Identification of Dehydroxytrichostatin A as a Novel Up-Regulator of the ATP-Binding Cassette Transporter A1 (ABCA1)

Abstract
:1. Introduction
2. Results and Discussion
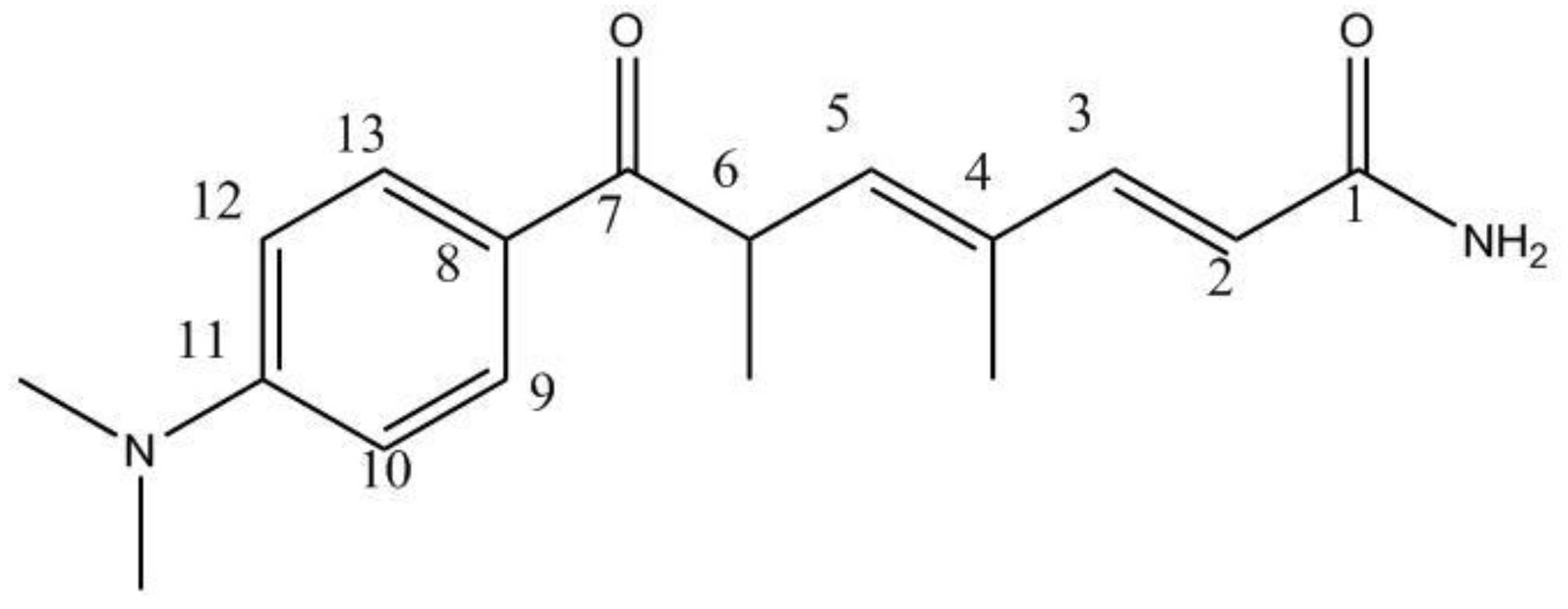
2.1. Isolation and Identification of 9179B from Strain 04-9179

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | 13C-NMR a | 1H-NMR b |
|---|---|---|
| 1 | n.d. c | |
| 2 | 120.1 | 5.97 (1H, d, J = 15.5) |
| 3 | 146.9 | 7.10 (1H, d, J = 15.5) |
| 4 | 134.3 | |
| 5 | 141.1 | 5.86 (1H, d, J = 9.5) |
| 6 | 41.6 | 4.48 (1H, m) |
| 7 | 201.4 | |
| 8 | 124.6 | |
| 9,13 | 131.8 | 7.81 (2H, d, J = 9.0) |
| 10,12 | 111.9 | 6.67 (2H,d, J = 9.0) |
| 11 | 155.4 | |
| 4-Me | 12.7 | 1.80 (3H, s) |
| 6-Me | 18.2 | 1.22 (3H, d, J = 6.5) |
| N-Me2 | 40.2 | 3.02 (6H, s) |

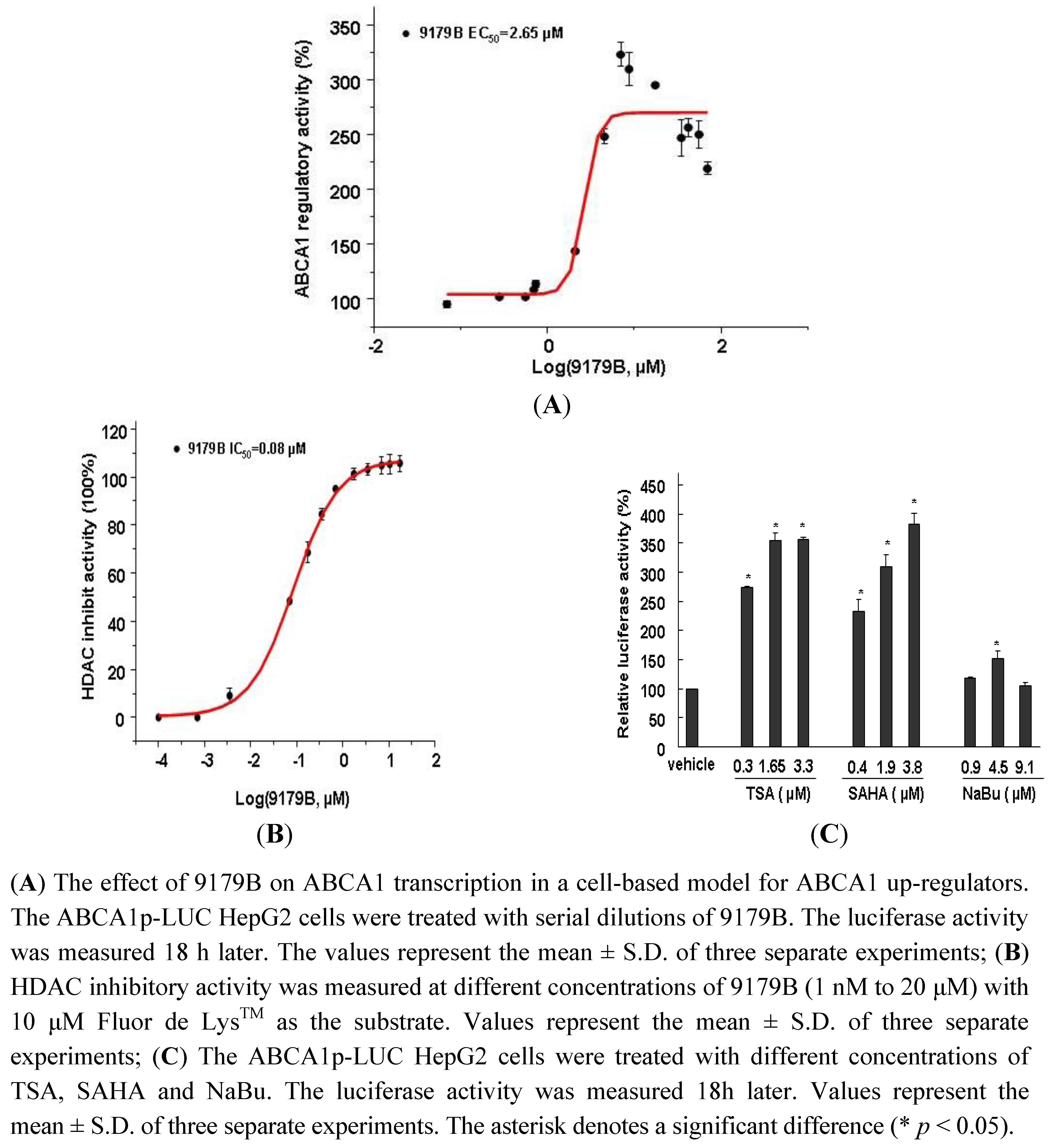
2.2. Effect of 9179B and Other HDAC Inhibitors on ABCA1 Promoter Activity

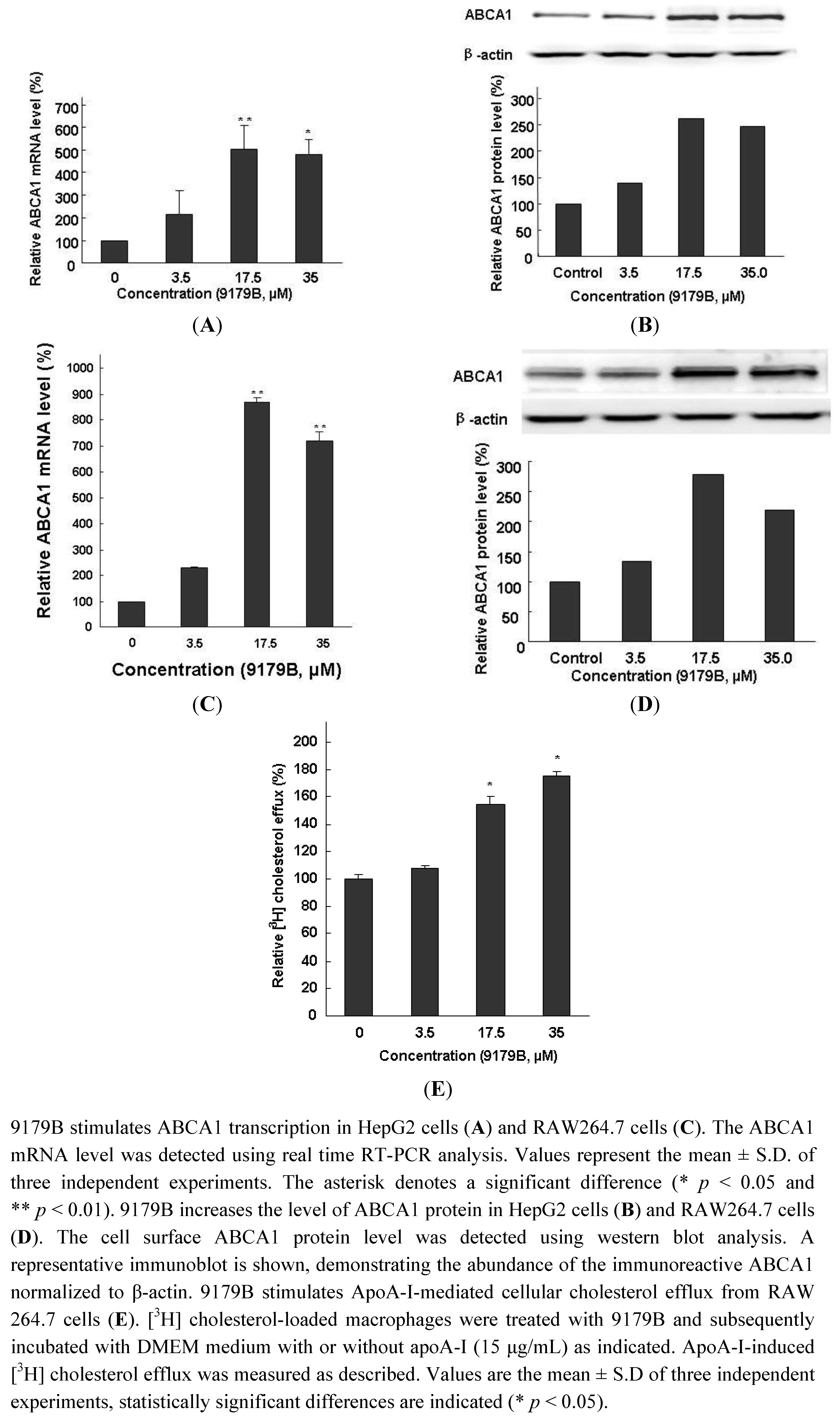
2.3. The Effects of 9179B on ABCA1 Expression in HepG2 and RAW 264.7 Cells and on Cholesterol Efflux in RAW 264.7 Cells

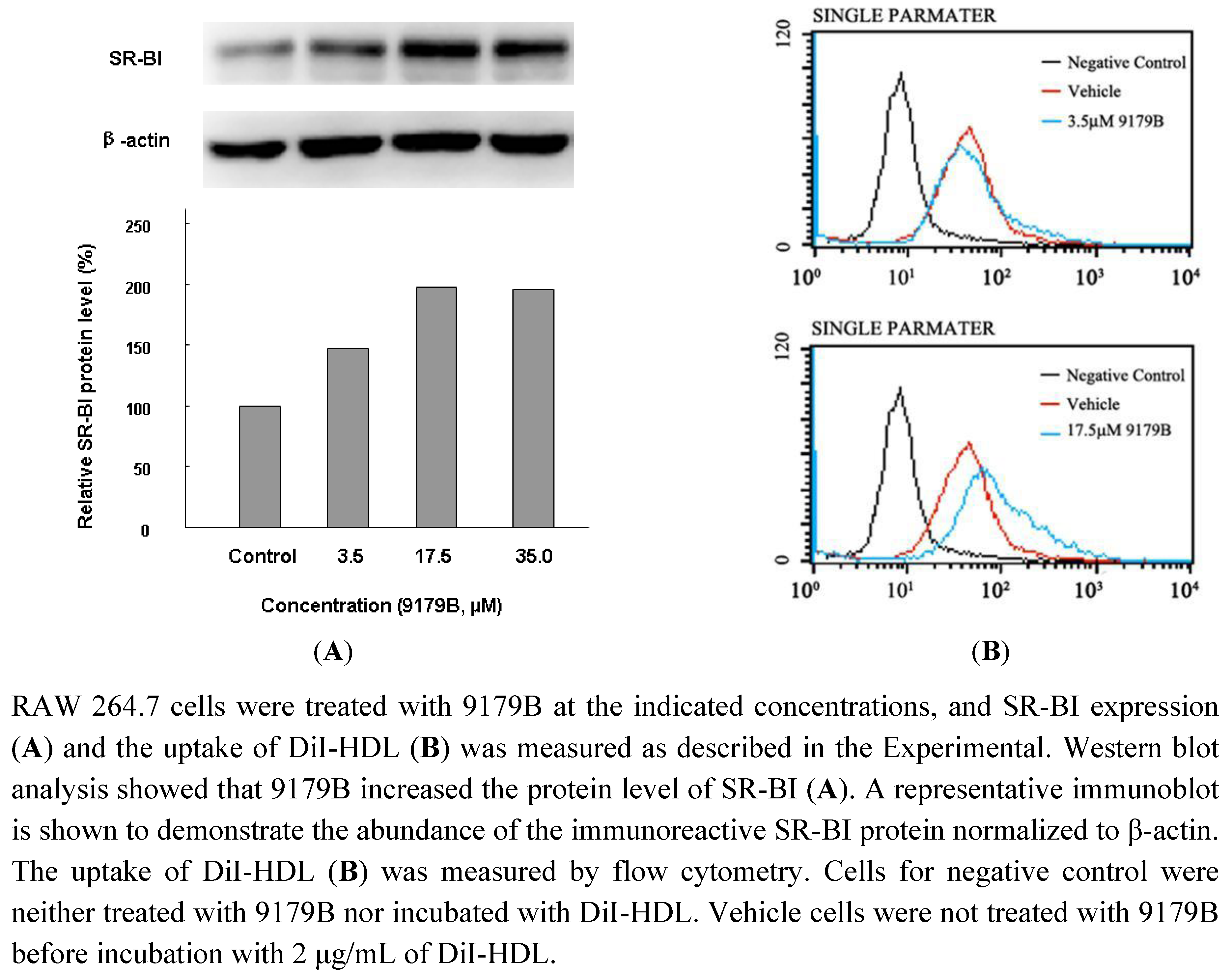
2.4. The Influence of 9179B on SR-BI and the Uptake of DiI-HDL in RAW 264.7 Cells

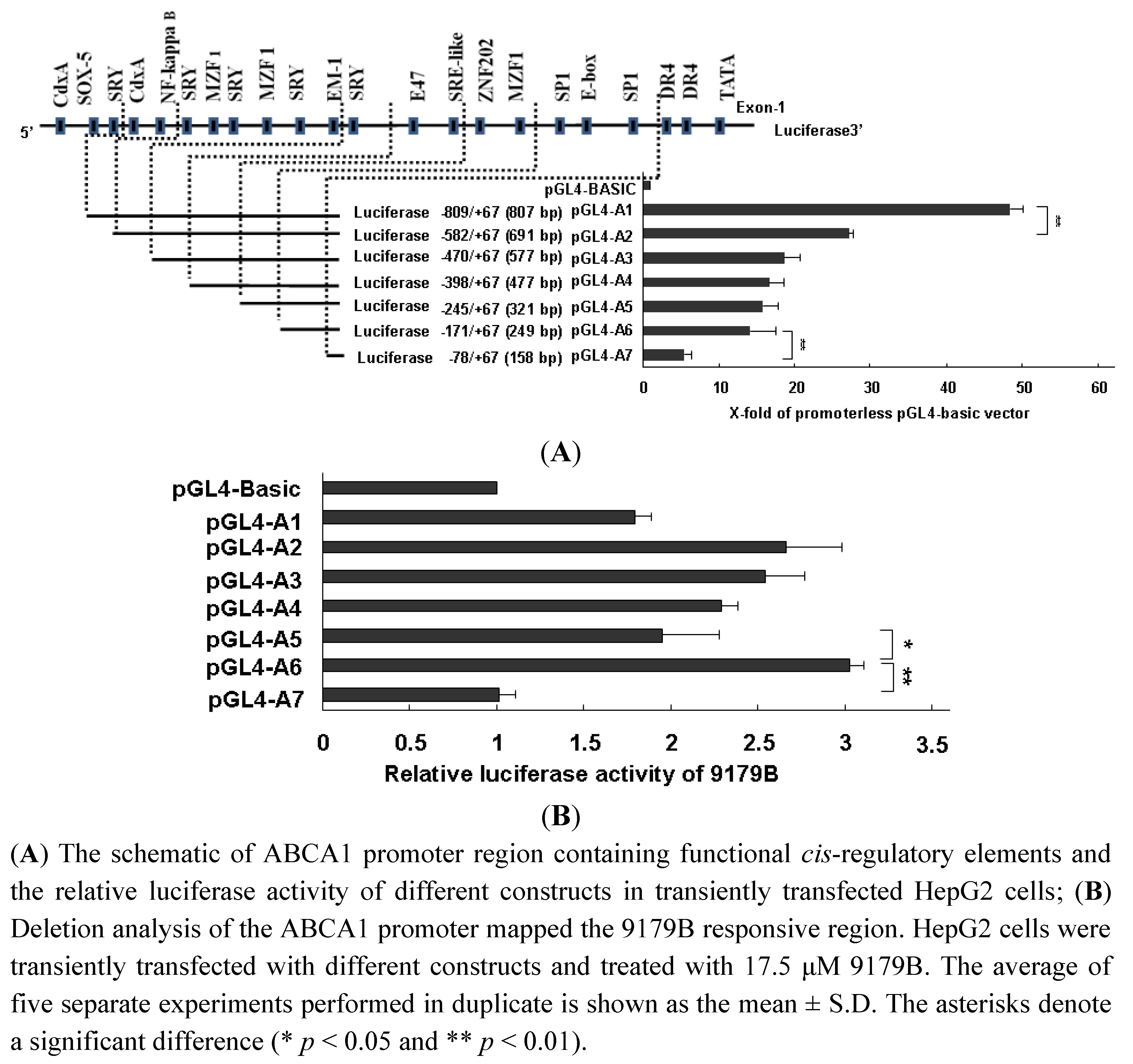
2.5. The Influence of 9179B on Different Cis-Regulatory Elements of ABCA1

3. Experimental
3.1. General
3.2. Cell Culture
3.3. The Isolation, Purification and Structure Elucidation of 9179B
3.4. ABCA1 Promoter Activity Assay
3.5. Enzyme Inhibitor Activity Assay and Other HDAC Inhibitor on ABCA1 Promoter Activity
3.6. The Effects of 9179B on ABCA1 Expression in HepG2 or RAW264.7 Cells
3.7. The Effects of 9179B on Cholesterol Efflux in RAW 264.7 Cells
3.8. The Effect of 9179B on SR-BI Expression in RAW264.7 Cells and the Analysis of Cellular Uptake of DiI-Labeled HDL by Flow Cytometry
3.9. Luciferase Reporter Assay
3.10. Statistical Analysis
4. Conclusions
Acknowledgments
- Sample Availability: Samples of the compounds are available from the authors and commercially available.
References
- Ross, R. Atherosclerosis-An inflammatory disease. N. Engl. J. Med. 1999, 340, 115–126. [Google Scholar] [CrossRef]
- Stein, O.; Stein, Y. Atheroprotective mechanisms of HDL. Atherosclerosis 1999, 144, 285–301. [Google Scholar] [CrossRef]
- Gordon, D.J.; Probstfield, J.L.; Garrison, R.J.; Neaton, J.D.; Castelli, W.P.; Knoke, J.D.; Jacobs, D.R., Jr.; Bangdiwala, S.; Tyroler, H.A. High-density lipoprotein cholesterol and cardiovascular disease. Four prospective American studies. Circulation 1989, 79, 8–15. [Google Scholar] [CrossRef]
- Goldbourt, U.; Yaari, J.S.; Medalie, J.H. Isolated low HDL cholesterol as a risk factor for coronary heart disease mortality: A 21-year follow-up of 8,000 men. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 107–113. [Google Scholar] [CrossRef]
- Langmann, T.; Klucken, J.; Reil, M.; Liebisch, G.; Luciani, M.F.; Chimini, G.; Kaminski, W.E.; Schmitz, G. Molecular cloning of the human ATP-binding cassette transporter 1 (hABC1): Evidence for sterol-dependent regulation in macrophages. Biochem. Biophys. Res. Commun. 1999, 257, 29–33. [Google Scholar] [CrossRef]
- Lee, J.Y.; Parks, J.S. ATP-binding cassette transporter AI and its role in HDL formation. Curr. Opin. Lipidol. 2005, 16, 19–25. [Google Scholar] [CrossRef]
- Oram, J.F.; Vaughan, A.M. ABCA1-mediated transport of cellular cholesterol and phospholipids to HDL apolipoproteins. Curr. Opin. Lipidol. 2000, 11, 253–260. [Google Scholar] [CrossRef]
- Zannis, V.I.; Chroni, A.; Krieger, M. Role of apoA-I, ABCA1, LCAT, and SR-BI in the biogenesis of HDL. J. Mol. Med. 2006, 84, 276–294. [Google Scholar] [CrossRef]
- Zhao, C.; Dahlman-Wright, K. Liver X receptor in cholesterol metabolism. J. Endocrinol. 2010, 204, 233–240. [Google Scholar] [CrossRef]
- Brooks-Wilson, A.; Marcil, M.; Clee, S.M.; Zhang, L.H.; Roomp, K.; van Dam, M.; Yu, L.; Brewer, C.; Collins, J.A.; Molhuizen, H.O.; et al. Mutations in ABC1 in Tangier disease and familial highdensity lipoprotein deficiency. Nat. Genet. 1999, 22, 336–345. [Google Scholar] [CrossRef]
- Bodzioch, M.; Orsó, E.; Klucken, J.; Langmann, T.; Böttcher, A.; Diederich, W.; Drobnik, W.; Barlage, S.; Büchler, C.; Porsch-Ozcürümez, M.; et al. The gene encoding ATP-binding cassette transporter 1 is mutated in Tangier disease. Nat. Genet. 1999, 22, 347–351. [Google Scholar] [CrossRef]
- Lu, L.; Liu, H.; Peng, J.; Gan, L.; Shen, L.; Zhang, Q.; Li, L.; Zhang, L.; Su, C.; Jiang, Y. Regulations of the key mediators in inflammation and atherosclerosis by aspirin in human macrophages. Lipids Health Dis. 2010, 9, 16. [Google Scholar] [CrossRef]
- Singaraja, R.R.; Fievet, C.; Castro, G.; James, E.R.; Hennuyer, N.; Clee, S.M.; Bissada, N.; Choy, J.C.; Fruchart, J.C.; McManus, B.M.; et al. Increased ABCA1 activity protects against atherosclerosis. J. Clin. Invest. 2002, 110, 35–42. [Google Scholar]
- Joyce, C.W.; Amar, M.J.; Lambert, G.; Vaisman, B.L.; Paigen, B.; Najib-Fruchart, J.; Hoyt, R.F., Jr.; Neufeld, E.D.; Remaley, A.T.; Fredrickson, D.S.; et al. The ATP binding cassette transporter A1 (ABCA1) modulates the development of aortic atherosclerosis in C57BL/6 and apoE-knockout mice. Proc. Natl. Acad. Sci. USA 2002, 99, 407–412. [Google Scholar]
- Timmins, J.M.; Lee, J.Y.; Boudyguina, E.; Kluckman, K.D.; Brunham, L.R.; Mulya, A.; Gebre, A.K.; Coutinho, J.M.; Colvin, P.L.; Smith, T.L.; et al. Targeted inactivation of hepatic Abca1 causes profound hypoalphalipoproteinemia and kidney hypercatabolism of apoA-I. J. Clin. Invest. 2005, 115, 1333–1342. [Google Scholar]
- Brunham, L.R.; Kruit, J.K.; Iqbal, J.; Fievet, C.; Timmins, J.M.; Pape, T.D.; Coburn, B.A.; Bissada, N.; Staels, B.; Groen, A.K.; et al. Intestinal ABCA1 directly contributes to HDL biogenesis in vivo. J. Clin. Invest. 2006, 116, 1052–1062. [Google Scholar] [CrossRef]
- Basso, F.; Freeman, L.; Knapper, C.L.; Remaley, A.; Stonik, J.; Neufeld, E.B.; Tansey, T.; Amar, M.J.; Fruchart-Najib, J.; Duverger, N.; et al. Role of the hepatic ABCA1 transporter in modulating intrahepatic cholesterol and plasma HDL cholesterol concentrations. J. Lipid Res. 2003, 44, 296–302. [Google Scholar] [CrossRef]
- Gao, J.; Xu, Y.; Yang, Y.; Yang, Y.; Zheng, Z.; Jiang, W.; Hong, B.; Yan, X.; Si, S. Identification of upregulators of human ATP-binding cassette transporter A1 via high-throughput screening of a synthetic and natural compound library. J. Biomol. Screen. 2008, 13, 648–656. [Google Scholar] [CrossRef]
- Tsuji, N.; Kobayashi, M.; Nagashima, K.; Wakisaka, Y.; Koizumi, K. A new antifungal antibiotic, trichostatin. J. Antibiot.(Tokyo) 1976, 29, 1–6. [Google Scholar] [CrossRef]
- Tsuji, N.; Kobayashi, M. Trichostatin C, a glucopyranosyl hydroxamate. J. Antibiot (Tokyo). 1978, 31, 939–944. [Google Scholar] [CrossRef]
- Yoshida, M.; Nomura, S.; Beppu, T. Effects of trichostatins on differentiation of murine erythroleukemia cells. Cancer Res. 1987, 47, 3688–3691. [Google Scholar]
- Rosenfeld, M.G.; Glass, C.K. Coregulator codes of transcriptional regulation by nuclear receptors. J. Biol. Chem. 2001, 276, 36865–36868. [Google Scholar] [CrossRef]
- Hermanson, O.; Glass, C.K.; Rosenfeld, M.G. Nuclear receptor coregulators: Multiple modes of modification. Trends Endocrinol. Metab. 2002, 13, 55–60. [Google Scholar] [CrossRef]
- Huang, E.Y.; Zhang, J.; Miska, E.A.; Guenther, M.G.; Kouzarides, T.; Lazar, M.A. Nuclear receptor corepressors partner with class II histone deacetylases in a Sin3-independent repression pathway. Genes Dev. 2000, 14, 45–54. [Google Scholar]
- Guenther, M.G.; Lane, W.S.; Fischle, W.; Verdin, E.; Lazar, M.A.; Shiekhattar, R. A core SMRT corepressor complex containing HDAC3 and TBL1, a WD40-repeat protein linked to deafness. Genes Dev. 2000, 14, 1048–1057. [Google Scholar]
- Wen, Y.D.; Perissi, V.; Staszewski, L.M.; Yang, W.M.; Krones, A.; Glass, C.K.; Rosenfeld, M.G.; Seto, E. The histone deacetylase-3 complex contains nuclear receptor corepressors. Proc. Natl. Acad. Sci. USA 2000, 97, 7202–7207. [Google Scholar]
- Laribee, R.N.; Klemsz, M.J. Loss of PU.1 expression following inhibition of histone deacetylases. J. Immunol. 2001, 167, 5160–5166. [Google Scholar]
- Klingenberg, R.; Hansson, G.K. Treating inflammation in atherosclerotic cardiovascular disease: Emerging therapies. Eur. Heart J. 2009, 30, 2838–2844. [Google Scholar] [CrossRef]
- Moubayed, S.P.; Heinonen, T.M.; Tardif, J.C. Anti-inflammatory drugs and atherosclerosis. Curr. Opin. Lipidol. 2007, 18, 638–644. [Google Scholar] [CrossRef]
- Schmitz, G.; Langmann, T. Transcriptional regulatory networks in lipid metabolism control ABCA1 expression. Biochim. Biophys. Acta 2005, 1735, 1–19. [Google Scholar] [CrossRef]
- Ohashi, R.; Mu, H.; Wang, X.; Yao, Q.; Chen, C. Reverse cholesterol transport and cholesterol efflux in atherosclerosis. QJM 2005, 98, 845–856. [Google Scholar] [CrossRef]
- Francis, G.A.; Knopp, R.H.; Oram, J.F. Defective removal of cellular cholesterol and phospholipids by apolipoprotein A-I in Tangier Disease. J. Clin. Invest. 1995, 96, 78–87. [Google Scholar] [CrossRef]
- Oram, J.F.; Lawn, R.M. ABCA1. The gatekeeper for eliminating excess tissue cholesterol. J. Lipid Res. 2001, 42, 1173–1179. [Google Scholar]
- Bao, Y.; Yang, Y.; Wang, L.; Gao, L.; Jiang, W.; Wang, L.; Si, S.; Hong, B. Identification of trichostatin A as a novel transcriptional up-regulator of scavenger receptor BI both in HepG2 and RAW264.7 cells. Atherosclerosis 2009, 204, 127–135. [Google Scholar] [CrossRef]
- Langmann, T.; Porsch-Ozcürümez, M.; Heimerl, S.; Probst, M.; Moehle, C.; Taher, M.; Borsukova, H.; Kielar, D.; Kaminski, W.E.; Dittrich-Wengenroth, E.; et al. Identification of sterol-independent regulatory elements in the human ATP-binding cassette transporter A1 promoter: Role of Sp1/3, E-box binding factors, and an oncostatin M-responsive element. J. Biol. Chem. 2002, 277, 14443–14450. [Google Scholar]
- Langmann, T.; Buechler, C.; Ries, S.; Schaeffler, A.; Aslanidis, C.; Schuierer, M.; Weiler, M.; Sandhoff, K.; de Jong, P.J.; Schmitz, G. Transcription factors Sp1 and AP-2 mediate induction of acid sphingomyelinase during monocytic differentiation. J. Lipid Res. 1999, 40, 870–880. [Google Scholar]
- Yang, X.P.; Freeman, L.A.; Knapper, C.L.; Amar, M.J.; Remaley, A.; Brewer, H.B., Jr.; Santamarina-Fojo, S. The E-box motif in the proximal ABCA1 promoter mediates transcriptional repression of the ABCA1 gene. J. Lipid Res. 2002, 43, 297–306. [Google Scholar]
- Baranova, I.; Vishnyakova, T.; Bocharov, A.; Chen, Z.; Remaley, A.T.; Stonik, J.; Eggerman, T.L.; Patterson, A.P. Lpopolysaccharide down regulates both scavenger receptor B1 and ATP binding cassette transporter A1 in RAW cells. Infect. Immun. 2002, 70, 2995–3003. [Google Scholar] [CrossRef]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xu, Y.; Xu, Y.; Bao, Y.; Hong, B.; Si, S. Identification of Dehydroxytrichostatin A as a Novel Up-Regulator of the ATP-Binding Cassette Transporter A1 (ABCA1). Molecules 2011, 16, 7183-7198. https://doi.org/10.3390/molecules16097183
Xu Y, Xu Y, Bao Y, Hong B, Si S. Identification of Dehydroxytrichostatin A as a Novel Up-Regulator of the ATP-Binding Cassette Transporter A1 (ABCA1). Molecules. 2011; 16(9):7183-7198. https://doi.org/10.3390/molecules16097183
Chicago/Turabian StyleXu, Yang, Yanni Xu, Yi Bao, Bin Hong, and Shuyi Si. 2011. "Identification of Dehydroxytrichostatin A as a Novel Up-Regulator of the ATP-Binding Cassette Transporter A1 (ABCA1)" Molecules 16, no. 9: 7183-7198. https://doi.org/10.3390/molecules16097183