
The Suppressive Effects of Cinnamomi Cortex and Its Phytocompound Coumarin on Oxaliplatin-Induced Neuropathic Cold Allodynia in Rats

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
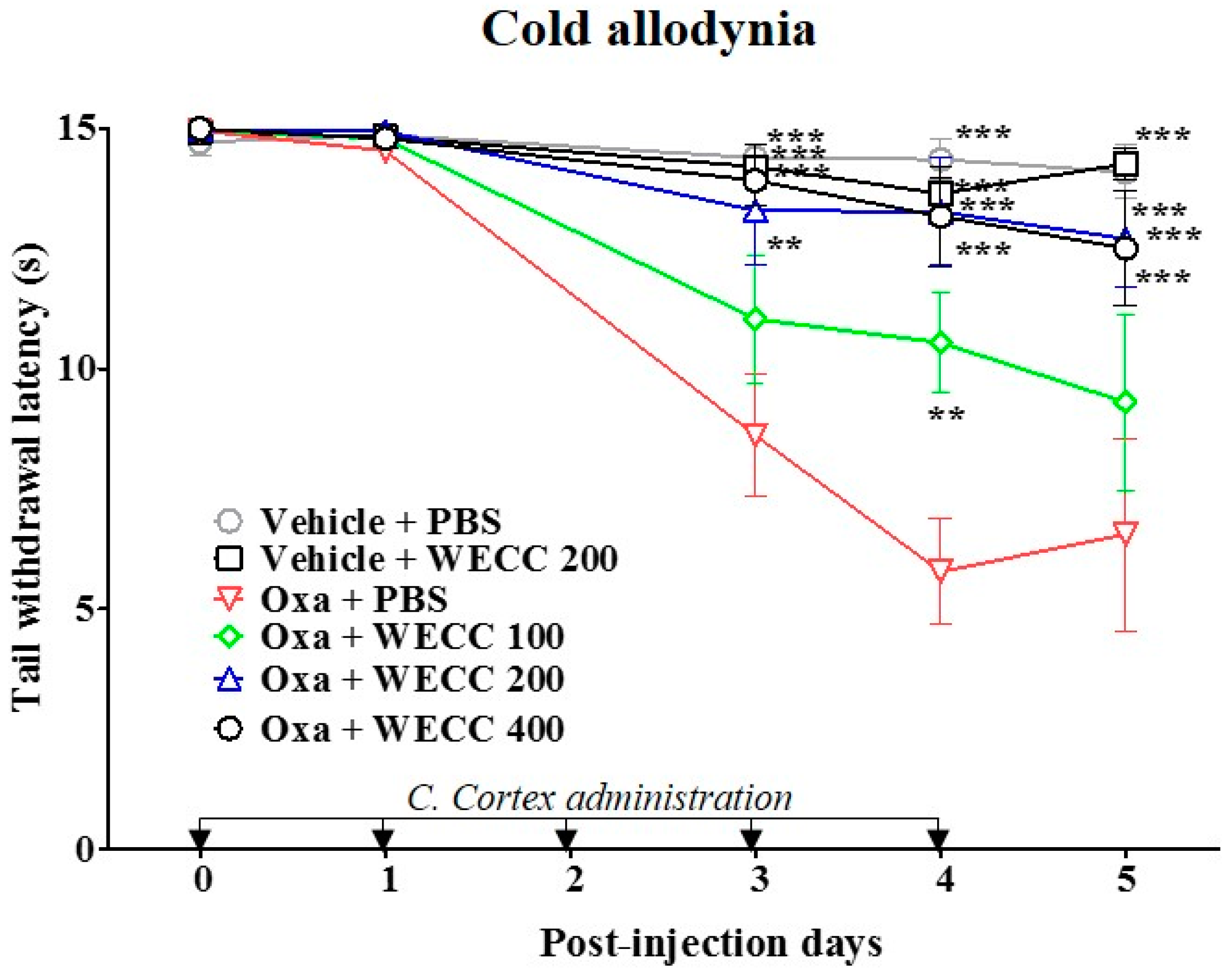
2.1. Suppresive Effect of WECC on Oxaliplatin-Induced Cold Allodynia in Rats
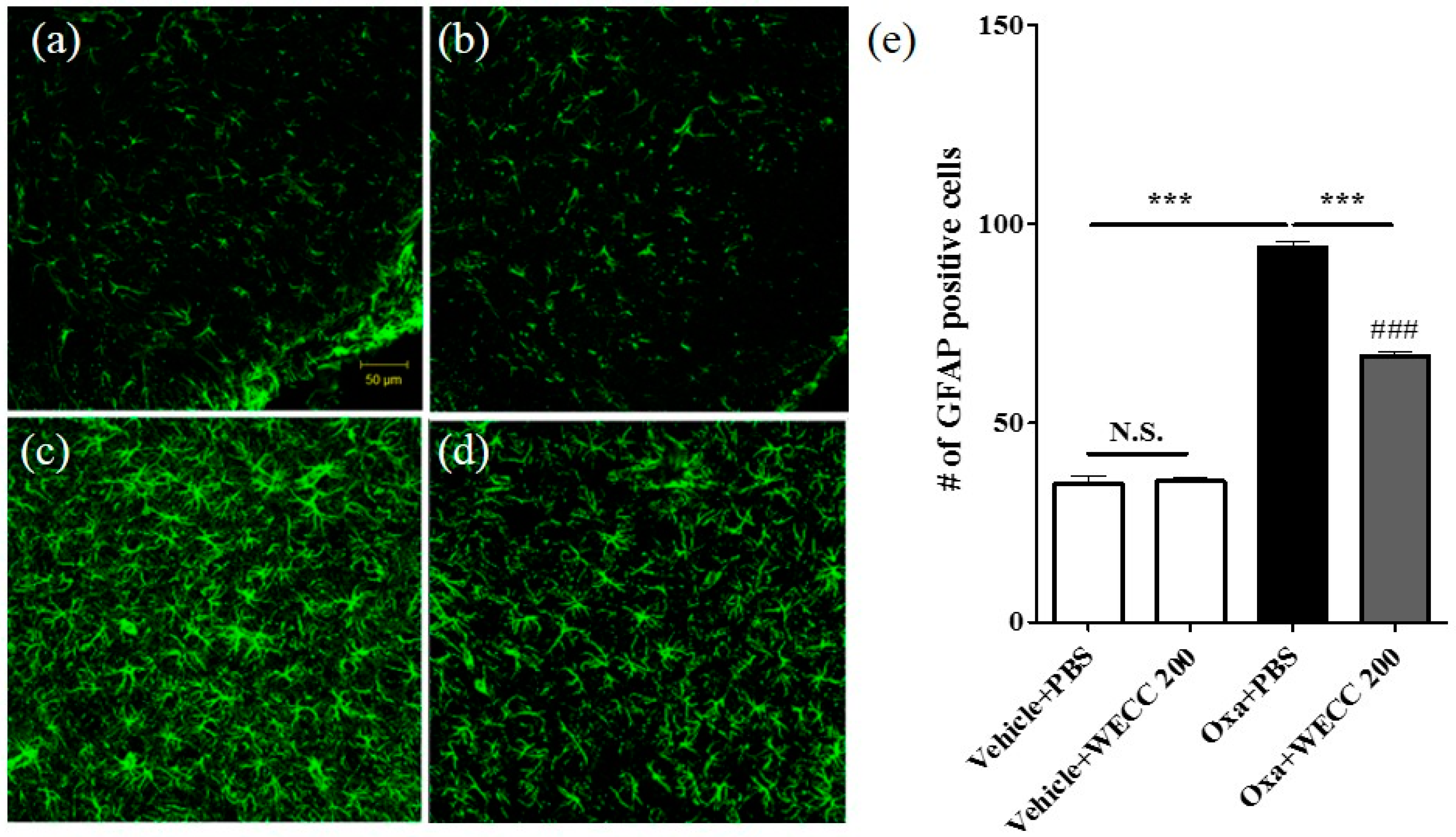
2.2. Inhibitory Effect of WECC Treatment on Activation of Spinal Astrocytes
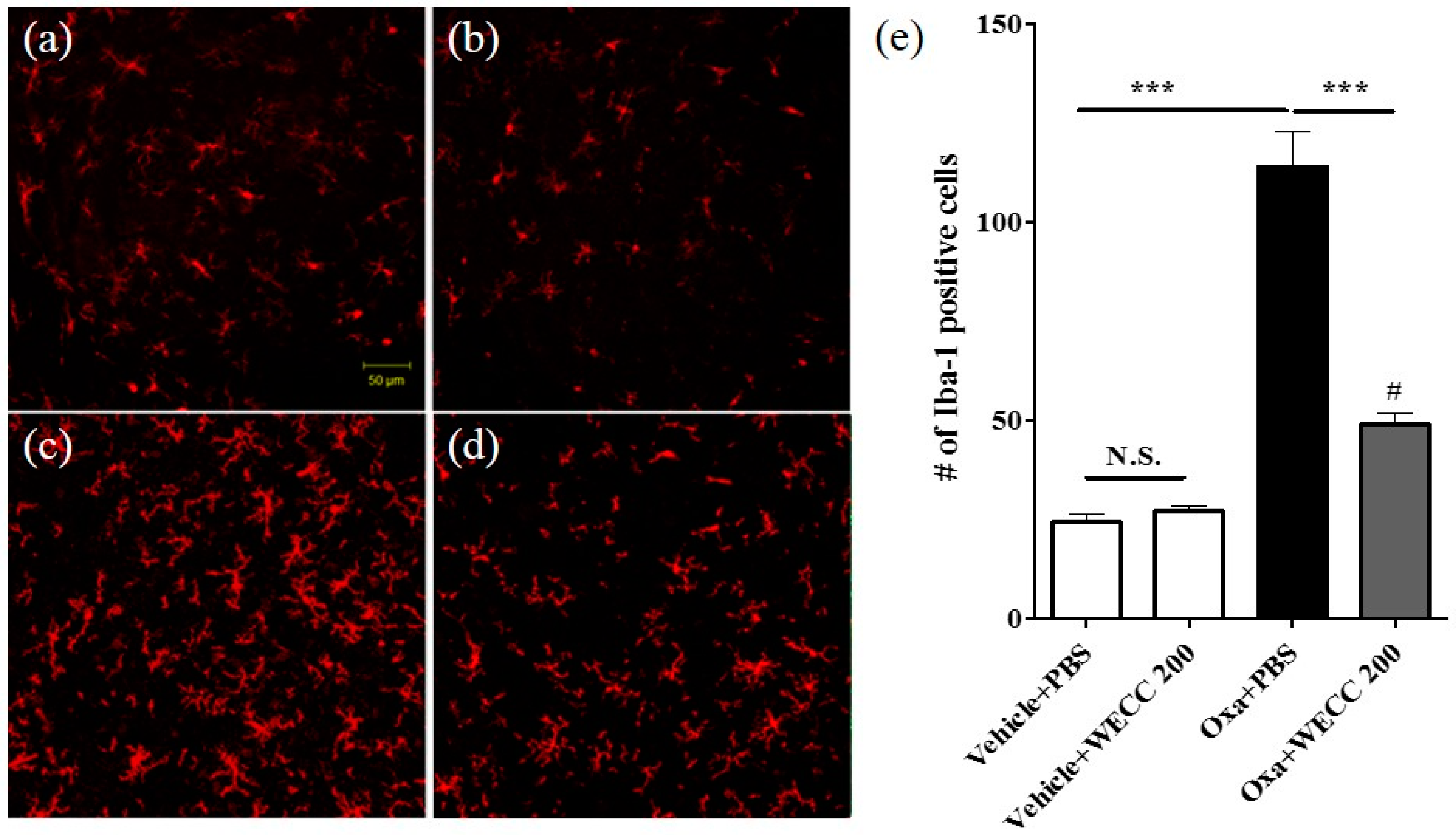
2.3. Inhibitory Effect of WECC Treatment on Spinal Microglia Activation
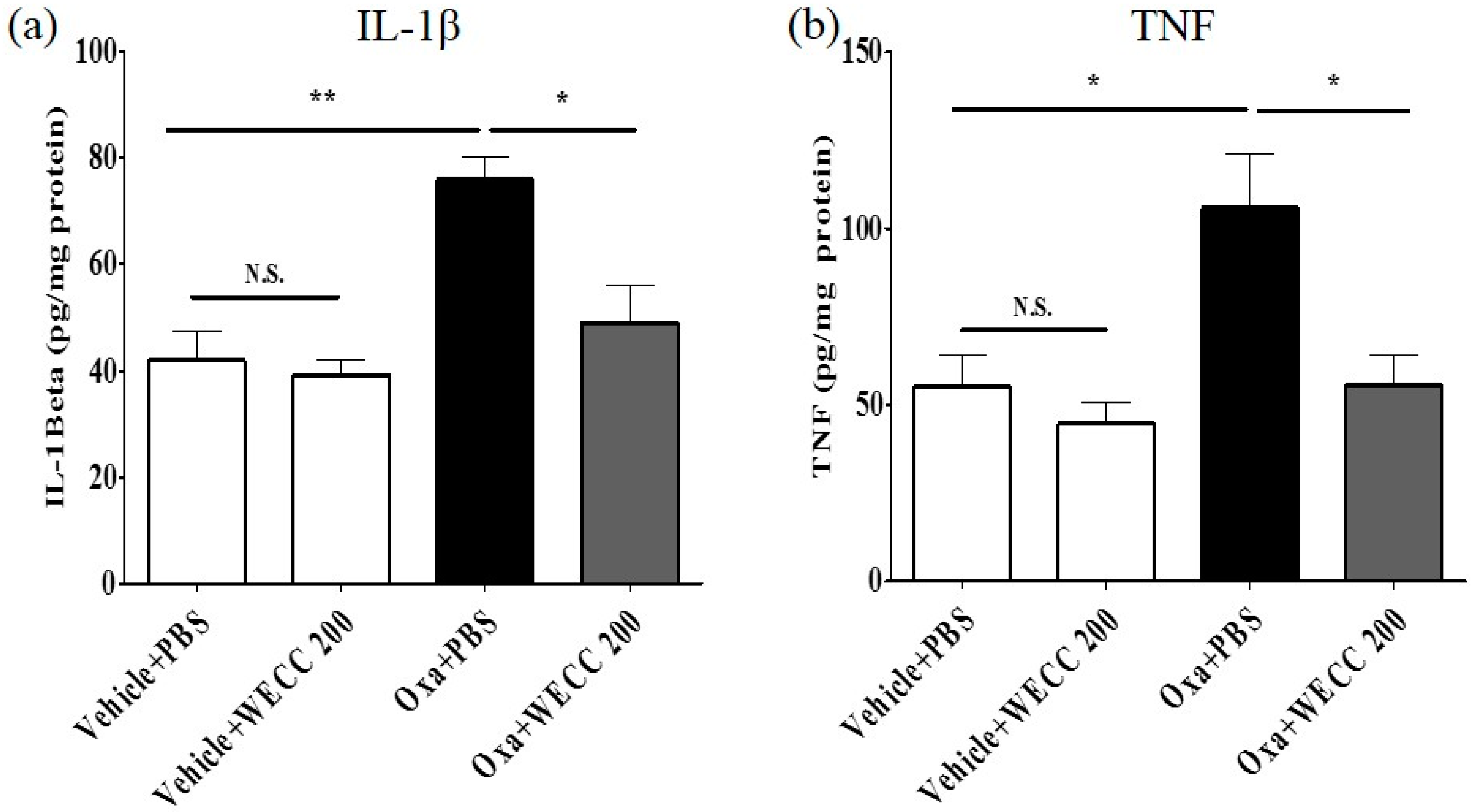
2.4. Modulatory Effect of WECC on Pro-Inflammatory Cytokines IL-1β and TNF
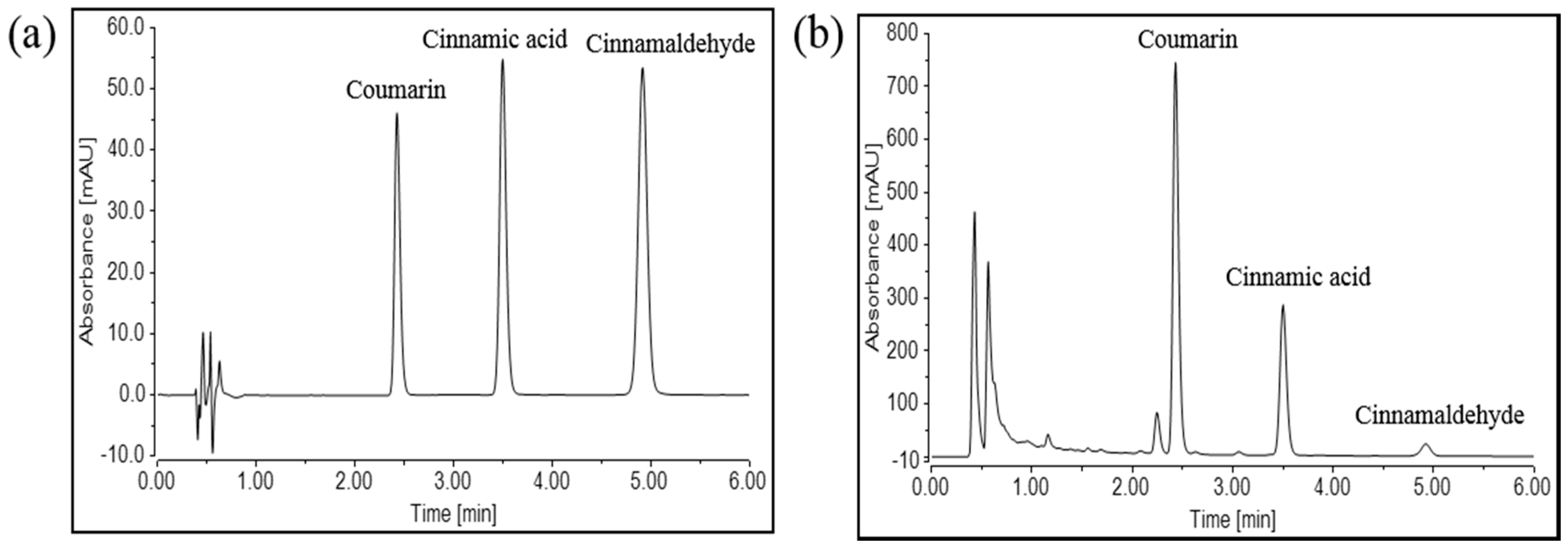
2.5. Qualitative and Quantitative Analysis of Three Chemicals in WECC
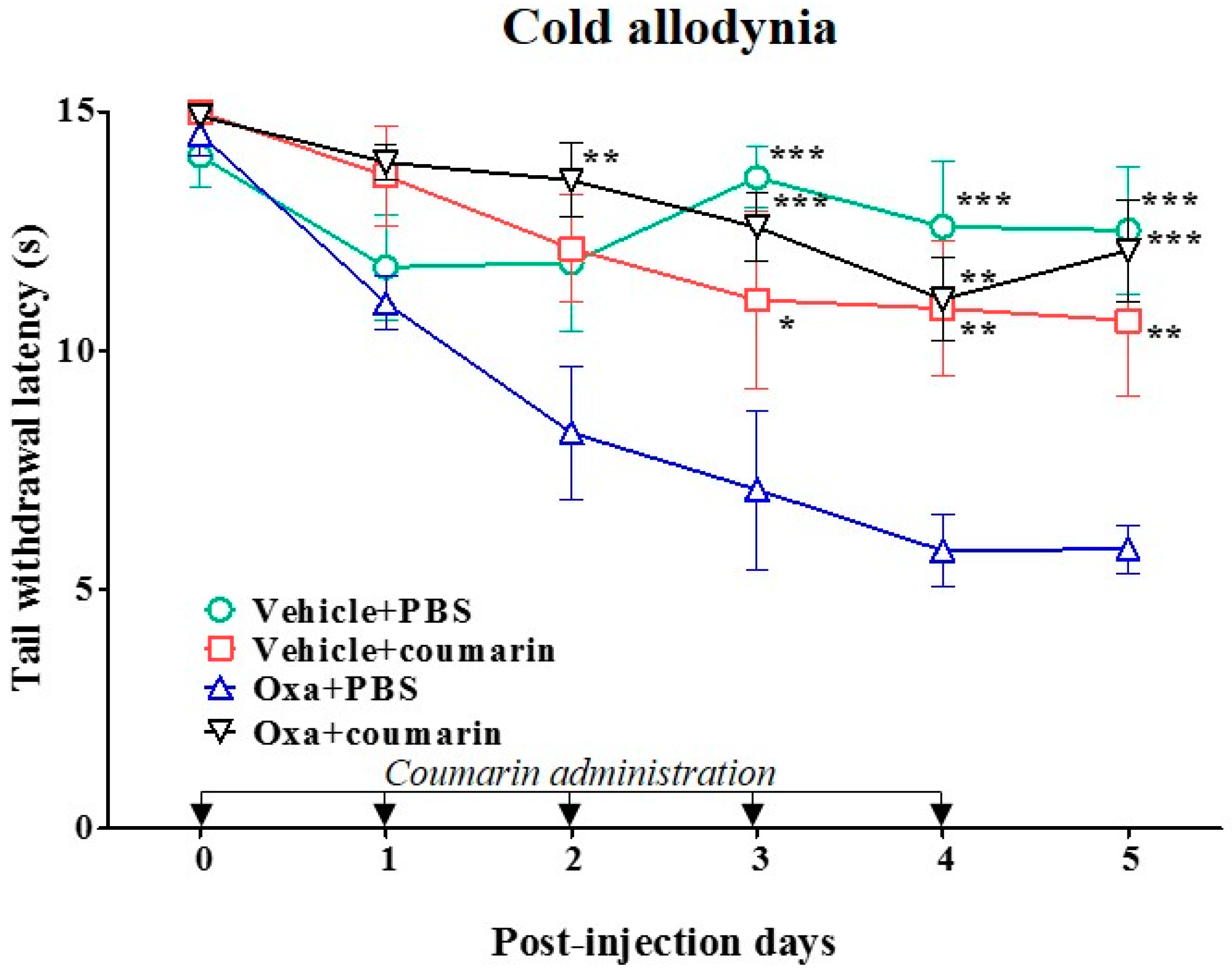
2.6. Anti-Allodynic Effect of Coumarin on Oxaliplatin-Induced Cold Allodynia in Rats
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Oxaliplatin Administration
4.3. Behavior Test
4.4. Experimental Schedule and Grouping and Treatment of WECC
4.5. Immunohistochemistry
4.6. ELISA
4.7. Ultra High-Performance Liquid Chromatography (UHPLC) Analysis
4.8. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Baker, D. Oxaliplatin: A new drug for the treatment of metastatic carcinoma of the colon or rectum. Rev. Gastroenterol. Disord. 2002, 3, 31–38. [Google Scholar]
- Argyriou, A.; Briani, C.; Cavaletti, G.; Bruna, J.; Alberti, P.; Velasco, R.; Lonardi, S.; Cortinovis, D.; Cazzaniga, M.; Campagnolo, M. Advanced age and liability to oxaliplatin-induced peripheral neuropathy: Post hoc analysis of a prospective study. Eur. J. Neurol. 2013, 20, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Petit, T.; Benider, A.; Yovine, A.; Bougnoux, P.; Spaeth, D.; Maindrault-Goebel, F.; Serin, D.; Tigaud, J.-D.; Eymard, J.C.; Simon, H. Phase II study of an oxaliplatin/vinorelbine combination in patients with anthracycline-and taxane-pre-treated metastatic breast cancer. Anticancer Drugs 2006, 17, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Desoize, B.; Madoulet, C. Particular aspects of platinum compounds used at present in cancer treatment. Crit. Rev. Oncol. Hematol. 2002, 42, 317–325. [Google Scholar] [CrossRef]
- Cassidy, J.; Misset, J.-L. Oxaliplatin-related side effects: Characteristics and management. Semin. Oncol. 2002, 29, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Shidahara, Y.; Ogawa, S.; Nakamura, M.; Nemoto, S.; Awaga, Y.; Takashima, M.; Hama, A.; Matsuda, A.; Takamatsu, H. Pharmacological comparison of a nonhuman primate and a rat model of oxaliplatin-induced neuropathic cold hypersensitivity. Pharmacol. Res. Perspect. 2016, 4, e00216. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.; Barton, D.; Kottschade, L.; Grothey, A.; Loprinzi, C. Chemotherapy-induced peripheral neuropathy: Prevention and treatment strategies. Eur. J. Cancer 2008, 44, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Tsuda, M.; Tozaki-Saitoh, H. Role of the glia in neuropathic pain caused by peripheral nerve injury. Brain Nerve 2012, 64, 1233–1239. (In Japanese) [Google Scholar] [PubMed]
- Old, E.A.; Clark, A.K.; Malcangio, M. The role of glia in the spinal cord in neuropathic and inflammatory pain. In Pain Control; Springer: Berlin, Germany, 2015; pp. 145–170. [Google Scholar]
- Mannelli, L.D.C.; Pacini, A.; Micheli, L.; Tani, A.; Zanardelli, M.; Ghelardini, C. Glial role in oxaliplatin-induced neuropathic pain. Exp. Neurol. 2014, 261, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Luongo, L.; Guida, F.; Boccella, S.; Bellini, G.; Gatta, L.; Rossi, F.; de Novellis, V.; Maione, S. Palmitoylethanolamide reduces formalin-induced neuropathic-like behaviour through spinal glial/microglial phenotypical changes in mice. CNS Neurol. Disord. Drug Targets 2013, 12, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Old, E.A.; Malcangio, M. Chemokine mediated neuron–glia communication and aberrant signalling in neuropathic pain states. Curr. Opin. Pharmacol. 2012, 12, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Ledeboer, A.; Sloane, E.M.; Milligan, E.D.; Frank, M.G.; Mahony, J.H.; Maier, S.F.; Watkins, L.R. Minocycline attenuates mechanical allodynia and proinflammatory cytokine expression in rat models of pain facilitation. Pain 2005, 115, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhang, J.; Shi, J.; Feng, Y.; Sun, Z.S.; Li, H. Antinociceptive synergistic effect of spinal mglur2/3 antagonist and glial cells inhibitor on peripheral inflammation-induced mechanical hypersensitivity. Brain Res. Bull. 2009, 79, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Scholz, J.; Woolf, C.J. The neuropathic pain triad: Neurons, immune cells and glia. Nat. Neurosci. 2007, 10, 1361–1368. [Google Scholar] [CrossRef] [PubMed]
- Miyoshi, K.; Obata, K.; Kondo, T.; Okamura, H.; Noguchi, K. Interleukin-18-mediated microglia/astrocyte interaction in the spinal cord enhances neuropathic pain processing after nerve injury. J. Neurosci. 2008, 28, 12775–12787. [Google Scholar] [CrossRef] [PubMed]
- Sommer, C.; Kress, M. Recent findings on how proinflammatory cytokines cause pain: Peripheral mechanisms in inflammatory and neuropathic hyperalgesia. Neurosci. Lett. 2004, 361, 184–187. [Google Scholar] [CrossRef] [PubMed]
- Ahn, B.-S.; Kim, S.-K.; Kim, H.N.; Lee, J.-H.; Lee, J.-H.; Hwang, D.S.; Bae, H.; Min, B.-I.; Kim, S.K. Gyejigachulbu-tang relieves oxaliplatin-induced neuropathic cold and mechanical hypersensitivity in rats via the suppression of spinal glial activation. Evid. Based Complement. Altern. Med. 2014, 2014, 436482. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.N. Gyejigachulbu-Tang Suppresses Oxaliplatin-Induced Neuropathic Mechanical Allodynia in Rats via Modulating Spinal tnf-α. Ph.D. Thesis, Kyung Hee University, Seoul, Korea, 2015. [Google Scholar]
- Hayashi, K.; Imanishi, N.; Kashiwayama, Y.; Kawano, A.; Terasawa, K.; Shimada, Y.; Ochiai, H. Inhibitory effect of cinnamaldehyde, derived from Cinnamomi Cortex, on the growth of influenza A/PR/8 virus in vitro and in vivo. Antivir. Res. 2007, 74, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kubo, M.; Ma, S.; Wu, J.; Matsuda, H. Anti-inflammatory activities of 70% methanolic extract from Cinnamomi Cortex. Biol. Pharm. Bull. 1996, 19, 1041–1045. [Google Scholar] [CrossRef] [PubMed]
- Takenaga, M.; Hirai, A.; Terano, T.; Tamura, Y.; Kitagawa, H.; Yoshida, S. In vitro effect of cinnamic aldehyde, a main component of Cinnamomi Cortex, on human platelet aggregation and arachidonic acid metabolism. J. Pharm. Dyn. 1987, 10, 201–208. [Google Scholar] [CrossRef]
- Moon, H.J.; Lim, B.-S.; Lee, D.-I.; Ye, M.S.; Lee, G.; Min, B.-I.; Bae, H.; Na, H.S.; Kim, S.K. Effects of electroacupuncture on oxaliplatin-induced neuropathic cold hypersensitivity in rats. J. Physiol. Sci. 2014, 64, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-H.; Sim, Y.-B.; Kang, Y.-J.; Kim, S.-S.; Kim, C.-H.; Kim, S.-J.; Lim, S.-M.; Suh, H.-W. Antinociceptive profiles and mechanisms of orally administered coumarin in mice. Biol. Pharm. Bull. 2013, 36, 925–930. [Google Scholar] [CrossRef] [PubMed]
- Lehky, T.; Leonard, G.; Wilson, R.; Grem, J.L.; Floeter, M. Oxaliplatin-induced neurotoxicity: Acute hyperexcitability and chronic neuropathy. Muscle Nerve 2004, 29, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Pasetto, L.M.; D’Andrea, M.R.; Rossi, E.; Monfardini, S. Oxaliplatin-related neurotoxicity: How and why? Crit. Rev. Oncol. Hematol. 2006, 59, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Ormseth, M.J.; Scholz, B.A.; Boomershine, C.S. Duloxetine in the management of diabetic peripheral neuropathic pain. Patient Prefer Adherence 2011, 5, 343–356. [Google Scholar] [PubMed]
- Serpell, M.; Neuropathic pain study group. Gabapentin in neuropathic pain syndromes: A randomised, double-blind, placebo-controlled trial. Pain 2002, 99, 557–566. [Google Scholar] [CrossRef]
- Sweitzer, S.; Martin, D.; DeLeo, J. Intrathecal interleukin-1 receptor antagonist in combination with soluble tumor necrosis factor receptor exhibits an anti-allodynic action in a rat model of neuropathic pain. Neuroscience 2001, 103, 529–539. [Google Scholar] [CrossRef]
- Austin, P.J.; Moalem-Taylor, G. The neuro-immune balance in neuropathic pain: Involvement of inflammatory immune cells, immune-like glial cells and cytokines. J. Neuroimmunol. 2010, 229, 26–50. [Google Scholar] [CrossRef] [PubMed]
- Chauvet, N.; Palin, K.; Verrier, D.; Poole, S.; Dantzer, R.; Lestage, J. Rat microglial cells secrete predominantly the precursor of interleukin-1β in response to lipopolysaccharide. Eur. J. Neurosci. 2001, 14, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Shigemoto-Mogami, Y.; Koizumi, S.; Tsuda, M.; Ohsawa, K.; Kohsaka, S.; Inoue, K. Mechanisms underlying extracellular ATP-evoked interleukin-6 release in mouse microglial cell line, MG-5. J. Neurochem. 2001, 78, 1339–1349. [Google Scholar] [CrossRef] [PubMed]
- Sachs, D.; Cunha, F.Q.; Poole, S.; Ferreira, S.H. Tumour necrosis factor-α, interleukin-1β and interleukin-8 induce persistent mechanical nociceptor hypersensitivity. Pain 2002, 96, 89–97. [Google Scholar] [CrossRef]
- Nadeau, S.; Filali, M.; Zhang, J.; Kerr, B.J.; Rivest, S.; Soulet, D.; Iwakura, Y.; de Rivero Vaccari, J.P.; Keane, R.W.; Lacroix, S. Functional recovery after peripheral nerve injury is dependent on the pro-inflammatory cytokines IL-1β and TNF: Implications for neuropathic pain. J. Neurosci. 2011, 31, 12533–12542. [Google Scholar] [CrossRef] [PubMed]
- Shamash, S.; Reichert, F.; Rotshenker, S. The cytokine network of wallerian degeneration: Tumor necrosis factor-α, interleukin-1α, and interleukin-1β. J. Neurosci. 2002, 22, 3052–3060. [Google Scholar] [PubMed]
- Homma, Y.; Brull, S.J.; Zhang, J.-M. A comparison of chronic pain behavior following local application of tumor necrosis factor α to the normal and mechanically compressed lumbar ganglia in the rat. Pain 2002, 95, 239–246. [Google Scholar] [CrossRef]
- Murata, Y.; Onda, A.; Rydevik, B.; Takahashi, I.; Takahashi, K.; Olmarker, K. Changes in pain behavior and histologic changes caused by application of tumor necrosis factor-alpha to the dorsal root ganglion in rats. Spine 2006, 31, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Schroder, S.; Beckmann, K.; Franconi, G.; Meyer-Hamme, G.; Friedemann, T.; Greten, H.J.; Rostock, M.; Efferth, T. Can medical herbs stimulate regeneration or neuroprotection and treat neuropathic pain in chemotherapy-induced peripheral neuropathy? Evid. Based Complement. Altern. Med. 2013, 2013, 423713. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Wu, E.Q.; Liang, C.; Chen, J.; Tran, M.N.; Hong, C.H.; Jang, Y.; Park, K.L.; Bae, K.; Kim, Y.H. Discrimination of cinnamon bark and cinnamon twig samples sourced from various countries using hplc-based fingerprint analysis. Food Chem. 2011, 127, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, J.; Ma, D.; Lin, H.; Xu, X.; Jiang, Y.H. Effects of different compatibilities of ramulus cinnamomi and peony in guizhi decoction on diabetic cardiac autonomic neuropathy. Zhongguo Zhong Xi Yi Jie He Za Zhi 2015, 35, 741. (In Chinese) [Google Scholar] [PubMed]
- Zhao, N.; Li, J.; Li, L.; Niu, X.-Y.; Jiang, M.; He, X.-J.; Bian, Z.-X.; Zhang, G.; Lu, A.-P. Molecular network-based analysis of guizhi-shaoyao-zhimu decoction, a TCM herbal formula, for treatment of diabetic peripheral neuropathy. Acta Pharmacol. Sin. 2015, 36, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Zhiling, Y.; Guangqin, Z.; Dai Yue, K.J.; Changgui, D.; Ruijian, L. Pharmacological study on compatibility of cortex cinnamomi with halloysitum rubrum. China J. Chin. Mater. Med. 1997, 22, 309–312. [Google Scholar]
- Saghir, S.; Sadikun, A.; Al-Suede, F.; Abdul, M.A.; Murugaiyah, V. Antihyperlipidemic, antioxidant and cytotoxic activities of methanolic and aqueous extracts of different parts of star fruit. Curr. Pharm. Biotechnol. 2016, 17, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Avula, B.; Nanayakkara, N.P.; Zhao, J.; Khan, I.A. Cassia cinnamon as a source of coumarin in cinnamon-flavored food and food supplements in the united states. J. Agric. Food Chem. 2013, 61, 4470–4476. [Google Scholar] [CrossRef] [PubMed]
- Meotti, F.C.; Missau, F.C.; Ferreira, J.; Pizzolatti, M.G.; Mizuzaki, C.; Nogueira, C.W.; Santos, A.R. Anti-allodynic property of flavonoid myricitrin in models of persistent inflammatory and neuropathic pain in mice. Biochem. Pharmacol. 2006, 72, 1707–1713. [Google Scholar] [CrossRef] [PubMed]
- De Lima, F.O.; Nonato, F.R.; Couto, R.D.; Barbosa Filho, J.M.; Nunes, X.P.; Ribeiro dos Santos, R.; Soares, M.B.P.; Villarreal, C.F. Mechanisms involved in the antinociceptive effects of 7-hydroxycoumarin. J. Nat. Prod. 2011, 74, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Hashizume, H.; DeLeo, J.A.; Colburn, R.W.; Weinstein, J.N. Spinal glial activation and cytokine expression after lumbar root injury in the rat. Spine 2000, 25, 1206–1217. [Google Scholar] [CrossRef] [PubMed]
- Shibata, K.; Sugawara, T.; Fujishita, K.; Shinozaki, Y.; Matsukawa, T.; Suzuki, T.; Koizumi, S. The astrocyte-targeted therapy by bushi for the neuropathic pain in mice. PLoS ONE 2011, 6, e23510. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef]
- Ling, B.; Coudoré-Civiale, M.-A.; Balayssac, D.; Eschalier, A.; Coudoré, F.; Authier, N. Behavioral and immunohistological assessment of painful neuropathy induced by a single oxaliplatin injection in the rat. Toxicology 2007, 234, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Ling, B.; Coudoré, F.; Decalonne, L.; Eschalier, A.; Authier, N. Comparative antiallodynic activity of morphine, pregabalin and lidocaine in a rat model of neuropathic pain produced by one oxaliplatin injection. Neuropharmacology 2008, 55, 724–728. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Sohn, S.-H.; Jung, K.-H.; Lee, K.-Y.; Yeom, Y.R.; Kim, G.-E.; Jung, S.; Jung, H.; Bae, H. The effects of maekmoondong-tang on cockroach extract-induced allergic asthma. Evid.-Based Complement. Altern. Med. 2014, 2014, 958965. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds are available from the authors.






© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, C.; Lee, J.H.; Kim, W.; Li, D.; Kim, Y.; Lee, K.; Kim, S.K. The Suppressive Effects of Cinnamomi Cortex and Its Phytocompound Coumarin on Oxaliplatin-Induced Neuropathic Cold Allodynia in Rats. Molecules 2016, 21, 1253. https://doi.org/10.3390/molecules21091253
Kim C, Lee JH, Kim W, Li D, Kim Y, Lee K, Kim SK. The Suppressive Effects of Cinnamomi Cortex and Its Phytocompound Coumarin on Oxaliplatin-Induced Neuropathic Cold Allodynia in Rats. Molecules. 2016; 21(9):1253. https://doi.org/10.3390/molecules21091253
Chicago/Turabian StyleKim, Changmin, Ji Hwan Lee, Woojin Kim, Dongxing Li, Yangseok Kim, Kyungjin Lee, and Sun Kwang Kim. 2016. "The Suppressive Effects of Cinnamomi Cortex and Its Phytocompound Coumarin on Oxaliplatin-Induced Neuropathic Cold Allodynia in Rats" Molecules 21, no. 9: 1253. https://doi.org/10.3390/molecules21091253