
Polyphenols from Erythrina crista-galli: Structures, Molecular Docking and Phytoestrogenic Activity


 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
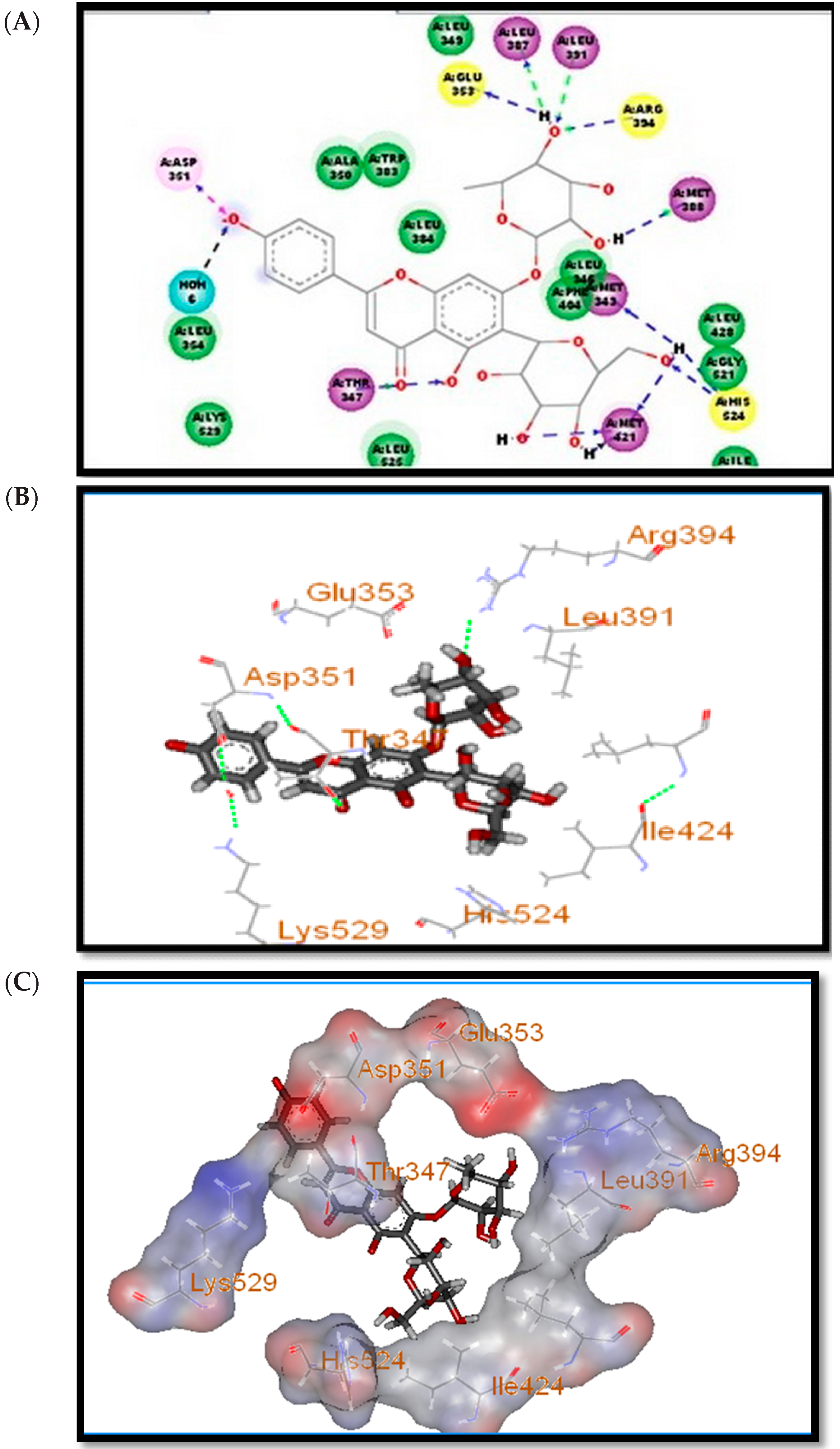
2.1. Docking Study
2.2. Phytoestrogenic Activity (pER8: GUS Reporter Assay)
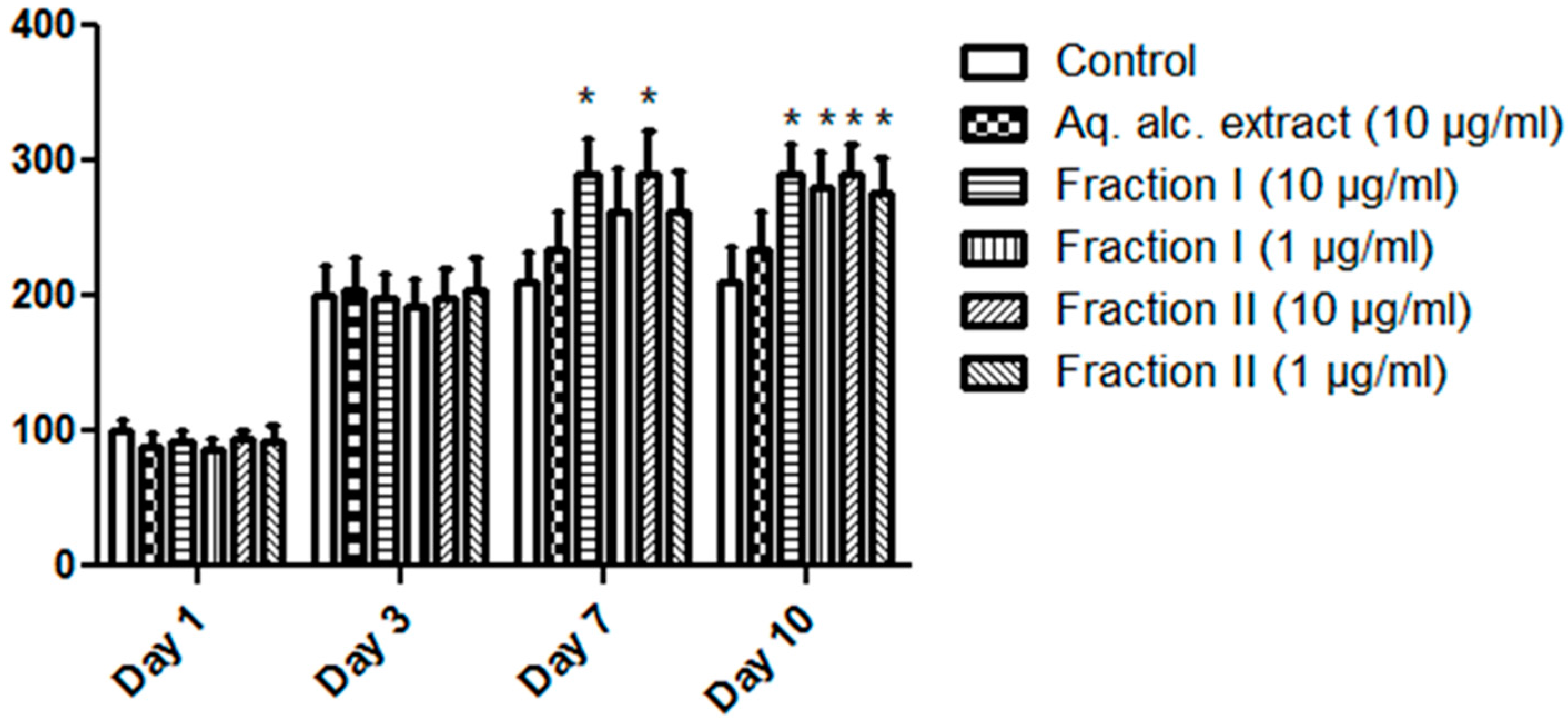
2.3. Proliferation-Enhancing Activity in MCF-7 Cells
3. Materials and Methods
3.1. Plant Material
3.2. Cell Lines and Chemicals
3.3. Phytochemistry
3.4. Extraction and Isolation
3.5. Molecular Docking Study
3.6. Biological Activity
3.6.1. Estrogenic Activity: pER8: GUS Reporter System
3.6.2. Cell Culture, Cytotoxicity and Cell Proliferation Assay
3.6.3. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wink, M. Evolutionary advantage and molecular modes of action of multi-component mixtures used in phytomedicine. Curr. Drug Metab. 2008, 9, 996–1009. [Google Scholar] [CrossRef] [PubMed]
- Psotova, J.; Kolář, M.; Soušek, J.; Švagera, Z.; Vičar, J.; Ulrichová, J. Biological activities of Prunella vulgaris extract. Phytother. Res. 2003, 17, 1082–1087. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, L.; Mazza, G. Assessing antioxidant and prooxidant activities of phenolic compounds. J. Agric. Food Chem. 2000, 48, 3597–3604. [Google Scholar] [CrossRef] [PubMed]
- Nomura, T.; Fukai, T.; Akiyama, T. Chemistry of phenolic compounds of licorice (Glycyrrhiza species) and their estrogenic and cytotoxic activities. Pure Appl. Chem. 2002, 74, 1199–1206. [Google Scholar] [CrossRef]
- Miksicek, R.J. Commonly occurring plant flavonoids have estrogenic activity. Mol. Pharmacol. 1993, 44, 37–43. [Google Scholar] [PubMed]
- Rao, Y.K.; Geethangili, M.; Fang, S.-H.; Tzeng, Y.-M. Antioxidant and cytotoxic activities of naturally occurring phenolic and related compounds: A comparative study. Food Chem. Toxicol. 2007, 45, 1770–1776. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Ninomiya, K.; Shimoda, H.; Nishida, N.; Matsuda, H. Hepatoprotective and antioxidative properties of Salacia reticulata: Preventive effects of phenolic constituents on CCl4-induced liver injury in mice. Biol. Pharm. Bull. 2002, 25, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Soni, M.; Deb, L.; Jain, A.; Rout, S.; Gupta, V.; Krishna, K. Antioxidant and hepatoprotective activity of ethanolic and aqueous extracts of Momordica dioica Roxb. Leaves. J. Ethnopharmacol. 2008, 115, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Hegnauer, R.; Renpe, J.; Gpayer-Barkmeijer, R.J. Relevance of seed polysaccharides and flavonoids for the classification of the leguminosae: A chemotaxonomic approach. Phytochemistry 1993, 34, 3–16. [Google Scholar] [CrossRef]
- Nestel, P.J.; Pomeroy, S.; Kay, S.; Komesaroff, P.; Behrsing, J.; Cameron, J.D.; West, L. Isoflavones from red clover improve systemic arterial compliance but not plasma lipids in menopausal women. J. Clin. Endocrinol. Metab. 1999, 84, 895–898. [Google Scholar] [CrossRef] [PubMed]
- Tindale, M.D.; Roux, D. A phytochemical survey of the Australian species of Acacia. Phytochemistry 1969, 8, 1713–1727. [Google Scholar] [CrossRef]
- Li, W.; Asada, Y.; Yoshikawa, T. Flavonoid constituents from Glycyrrhiza glabra hairy root cultures. Phytochemistry 2000, 55, 447–456. [Google Scholar] [CrossRef]
- Wang, G.-S.; Han, Z.-W. Effects of flavonoids of glycyrrhiza on ethanol-induced liver injury in mice. Chin. Pharmacol. Bull. 1993, 9, 271–273. [Google Scholar]
- Kone, W.M.; Solange, K.-N.E.; Dosso, M. Assessing sub-Saharan Erythrina for efficacy: Traditional uses, biological activities and phytochemistry. Pak. J. Biol. Sci. 2011, 14, 560–571. [Google Scholar] [PubMed]
- De Lima, M.R.F.; de Souza Luna, J.; dos Santos, A.F.; de Andrade, M.C.C.; Sant’Ana, A.E.G.; Genet, J.-P.; Marquez, B.; Neuville, L.; Moreau, N. Anti-bacterial activity of some Brazilian medicinal plants. J. Ethnopharmacol. 2006, 105, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Flausino, O.A., Jr.; Pereira, A.M.; da Silva Bolzani, V.; Nunes-de-Souza, R.L. Effects of erythrinian alkaloids isolated from Erythrina mulungu (Papilionaceae) in mice submitted to animal models of anxiety. Biol. Pharm. Bull. 2007, 30, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Juma, B.F.; Majinda, R.R.T. Erythrinaline alkaloids from the flowers and pods of Erythrina lysistemon and their DPPH radical scavenging properties. Phytochemistry 2004, 65, 1397–1404. [Google Scholar] [CrossRef] [PubMed]
- Wanjala, C.C.; Juma, B.F.; Bojase, G.; Gashe, B.A.; Majinda, R.R. Erythrinaline alkaloids and antimicrobial flavonoids from Erythrina latissima. Planta Med. 2002, 68, 640–642. [Google Scholar] [CrossRef] [PubMed]
- Djiogue, S.; Halabalaki, M.; Njamen, D.; Kretzschmar, G.; Lambrinidis, G.; Hoepping, J.; Raffaelli, F.M.; Mikros, E.; Skaltsounis, A.L.; Vollmer, G. Erythroidine alkaloids: A novel class of phytoestrogens. Planta Med. 2014, 80, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Hikita, K.; Tanaka, H.; Murata, T.; Kato, K.; Hirata, M.; Sakai, T.; Kaneda, N. Phenolic constituents from stem bark of Erythrina poeppigiana and their inhibitory activity on human glyoxalase I. J. Nat. Med. 2014, 68, 636–642. [Google Scholar] [CrossRef] [PubMed]
- KamdemWaffo, A.F.; Coombes, P.H.; Mulholland, D.A.; Nkengfack, A.E.; Fomum, Z.T. Flavones and isoflavones from the west African Fabaceae Erythrina vogelii. Phytochemistry 2006, 67, 459–463. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Atsumi, I.; Shirota, O.; Sekita, S.; Sakai, E.; Sato, M.; Murata, J.; Murata, H.; Darnaedi, D.; Chen, I.S. Three new constituents from the roots of Erythrina variegata and their antibacterial activity against methicillin-resistant Staphylococcus aureus. Chem. Biodivers. 2011, 8, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Chukwujekwu, J.C.; van Heerden, F.R.; van Staden, J. Antibacterial activity of flavonoids from the stem bark of Erythrina caffrathunb. Phytother. Res. 2011, 25, 46–48. [Google Scholar] [CrossRef] [PubMed]
- Innok, P.; Rukachaisirikul, T.; Phongpaichit, S.; Suksamrarn, A. Fuscacarpans A–C, new pterocarpans from the stems of Erythrina fusca. Fitoterapia 2010, 81, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Dao, T.T.; Nguyen, P.H.; Thuong, P.T.; Kang, K.W.; Na, M.K.; Ndinteh, D.T.; Mbafor, J.T.; Oh, W.K. Pterocarpans with inhibitory effects on protein tyrosine phosphatase 1B from Erythrina lysistemon Hutch. Phytochemistry 2009, 70, 2053–2057. [Google Scholar] [CrossRef] [PubMed]
- Innok, P.; Rukachaisirikul, T.; Suksamrarn, A. Flavanoids and pterocarpans from the bark of Erythrinafusca. Chem. Pharm. Bull. 2009, 57, 993–996. [Google Scholar] [CrossRef] [PubMed]
- Watjen, W.; Suckow-Schnitker, A.K.; Rohrig, R.; Kulawik, A.; Addae-Kyereme, J.; Wright, C.W.; Passreiter, C.M. Prenylated flavonoid derivatives from the bark of Erythrina addisoniae. J. Nat. Prod. 2008, 71, 735–738. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Na, M.; Thuong, P.T.; Njamen, D.; Mbafor, J.T.; Fomum, Z.T.; Woo, E.R.; Oh, W.K. Prenylated flavonoids with PTP1B inhibitory activity from the root bark of Erythrina mildbraedii. Chem. Pharm. Bull. 2008, 56, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Thuong, P.T.; Lee, H.S.; Njamen, D.; Mbafor, J.T.; Fomum, Z.T.; Lee, J.; Kim, Y.H.; Oh, W.K. Four new chalcones from Erythrina abyssinica. Planta Med. 2008, 74, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Na, M.; Jang, J.; Njamen, D.; Mbafor, J.T.; Fomum, Z.T.; Kim, B.Y.; Oh, W.K.; Ahn, J.S. Protein tyrosine phosphatase-1B inhibitory activity of isoprenylated flavonoids isolated from Erythrina mildbraedii. J. Nat. Prod. 2006, 69, 1572–1576. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Tanaka, H.; Yamaguchi, R.; Oh-Uchi, T.; Etoh, H. Erythrinapoeppigiana-derived phytochemical exhibiting antimicrobial activity against Candida albicans and methicillin-resistant Staphylococcus aureus. Lett. Appl. Microbiol. 2003, 37, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Iinuma, M.; Okawa, Y.; Tanaka, T. Three new cinnamylphenols in heartwood of Erythrina crista-galli. Phytochemistry 1994, 37, 1153–1155. [Google Scholar] [CrossRef]
- Tanaka, H.; Sudo, M.; Kawamura, T.; Sato, M.; Yamaguchi, R.; Fukai, T.; Sakai, E.; Tanaka, N. Antibacterial constituents from the roots of Erythrina herbacea against methicillin-resistant Staphylococcus aureus. Planta Med. 2010, 76, 916–919. [Google Scholar] [CrossRef] [PubMed]
- Andayi, A.W.; Yenesew, A.; Derese, S.; Midiwo, J.O.; Gitu, P.M.; Jondiko, O.J.I.; Akala, H.; Liyala, P.; Wangui, J.; Waters, N.C. Antiplasmodial flavonoids from Erythrina sacleuxii. Planta Med. 2006, 72, 187–188. [Google Scholar] [CrossRef] [PubMed]
- Sakat, S.; Juvekar, A. Comparative study of Erythrina indica Lam. (Febaceae) leaves extracts for antioxidant activity. J. Young Pharm. 2010, 2, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Njamen, D.; Mbafor, J.T.; Fomum, Z.T.; Kamanyi, A.; Mbanya, J.C.; Recio, M.C.; Giner, R.M.; Manez, S.; Rios, J.L. Anti-inflammatory activities of two flavanones, sigmoidin A and sigmoidin B, from Erythrina sigmoidea. Planta Med. 2004, 70, 104–107. [Google Scholar] [PubMed]
- Agrawal, S.K.; Agrawal, M.; Sharma, P.R.; Gupta, B.D.; Arora, S.; Saxena, A.K. Induction of apoptosis in human promyelocytic leukemia HL60 cells by an extract from Erythrina suberosa stem bark. Nutr. Cancer 2011, 63, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Kuete, V.; Sandjo, L.P.; Djeussi, D.E.; Zeino, M.; Kwamou, G.M.; Ngadjui, B.; Efferth, T. Cytotoxic flavonoids and isoflavonoids from Erythrina sigmoidea towards multi-factorial drug resistant cancer cells. Investig. New Drugs 2014, 32, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- RathiSre, P.R.; Reka, M.; Poovazhagi, R.; Arul Kumar, M.; Murugesan, K. Antibacterial and cytotoxic effect of biologically synthesized silver nanoparticles using aqueous root extract of Erythrina indica Lam. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2015, 135, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.H.; Le, T.V.; Thuong, P.T.; Dao, T.T.; Ndinteh, D.T.; Mbafor, J.T.; Kang, K.W.; Oh, W.K. Cytotoxic and PTP1B inhibitory activities from Erythrina abyssinica. Bioorg. Med. Chem. Lett. 2009, 19, 6745–6749. [Google Scholar] [CrossRef] [PubMed]
- Djiogue, S.; Njamen, D.; Halabalaki, M.; Kretzschmar, G.; Beyer, A.; Mbanya, J.C.; Skaltsounis, A.L.; Vollmer, G. Estrogenic properties of naturally occurring prenylated isoflavones in U2OS human osteosarcoma cells: Structure-activity relationships. J. Steroid. Biochem. Mol. Biol. 2010, 120, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Njamen, D.; Djiogue, S.; Zingue, S.; Mvondo, M.A.; Nkeh-Chungag, B. In vivo and in vitro estrogenic activity of extracts from Erythrina poeppigiana (Fabaceae). J. Complement. Integr. Med. 2013, 10, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, Q.; Li, X.; Wan, H.Y.; Wong, M.S. Erythrina variegata extract exerts osteoprotective effects by suppression of the process of bone resorption. Br. J. Nutr. 2010, 104, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, S.M.; Macedo, D.S.; de Melo, C.T.; Paiva Monteiro, A.; Rodrigues, A.C.; Silveira, E.R.; Cunha, G.M.; Sousa, F.C.; Viana, G.S. Central activity of hydro alcoholic extracts from Erythrina velutina and Erythrin amulungu in mice. J. Pharm. Pharmacol. 2004, 56, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, G. Illustrated Cyclopedia of Brazilian Medicinal Plants; ABOC-SHA: Kamakura, Japan, 1996; pp. 678–679. [Google Scholar]
- Ozawa, M.; Kawamata, S.; Etoh, T.; Hayashi, M.; Komiyama, K.; Kishida, A.; Kuroda, C.; Ohsaki, A. Structures of new erythrinan alkaloids and nitric oxide production inhibitors from Erythrina crista-galli. Chem. Pharm. Bull. 2010, 58, 1119–1122. [Google Scholar] [CrossRef] [PubMed]
- Ramarathnam, N.; Osawa, T.; Namiki, M.; Kawakishi, S. Chemical studies on novel rice hull antioxidants. 2. Identification of isovitexin, a C-glycosyl flavonoid. J. Agric. Food Chem. 1989, 37, 316–319. [Google Scholar] [CrossRef]
- Koeppen, B.; Roux, D. C-Glycosylflavonoids. The chemistry of orientin and iso-orientin. Biochem. J. 1965, 97, 444–448. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Fan, G.; Hong, Z.; Chai, Y.; Wu, Y. Preparative separation of isovitexin and isoorientin from Patrinia villosa Juss by high-speed counter-current chromatography. J. Chromatogr. A 2005, 1074, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Mabry, T.J.; Markham, K.R.; Thomas, M.B. The Systematic Identification of Flavonoids; Springer Verlag: New York, NY, USA; Heidelberg/Berlin, Germany, 1970; pp. 253–273. [Google Scholar]
- Celik, L.; Lund, J.D.; Schiøtt, B. Conformational dynamics of the estrogen receptor α: Molecular dynamics simulations of the influence of binding site structure on protein dynamics. Biochem. J. 2007, 46, 1743–1758. [Google Scholar] [CrossRef] [PubMed]
- Chang, F.R.; Hayashi, K.; Chua, N.H.; Kamio, S.; Huang, Z.Y.; Nozaki, H.; Wu, Y.C. The transgenic Arabidopsis plant system, pER8-GFP, as a powerful tool in searching for natural product estrogen-agonists/antagonists. J. Nat. Prod. 2005, 68, 971–973. [Google Scholar] [CrossRef] [PubMed]
- MagneNde, C.B.; Njamen, D.; TaneeFomum, S.; Wandji, J.; Simpson, E.; Clyne, C.; Vollmer, G. In vitro estrogenic activity of two major compounds from the stem bark of Erythrina lysistemon (Fabaceae). Eur. J. Pharmacol. 2012, 674, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Hillerns, P.I.; Wink, M. Binding of flavonoids from Sophora flavescens to the rat uterine estrogen receptor. Planta Med. 2005, 71, 1065–1068. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.C.; Chiang, S.Y.; El-Shazly, M.; Wu, C.C.; Beerhues, L.; Lai, W.C.; Wu, S.F.; Yen, M.H.; Wu, Y.C.; Chang, F.R. The oestrogenic and anti-platelet activities of dihydrobenzofuroisocoumarins and homoisoflavonoids from Liriope platyphylla roots. Food Chem. 2013, 140, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, M.M.; Ibrahim, N.A.; Awad, N.E.; Matloub, A.A.; Mohamed-Ali, A.G.; Barakat, E.E.; Mohamed, A.E.; Colla, P.L. Anti-HIV-1 and cytotoxicity of the alkaloids of Erythrina abyssinica Lam. growing in Sudan. Nat. Prod. Res. 2012, 26, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.; Bladt, S. Plant Drug Analysis, A Thin Layer Chromatography Atlas, 2nd ed.; Springer-Verlag: Berlin/Heidelberg, Germany, 2009; pp. 3–51. [Google Scholar]
- Hassaan, Y.; Handoussa, H.; El-Khatib, A.H.; Linscheid, M.W.; El Sayed, N.; Ayoub, N. Evaluation of plant phenolic metabolites as a source of Alzheimer’s drug leads. Biomed. Res. Int. 2014, 2014, 843263. [Google Scholar] [CrossRef] [PubMed]
- Tanenbaum, D.M.; Wang, Y.; Williams, S.P.; Sigler, P.B. Crystallographic comparison of the estrogen and progesterone receptor’s ligand binding domains. Proc. Natl. Acad. Sci. USA 1998, 95, 5998–6003. [Google Scholar] [CrossRef] [PubMed]
- Brand, L.; Hörler, M.; Nüesch, E.; Vassalli, S.; Barrell, P.; Yang, W.; Jefferson, R.A.; Grossniklaus, U.; Curtis, M.D. A versatile and reliable two-component system for tissue-specific gene induction in Arabidopsis. Plant Physiol. 2006, 141, 1194–1204. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.C.; Wang, H.C.; Chen, G.Y.; Yang, J.C.; Korinek, M.; Hsieh, C.J.; Nozaki, H.; Hayashi, K.I.; Wu, C.C.; Wu, Y.C.; et al. Using the pER8: GUS reporter system to screen for phytoestrogens from Caesalpinia sappan. J. Nat. Prod. 2011, 74, 1698–1706. [Google Scholar] [CrossRef] [PubMed]
- Skehan, P.; Storeng, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New colorimetric cytotoxicity assay for anticancer-drug screening. J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef] [PubMed]
- Mense, S.M.; Hei, T.K.; Ganju, R.K.; Bhat, H.K. Phytoestrogens and breast cancer prevention: Possible mechanisms of action. Environ. Health Perspect. 2008, 116, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Miller, P.E.; Snyder, D.C. Phytochemicals and cancer risk: A review of the epidemiological evidence. Nutr. Clin. Pract. 2012, 27, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the compounds 2–5 are available from the authors.

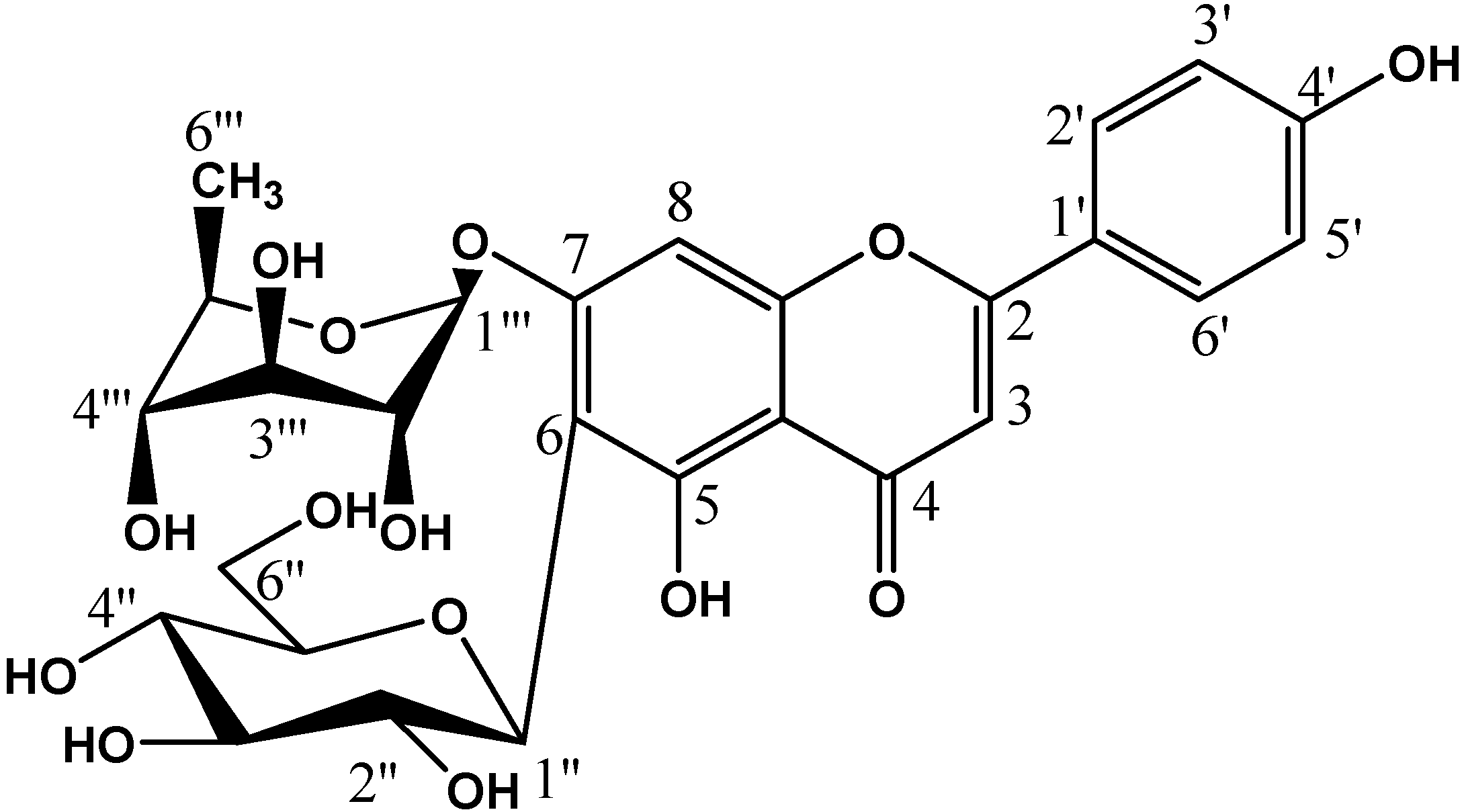
| Compound | R1 | R2 | R3 |
|---|---|---|---|
| Apigenin-7-O-rhamnosyl-6-C-glucoside (1) | H | glucose | rhamnose |
| Apigenin-6-C-glucoside (isovitexin) (2) | H | glucose | H |
| Luteolin-6-C-glucoside (Isoorientin) (3) | OH | glucose | H |
| Apigenin (4) | H | H | H |
| Luteolin (5) | OH | H | H |

 |  |  |  |  |  |  |
| Blank | 0.3125 nM | 0.625 nM | 1.25 nM | 2.5 nM | 5 nM | 10 nM |
| (a) | ||||||
| 6.25 µg/mL | 12.5 µg/mL | 25 µg/mL | 50 µg/mL | 100 µg/mL | 200 µg/mL | |
| Aqueous extract |  |  |  |  |  |  |
| Aqueous methanol extract |  |  |  |  |  |  |
| Fraction I |  |  |  |  |  |  |
| Fraction II |  |  |  |  |  |  |
| (b) | ||||||


{kind=link}
{kind=link}
{kind=link}
| Compound Name | Binding Energy (ΔGbinding) |
|---|---|
| Apigenin-7-O-rhamnosyl-6-C-glucoside | −44.7 |
| Apigenin-6-C-glucoside | −39.11 |
| Luteolin-6-C-glucoside | −41.91 |
| Apigenin | −43.33 |
| Luteolin | −40.97 |
| 17β-estradiol | −36.38 |
| Addition | IC50 (µg/mL) |
|---|---|
| Aqueous extract | 187.0 ± 11.2 |
| Aqueous methanol extract | >1000 |
| Fraction I | 23.3 ± 1.9 |
| Fraction II | 51.5 ± 3.5 |
| Drug | Viability % | |||
|---|---|---|---|---|
| Day 1 | Day 3 | Day 7 | Day 10 | |
| Control | 100.00 ± 9.2 | 200.00± 22.92 | 210.97 ± 22.36 | 210.97 ± 25.36 |
| Aq.alc. extract (10 µg/mL) | 88.37 ± 9.63 | 204.87 ± 24.14 | 234.14 ± 29.02 | 234.14 ± 29.02 |
| Fraction I (10 µg/mL) | 91.46 ± 8.41 | 198.78 ± 18.7 | 290.24 ± 27.22 * | 290.24 ± 21.22 * |
| (1 µg/mL) | 86.58 ± 7.19 | 193.90 ± 20.26 | 261.95 ± 32.12 | 279.83 ± 26.34 * |
| Fraction II (10 µg/mL) | 93.41 ± 5.97 | 198.78 ± 21.66 | 290.24 ± 31.22 * | 290.24 ± 21.22 * |
| (1 µg/mL) | 92.68 ± 10.85 | 203.66 ± 24.14 | 262.73 ± 30.12 | 276.83 ± 26.34 * |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashmawy, N.S.; Ashour, M.L.; Wink, M.; El-Shazly, M.; Chang, F.-R.; Swilam, N.; Abdel-Naim, A.B.; Ayoub, N. Polyphenols from Erythrina crista-galli: Structures, Molecular Docking and Phytoestrogenic Activity. Molecules 2016, 21, 726. https://doi.org/10.3390/molecules21060726
Ashmawy NS, Ashour ML, Wink M, El-Shazly M, Chang F-R, Swilam N, Abdel-Naim AB, Ayoub N. Polyphenols from Erythrina crista-galli: Structures, Molecular Docking and Phytoestrogenic Activity. Molecules. 2016; 21(6):726. https://doi.org/10.3390/molecules21060726
Chicago/Turabian StyleAshmawy, Naglaa S., Mohamed L. Ashour, Michael Wink, Mohamed El-Shazly, Fang-Rong Chang, Noha Swilam, Ashraf B. Abdel-Naim, and Nahla Ayoub. 2016. "Polyphenols from Erythrina crista-galli: Structures, Molecular Docking and Phytoestrogenic Activity" Molecules 21, no. 6: 726. https://doi.org/10.3390/molecules21060726