A Screening of a Library of T7 Phage-Displayed Peptide Identifies E2F-4 as an Etoposide-Binding Protein

Abstract
:1. Introduction


2. Results and Discussion
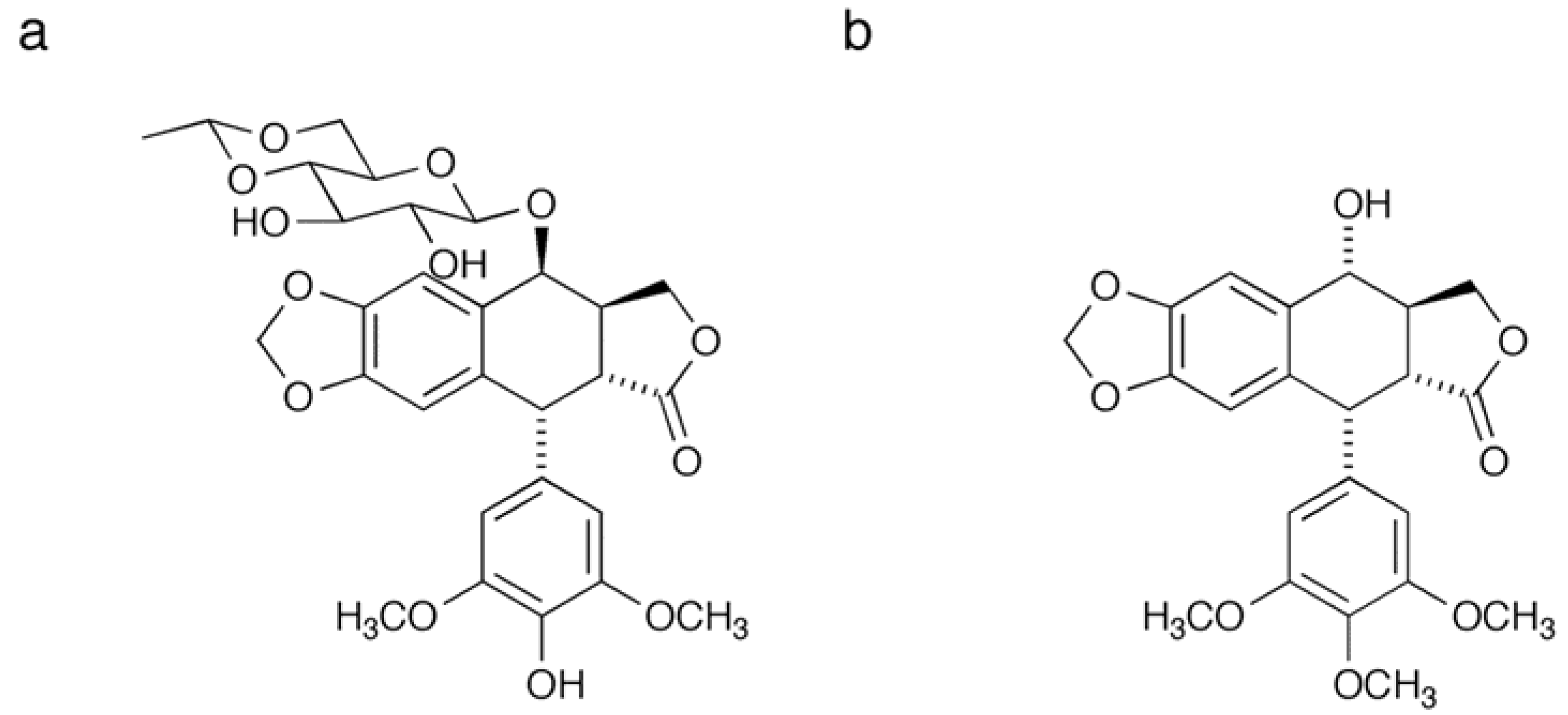
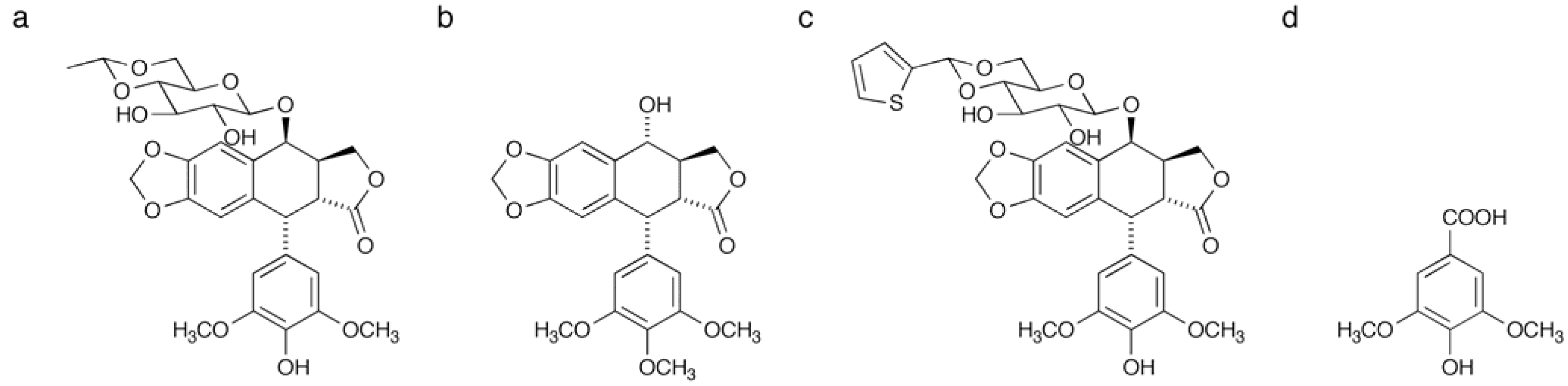
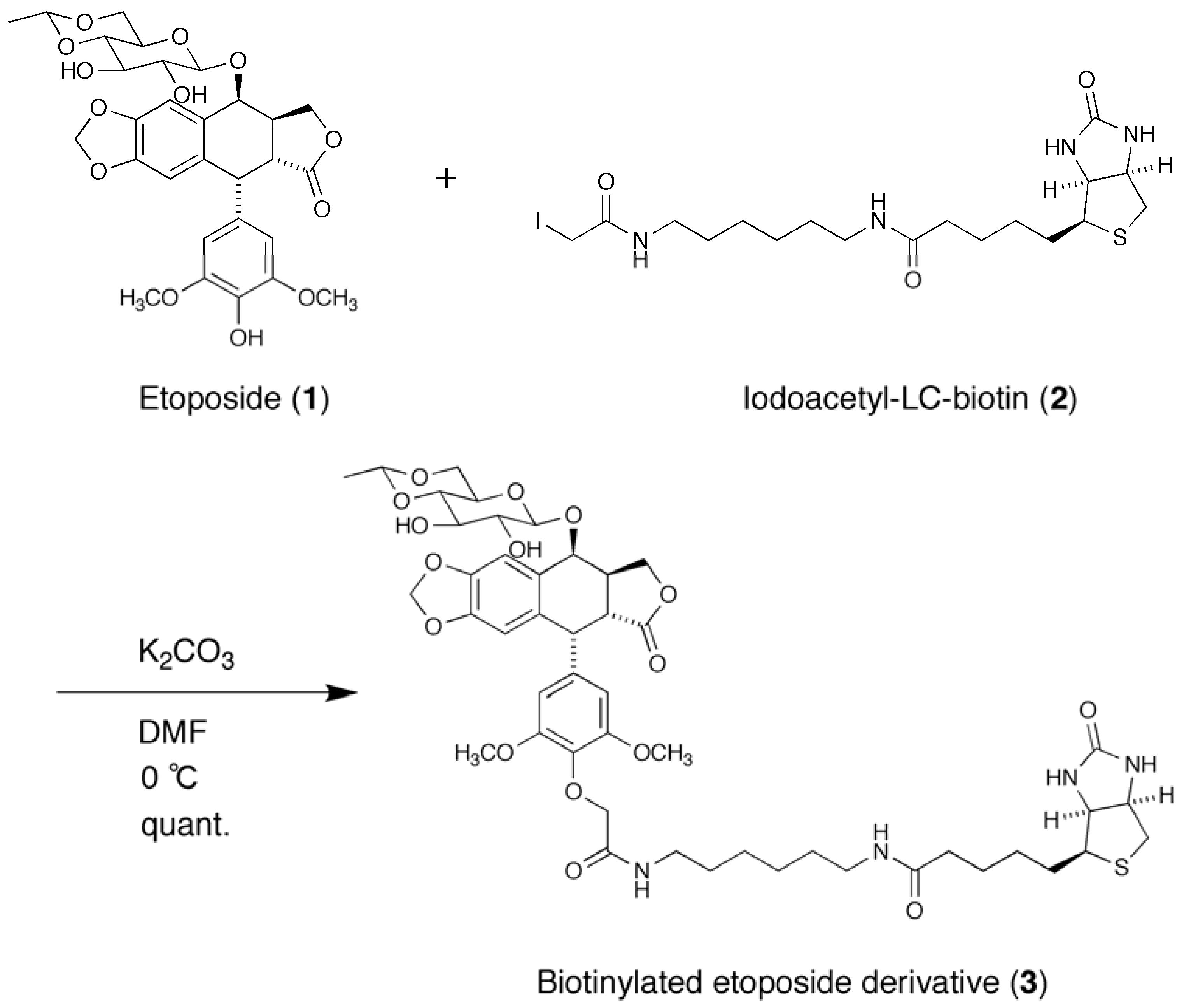
2.1. Synthesis of a Biotinylated Etoposide Derivative

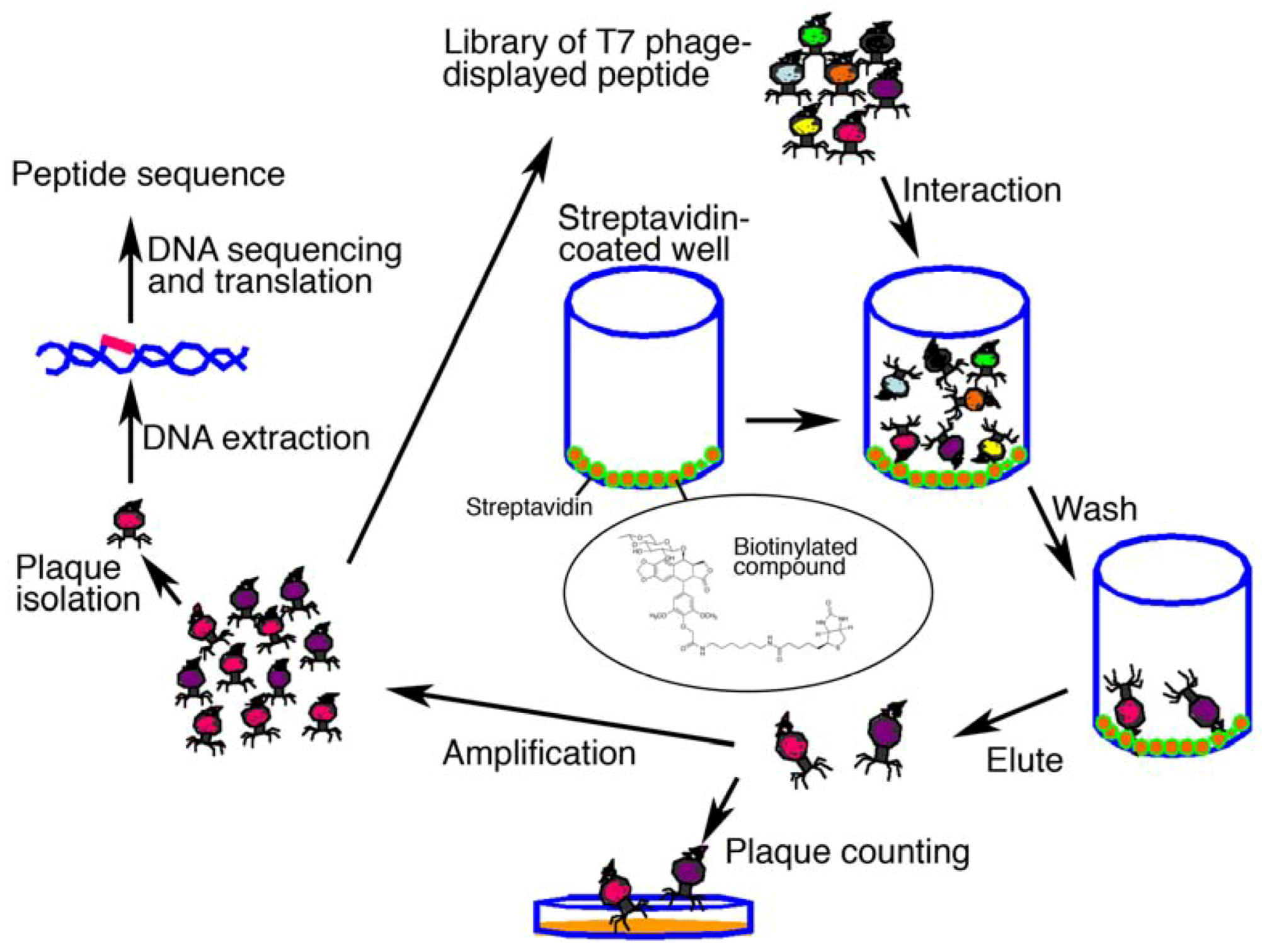
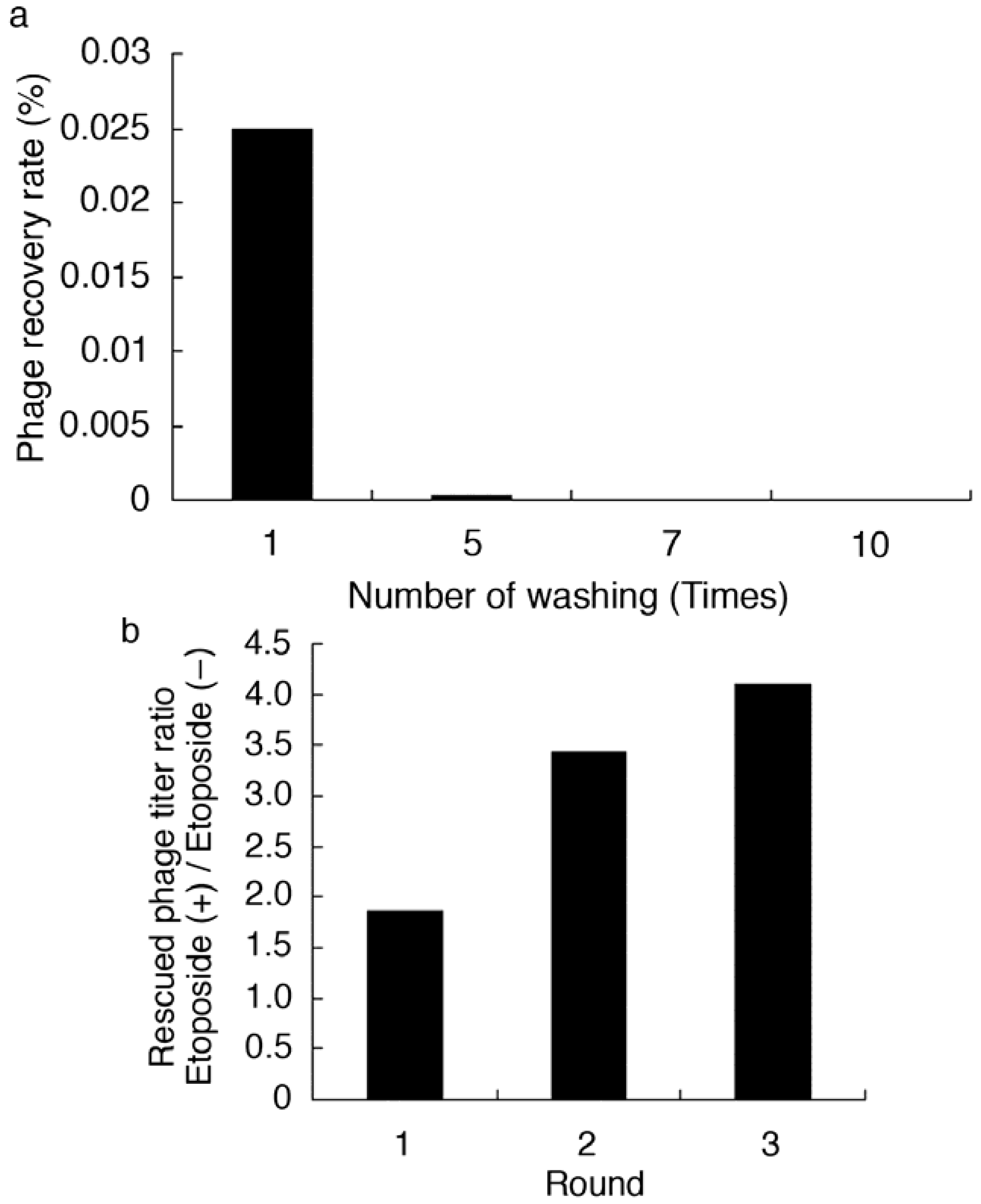
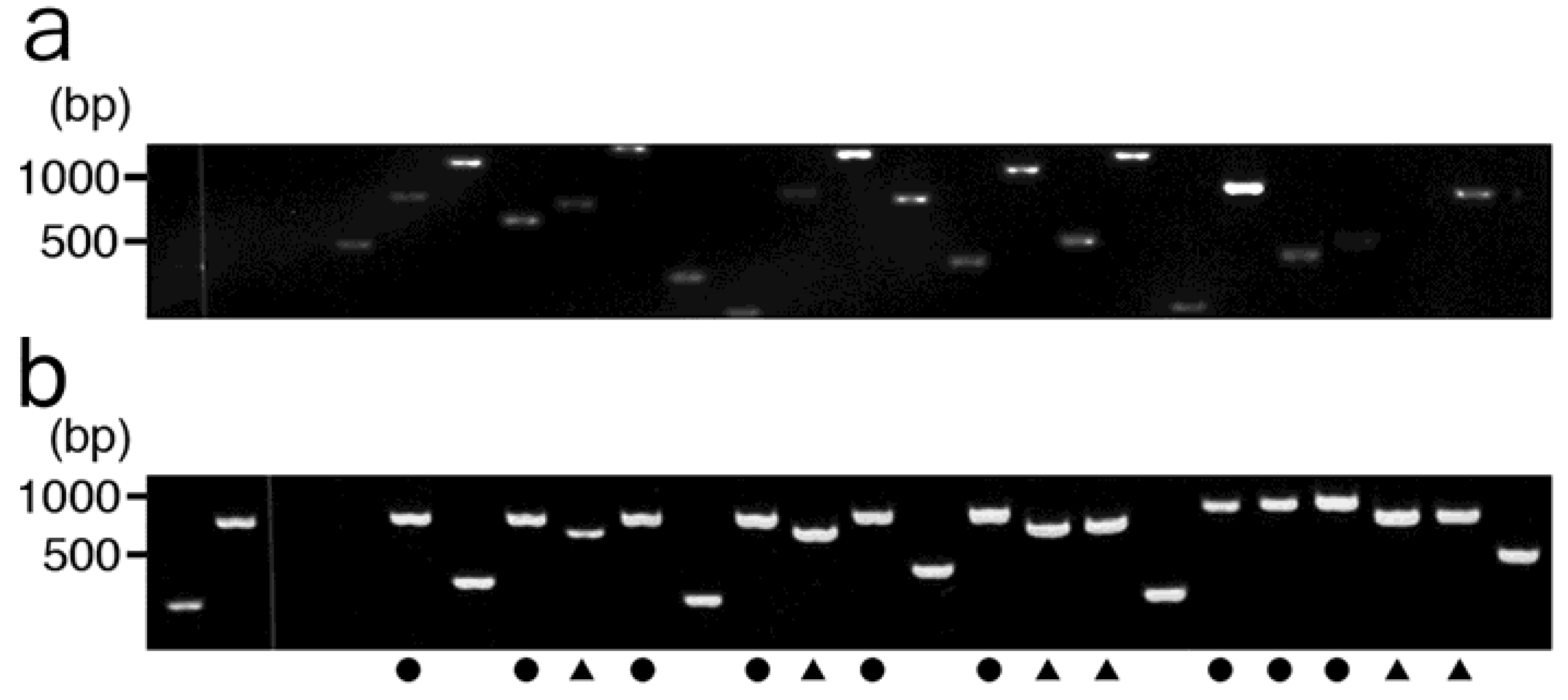
2.2. Affinity Selection of a Library of T7 Phage-displayed Peptides


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immobil. etoposide | 50 µL of 20 µM in 100 mM Tris-HCl (pH 8.0) − 2% DMSO soln. (4 °C, O/N) | |
| Blocking | 200 µL of 3% skimmed milk (r.t., 1 h) | |
| Input of library | 1.0 × 108 pfu | |
| Incubation | r.t., 3 h | |
| Wash condition | 200 µL of wash buffer for 5 times (5 min each on shaker) | |
| Wash buffer | 200 µL of 100 mM Tris-HCl (pH 8.0) containing 60 mM NaCl and 0.3% Tween 20 | |
| Elution condition | 100 µL of elution buffer (O/N on shaker) | |
| 100 µL of elution buffer for 4 times (5 min each on shaker) | ||
| (Total 500 µL) | ||
| Elution buffer | Round 1 | 3 M NaCl |
| 2 | A mixture containing 3 M NaCl, 2 M urea and 3% Tween 20 | |
| 3 | A mixture containing 4 M NaCl, 2 M urea and 3% Tween 20 | |
| 4 | Host E. coli (BLT5615) culture (100 µL for 30 min on shaker) | |

2.3. Interaction Analysis Between Etoposide and Synthetic ETBP Using SPR Biosensor
2.4. Prediction of the Binding Partner for Etoposide by Similarity Search and Biological Data
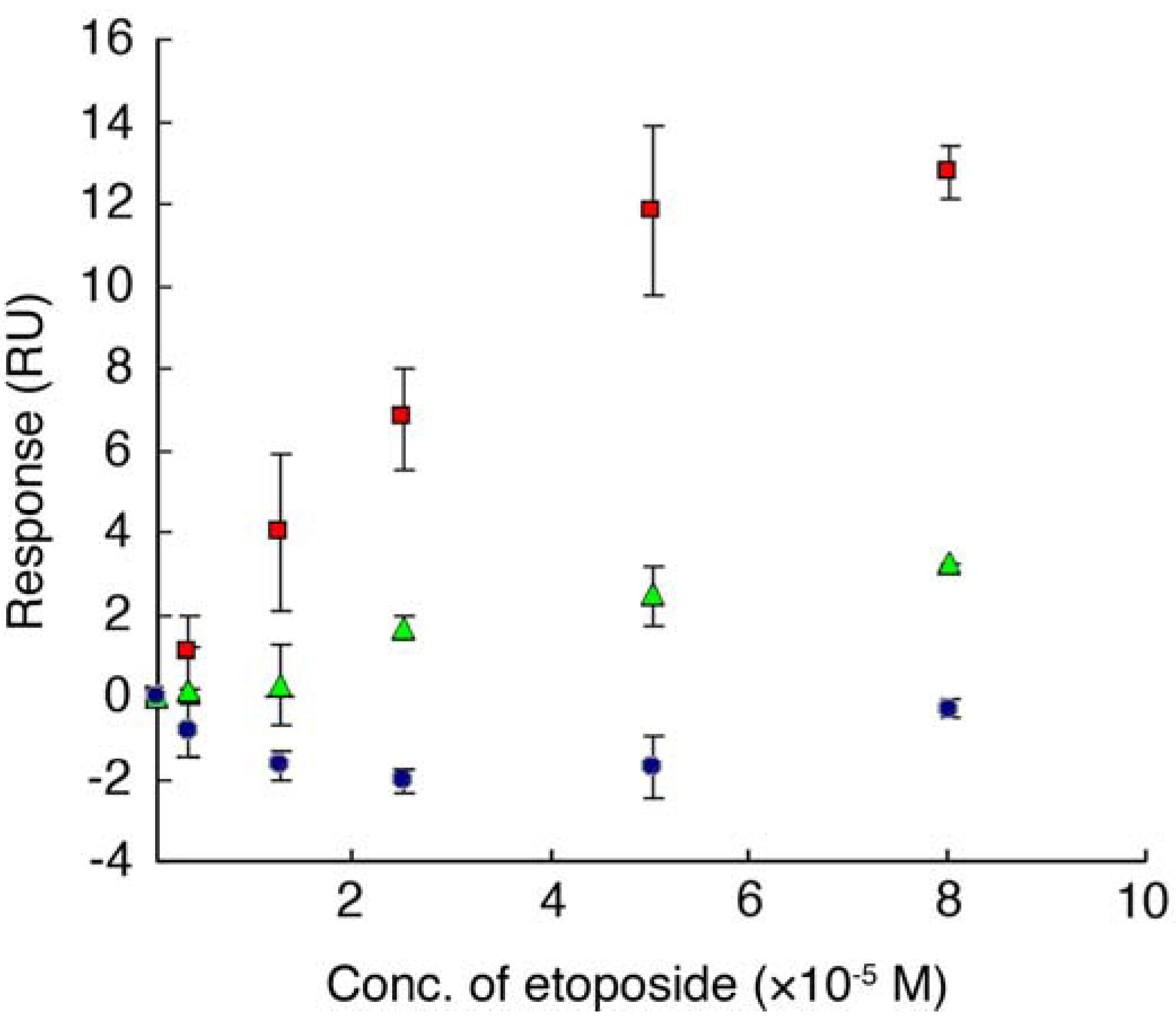
 , etoposide and ETBP16,
, etoposide and ETBP16,  , etoposide and ETBP10,
, etoposide and ETBP10,  , etoposide and a control peptide HG102. Data are means ± S.D. of at least three independent experiments.
, etoposide and ETBP16, , etoposide and ETBP10, , etoposide and a control peptide HG102. Data are means ± S.D. of at least three independent experiments.
, etoposide and a control peptide HG102. Data are means ± S.D. of at least three independent experiments.
, etoposide and ETBP16, , etoposide and ETBP10, , etoposide and a control peptide HG102. Data are means ± S.D. of at least three independent experiments.

2.5. Interaction Analysis between Etoposide and E2F-4 Using an SPR Biosensor
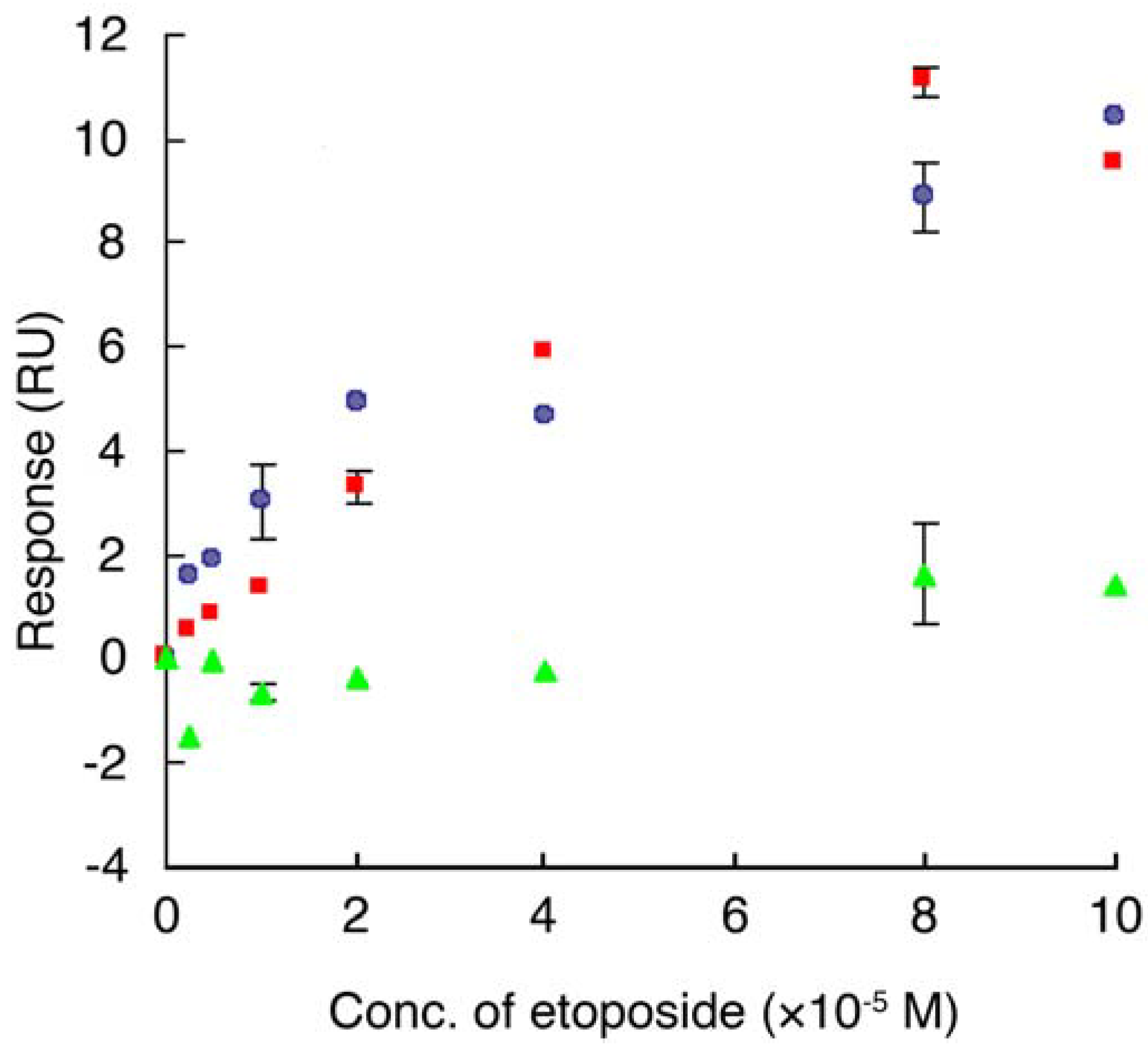
, etoposide and GST-E2F-4, , etoposide and top II, , etoposide and GST. Data are means ± S.D. of at least three independent experiments.
, etoposide and GST-E2F-4, , etoposide and top II, , etoposide and GST. Data are means ± S.D. of at least three independent experiments.
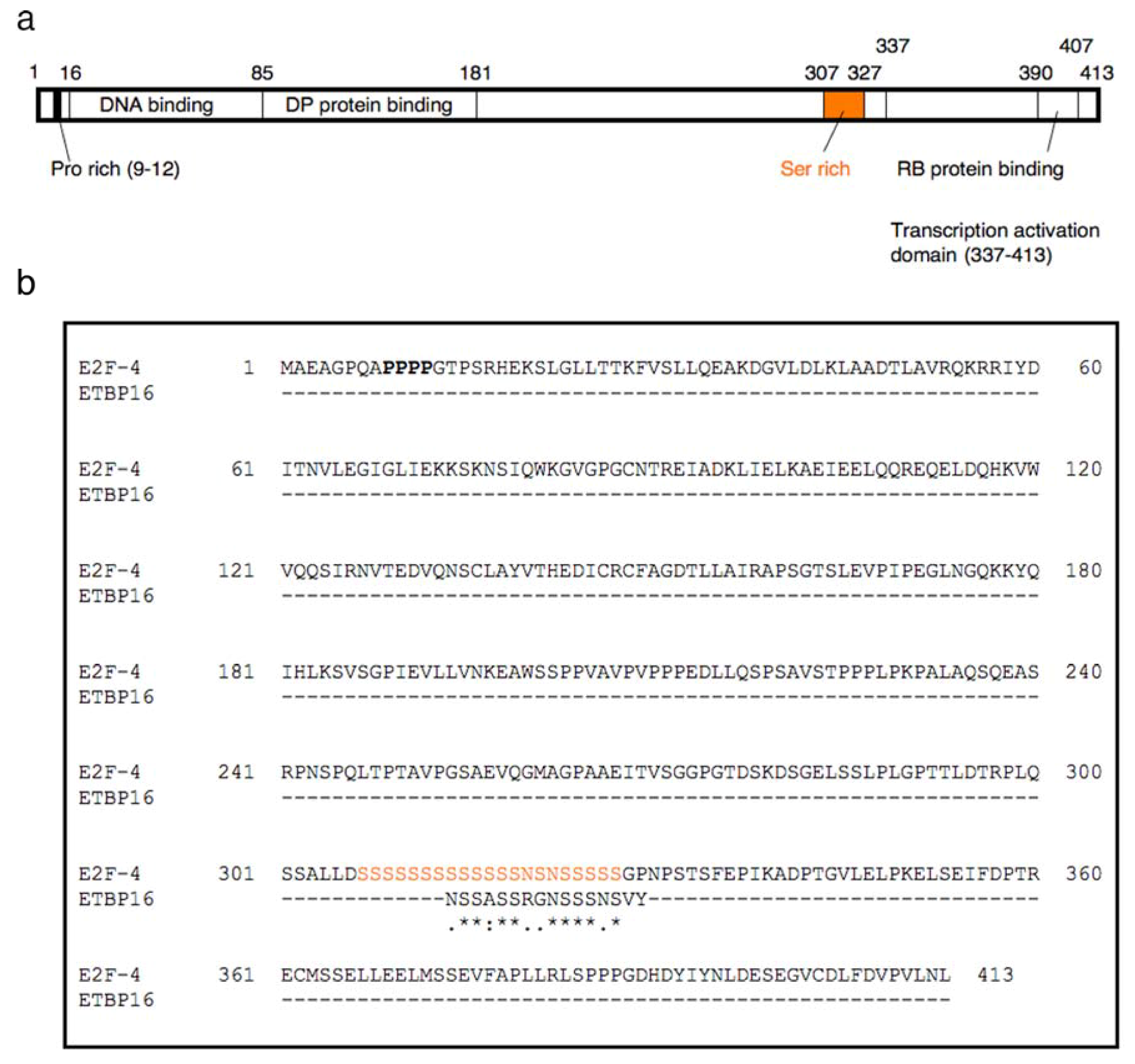
2.5. Structure-binding Relationships of Etoposide for E2F-4

| Compound: | Etoposide | Podophyllotoxin | Teniposide | Syringic acid |
|---|---|---|---|---|
| KD (×10−5 M): | 7.5 | 30.8 | 65.4 | ND |
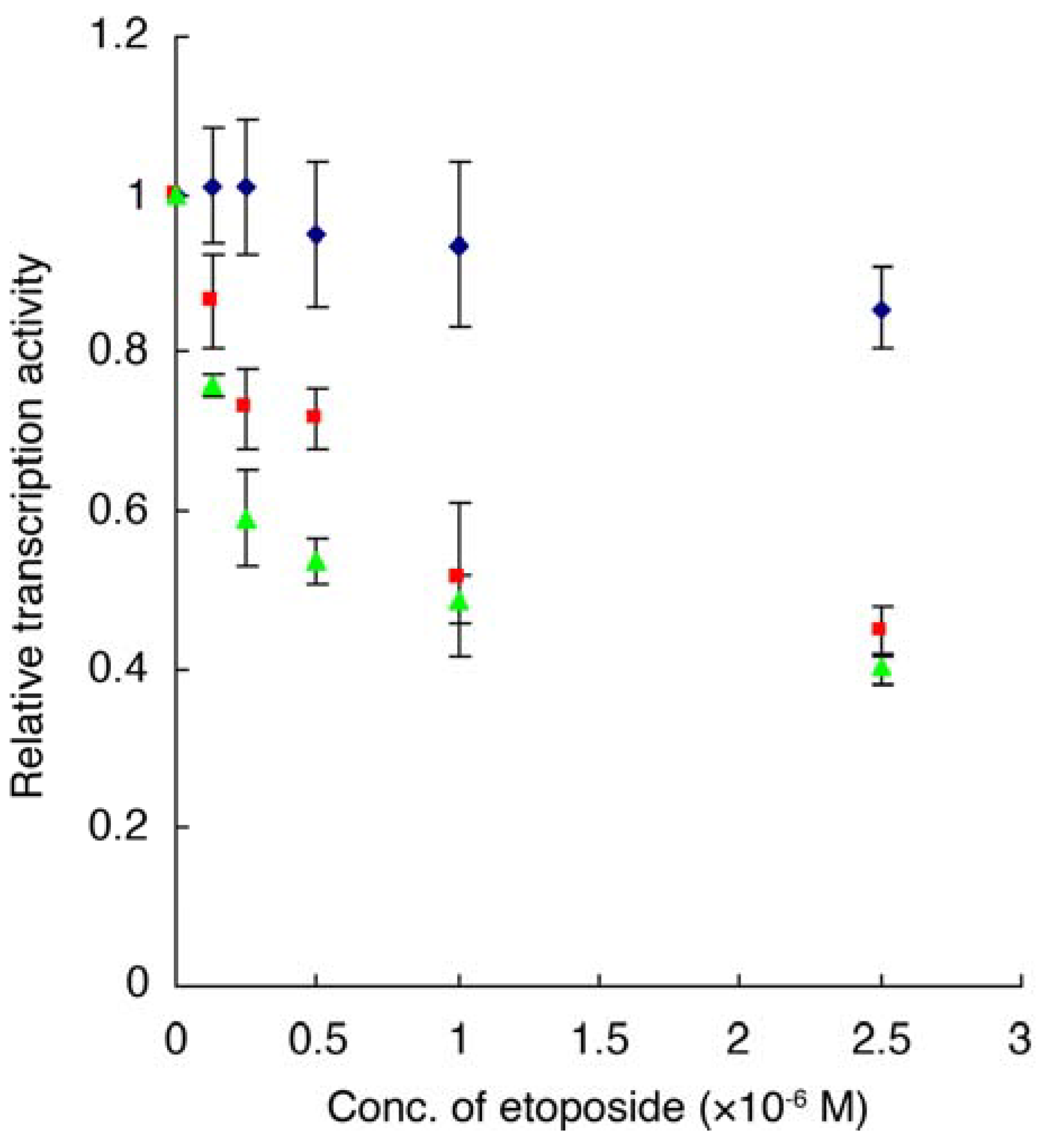
2.6. Luciferase Reporter Gene Assay
, E2F4/DP1 expressing cell, , E2F-4/DP2 expressing cell,  , control cell transfected with vehicle. Data are means ± S.D. of three independent experiments.
, E2F4/DP1 expressing cell, , E2F-4/DP2 expressing cell, , control cell transfected with vehicle. Data are means ± S.D. of three independent experiments.
, control cell transfected with vehicle. Data are means ± S.D. of three independent experiments.
, E2F4/DP1 expressing cell, , E2F-4/DP2 expressing cell, , control cell transfected with vehicle. Data are means ± S.D. of three independent experiments.
| Complex: | E2F-4/DP1 | E2F-4/DP2 | E2F-1/DP1 |
|---|---|---|---|
| IC50 (µM) of etoposide | 1.56 | 1.16 | 3.59 |
3. Experimental
3.1. Instrumentation
 20 × 250 mm, UG120Å, Shiseido, Tokyo, Japan). Synthetic peptide was verified by a LC-MS using Bruker Daltonics Esquire 3000 plus (Bruker Daltonics K. K., Kanagawa, Japan). Recombinant GST-E2F-4 was purified using a fast performance liquid chromatography (FPLC) instrument (ÄKTA explorer 10s, GE Healthcare, Piscataway, NJ, USA) with 1 mL of GSTrap column (GE Healthcare). The SPR biosensor (Biacore®3000), CM5 sensor chip and an amine coupling kit were from GE Healthcare. Optical density (OD) was determined using Wallac VictorTM 1420 multilabel counter (PerkinElmer, Waltham, MA, USA).
20 × 250 mm, UG120Å, Shiseido, Tokyo, Japan). Synthetic peptide was verified by a LC-MS using Bruker Daltonics Esquire 3000 plus (Bruker Daltonics K. K., Kanagawa, Japan). Recombinant GST-E2F-4 was purified using a fast performance liquid chromatography (FPLC) instrument (ÄKTA explorer 10s, GE Healthcare, Piscataway, NJ, USA) with 1 mL of GSTrap column (GE Healthcare). The SPR biosensor (Biacore®3000), CM5 sensor chip and an amine coupling kit were from GE Healthcare. Optical density (OD) was determined using Wallac VictorTM 1420 multilabel counter (PerkinElmer, Waltham, MA, USA).3.2. Materials
3.2.1. Chemistry
3.2.2. Biology
3.3. Synthesis of Biotinylated Etoposide Derivative
3.4. Screening of a Library of T7 Phage-displayed Peptide
3.5. Agarose Gel Electrophoresis and DNA Sequencing
3.6. Synthesis and Purification of Peptide
20 × 250 nm, UG120Å, Shiseido) and verified by LC-MS using the Bruker Daltonics Esquire 3000 plus.3.7. Protein Expression and Purification
3.8. Interaction Analysis Using an SPR Biosensor
3.9. Cell Culture
3.10. MTS Assay
3.11. Transfection
3.12. Luciferase Reporter Gene Assay
3.13. Bioinformatics Tool
4. Conclusions
Acknowledgements
References and Notes
- Keller-Juslen, C.; Kuhn, M.; Stahelin, H.; von Wartburg, A. Synthesis and antimitotic activity of glycosidic lignan derivatives related to podophyllotoxin. J. Med. Chem. 1971, 14, 936–940. [Google Scholar] [CrossRef]
- Baldwin, E.L.; Osheroff, N. Etoposide, topoisomerase II and cancer. Curr. Med. Chem. Anticancer Agents 2005, 5, 363–372. [Google Scholar] [CrossRef]
- Chiu, C.C.; Lin, C.H.; Fang, K. Etoposide (VP-16) sensitizes p53-deficient human non-small cell lung cancer cells to caspase-7-mediated apoptosis. Apoptosis 2005, 10, 643–650. [Google Scholar] [CrossRef]
- Burden, D.A.; Kingma, P.S.; Froelich-Ammon, S.J.; Bjornsti, M.A.; Patchan, M.W.; Thompson, R.B.; Osheroff, N. Topoisomerase II.etoposide interactions direct the formation of drug-induced enzyme-DNA cleavage complexes. J. Biol. Chem. 1996, 271, 29238–29244. [Google Scholar]
- Chen, G.L.; Yang, L.; Rowe, T.C.; Halligan, B.D.; Tewey, K.M.; Liu, L.F. Nonintercalative antitumor drugs interfere with the breakage-reunion reaction of mammalian DNA topoisomerase II. J. Biol. Chem. 1984, 259, 13560–13566. [Google Scholar]
- Soues, S.; Wiltshire, M.; Smith, P.J. Differential sensitivity to etoposide (VP-16)-induced S phase delay in a panel of small-cell lung carcinoma cell lines with G1/S phase checkpoint dysfunction. Cancer Chemother. Pharmacol. 2001, 47, 133–140. [Google Scholar] [CrossRef]
- Davis, M.P.; Murthy, M.S.; Simon, J.; Wise, H., Minton. Successful management of small cell carcinoma of the bladder with cisplatin and etoposide. J. Urol. 1989, 142, 817. [Google Scholar]
- Walther, P.J.; Williams, S.D.; Troner, M.; Greco, F.A.; Birch, R.; Einhorn, L.H. Phase II study of etoposide for carcinoma of the bladder: The Southeastern Cancer Study Group experience. Cancer Treat. Rep. 1986, 70, 1337–1338. [Google Scholar]
- Morris, M.; Brader, K.R.; Burke, T.W.; Levenback, C.F.; Gershenson, D.M. A phase II study of prolonged oral etoposide in advanced or recurrent carcinoma of the cervix. Gynecol. Oncol. 1998, 70, 215–218. [Google Scholar] [CrossRef]
- Kredentser, D.C. Etoposide (VP-16), ifosfamide/mesna, and cisplatin chemotherapy for advanced and recurrent carcinoma of the cervix. Gynecol. Oncol. 1991, 43, 145–148. [Google Scholar] [CrossRef]
- Slayton, R.E.; Blessing, J.A.; Homesley, H.D. Phase II trial of etoposide in the management of advanced or recurrent non-squamous cell carcinoma of the cervix: A Gynecologic Oncology Group study. Cancer Treat. Rep. 1984, 68, 1513–1514. [Google Scholar]
- Dubrez, L.; Goldwasser, F.; Genne, P.; Pommier, Y.; Solary, E. The role of cell cycle regulation and apoptosis triggering in determining the sensitivity of leukemic cells to topoisomerase I and II inhibitors. Leukemia 1995, 9, 1013–1024. [Google Scholar]
- Lee, Y.S.; Lee, E.K.; Han, I.O.; Park, S.H. Etoposide-induced Smad6 expression is required for the G1 to S phase transition of the cell cycle in CMT-93 mouse intestinal epithelial cells. Exp. Mol. Med. 2008, 40, 43–51. [Google Scholar] [CrossRef]
- Kurosu, T.; Takahashi, Y.; Fukuda, T.; Koyama, T.; Miki, T.; Miura, O. p38 MAP kinase plays a role in G2 checkpoint activation and inhibits apoptosis of human B cell lymphoma cells treated with etoposide. Apoptosis 2005, 10, 1111–1120. [Google Scholar] [CrossRef]
- Nakada, S.; Katsuki, Y.; Imoto, I.; Yokoyama, T.; Nagasawa, M.; Inazawa, J.; Mizutani, S. Early G2/M checkpoint failure as a molecular mechanism underlying etoposide-induced chromosomal aberrations. J. Clin. Invest. 2006, 116, 80–89. [Google Scholar]
- Takakusagi, Y.; Takakusagi, K.; Sugawara, F.; Sakaguchi, K. Use of phage display technology for the determination of the targets for small-molecule therapeutics. Expert Opin. Drug Discov. 2010, 5, 361–389. [Google Scholar]
- Homola, J. Surface plasmon resonance sensors for detection of chemical and biological species. Chem. Rev. 2008, 108, 462–493. [Google Scholar]
- Komatsu, N.; Takeuchi, S.; Ikezoe, T.; Tasaka, T.; Hatta, Y.; Machida, H.; Williamson, I.K.; Bartram, C.R.; Koeffler, H.P.; Taguchi, H. Mutations of the E2F4 gene in hematological malignancies having microsatellite instability. Blood 2000, 95, 1509–1510. [Google Scholar]
- Litovchick, L.; Sadasivam, S.; Florens, L.; Zhu, X.; Swanson, S.K.; Velmurugan, S.; Chen, R.; Washburn, M.P.; Liu, X.S.; DeCaprio, J.A. Evolutionarily conserved multisubunit RBL2/p130 and E2F4 protein complex represses human cell cycle-dependent genes in quiescence. Mol. Cell 2007, 26, 539–551. [Google Scholar] [CrossRef]
- Chang, W.Y.; Bryce, D.M.; D’Souza, S.J.; Dagnino, L. The DP-1 transcription factor is required for keratinocyte growth and epidermal stratification. J. Biol. Chem. 2004, 279, 51343–51353. [Google Scholar]
- Maehara, K.; Yamakoshi, K.; Ohtani, N.; Kubo, Y.; Takahashi, A.; Arase, S.; Jones, N.; Hara, E. Reduction of total E2F/DP activity induces senescence-like cell cycle arrest in cancer cells lacking functional pRB and p53. J. Cell Biol. 2005, 168, 553–560. [Google Scholar]
- Zheng, N.; Fraenkel, E.; Pabo, C.O.; Pavletich, N.P. Structural basis of DNA recognition by the heterodimeric cell cycle transcription factor E2F-DP. Genes Dev. 1999, 13, 666–674. [Google Scholar]
- Matsubara, N.; Yoshitaka, T.; Matsuno, T.; Ikeda, M.; Isozaki, H.; Tanaka, N.; Shimizu, K. Multiple tumors and a novel E2F-4 mutation. A case report. Digestion 2000, 62, 213–216. [Google Scholar] [CrossRef]
- Takashima, H.; Matsumoto, Y.; Matsubara, N.; Shirakawa, Y.; Kawashima, R.; Tanino, M.; Ito, S.; Isozaki, H.; Ouchida, M.; Meltzer, S.J.; Shimizu, K.; Tanaka, N. Effect of naturally occurring E2F-4 alterations on transcriptional activation and proliferation in transfected cells. Lab. Invest. 2001, 81, 1565–1573. [Google Scholar] [CrossRef]
- Yoshitaka, T.; Matsubara, N.; Ikeda, M.; Tanino, M.; Hanafusa, H.; Tanaka, N.; Shimizu, K. Mutations of E2F-4 trinucleotide repeats in colorectal cancer with microsatellite instability. Biochem. Biophys. Res. Commun. 1996, 227, 553–557. [Google Scholar]
- Smith, P.J.; Soues, S.; Gottlieb, T.; Falk, S.J.; Watson, J.V.; Osborne, R.J.; Bleehen, N.M. Etoposide-induced cell cycle delay and arrest-dependent modulation of DNA topoisomerase II in small-cell lung cancer cells. Br. J. Cancer 1994, 70, 914–921. [Google Scholar] [CrossRef]
- Takakusagi, Y.; Takakusagi, K.; Kuramochi, K.; Kobayashi, S.; Sugawara, F.; Sakaguchi, K. Identification of C10 biotinylated camptothecin (CPT-10-B) binding peptides using T7 phage display screen on a QCM device. Bioorg. Med. Chem. 2007, 15, 7590–7598. [Google Scholar]
- Takakusagi, Y.; Kuroiwa, Y.; Sugawara, F.; Sakaguchi, K. Identification of a methotrexate-binding peptide from a T7 phage display screen using a QCM device. Bioorg. Med. Chem. 2008, 16, 7410–7414. [Google Scholar] [CrossRef]
- Sample Availability: Sample of the biotinylated etoposide derivative is available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Takami, M.; Takakusagi, Y.; Kuramochi, K.; Tsukuda, S.; Aoki, S.; Morohashi, K.; Ohta, K.; Kobayashi, S.; Sakaguchi, K.; Sugawara, F. A Screening of a Library of T7 Phage-Displayed Peptide Identifies E2F-4 as an Etoposide-Binding Protein. Molecules 2011, 16, 4278-4294. https://doi.org/10.3390/molecules16054278
Takami M, Takakusagi Y, Kuramochi K, Tsukuda S, Aoki S, Morohashi K, Ohta K, Kobayashi S, Sakaguchi K, Sugawara F. A Screening of a Library of T7 Phage-Displayed Peptide Identifies E2F-4 as an Etoposide-Binding Protein. Molecules. 2011; 16(5):4278-4294. https://doi.org/10.3390/molecules16054278
Chicago/Turabian StyleTakami, Mihoko, Yoichi Takakusagi, Kouji Kuramochi, Senko Tsukuda, Satoko Aoki, Kengo Morohashi, Keisuke Ohta, Susumu Kobayashi, Kengo Sakaguchi, and Fumio Sugawara. 2011. "A Screening of a Library of T7 Phage-Displayed Peptide Identifies E2F-4 as an Etoposide-Binding Protein" Molecules 16, no. 5: 4278-4294. https://doi.org/10.3390/molecules16054278