012, 32, 1928–1943. [Google Scholar] [CrossRef] [PubMed]
- Tatsuta, T.; Hosono, M.; Sugawara, S.; Kariya, Y.; Ogawa, Y.; Hakomori, S.; Nitta, K. Sialic acid-binding lectin (leczyme) induces caspase-dependent apoptosis-mediated mitochondrial perturbation in Jurkat cells. Int. J. Oncol. 2013, 43, 1402–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatsuta, T.; Hosono, M.; Takahashi, K.; Omoto, T.; Kariya, Y.; Sugawara, S.; Hakomori, S.; Nitta, K. Sialic acid-binding lectin (leczyme) induces apoptosis to malignant mesothelioma and exerts synergistic antitumor effects with TRAIL. Int. J. Oncol. 2014, 44, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Dhamad, A.E.; Zhou, Z.; Zhou, J.; Du, Y. Systematic proteomic identification of the heat shock proteins (Hsp) that interact with estrogen receptor alpha (ERα) and biochemical characterization of the ERα-Hsp70 Interaction. PLoS ONE 2016, 11, e0160312. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.; Garcia, Y.A.; Zierer, B.; Patwardhan, C.; Gutierrez, O.; Hildenbrand, Z.; Harris, D.C.; Balsiger, H.A.; Sivils, J.C.; Johnson, J.L.; et al. The cochaperone SGTA (small glutamine-rich tetratricopeptide repeat-containing protein alpha) demonstrates regulatory specificity for the androgen, glucocorticoid, and progesterone receptors *. J. Biol. Chem. 2014, 289, 15297–15306. [Google Scholar] [CrossRef] [PubMed]
- Tatsuta, T.; Hosono, M.; Ogawa, Y.; Inage, K.; Sugawara, S.; Nitta, K. Downregulation of Hsp70 inhibits apoptosis induced by sialic acid-binding lectin (leczyme). Oncol. Rep. 2013, 1, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Sigismund, S.; Argenzio, E.; Tosoni, D.; Cavallaro, E.; Polo, S.; Di Fiore, P.P. Clathrin-Mediated Internalization Is Essential for Sustained EGFR Signaling but Dispensable for Degradation. Dev. Cell. 2008, 15, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Hatton, N.; Lintz, E.; Mahankali, M.; Henkels, K.; Gomez-Cambronero, J. Phosphatidic acid (PA) increases EGF receptor (EGFR) expression by stabilizing mRNA, inhibiting RNAse-A, and by inhibiting lysosomal and proteasomal degradation of the internalized EGFR. Mol. Cell. Biol. 2015, 35, MCB.00286-15. [Google Scholar] [CrossRef] [PubMed]
- Esser, C.; Alberti, S.; Höhfeld, J. Cooperation of molecular chaperones with the ubiquitin/proteasome system. Biochim. Biophys. Acta Mol. Cell. Res. 2004, 1695, 171–188. [Google Scholar] [CrossRef] [PubMed]
- Leu, J.I.-J.; Pimkina, J.; Pandey, P.; Murphy, M.E.; George, D.L. HSP70 inhibition by the small-molecule 2-phenylethynesulfonamide impairs protein clearance pathways in tumor cells. Mol. Cancer Res. 2011, 9, 936–947. [Google Scholar] [CrossRef] [PubMed]
- Sawai, A.; Chandarlapaty, S.; Greulich, H.; Gonen, M.; Ye, Q.; Arteaga, C.L.; Sellers, W.; Rosen, N.; Solit, D.B. Inhibition of Hsp90 down-regulates mutant epidermal growth factor receptor (EGFR) expression and sensitizes EGFR mutant tumors to paclitaxel. Cancer Res. 2008, 68, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.D.; Huang, H.C.; Chan, H.J.; Kuo, S.J. Large-scale preparation of a ribonuclease from Rana catesbeiana (bullfrog) oocytes and characterization of its specific cytotoxic activity against tumor cells. Protein Expr. Purif. 1996, 7, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.C.; Tang, C.H.; Wang, J.J. Caspase activation in response to cytotoxic Rana catesbeiana ribonuclease in MCF-7 cells. FEBS Lett. 2001, 503, 65–68. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.C.; Lee, Y.H.; Tang, C.H.; Cheng, J.T.; Wang, J.J. Synergistic cytotoxicity of Rana catesbeiana ribonuclease and IFN-gamma on hepatoma cells. Biochem. Biophys. Res. Commun. 2001, 280, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.W.; Hu, C.C.A.; Tang, C.H.A.; Lee, M.C.; Wang, J.J. Induction of differentiation rescues HL-60 cells from Rana catesbeiana ribonuclease-induced cell death. FEBS Lett. 2002, 531, 421–426. [Google Scholar] [CrossRef] [Green Version]
- Tang, P.-C.; Huang, H.-C.; Wang, S.-C.; Jeng, J.-C.; Liao, Y.-D. Regulation of ribonuclease expression by estradiol in Rana catesbeiana (Bullfrog). Nucleic Acids Res. 2002, 30, 3286–3293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, C.-H.A.; Hu, C.-C.A.; Wei, C.-W.; Wang, J.-J. Synergism of Rana catesbeiana ribonuclease and IFN-gamma triggers distinct death machineries in different human cancer cells. FEBS Lett. 2005, 579, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yiang, G.; Lin, Y.; Chou, P.; Wu, T.; Chang, W.; Chen, C.; Yu, Y. Rana catesbeiana ribonuclease induces cell apoptosis via the caspase-9/-3 signaling pathway in human glioblastoma DBTRG, GBM8901 and GBM8401 cell lines. Oncol. Lett. 2015, 2471–2476. [Google Scholar] [CrossRef] [PubMed]
- Satoh, T.; Tatsuta, T.; Sugawara, S.; Hara, A.; Hosono, M. Synergistic anti-tumor effect of bullfrog sialic acid-binding lectin and pemetrexed in malignant mesothelioma. Oncotarget 2017, 8, 42466–42477. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-H.; Wei, C.-W.; Wang, J.-J.; Chiou, C.-T. Rana catesbeiana ribonuclease inhibits Japanese encephalitis virus (JEV) replication and enhances apoptosis of JEV-infected BHK-21 cells. Antiviral Res. 2011, 89, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, Y.; Sugawara, S.; Tatsuta, T.; Hosono, M.; Nitta, K.; Fujii, Y.; Kobayashi, H.; Fujimura, T.; Taka, H.; Koide, Y.; et al. Sialyl-glycoconjugates in cholesterol-rich microdomains of P388 cells are the triggers for apoptosis induced by Rana catesbeiana oocyte ribonuclease. Glycoconj. J. 2014, 31, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Tatsuta, T.; Satoh, T.; Sugawara, S.; Hara, A.; Hosono, M. Sialic acid-binding lectin from bullfrog eggs inhibits human malignant mesothelioma cell growth in vitro and in vivo. PLoS ONE 2018, 13, e0190653. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F.; Zhang, J.; Liao, Y.-D.; Dyer, K.D. Rapid diversification of RNase A superfamily ribonucleases from the bullfrog, Rana catesbeiana. J. Mol. Evol. 2001, 53, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Notomista, E.; Catanzano, F.; Graziano, G.; Piaz, F.D.; Barone, G.; D’Alessio, G.; Di Donato, A. Onconase: An unusually stable protein. Biochemistry 2000, 39, 8711–8718. [Google Scholar] [CrossRef] [PubMed]
- Costanzi, J.; Sidransky, D.; Navon, A.; Goldsweig, H. Ribonucleases as a novel pro-apoptotic anticancer strategy: Review of the preclinical and clinical data for ranpirnase. Cancer Invest. 2005, 23, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Tatsuta, T.; Sugawara, S.; Takahashi, K.; Ogawa, Y.; Hosono, M.; Nitta, K. Cancer-selective induction of apoptosis by leczyme. Front. Oncol. 2014, 4, 139. [Google Scholar] [CrossRef] [PubMed]
- Serra, V.; Scaltriti, M.; Prudkin, L.; Eichhorn, P.J.A.; Ibrahim, Y.H.; Chandarlapaty, S.; Markman, B.; Rodriguez, O.; Guzman, M.; Rodriguez, S.; et al. PI3K inhibition results in enhanced HER signaling and acquired ERK dependency in HER2-overexpressing breast cancer. Oncogene 2011, 30, 2547–2557. [Google Scholar] [CrossRef] [PubMed]
- Rodrik-Outmezguine, V.S.; Chandarlapaty, S.; Pagano, N.C.; Poulikakos, P.I.; Scaltriti, M.; Moskatel, E.; Baselga, J.; Guichard, S.; Rosen, N. mTOR kinase inhibition causes feedback-dependent biphasic regulation of AKT signaling. Cancer Discov. 2011, 1, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.J.; Castel, P.; Radosevic-robin, N.; Elkabets, M.; Aceto, N.; Weitsman, G.; Barber, P.; Vojnovic, B.; Morse, N.; Viola-villegas, N.T.; et al. Antagonism of EGFR and HER3 Enhances the Response to Inhibitors of the PI3K-Akt Pathway in Triple-Negative Breast Cancer. Sci. Signal. 2015, 7, ra29. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of cSBL are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | ER 1 | PgR 1 | HER2 2 | Subtype |
|---|---|---|---|---|
| ZR-75-1 | + | + | 2+ | Luminal A |
| BT-474 | + | + | 3+ | Luminal B |
| MCF7 | + | + | 0-1+ | Luminal A |
| SK-BR-3 | - | - | 3+ | HER2 |
| MDA-MB-231 | - | - | 0-1+ | Basal |
| MDA-MB-468 | - | - | - | Basal |
| MCF 10A | - | - | 0-1+ | Basal |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tatsuta, T.; Sato, S.; Sato, T.; Sugawara, S.; Suzuki, T.; Hara, A.; Hosono, M. Sialic Acid-Binding Lectin from Bullfrog Eggs Exhibits an Anti-Tumor Effect Against Breast Cancer Cells Including Triple-Negative Phenotype Cells. Molecules 2018, 23, 2714. https://doi.org/10.3390/molecules23102714
Tatsuta T, Sato S, Sato T, Sugawara S, Suzuki T, Hara A, Hosono M. Sialic Acid-Binding Lectin from Bullfrog Eggs Exhibits an Anti-Tumor Effect Against Breast Cancer Cells Including Triple-Negative Phenotype Cells. Molecules. 2018; 23(10):2714. https://doi.org/10.3390/molecules23102714
Chicago/Turabian StyleTatsuta, Takeo, Shoko Sato, Toshiyuki Sato, Shigeki Sugawara, Tsuneyoshi Suzuki, Akiyoshi Hara, and Masahiro Hosono. 2018. "Sialic Acid-Binding Lectin from Bullfrog Eggs Exhibits an Anti-Tumor Effect Against Breast Cancer Cells Including Triple-Negative Phenotype Cells" Molecules 23, no. 10: 2714. https://doi.org/10.3390/molecules23102714