Astragalus Polysaccharide Improves Insulin Sensitivity via AMPK Activation in 3T3-L1 Adipocytes

Abstract
:1. Introduction
2. Results
2.1. APS Promoted 3T3-L1 Pre-Adipocytes Proliferation
2.2. Effect of APS on Adipogenesis in 3T3-L1 Pre-Adipocytes
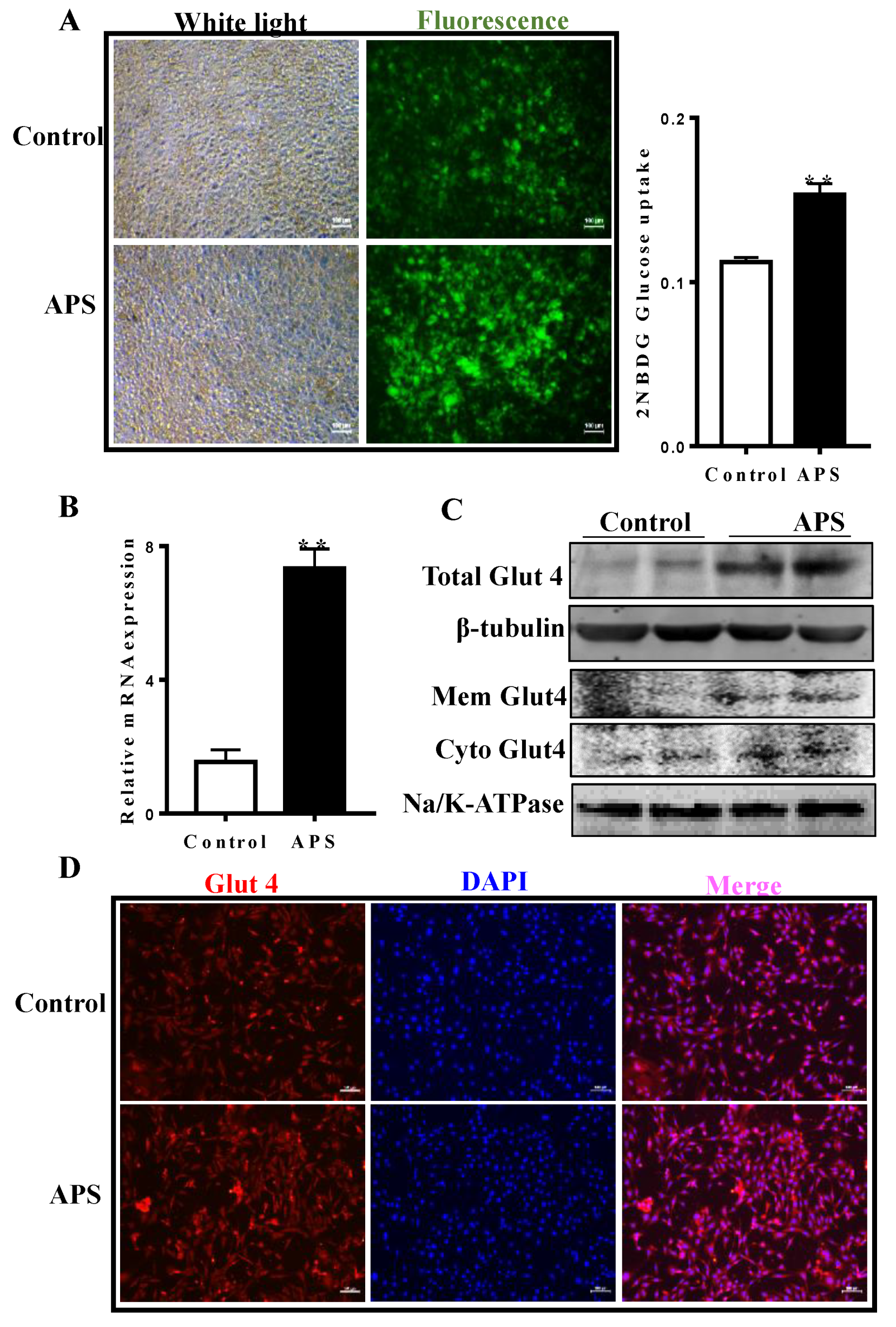
2.3. APS Stimulated Glucose Uptake and Increased the Expression of Glut4
2.4. Effect of APS Treatment on Insulin Sensitivity and AMPK Activity
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Lactate Dehydrogenase (LDH) Assay
4.3. CCK-8 Assays
4.4. EdU Staining
4.5. Oil-Red-O Staining
4.6. Western Blotting
4.7. Real-Time Quantitative PCR (qRT-PCR)
4.8. Plasma Membrane and Cytosol Fractionation
4.9. Glucose Uptake Assay
4.10. Immunocytochemical Staining
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hameed, I.; Masoodi, S.R.; Mir, S.A.; Nabi, M.; Ghazanfar, K.; Ganai, B.A. Type 2 diabetes mellitus: From a metabolic disorder to an inflammatory condition. World J. Diabetes 2015, 6, 598–612. [Google Scholar] [CrossRef] [PubMed]
- Kahn, S.E.; Cooper, M.E.; Del Prato, S. Pathophysiology and treatment of type 2 diabetes: Perspectives on the past, present, and future. Lancet 2014, 383, 1068–1083. [Google Scholar] [CrossRef]
- Reaven, G.M. Insulin resistance: The link between obesity and cardiovascular disease. Endocrinol. Metab. Clin. North Am. 2008, 37, 581–601. [Google Scholar] [CrossRef] [PubMed]
- Kahn, B.B.; Alquier, T.; Carling, D.; Hardie, D.G. AMP-activated protein kinase: Ancient energy gauge provides clues to modern understanding of metabolism. Cell Metab. 2005, 1, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Tom, R.Z.; Garcia-Roves, P.M.; Sjögren, R.J.; Jiang, L.Q.; Holmström, M.H.; Deshmukh, A.S.; Vieira, E.; Chibalin, A.V.; Björnholm, M.; Zierath, J.R. Effects of AMPK activation on insulin sensitivity and metabolism in leptin-deficient ob/ob mice. Diabetes 2014, 63, 1560–1571. [Google Scholar] [CrossRef]
- Viollet, B.; Andreelli, F. AMP-activated protein kinase and metabolic control. In Diabetes—Perspectives in Drug Therapy; Handbook of Experimental Pharmacology; Schwanstecher, M., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; Volume 203, pp. 303–330. [Google Scholar]
- Leverve, X.; Guigas, B.; Detaille, D.; Batandier, C.; Koceir, E.; Chauvin, C.; Fontaine, E.; Wiernsperger, N. Mitochondrial metabolism and type-2 diabetes: A specific target of metformin. Diabetes Metab. 2003, 29, 6S88–6S94. [Google Scholar] [CrossRef]
- Thong, F.S.; Bilan, P.J.; Klip, A. The Rab GTPase-activating protein AS160 integrates Akt, protein kinase C, and AMP-activated protein kinase signals regulating GLUT4 traffic. Diabetes 2007, 56, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Calabrese, E.J.; Lian, B.; Lin, Z.; Calabrese, V. Hormesis as a mechanistic approach to understanding herbal treatments in traditional Chinese medicine. Pharmacol. Ther. 2017, 184, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.; Dai, Y.; Verpoorte, R.; Lam, W.; Cheng, Y.-C.; Pao, L.-H.; Zhang, W.; Chen, S. Major achievements of evidence-based traditional Chinese medicine in treating major diseases. Biochem. Pharmacol. 2017, 139, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Wang, Z.; Huang, L.; Zheng, S.; Wang, D.; Chen, S.; Zhang, H.; Yang, S. Review of the botanical characteristics, phytochemistry, and pharmacology of Astragalus membranaceus (Huangqi). Phytother. Res. 2014, 28, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Zhang, Z.-F.; Ding, Y.; Wang, J.-B.; Li, Y. Astragalus polysaccharide improves palmitate-induced insulin resistance by inhibiting PTP1B and NF-κB in C2C12 myotubes. Molecules 2012, 17, 7083–7092. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Weng, S.; Wang, L.; Mao, Z. Mechanism of Astragalus polysaccharides in attenuating insulin resistance in Rats with type 2 diabetes mellitus via the regulation of liver microRNA-203a-3p. Mol. Med. Rep. 2018, 17, 1617–1624. [Google Scholar] [CrossRef] [PubMed]
- Abel, E.D.; Peroni, O.; Kim, J.K.; Kim, Y.B.; Boss, O.; Hadro, E.; Minnemann, T.; Shulman, G.I.; Kahn, B.B. Adipose-selective targeting of the GLUT4 gene impairs insulin action in muscle and liver. Nature 2001, 409, 729–733. [Google Scholar] [CrossRef] [PubMed]
- Leonardini, A.; Laviola, L.; Perrini, S.; Natalicchio, A.; Giorgino, F. Cross-talk between PPAR and insulin signaling and modulation of insulin sensitivity. PPAR Res. 2009, 2009, 818945. [Google Scholar] [CrossRef]
- Patel, D.K.; Prasad, S.K.; Kumar, R.; Hemalatha, S. An overview on antidiabetic medicinal plants having insulin mimetic property. Asian Pac. J. Trop. Biomed. 2012, 2, 320–330. [Google Scholar] [CrossRef] [Green Version]
- Hiebl, B.; Peters, S.; Gemeinhardt, O.; Niehues, S.M.; Jung, F. Impact of serum in cell culture media on in vitro lactate dehydrogenase (LDH) release determination. J. Cell. Biochem. 2017, 3, 9–13. [Google Scholar] [CrossRef]
- Juríková, M.; Danihel, Ľ.; Polák, Š.; Varga, I. Ki67, PCNA, and MCM proteins: Markers of proliferation in the diagnosis of breast cancer. Acta Histochem. 2016, 118, 544–552. [Google Scholar] [CrossRef] [PubMed]
- Maga, G.; Hubscher, U. Proliferating cell nuclear antigen (PCNA): A dancer with many partners. J. Cell. Sci. 2003, 116, 3051–3060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Liu, H.T. MAPK signal pathways in the regulation of cell proliferation in mammalian cells. Cell Res. 2002, 12, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Q.Q.; Lane, M.D. Adipogenesis: From Stem Cell to Adipocyte. Annu. Rev. Biochem. 2012, 81, 715–736. [Google Scholar] [CrossRef] [PubMed]
- Siersbaek, R.; Nielsen, R.; Mandrup, S. Transcriptional networks and chromatin remodeling controlling adipogenesis. Trends Endocrinol. Metab. 2012, 23, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Ke, B.; Ke, X.; Wan, X.; Yang, Y.; Huang, Y.; Qin, J.; Hu, C.; Shi, L. Astragalus polysaccharides attenuates TNF-alpha-induced insulin resistance via suppression of miR-721 and activation of PPAR-gamma and PI3K/AKT in 3T3-L1 adipocytes. Am. J. Transl. Res. 2017, 9, 2195–2206. [Google Scholar] [PubMed]
- Shan, T.; Liu, W.; Kuang, S. Fatty acid binding protein 4 expression marks a population of adipocyte progenitors in white and brown adipose tissues. FASEB J. 2013, 27, 277–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Mariman, E.; Renes, J.; Keijer, J. The secretory function of adipocytes in the physiology of white adipose tissue. J. Cell. Physiol. 2008, 216, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Satoh, T. Molecular mechanisms for the regulation of insulin-stimulated glucose uptake by small guanosine triphosphatases in skeletal muscle and adipocytes. Int. J. Mol. Sci. 2014, 15, 18677–18692. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, I.; Dörpholz, G.; Ott, C.-E.; Kragesteen, B.; Schanze, N.; Lee, C.T.; Köhrle, J.; Mundlos, S.; Ruschke, K.; Knaus, P. BMPs as new insulin sensitizers: Enhanced glucose uptake in mature 3T3-L1 adipocytes via PPARγ and GLUT4 upregulation. Sci. Rep. 2017, 7, 17192. [Google Scholar] [CrossRef] [PubMed]
- Zou, F.; Mao, X.-Q.; Wang, N.; Liu, J.; Ou-Yang, J.-P. Astragalus polysaccharides alleviates glucose toxicity and restores glucose homeostasis in diabetic states via activation of AMPK. Acta Pharmacol. Sin. 2009, 30, 1607–1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leto, D.; Saltiel, A.R. Regulation of glucose transport by insulin: Traffic control of GLUT4. Nat. Rev. Mol. Cell Bio. 2012, 13, 383. [Google Scholar] [CrossRef] [PubMed]
- Watson, R.T.; Pessin, J.E. Bridging the GAP between insulin signaling and GLUT4 translocation. Trends Biochem. Sci. 2006, 31, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Mao, X.; Wang, L.; Liu, M.; Wetzel, M.D.; Guan, K.-L.; Dong, L.Q.; Liu, F. Adiponectin sensitizes insulin signaling by reducing p70 S6 kinase-mediated serine phosphorylation of IRS-1. J. Biol. Chem. 2007, 282, 7991–7996. [Google Scholar] [CrossRef] [PubMed]
- Russo, G.L.; Russo, M.; Ungaro, P. AMP-activated protein kinase: A target for old drugs against diabetes and cancer. Biochem. Pharmacol. 2013, 86, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Zou, F.; Liu, J.; Jin, H.; Ouyang, J. APS Actives AMPK and alleviates glucose toxicity in high-glucose treated C2C12 cells. FASEB J. 2009, 23, 753–756. [Google Scholar] [CrossRef]
- McGee, S.L.; Van Denderen, B.J.; Howlett, K.F.; Mollica, J.; Schertzer, J.D.; Kemp, B.E.; Hargreaves, M. AMP-activated protein kinase regulates GLUT4 transcription by phosphorylating histone deacetylase 5. Diabetes 2008, 57, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Katahira, H.; Ozawa, S.; Nakamichi, Y.; Tanaka, T.; Shimoyama, T.; Takahashi, K.; Yoshimoto, K.; Imaizumi, M.O.; Nagamatsu, S. Activators of AMP-activated protein kinase enhance GLUT4 translocation and its glucose transport activity in 3T3-L1 adipocytes. AJP: Endocrinol. Metab. 2005, 289, E643–E649. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.X.; Hu, J.; Zhu, M.J.; Du, M. Trenbolone enhances myogenic differentiation via enhancing β-catenin signaling in muscle-derived stem cells of cattle. Domest. Anim. Endocrinol. 2011, 40, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kang, S.M.; Ko, S.C.; Lee, D.H.; Jeon, Y.J. Octaphlorethol A, a novel phenolic compound isolated from a brown alga, Ishige foliacea, increases glucose transporter 4-mediated glucose uptake in skeletal muscle cells. Biochem. Biophys. Res. Commun. 2012, 420, 576–581. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′–3′) | Accession Number |
|---|---|---|
| PCNA | GAACCTCACCAGCATGTCCA TGGGATTCCAAGTTGCTCCA | NM_011045.2 |
| PPARγ | ACCTCTGCTGGGGATCTGAA ATCACGGAGAGGTCCACAGA | NM_001127330.2 |
| C/EBPα | CCCTTGCTTTTTGCACCTCC TGCCCCCATTCTCCATGAAC | NM_001287514.1 |
| AP2 | GGATTTGGTCACCATCCGGT TTCCATCCCACTTCTGCACC | NM_024406.3 |
| Glut4 | CTAGGCATCAATGCTGTTTTCTA CGAGACCAACGTGAAGACCGTATT | AB008453.1 |
| 18S | GTAACCCGTTGAACCCCATT CCATCCAATCGGTAGTAGCG | M35283.1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, R.; Qin, X.; Zhang, T.; Li, Q.; Zhang, J.; Zhao, J. Astragalus Polysaccharide Improves Insulin Sensitivity via AMPK Activation in 3T3-L1 Adipocytes. Molecules 2018, 23, 2711. https://doi.org/10.3390/molecules23102711
Zhang R, Qin X, Zhang T, Li Q, Zhang J, Zhao J. Astragalus Polysaccharide Improves Insulin Sensitivity via AMPK Activation in 3T3-L1 Adipocytes. Molecules. 2018; 23(10):2711. https://doi.org/10.3390/molecules23102711
Chicago/Turabian StyleZhang, Ruixin, Xuze Qin, Ting Zhang, Qian Li, Jianxin Zhang, and Junxing Zhao. 2018. "Astragalus Polysaccharide Improves Insulin Sensitivity via AMPK Activation in 3T3-L1 Adipocytes" Molecules 23, no. 10: 2711. https://doi.org/10.3390/molecules23102711