
Anti-Inflammatory and Antioxidant Properties of Casein Hydrolysate Produced Using High Hydrostatic Pressure Combined with Proteolytic Enzymes

Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of Different Enzymes and High Hydrostatic Pressure on Casein Hydrolysis
2.2. Effect of Incubation Time on Casein Hydrolysis
2.3. Effect of Enzyme-to-Substrate (E:S) Ratio on Casein Hydrolysis
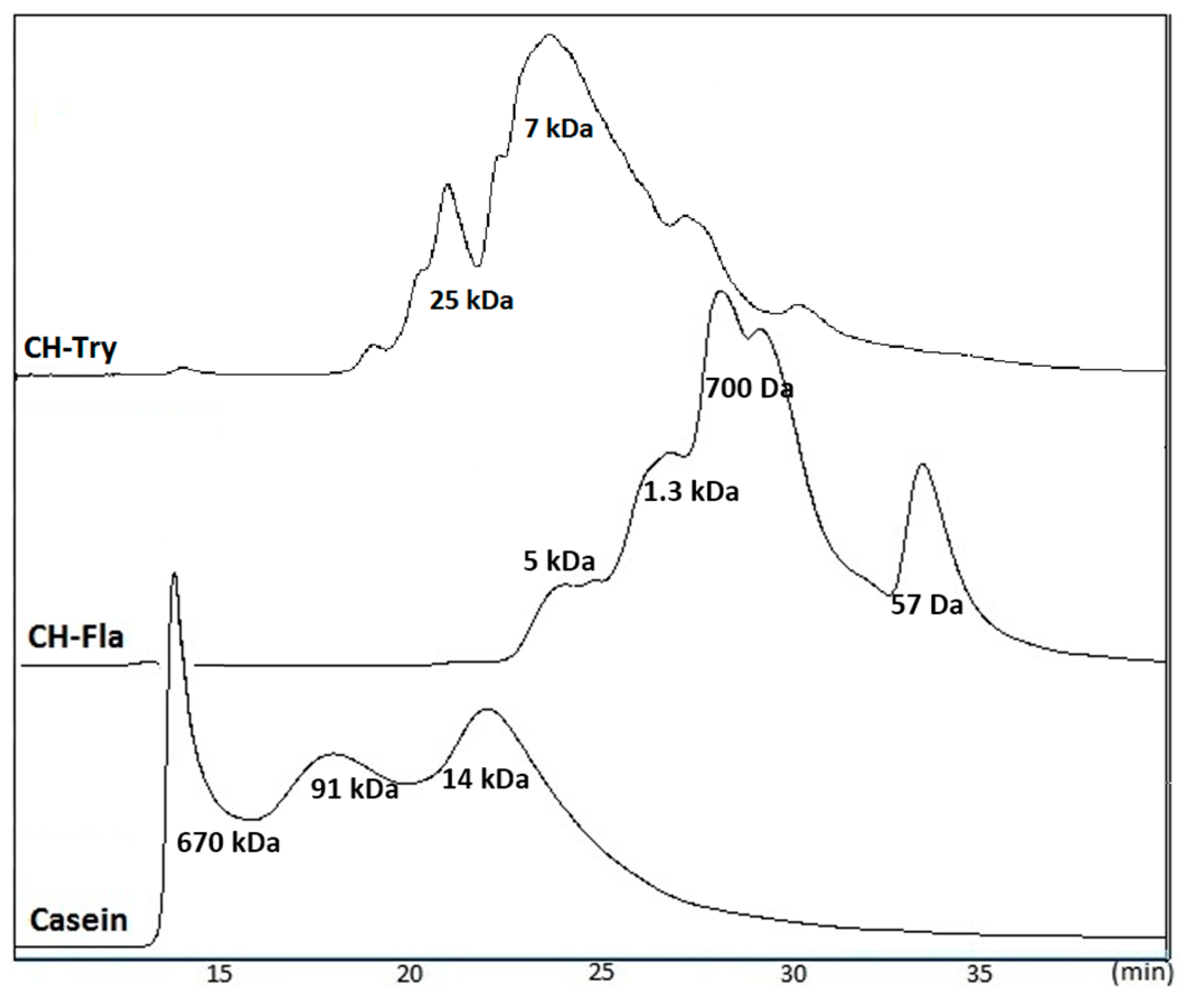
2.4. Molecular Weight Distribution of CH
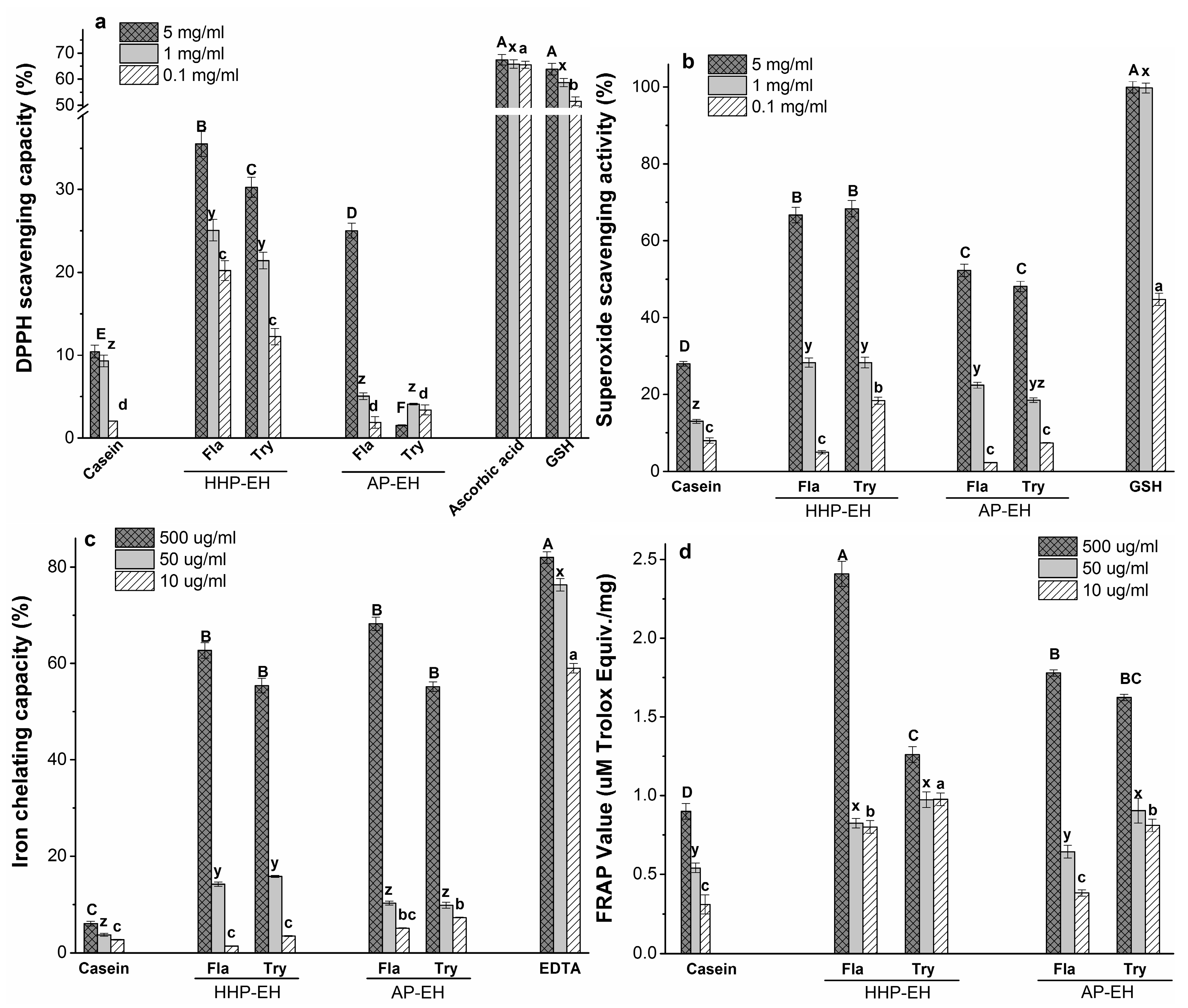
2.5. Antioxidant Capacity of CH
2.5.1. 1,1-Diphenyl-2-picryl Hydrazyl (DPPH) Radical Scavenging Capacity
2.5.2. Superoxide Radical Scavenging Activity
2.5.3. Iron Chelating Activity
2.5.4. Reducing Capacity (FRAP assay)
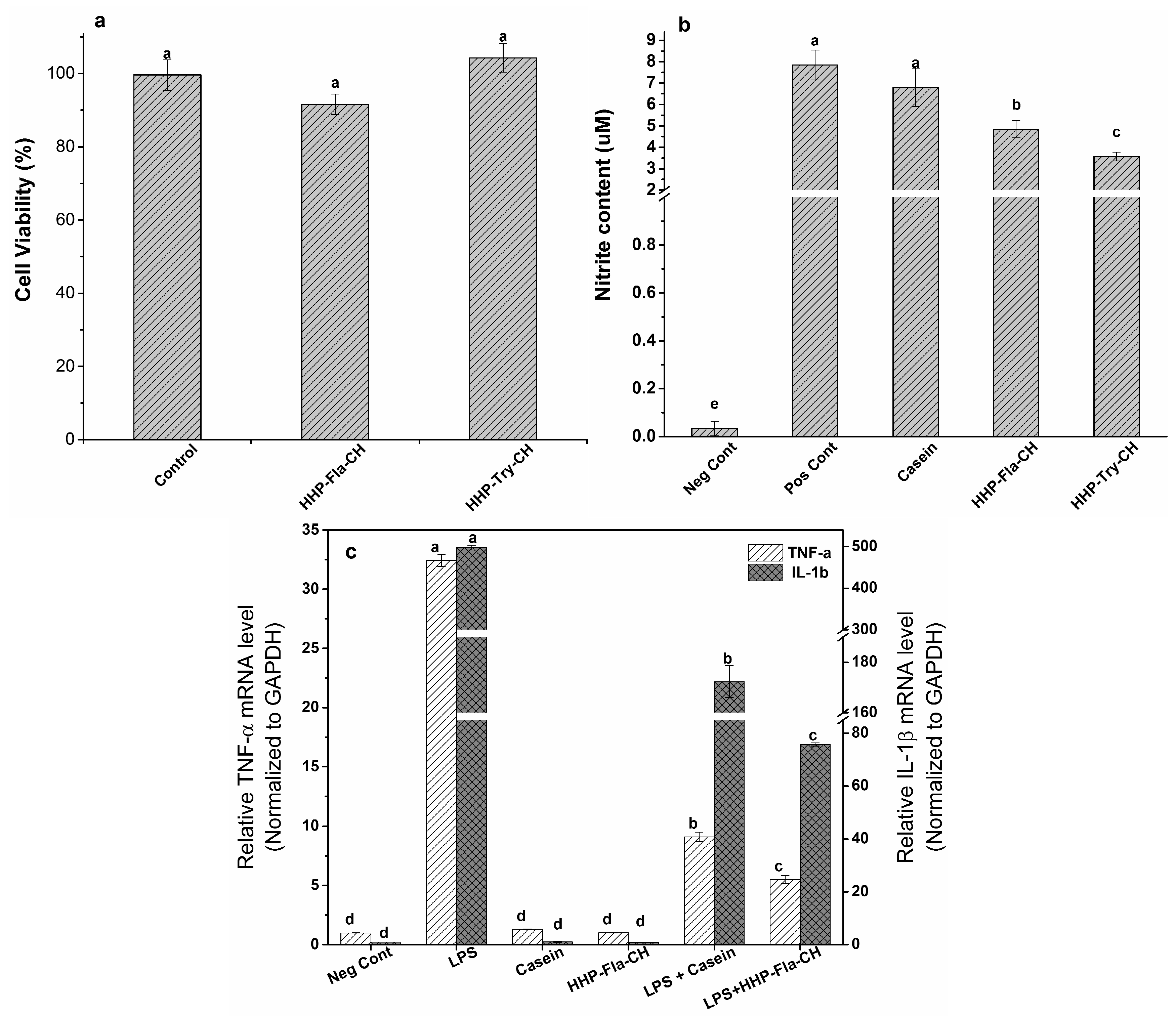
2.6. Anti-Inflammatory Properties of CH
2.6.1. Cell Viability Assay
2.6.2. Determination of NO Production by Macrophage Cells
2.6.3. Gene Expression of Pro-Inflammatory Cytokines in LPS-Stimulated Macrophages
2.7. Peptide Sequencing of the Most Potent CH
3. Materials and Methods
3.1. Materials
3.2. Apparatus
3.3. Enzymatic Hydrolysis of Casein
3.4. Determination of Degree of Hydrolysis (DH)
3.5. Size Exclusion High Performance Liquid Chromatography (SE-HPLC)
3.6. Matrix-Assisted Laser Desorption/Ionization–Time of Flight (MALDI–TOF) Spectrometry
3.7. Antioxidant Capacity Analysis
3.7.1. 1,1-Diphenyl-2-picryl Hydrazyl (DPPH) Radical Scavenging Assay
3.7.2. Superoxide Radical Scavenging Assay
3.7.3. Iron Chelating Capacity
3.7.4. Ferric Reducing Antioxidant Power (FRAP) Assay
3.8. Anti-Inflammatory Properties of CH
3.8.1. Cell Viability Assay
3.8.2. Determination of Nitric Oxide (NO) Production (Griess Assay)
3.8.3. Real-Time Polymerase Chain Reaction (RT-PCR) Analysis for Cytokine Gene Expression
3.9. Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kitts, D.D.; Weiler, K. Bioactive proteins and peptides from food sources. Applications of bioprocesses used in isolation and recovery. Curr. Pharm. Des. 2003, 9, 1309–1323. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Zhong, Y. Bioactive peptides. J. AOAC Int. 2008, 91, 914–931. [Google Scholar] [PubMed]
- Halliwell, B. Free radicals and antioxidants: Updating a personal view. Nutr. Rev. 2012, 70, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Jahandideh, F.; Wu, J.P. Food-derived bioactive peptides on inflammation and oxidative stress. BioMed Res. Int. 2014, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Clare, D.A.; Swaisgood, H.E. Bioactive milk peptides: A prospectus. J. Dairy Sci. 2000, 83, 1187–1195. [Google Scholar] [CrossRef]
- Farrell, H.M.; Jimenez-Flores, R.; Bleck, G.T.; Brown, E.M.; Butler, J.E.; Creamer, L.K.; Hicks, C.L.; Hollar, C.M.; Ng-Kwai-Hang, K.F.; Swaisgood, H.E. Nomenclature of the proteins of cows’ milk—Sixth revision. J. Dairy Sci. 2004, 87, 1641–1674. [Google Scholar] [CrossRef]
- Kitts, D.D. Antioxidant properties of casein-phosphopeptides. Trends Food Sci. Technol. 2005, 16, 549–554. [Google Scholar] [CrossRef]
- Garriga, M.; Grebol, N.; Aymerich, M.T.; Monfort, J.M.; Hugas, M. Microbial inactivation after high-pressure processing at 600 Mpa in commercial meat products over its shelf life. Innov. Food Sci. Emerg. Technol. 2004, 5, 451–457. [Google Scholar] [CrossRef]
- Barba, F.; Esteva, M.J.; Frigola, A. High pressure treatment effect of physicochemical and nutritional properties of fluid foods during storage. Compr. Rev. Food Sci. Food Saf. 2012, 11, 307–322. [Google Scholar] [CrossRef]
- Sunwoo, H.H.; Gujral, N.; Huebl, A.C.; Kim, C.T. Application of high hydrostatic pressure and enzymatic hydrolysis for the extraction of ginsenosides from fresh ginseng root (Panax ginseng CA Myer). Food Bioprocess Technol. 2014, 7, 1246–1254. [Google Scholar] [CrossRef]
- Barba, F.; Terefe, N.S.; Buckow, R.; Knorr, D.; Orlien, V. New opportunities and perspectives of high pressure treatment to improve health and safety attributes of foods. Food Res. Int. 2015, 77, 725–742. [Google Scholar] [CrossRef]
- Schettino, V.; Bini, R. Constraining molecules at the closest approach: Chemistry at high pressure. Chem. Soc. Rev. 2007, 36, 869–880. [Google Scholar] [CrossRef] [PubMed]
- Rivalain, N.; Roquain, J.; Demazeau, G. Development of high hydrostatic pressure in biosciences: Pressure effect on biological structures and potential applications in biotechnologies. Biotechnol. Adv. 2010, 28, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Hendrickx, M.; Denys, S.; Indrawati; Ludikhuyze, L.; Van den Broeck, I.; Weemaes, C. Effect of combined high pressure-elevated temperatures on food quality affecting enzymes. Vtt Symp. 1998, 186, 21–25. [Google Scholar]
- Weingand-Ziade, A.; Ribes, F.; Renault, F.; Masson, P. Pressure- and heat-induced inactivation of butyrylcholinesterase: Evidence for multiple intermediates and the remnant inactivation process. Biochem. J. 2001, 356, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Ludikhuyze, L.; van Loey, A.; Indrawati; Smout, C.; Hendrickx, M. Effects of combined pressure and temperature on enzymes related to quality of fruits and vegetables: From kinetic information to process engineering aspects. Crit. Rev. Food Sci. Nutr. 2003, 43, 527–586. [Google Scholar] [CrossRef] [PubMed]
- Van Willige, R.W.G.; Fitzgerald, R.J. Tryptic and chymotryptic hydrolysis of beta-lactoglobulin A, B and AB at ambient and high-pressure. Milchwissenschaft 1995, 50, 183–186. [Google Scholar]
- Quiros, A.; Chichon, R.; Recio, I.; Lopez-Fandino, R. The use of high hydrostatic pressure to promote the proteolysis and release of bioactive peptides from ovalbumin. Food Chem. 2007, 104, 1734–1739. [Google Scholar] [CrossRef]
- Zhang, T.; Jiang, B.; Miao, M.; Mu, W.M.; Li, Y.H. Combined effects of high-pressure and enzymatic treatments on the hydrolysis of chickpea protein isolates and antioxidant activity of the hydrolysates. Food Chem. 2012, 135, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mora, P.; Penas, E.; Frias, J.; Zielinski, H.; Wiczkowski, W.; Zielinska, D.; Martinez-Villaluenga, C. High-pressure-assisted enzymatic release of peptides and phenolics increases angiotensin converting enzyme I inhibitory and antioxidant activities of pinto bean hydrolysates. J. Agric. Food Chem. 2016, 64, 1730–1740. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.; Bamdad, F.; Gujral, N.; Suh, J.W.; Sunwoo, H. High hydrostatic pressure-assisted enzymatic treatment improves antioxidant and anti-inflammatory properties of phosvitin. Curr. Pharm. Biotechnol. 2017, 18, 158–167. [Google Scholar] [CrossRef]
- Lopez-Fandino, R. High pressure-induced changes in milk proteins and possible applications in dairy technology. Int. Dairy J. 2006, 16, 1119–1131. [Google Scholar] [CrossRef]
- Mozhaev, V.V.; Heremans, K.; Frank, J.; Masson, P.; Balny, C. High pressure effects on protein structure and function. Proteins 1996, 24, 81–91. [Google Scholar] [CrossRef]
- Zeece, M.; Huppertz, T.; Kelly, A. Effect of high-pressure treatment on in vitro digestibility of beta-lactoglobulin. Innov. Food Sci. Emerg. Technol. 2008, 9, 62–69. [Google Scholar] [CrossRef]
- Garcia-Mora, P.; Penas, E.; Frias, J.; Gomez, R.; Martinez-Villaluenga, C. High-pressure improves enzymatic proteolysis and the release of peptides with angiotensin I converting enzyme inhibitory and antioxidant activities from lentil proteins. Food Chem. 2015, 171, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Hendrickx, M.; Ludikhuyze, L.; van den Broeck, I.; Weemaes, C. Effects of high pressure on enzymes related to food quality. Trends Food Sci. Technol. 1998, 9, 197–203. [Google Scholar] [CrossRef]
- Rastogi, N.K.; Raghavarao, K.S.M.S.; Balasubramaniam, V.M.; Niranjan, K.; Knorr, D. Opportunities and challenges in high pressure processing of foods. Crit. Rev. Food Sci. 2007, 47, 69–112. [Google Scholar] [CrossRef] [PubMed]
- Dalgleish, D.G.; Corredig, M. The structure of the casein micelle of milk and its changes during processing. Annu. Rev. Food Sci. T 2012, 3, 449–467. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.S.; Su, Y.J.; Jia, F.; Jin, H.L. Characterization of casein hydrolysates derived from enzymatic hydrolysis. Chem. Cent. J. 2013, 7, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Morato, A.F.; Carreira, R.L.; Junqueira, R.G.; Silvestre, M.P.C. Optimization of casein hydrolysis for obtaining high contents of small peptides: Use of subtilisin and trypsin. J. Food Compos. Anal. 2000, 13, 843–857. [Google Scholar] [CrossRef]
- Rossini, K.; Norena, C.P.Z.; Cladera-Olivera, F.; Brandelli, A. Casein peptides with inhibitory activity on lipid oxidation in beef homogenates and mechanically deboned poultry meat. LWT Food Sci. Technol. 2009, 42, 862–867. [Google Scholar] [CrossRef]
- Zhang, J.H.; Zhang, H.; Wang, L.; Guo, X.N.; Wang, X.G.; Yao, H.Y. Antioxidant activities of the rice endosperm protein hydrolysate: Identification of the active peptide. Eur. Food Res. Technol. 2009, 229, 709–719. [Google Scholar] [CrossRef]
- Bamdad, F.; Chen, L.Y. Antioxidant capacities of fractionated barley hordein hydrolysates in relation to peptide structures. Mol. Nutr. Food Res. 2013, 57, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Rajapakse, N.; Mendis, E.; Byun, H.G.; Kim, S.K. Purification and in vitro antioxidative effects of giant squid muscle peptides on free radical-mediated oxidative systems. J. Nutr. Biochem. 2005, 16, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Phelan, M.; Aherne, A.; FitzGerald, R.J.; O’Brien, N.M. Casein-derived bioactive peptides: Biological effects, industrial uses, safety aspects and regulatory status. Int. Dairy J. 2009, 19, 643–654. [Google Scholar] [CrossRef]
- Doorten, A.Y.P.S.; vd Wiel, J.A.G.; Jonker, D. Safety evaluation of an IPP tripeptide-containing milk protein hydrolysate. Food Chem. Toxicol. 2009, 47, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Jin, D.H.; Ogawa, T.; Muramoto, K.; Hatakeyama, E.; Yasuhara, T.; Nokihara, K. Antioxidative properties of tripeptide libraries prepared by the combinatorial chemistry. J. Agric. Food Chem. 2003, 51, 3668–3674. [Google Scholar] [CrossRef] [PubMed]
- Suetsuna, K.; Ukeda, H.; Ochi, H. Isolation and characterization of free radical scavenging activities peptides derived from casein. J. Nutr. Biochem. 2000, 11, 128–131. [Google Scholar] [CrossRef]
- Bamdad, F.; Wu, J.P.; Chen, L.Y. Effects of enzymatic hydrolysis on molecular structure and antioxidant activity of barley hordein. J. Cereal Sci. 2011, 54, 20–28. [Google Scholar] [CrossRef]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Smolskaite, L.; Venskutonis, P.R.; Talou, T. Comprehensive evaluation of antioxidant and antimicrobial properties of different mushroom species. LWT Food Sci. Technol. 2015, 60, 462–471. [Google Scholar] [CrossRef]
- Lin, S.Y.; Jin, Y.; Liu, M.Y.; Yang, Y.; Zhang, M.S.; Guo, Y.; Jones, G.; Liu, J.B.; Yin, Y.G. Research on the preparation of antioxidant peptides derived from egg white with assisting of high-intensity pulsed electric field. Food Chem. 2013, 139, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Iskandar, M.M.; Dauletbaev, N.; Kubow, S.; Mawji, N.; Lands, L.C. Whey protein hydrolysates decrease IL-8 secretion in lipopolysaccharide (LPS)-stimulated respiratory epithelial cells by affecting LPS binding to toll-like receptor 4. Br. J. Nutr. 2013, 110, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Elmarakby, A.A.; Sullivan, J.C. Relationship between oxidative stress and inflammatory cytokines in diabetic nephropathy. Cardiovasc. Ther. 2012, 30, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.N.; Al-Omran, A.; Parvathy, S.S. Role of nitric oxide in inflammatory diseases. Inflammopharmacology 2007, 15, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Villaluenga, C.; Dia, V.P.; Berhow, M.; Bringe, N.A.; de Mejia, E.G. Protein hydrolysates from beta-conglycinin enriched soybean genotypes inhibit lipid accumulation and inflammation in vitro. Mol. Nutr. Food Res. 2009, 53, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Elass-Rochard, E.; Legrand, D.; Salmon, V.; Roseanu, A.; Trif, M.; Tobias, P.S.; Mazurier, J.; Spik, G. Lactoferrin inhibits the endotoxin interaction with CD14 by competition with the lipopolysaccharide-binding protein. Infect. Immun. 1998, 66, 486–491. [Google Scholar] [PubMed]
- Maiolo, J.R.; Ferrer, M.; Ottinger, E.A. Effects of cargo molecules on the cellular uptake of arginine-rich cell-penetrating epeptides. BBA Biomembranes 2005, 1712, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Ranathunga, S.; Rajapakse, N.; Kim, S.K. Purification and characterization of antioxidative peptide derived from muscle of conger eel (Conger myriaster). Eur. Food Res. Technol. 2006, 222, 310–315. [Google Scholar] [CrossRef]
- Alashi, A.M.; Blanchard, C.L.; Mailer, R.J.; Agboola, S.O.; Mawson, A.J.; He, R.; Girgih, A.; Aluko, R.E. Antioxidant properties of australian canola meal protein hydrolysates. Food Chem. 2014, 146, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Mendis, E.; Rajapakse, N.; Byun, H.G.; Kim, S.K. Investigation of jumbo squid (Dosidicus gigas) skin gelatin peptides for their in vitro antioxidant effects. Life Sci. 2005, 77, 2166–2178. [Google Scholar] [CrossRef] [PubMed]
- Bamdad, F.; Ahmed, S.; Chen, L.Y. Specifically designed peptide structures effectively suppressed oxidative reactions in chemical and cellular systems. J. Funct. Foods 2015, 18, 35–46. [Google Scholar] [CrossRef]
- Vitali, A. Proline-rich peptides: Multifunctional bioactive molecules as new potential therapeutic drugs. Curr. Protein Pept. Sci. 2015, 16, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, S.; Prakash, V. Bioactive peptides from bovine milk alpha-casein: Isolation, characterization and multifunctional properties. Int. J. Pept. Res. Ther. 2010, 16, 7–15. [Google Scholar] [CrossRef]
- Lopez-Exposito, I.; Quiros, A.; Amigo, L.; Recio, I. Casein hydrolysates as a source of antimicrobial, antioxidant and antihypertensive peptides. Lait 2007, 87, 241–249. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Chemometric analysis of the amino acid requirements of antioxidant food protein hydrolysates. Int. J. Mol. Sci. 2011, 12, 3148–3161. [Google Scholar] [CrossRef] [PubMed]
- Benkhelifa, H.; Bengoa, C.; Larre, C.; Guibal, E.; Popineau, Y.; Legrand, J. Casein hydrolysis by immobilized enzymes in a torus reactor. Process Biochem. 2005, 40, 461–467. [Google Scholar] [CrossRef]
- Tang, X.Y.; He, Z.Y.; Dai, Y.F.; Xiong, Y.L.L.; Xie, M.Y.; Chen, J. Peptide fractionation and free radical scavenging activity of zein hydrolysate. J. Agric. Food Chem. 2010, 58, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Sakanaka, S.; Tachibana, Y. Active oxygen scavenging activity of egg-yolk protein hydrolysates and their effects on lipid oxidation in beef and tuna homogenates. Food Chem. 2006, 95, 243–249. [Google Scholar] [CrossRef]
- Fogarasi, A.L.; Kun, S.; Tanko, G.; Stefanovits-Banyai, E.; Hegyesne-Vecseri, B. A comparative assessment of antioxidant properties, total phenolic content of einkorn, wheat, barley and their malts. Food Chem. 2015, 167, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, J.; Degraff, W.B.; Gamson, J.; Gazdar, A.F.; Minna, J.D.; Mitchell, J.B. Chemosentivity testing of human-lung cancer cell-lines using a colorimetric assay. Br. J. Cancer 1987, 55, 335. [Google Scholar]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [n-15]-labeled nitrate in biological-fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Chanput, W.; Mes, J.; Vreeburg, R.A.M.; Sayelkoul, H.F.J.; Wichers, H.J. Transcription profiles of LPS-stimulated THP-1 monocytes and macrophages: A tool to study inflammation modulating effects of food-derived compounds. Food Funct. 2010, 1, 254–261. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Sources | Proteolytic Activity a | Optimum Conditions |
|---|---|---|---|
| Elastase | Elastase from hog pancreas | ≥4 U/mg | pH 8; 37 °C |
| Flavourzyme | Protease from Aspergillus oryzae | ≥500 U/g | pH 7; 50 °C |
| Savinase | Protease from Bacillus sp. | ≥16 U/g | pH 7; 55 °C |
| Thermolysin | Protease from Bacillus thermoproteolyticus | 14 U/mg | pH 7; 50 °C |
| Trypsin | Protease derived from porcine pancreas | 30 U/g | pH 7; 37 °C |
| Samples | Atmospheric Hydrolysis | High Pressure Hydrolysis | ||||
|---|---|---|---|---|---|---|
| 500–1000 Da | 1000–2000 Da | >2000 Da | 500–1000 Da | 1000–2000 Da | >2000 Da | |
| Fla-CH | 0.3 | 98.6 | 1.1 | 1.9 | 95.9 | 2.2 |
| Try-CH | 4.1 | 46.5 | 49.4 | 26.5 | 23.7 | 49.8 |
| Peptide Sequence | Ion (m/z) | Observed Mass | Calculated Mass | Source (Fragment) |
|---|---|---|---|---|
| PGPIPN | 594.33 | 593.33 | 593.32 | β-Casein (78–83) |
| PFPGPIPN | 838.44 | 837.43 | 837.44 | β-Casein (76–83) |
| YPFPGPIP | 887.47 | 886.47 | 886.46 | β-Casein (75–82) |
| VYPFPGPIPN | 1100.55 | 1099.55 | 1099.57 | β-Casein (74–83) |
| MPFPKYPVEP | 610.82 (2) | 1219.62 | 1219.59 | β-Casein (124–133) |
| EPVLGPVRGPFP | 632.87 (2) | 1263.73 | 1263.70 | β-Casein (210–221) |
| QEPVLGPVRGPFP | 696.90 (2) | 1391.78 | 1391.76 | β-Casein (209–221) |
| TPVVVPPFLQPE | 661.87 (2) | 1321.73 | 1321.72 | β-Casein (95–106) |
| TQTPVVVPPFLQPE | 776.42 (2) | 1550.83 | 1550.83 | β -Casein (93–106) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bamdad, F.; Shin, S.H.; Suh, J.-W.; Nimalaratne, C.; Sunwoo, H. Anti-Inflammatory and Antioxidant Properties of Casein Hydrolysate Produced Using High Hydrostatic Pressure Combined with Proteolytic Enzymes. Molecules 2017, 22, 609. https://doi.org/10.3390/molecules22040609
Bamdad F, Shin SH, Suh J-W, Nimalaratne C, Sunwoo H. Anti-Inflammatory and Antioxidant Properties of Casein Hydrolysate Produced Using High Hydrostatic Pressure Combined with Proteolytic Enzymes. Molecules. 2017; 22(4):609. https://doi.org/10.3390/molecules22040609
Chicago/Turabian StyleBamdad, Fatemeh, Seulki Hazel Shin, Joo-Won Suh, Chamila Nimalaratne, and Hoon Sunwoo. 2017. "Anti-Inflammatory and Antioxidant Properties of Casein Hydrolysate Produced Using High Hydrostatic Pressure Combined with Proteolytic Enzymes" Molecules 22, no. 4: 609. https://doi.org/10.3390/molecules22040609