
Antibacterial Peptide CecropinB2 Production via Various Host and Construct Systems


Abstract
:1. Introduction
2. Results and Discussion
2.1. CecropinB2 Production in E. coli


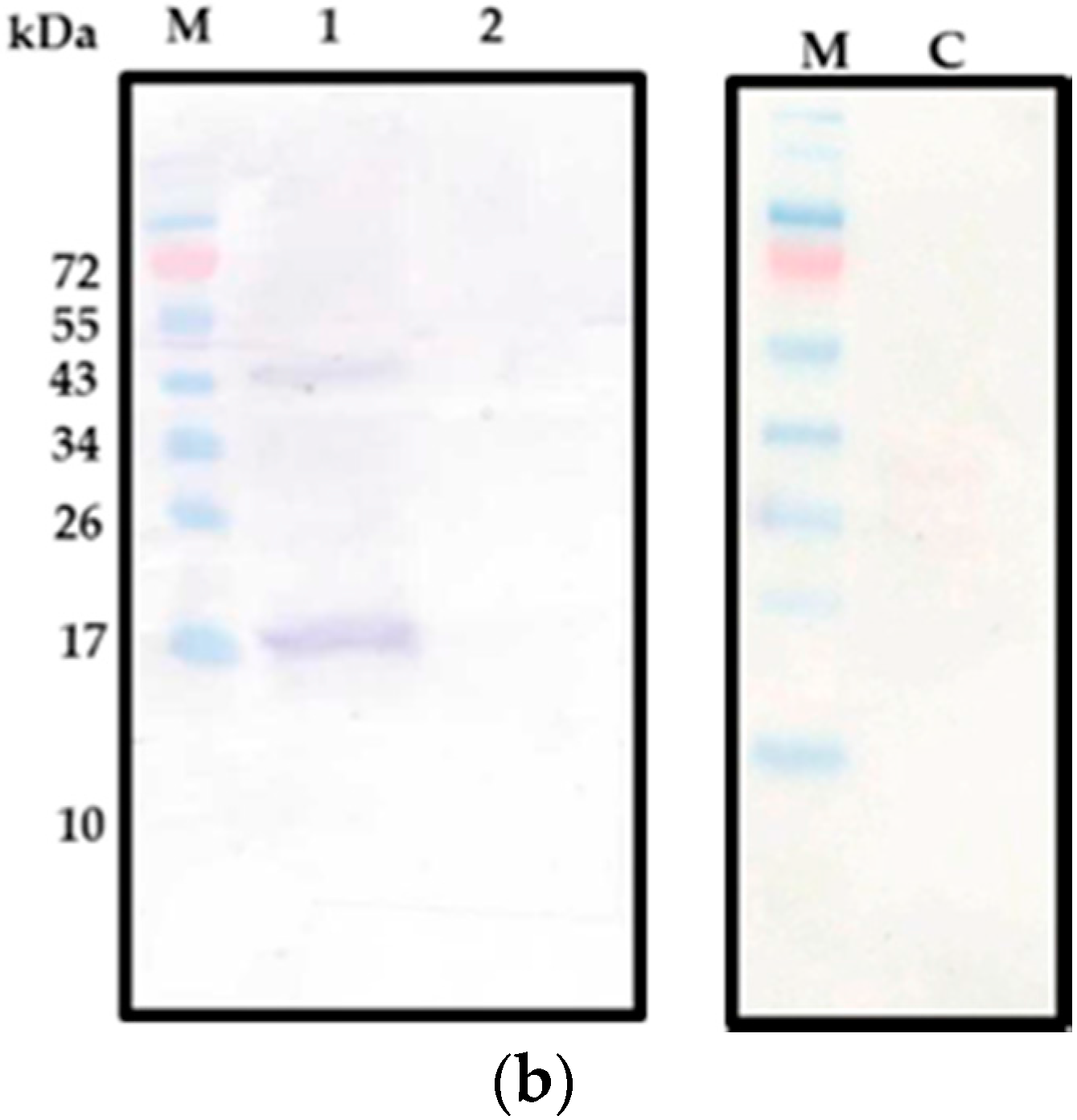
2.2. CecropinB2 Production in B. subtilis
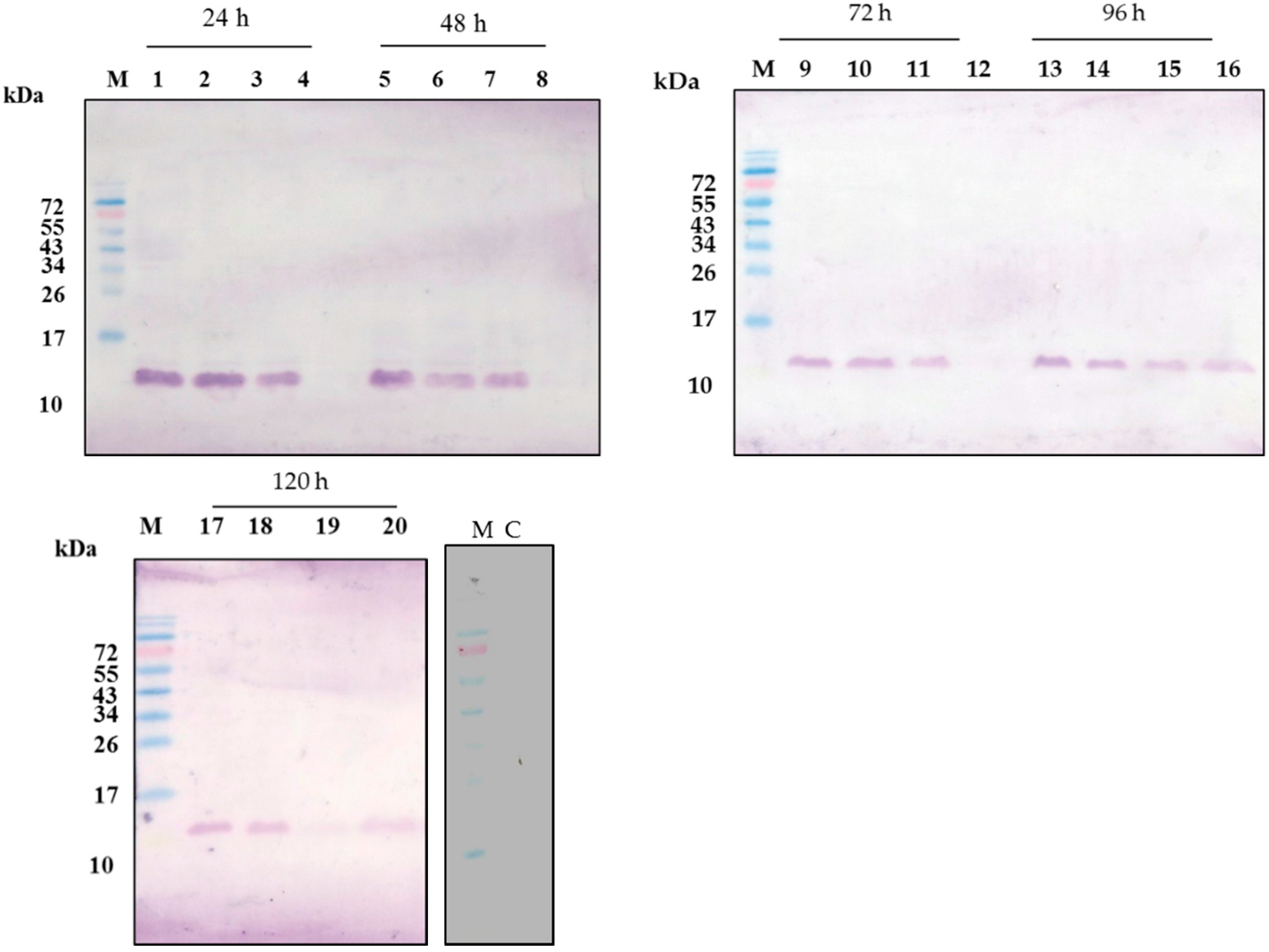
2.3. CecropinB2 Production in P. pastoris

2.4. Comparison of the Host Systems

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Construct | Total Protein Content (mg/L) 1 | Purity (%) 2 | Yield of CecropinB2 3 |
|---|---|---|---|
| (a) E. coli | |||
| pET26b-cecB2/E. coli ER2566 | - | - | - |
| pET-28a-cecB2/E. coli ER2566 | - | - | - |
| pET28a-INT-cecB2/E. coli ER2566 | - | - | - |
| pET26b-INT-cecB2/E. coli ER2566 | 1022 | 0.8 | + |
| pET26b-10K-INT-cecB2/E. coli ER2566 | 987 | 0.8 | + |
| pET26b-10R-INT-cecB2/E. coli ER2566 | 1055 | 0.8 | + |
| (b) B. subtilis | |||
| PRPA-cecB2/B. subtilis WB800 | 1128 | 1.8 | + |
| PRPA-INT-cecB2/B. subtilis WB800 | - | - | - |
| (c) P. pastoris | |||
| pGAPZαC-cecB2/P. pastoris SMD1168 | 1980 | 2.4 | ++ |
| pGAPZαC-INT-cecB/P. pastoris SMD1168 | - | - | - |
| pGAPZαC-prepro-cecB2/P. pastoris SMD1168 | 2048 | 4.3 | +++ |
2.5. Antibacterial Tests

| Bacteria Strain | Antibacterial Activity 2 | |
|---|---|---|
| Purified CecropinB2 (10 μg/mL) | Synthetic CecropinB2 (4 μg/mL) | |
| E. coli ER2566 | 6.5 | 6.8 |
| E. coli BL21 | 6.8 | 7.2 |
| E. coli Rosetta | 7.1 | 7.2 |
| E. coli JM109 | 6.5 | 6.6 |
| E. coli DH1 | 6.5 | 6.7 |
| B. subtilis WB800 | - | - |
| P. pastoris SMD1168 | - | - |
| A. baumannii BCRC 15884 | 7.2 | 6.8 |
| A. baumannii E1359 | 7.3 | 6.9 |
3. Materials and Methods
3.1. Bacterial Strains, Plasmids and Synthetic CecropinB2
| Protein | Number of Amino Acid | Theoretical Molecular Mass (Da) |
|---|---|---|
| CecropinB2 | 36 | 3749 |
| INT-cecB2 | 193 | 17,569 |
| signal peptide (SP) | 18 | 2085 |
| Propeptide | 5 | 468 |
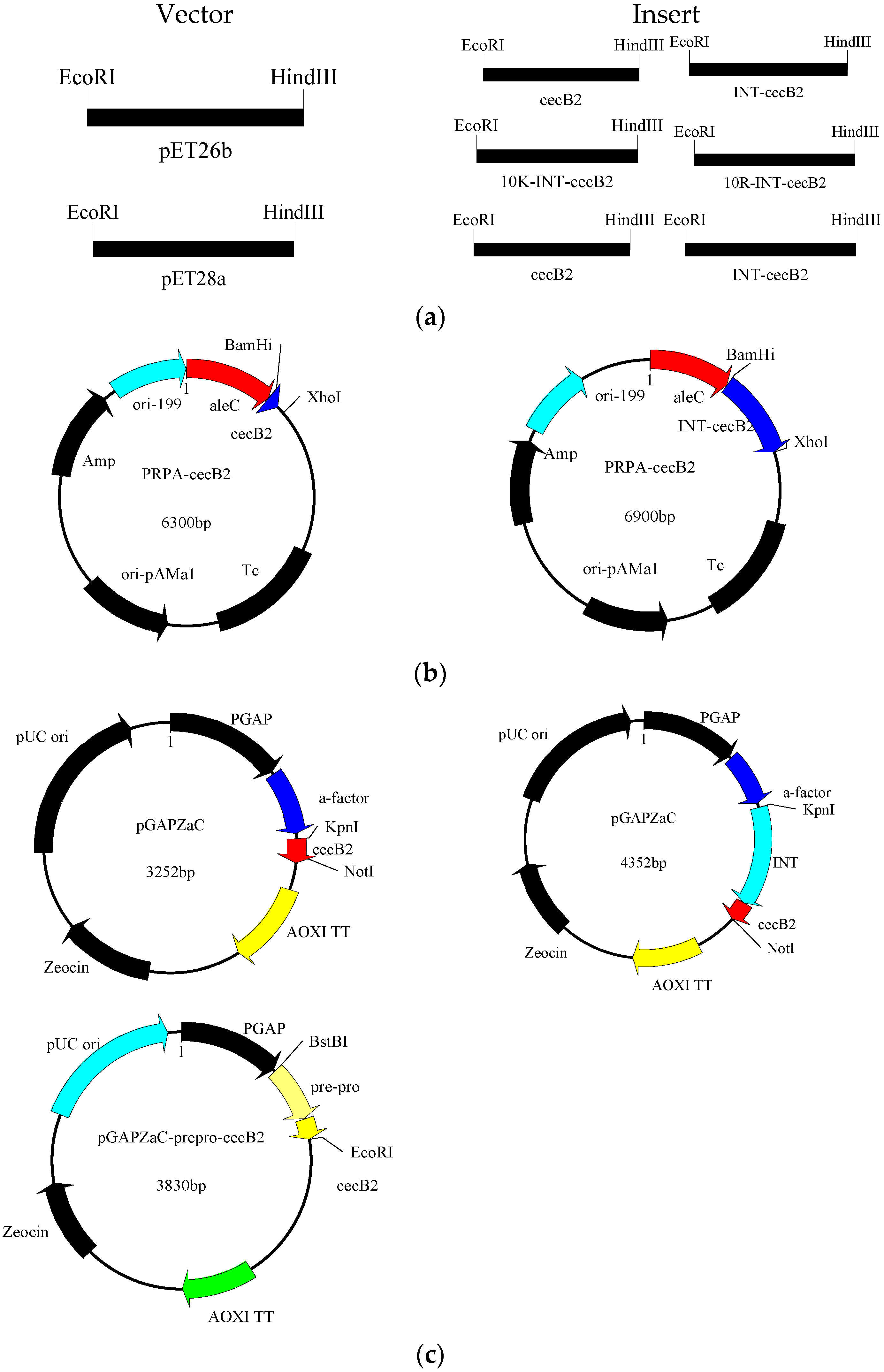
3.2. Construction of Expression Systems

3.3. Production of the Antibacterial Peptide
3.3.1. Culture of E. coli
3.3.2. Culture of B. subtilis
3.3.3. Culture of P. pastoris
3.4. Purification of Target Protein
3.5. Analytical Methods
3.6. Antibacterial Assay
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fleming, A. In-vitro test of pencillin potency. Lancet 1942, 239, 732–733. [Google Scholar] [CrossRef]
- Nikaido, H. Multidrug Resistance in Bacteria. Annu. Rev. Biochem. 2009, 78, 119–146. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.M.; Jan, P.S.; Yu, H.C.; Haung, H.Y.; Fang, H.J.; Chang, Y.I.; Cheng, J.W.; Chen, H.M. Structure and function of a custom anticancer peptide, CB1a. Peptides 2009, 30, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Hoskin, D.W.; Ramamoorthy, A. Studies on anticancer activities of antimicrobial peptides. Biochim. Biophys. Acta 2008, 1778, 357–375. [Google Scholar] [CrossRef] [PubMed]
- Bahar, A.; Ren, D. Antimicrobial Peptides. Pharmaceuticals 2013, 6, 1543. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Wang, W.; Smith, D.; Chan, S.C. Effects of the anti-bacterial peptide cecropin B and its analogs, cecropins B-1 and B-2, on liposomes, bacteria, and cancer cells. Biochim. Biophys. Acta Gen. Subj. 1997, 1336, 171–179. [Google Scholar] [CrossRef]
- Steiner, H.; Hultmark, D.; Engstrom, A.; Bennich, H.; Boman, H.G. Sequence and specificity of two antibacterial proteins involved in insect immunity. Nature 1981, 292, 246–248. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, F. Cationic amphiphilic peptides with cancer-selective toxicity. Eur. J. Pharmacol. 2009, 625, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Jin, F.; Yu, X.; Ji, S.; Wang, J.; Cheng, H.; Wang, C.; Zhang, W. Expression and purification of a recombinant antibacterial peptide, cecropin, from Escherichia coli. Protein Expr. Purif. 2007, 53, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhong, Z.; Huang, L.; Peng, L.; Wang, F.; Cen, P. High-level production of bioactive human beta-defensin-4 in Escherichia coli by soluble fusion expression. Appl. Microbiol. Biotechnol. 2006, 72, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Schumann, W. Production of Recombinant Proteins in Bacillus subtilis. Adv. Appl. Microbiol. 2007, 62, 137–189. [Google Scholar] [PubMed]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef] [PubMed]
- Cregg, J.; Cereghino, J.; Shi, J.; Higgins, D. Recombinant protein expression in Pichia pastoris. Mol. Biotechnol. 2000, 16, 23–52. [Google Scholar] [CrossRef]
- Skosyrev, V.S.; Rudenko, N.V.; Yakhnin, A.V.; Zagranichny, V.E.; Popova, L.I.; Zakharov, M.V.; Gorokhovatsky, A.Y.; Vinokurov, L.M. EGFP as a fusion partner for the expression and organic extraction of small polypeptides. Protein Expr. Purif. 2003, 27, 55–62. [Google Scholar] [CrossRef]
- Zhang, J.; Movahedi, A.; Xu, J.; Wang, M.; Wu, X.; Xu, C.; Yin, T.; Zhuge, Q. In vitro production and antifungal activity of peptide ABP-dHC-cecropin A. J. Biotechnol. 2015, 199, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Su, Y.; Li, J.; Sun, J.; Yang, Y. Expression and purification of the antimicrobial peptide cecropin AD by fusion with cationic elastin-like polypeptides. Protein Expr. Purif. 2012, 85, 200–203. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.-J.; Kim, S.-K.; Min, W.-K.; Lee, S.-S.; Park, K.; Park, Y.-C.; Seo, J.-H. Polycationic amino acid tags enhance soluble expression of Candida antarctica lipase B in recombinant Escherichia coli. Bioprocess Biosyst. Eng. 2011, 34, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.-M.; Wang, J.-P.; Su, F.-S. Extracellular Production of a Novel Ice Structuring Protein byBacillus subtilis—A Case of Recombinant Food Peptide Additive Production. Food Biotechnol. 2007, 21, 119–128. [Google Scholar] [CrossRef]
- Sambrook, J.R. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Meth. 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Ekengren, S.; Hultmark, D. Drosophila cecropin as an antifungal agent. Insect Biochem. Mol. Biol. 1999, 29, 965–972. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, W.-S.; Kan, S.-C.; Lin, C.-C.; Shieh, C.-J.; Liu, Y.-C. Antibacterial Peptide CecropinB2 Production via Various Host and Construct Systems. Molecules 2016, 21, 103. https://doi.org/10.3390/molecules21010103
Lai W-S, Kan S-C, Lin C-C, Shieh C-J, Liu Y-C. Antibacterial Peptide CecropinB2 Production via Various Host and Construct Systems. Molecules. 2016; 21(1):103. https://doi.org/10.3390/molecules21010103
Chicago/Turabian StyleLai, Wei-Shiang, Shu-Chen Kan, Chia-Chi Lin, Chwen-Jen Shieh, and Yung-Chuan Liu. 2016. "Antibacterial Peptide CecropinB2 Production via Various Host and Construct Systems" Molecules 21, no. 1: 103. https://doi.org/10.3390/molecules21010103