
Fatty Acid Profile and Biological Activities of Linseed and Rapeseed Oils


Abstract
:1. Introduction
2. Results and Discussion

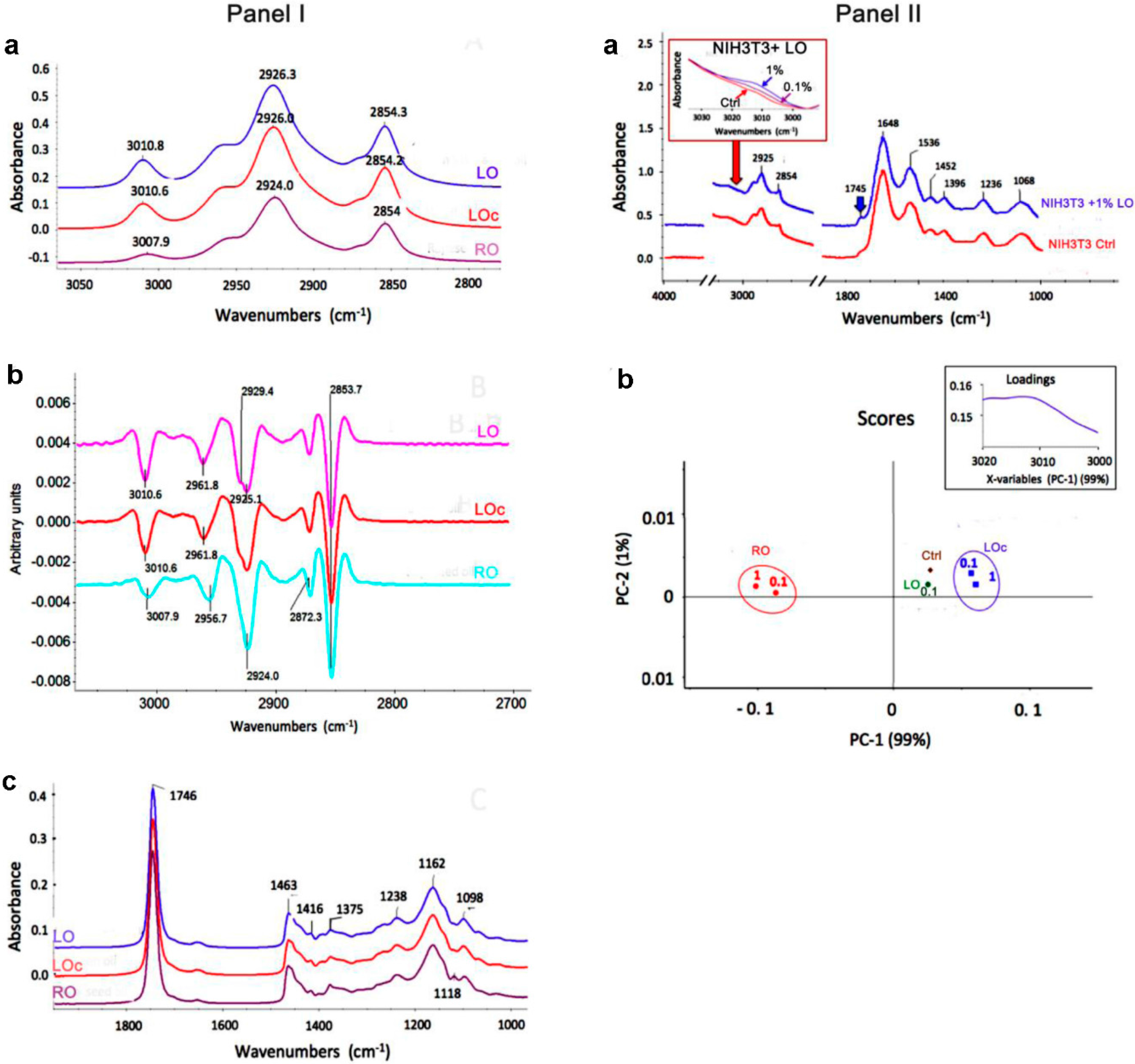{kind=link}
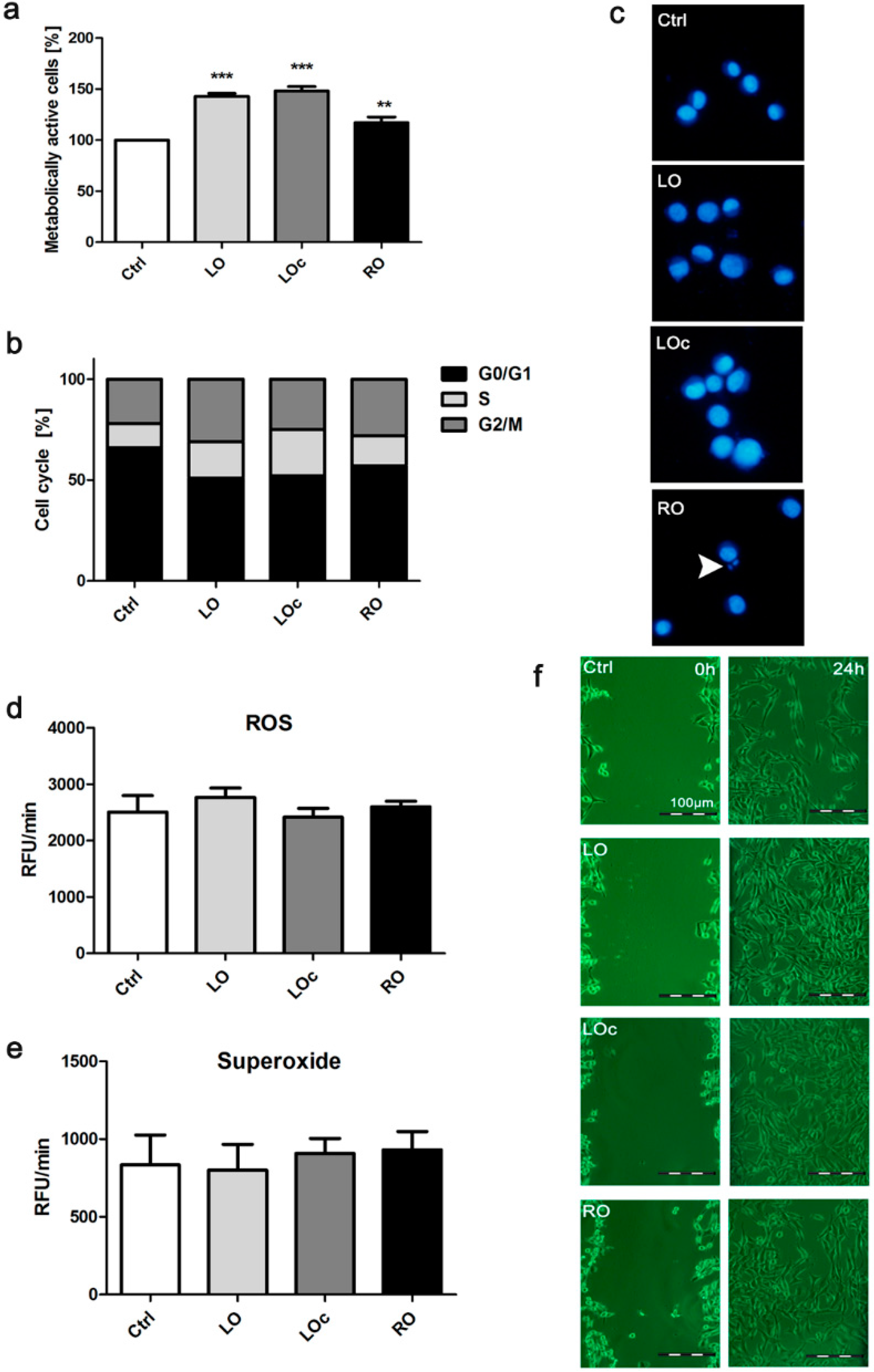{kind=link}
| Oils | MUFA [%] | PUFA [%] | SFA [%] | Unsaturated FA/Saturated FA | MUFA/PUFA |
|---|---|---|---|---|---|
| LO | 15.8 | 74.8 | 9.4 | 9.64 | 0.21 |
| LOc | 20.3 | 68.5 | 11 | 8.07 | 0.30 |
| RO | 62.9 | 29.6 | 7.5 | 12.33 | 2.10 |
| Common Name, C:D a | Oils | ||
|---|---|---|---|
| LO | LOc | RO | |
| Myristic Acid, 14:0 | <0.1 | 0.1 | ND b |
| Palmitic Acid, 16:0 | 5.1 | 5.3 | 4.3 |
| Stearic Acid, 18:0 | 4.3 | 5.1 | 2.0 |
| Arachidic Acid, 20:0 | <0.1 | 0.3 | 0.8 |
| Behenic Acid, 22:0 | ND b | 0.2 | 0.4 |
| Oleic acid, 18:1, n-9 | 15.8 | 20.0 | 62.5 |
| Linoleic Acid, 18:2, n-6 | 16.5 | 17.8 | 19.6 |
| α-Linolenic Acid, 18:3, n-3 | 58.3 | 50.7 | 10.0 |
| Erucic Acid, 22:1, n-9 | ND b | 0.3 | 0.4 |


3. Materials and Methods
3.1 Fatty Acid Profiles
3.2 Cell Culture
3.3 Cytotoxicity, Cell Cycle and Genotoxicity
3.4 Oxidative Stress
3.5 Scratch Wound Healing Assay
3.6 Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Basch, E.; Bent, S.; Collins, J.; Dacey, C.; Hammerness, P.; Harrison, M.; Smith, M.; Szapary, P.; Ulbricht, C.; Vora, M.; et al. Flax and flaxseed oil (Linum usitatissimum): A review by the natural standard research collaboration. J. Soc. Integr. Oncol. 2007, 5, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Allemekinders, H.; Dansby, A.; Campbell, L.; Durance-Tod, S.; Berger, A.; Jones, P.J. Evidence of health benefits of canola oil. Nutr. Rev. 2013, 71, 370–385. [Google Scholar] [CrossRef] [PubMed]
- Dupont, J.; White, P.J.; Johnston, K.M.; Heggtveit, H.A.; McDonald, B.E.; Grundy, S.M.; Bonanome, A. Food safety and health effects of canola oil. J. Am. Coll. Nutr. 1989, 8, 360–375. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.H.; Keast, D.R.; Kris-Etherton, P.M. Dietary modeling shows that the substitution of canola oil for fats commonly used in the united states would increase compliance with dietary recommendations for fatty acids. J. Am. Diet. Assoc. 2007, 107, 1726–1734. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, R.; Gerber, M. Can rapeseed oil replace olive oil as part of a mediterranean-style diet? Br. J. Nutr. 2014, 112, 1882–1895. [Google Scholar] [CrossRef] [PubMed]
- Ruthig, D.J.; Meckling-Gill, K.A. Both (n-3) and (n-6) fatty acids stimulate wound healing in the rat intestinal epithelial cell line, IEC-6. J. Nutr. 1999, 129, 1791–1798. [Google Scholar] [PubMed]
- Hankenson, K.D.; Watkins, B.A.; Schoenlein, I.A.; Allen, K.G.; Turek, J.J. Omega-3 fatty acids enhance ligament fibroblast collagen formation in association with changes in interleukin-6 production. Proc. Soc. Exp. Biol. Med. 2000, 223, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, R.; Webster, F.; Kiemle, D. Spectrometric Identification of Organic Compounds; John Wiley & Sons, Inc.: New York, NY, USA, 2005. [Google Scholar]
- Yoshida, S.; Yoshida, H. Noninvasive analyses of polyunsaturated fatty acids in human oral mucosa in vivo by fourier-transform infrared spectroscopy. Biopolymers 2004, 74, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Nykter, M.; Kymalainen, H.; Gates, F.; Sjoberg, A. Quality characteristics of edible linseed oil. Agric. Food Sci. 2006, 15, 402–413. [Google Scholar] [CrossRef]
- Ramos, M.J.; Fernandez, C.M.; Casas, A.; Rodriguez, L.; Perez, A. Influence of fatty acid composition of raw materials on biodiesel properties. Bioresour. Technol. 2009, 100, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Stuart, B. Infrared spectroscopy. In Kirk-Othmer Encyclopedia of Chemical Technology; Seidel, A., Ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2005; Volume 14, pp. 224–243. [Google Scholar]
- Russo, G.L. Dietary n-6 and n-3 polyunsaturated fatty acids: From biochemistry to clinical implications in cardiovascular prevention. Biochem. Pharmacol. 2009, 77, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. N-3 polyunsaturated fatty acids and inflammation: From molecular biology to the clinic. Lipids 2003, 38, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Fetterman, J.W., Jr.; Zdanowicz, M.M. Therapeutic potential of n-3 polyunsaturated fatty acids in disease. Am. J. Health Syst. Pharm. 2009, 66, 1169–1179. [Google Scholar] [CrossRef] [PubMed]
- Otranto, M.; Do Nascimento, A.P.; Monte-Alto-Costa, A. Effects of supplementation with different edible oils on cutaneous wound healing. Wound Repair Regen. 2010, 18, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, C.R.; Souza, M.A.; Ferro, E.A.; Favoreto, S., Jr.; Pena, J.D. Influence of topical administration of n-3 and n-6 essential and n-9 nonessential fatty acids on the healing of cutaneous wounds. Wound Repair Regen. 2004, 12, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Turek, J.J. Inducible nitric oxide synthase links NF-kappaB to PGE2 in polyunsaturated fatty acid altered fibroblast in vitro wound healing. Lipids Health Dis. 2005, 4, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.P.; Most, D.; Efron, D.T.; Tantry, U.; Fischel, M.H.; Barbul, A. The role of iNOS in wound healing. Surgery 2001, 130, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Turek, J.J. Altered NF-kappaB gene expression and collagen formation induced by polyunsaturated fatty acids. J. Nutr. Biochem. 2005, 16, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Altavilla, D.; Saitta, A.; Cucinotta, D.; Galeano, M.; Deodato, B.; Colonna, M.; Torre, V.; Russo, G.; Sardella, A.; Urna, G.; et al. Inhibition of lipid peroxidation restores impaired vascular endothelial growth factor expression and stimulates wound healing and angiogenesis in the genetically diabetic mouse. Diabetes 2001, 50, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Sola, R.; la Ville, A.E.; Richard, J.L.; Motta, C.; Bargallo, M.T.; Girona, J.; Masana, L.; Jacotot, B. Oleic acid rich diet protects against the oxidative modification of high density lipoprotein. Free Radic. Biol. Med. 1997, 22, 1037–1045. [Google Scholar] [CrossRef]
- Siwak, J.; Lewinska, A.; Wnuk, M.; Bartosz, G. Protection of flavonoids against hypochlorite-induced protein modifications. Food Chem. 2013, 141, 1227–1241. [Google Scholar] [CrossRef] [PubMed]
- Lewinska, A.; Adamczyk, J.; Pajak, J.; Stoklosa, S.; Kubis, B.; Pastuszek, P.; Slota, E.; Wnuk, M. Curcumin-mediated decrease in the expression of nucleolar organizer regions in cervical cancer (HeLa) cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2014, 771, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Lewinska, A.; Jarosz, P.; Czech, J.; Rzeszutek, I.; Bielak-Zmijewska, A.; Grabowska, W.; Wnuk, M. Capsaicin-induced genotoxic stress does not promote apoptosis in A549 human lung and DU145 prostate cancer cells. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2015, 779, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: The oil samples are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lewinska, A.; Zebrowski, J.; Duda, M.; Gorka, A.; Wnuk, M. Fatty Acid Profile and Biological Activities of Linseed and Rapeseed Oils. Molecules 2015, 20, 22872-22880. https://doi.org/10.3390/molecules201219887
Lewinska A, Zebrowski J, Duda M, Gorka A, Wnuk M. Fatty Acid Profile and Biological Activities of Linseed and Rapeseed Oils. Molecules. 2015; 20(12):22872-22880. https://doi.org/10.3390/molecules201219887
Chicago/Turabian StyleLewinska, Anna, Jacek Zebrowski, Magdalena Duda, Anna Gorka, and Maciej Wnuk. 2015. "Fatty Acid Profile and Biological Activities of Linseed and Rapeseed Oils" Molecules 20, no. 12: 22872-22880. https://doi.org/10.3390/molecules201219887