Purification of a Lectin from Arisaema erubescens (Wall.) Schott and Its Pro-Inflammatory Effects

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
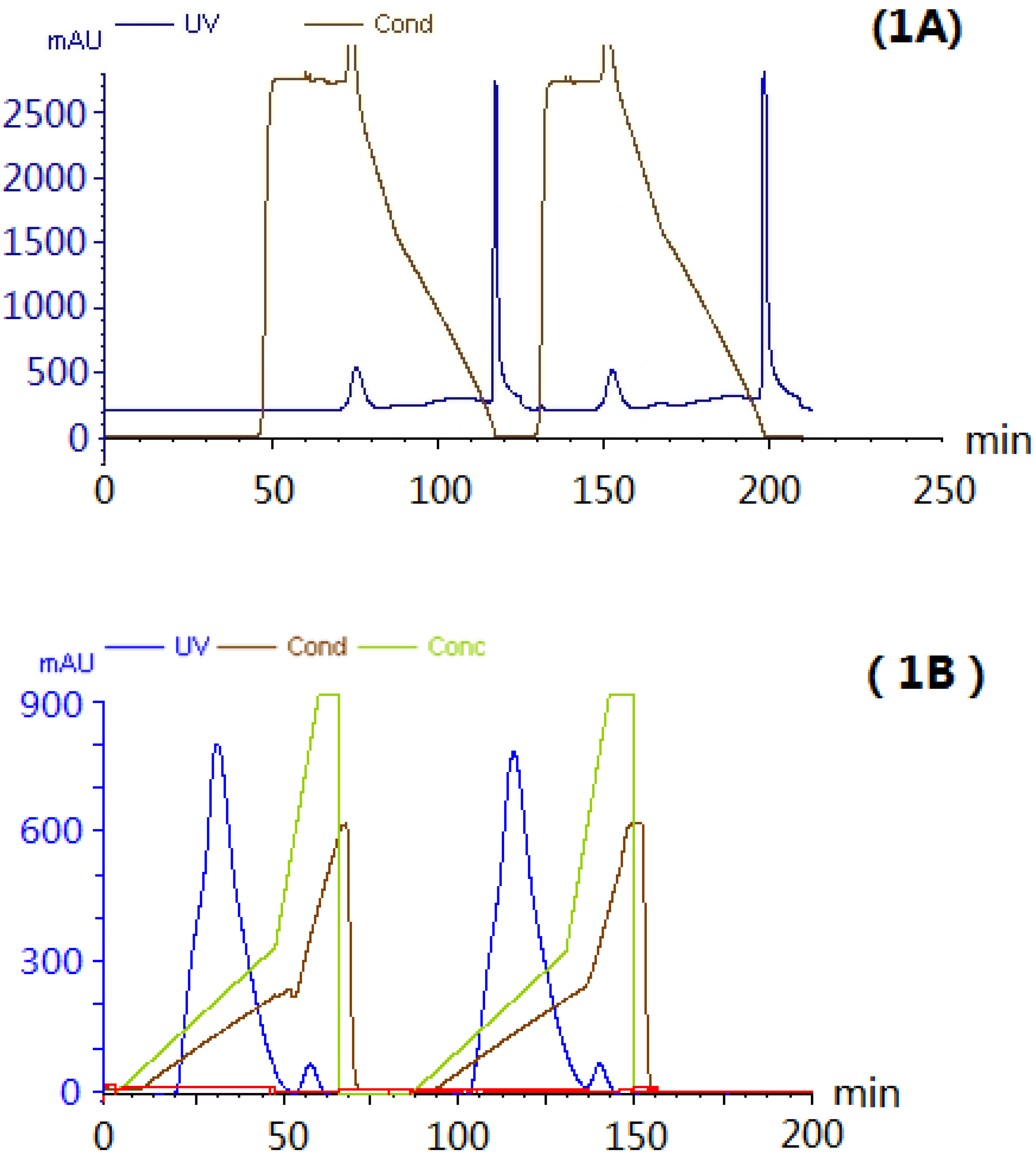
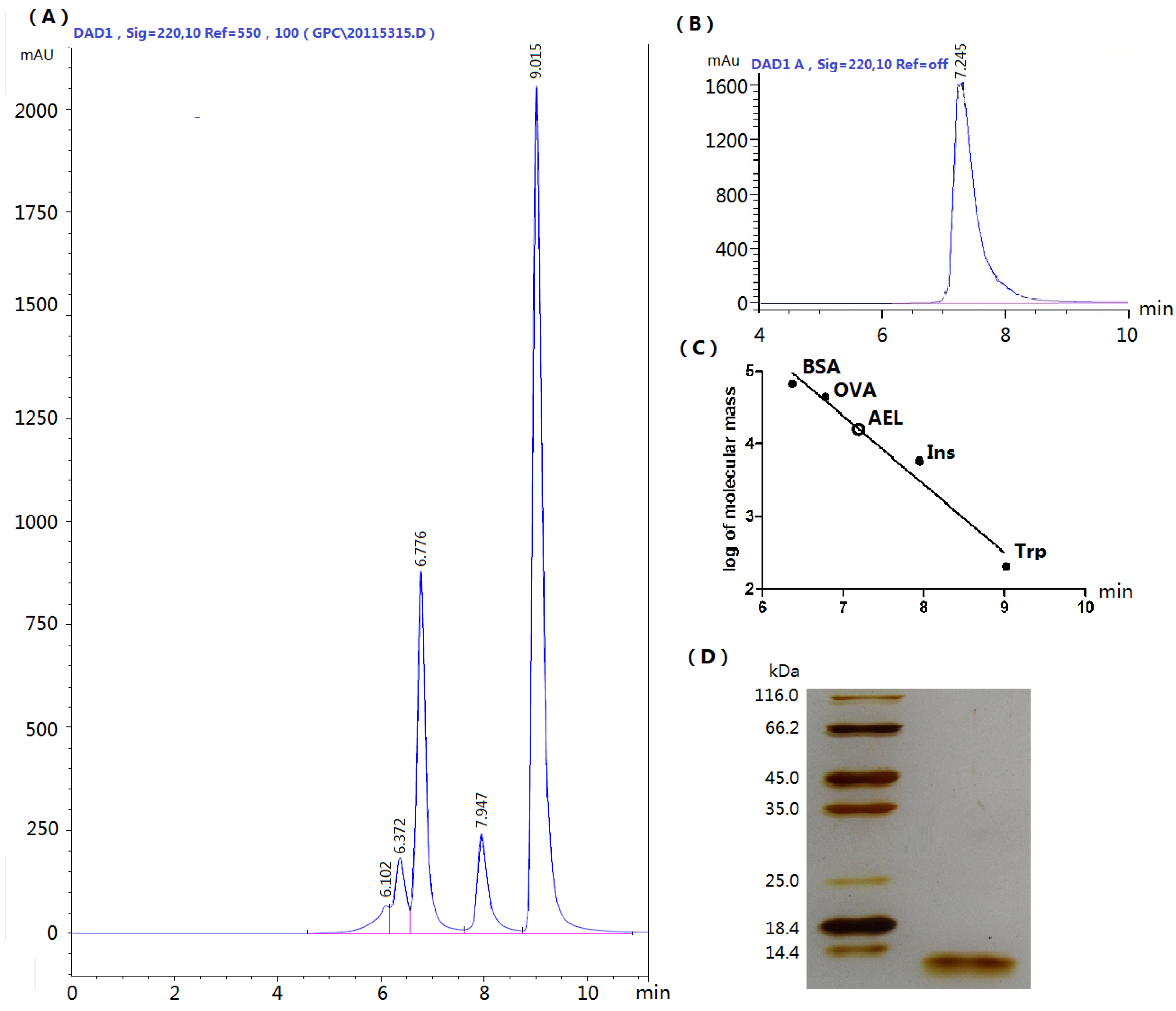
2.1. Extraction and Purification of AEL


2.2. Haemagglutination Assays
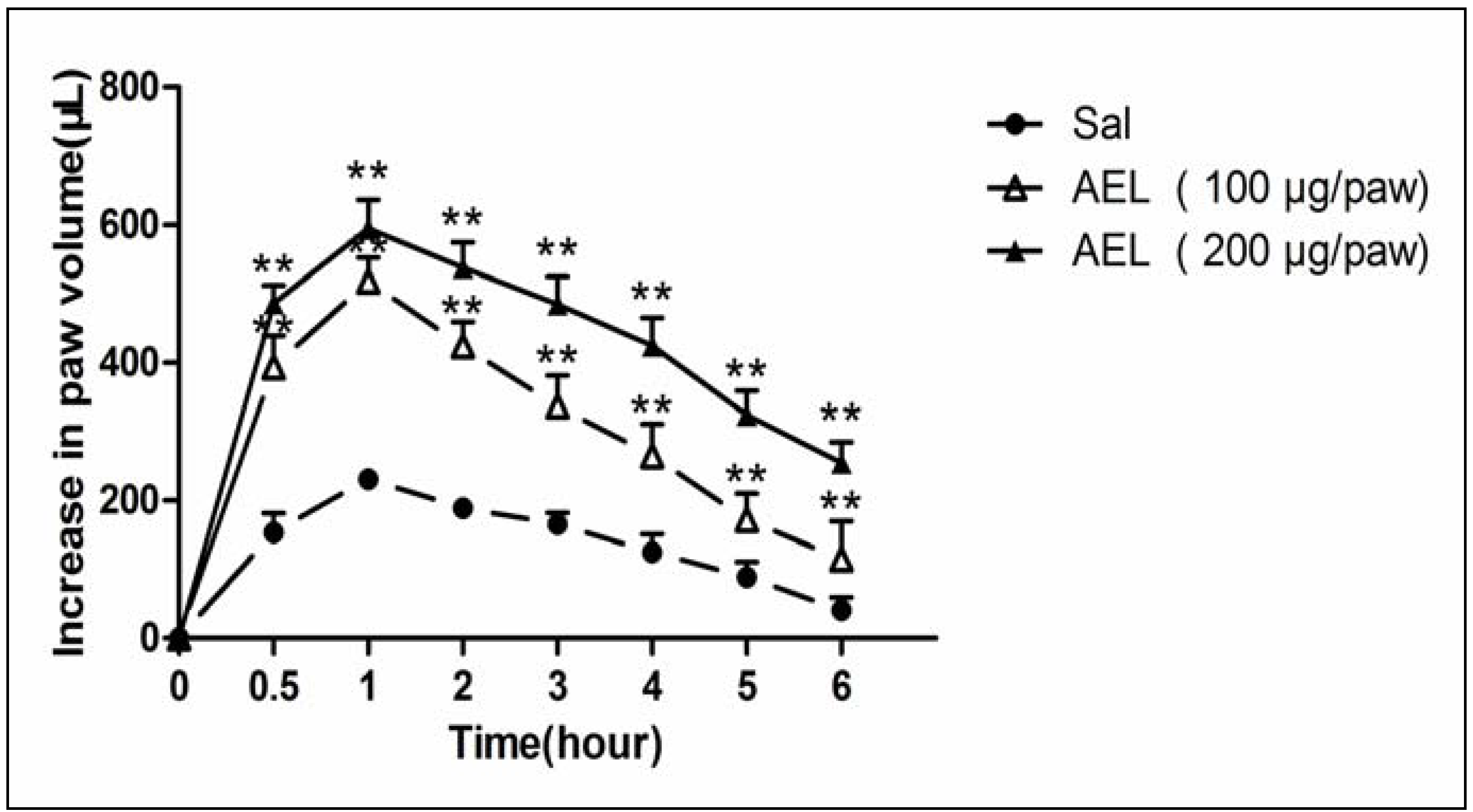
2.3. AEL-Induced Rat Paw Edema

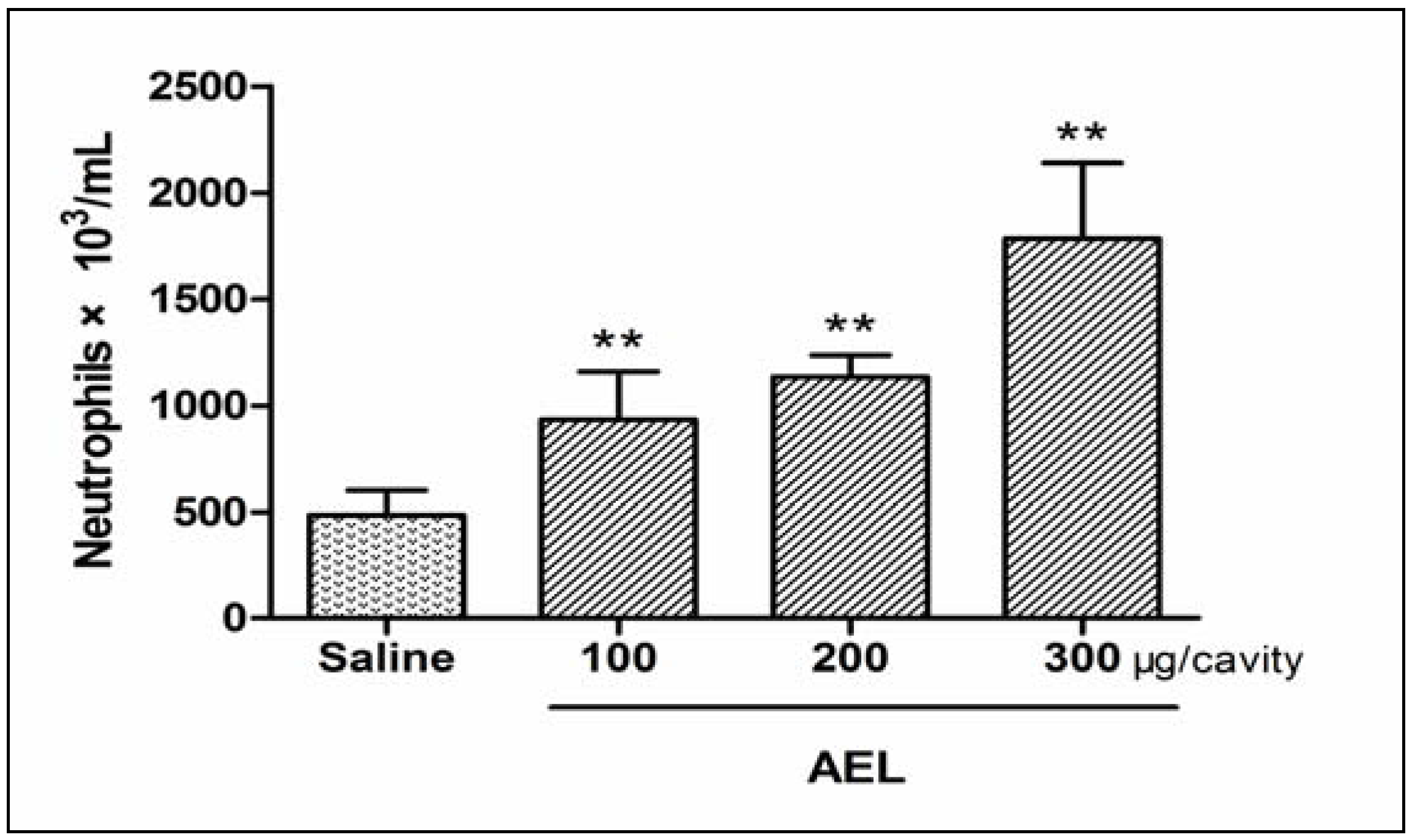
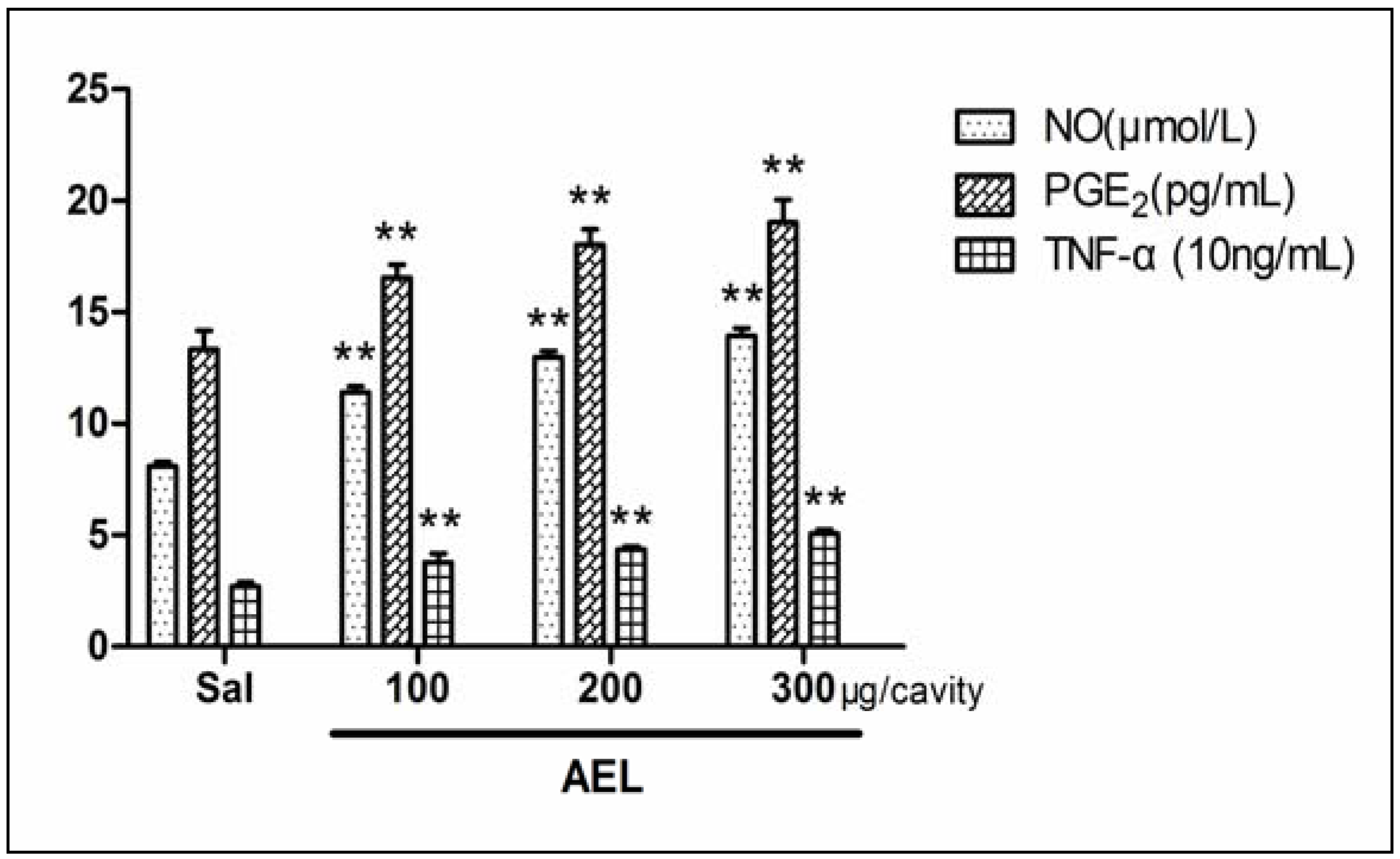
2.4. AEL-Induced Neutrophil Migration and NO, PGE2, TNF-α in Vivo


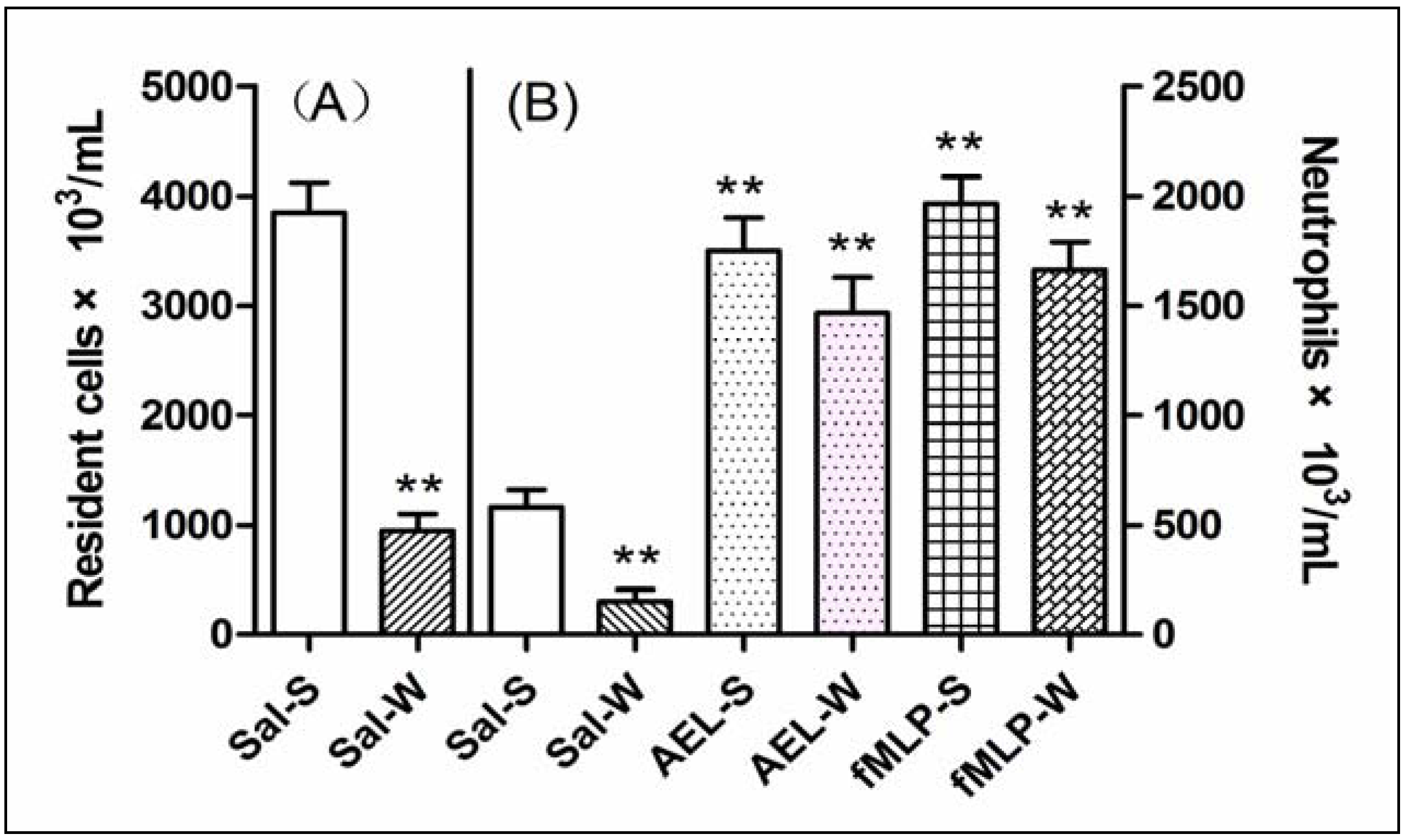
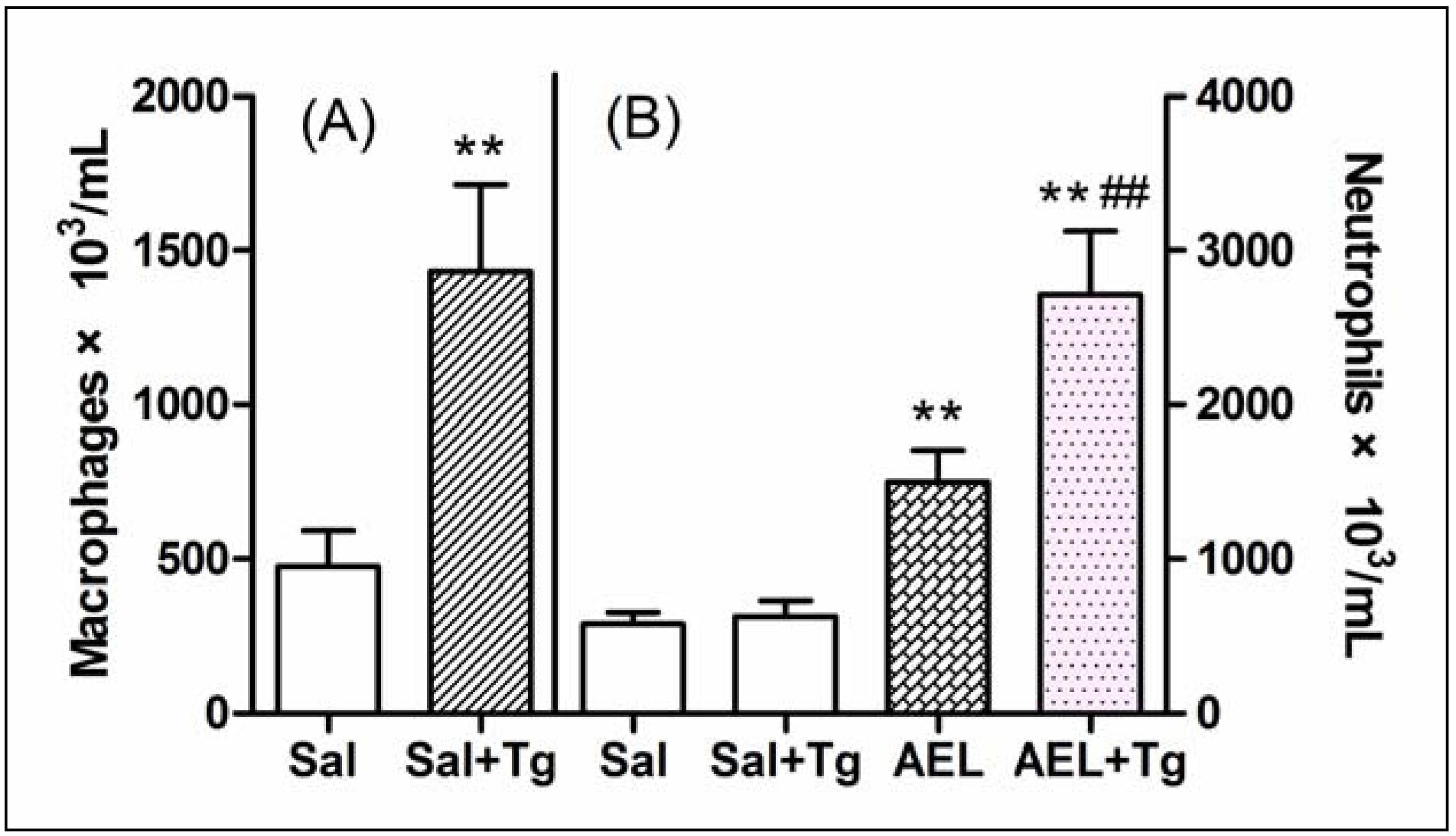
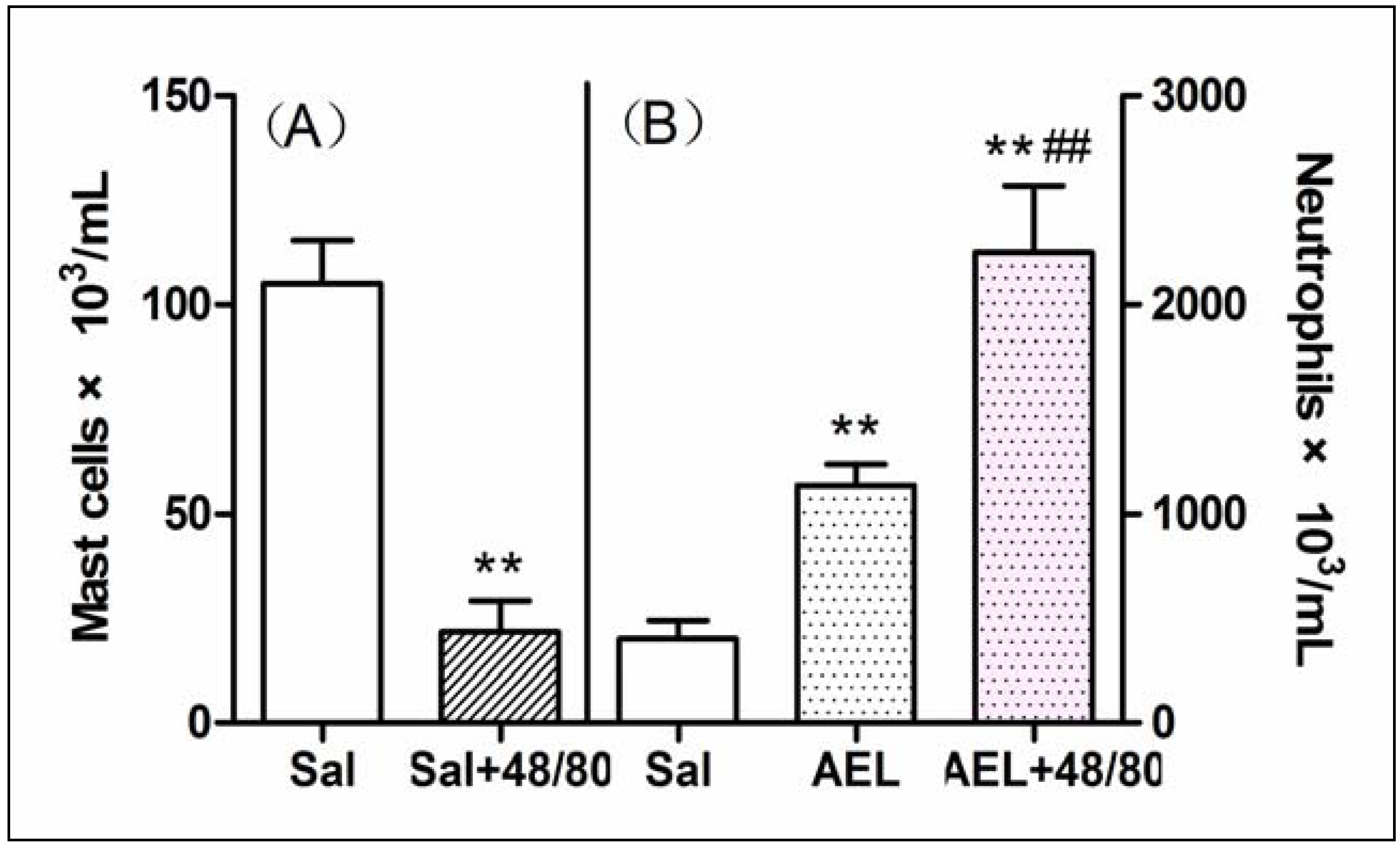
2.5. AEL-Induced Neutrophil Migration with Resident Cells



3. Experimental Section
3.1. Plant Material
3.2. Animals
3.3. Apparatus, Chemicals and Reagents
3.4. Extraction and Purification of AEL
3.5. SDS-PAGE and SEC-HPLC
3.6. Haemagglutination and Inhibition Assays
3.7. Thermal Stability
3.8. pH Stability
3.9. Rat Paw Edema
3.10. Stimulation of Neutrophil Migration into Peritoneal Cavities by AEL
3.11. Content of NO, PGE2, TNF-α Induced by AEL
3.12. Depletion of Total Resident Cell Population by Peritoneal Lavage
3.13. Increase of the Peritoneal Macrophage Population by Treatment with Tg
3.14. Depletion of Peritoneal Mast Cell Population by Chronic Treatment with Compound 48/80
3.15. Statistical Analysis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Jeelani, S.; Akbar Khuroo, M.; Razadan, T.K. New triterpenoids from Arisaema jacquemontii. J. Asian Nat. Prod. Res. 2010, 12, 157–161. [Google Scholar] [CrossRef]
- Shangary, S.; Singh, J.; Kamboj, S.S.; Kamboj, K.K.; Sandhu, R.S. Purification and properties of four monocot lectins from the family Araceae. Phytochemistry 1995, 40, 449–455. [Google Scholar] [CrossRef]
- Ge, X.Y.; Wu, H. Analysis of the composition of poisonous raphides in Araceae plant. Chin. J. Pharm. Anal. 2010, 30, 190–193. [Google Scholar]
- Wu, H.; Ge, X.Y.; Yu, H.L.; Chen, L. Comparisons of crystal form of raphides to toxicity raphides in four poisonous herbs of Araceae family. Chin. J. Chin. Mater. Med. 2010, 35, 1152–1155. [Google Scholar]
- Yu, H.L.; Zhu, F.G.; Wu, H. Study of toxic proteins on raphides from Pinellia ternata and Pinellia pedatisecta Schott. China J. Tradit. Chin. Med. Pharm. 2011, 26, 1037–1042. [Google Scholar]
- Liu, X.Q.; Wu, H.; Yu, H.L.; Pan, Y.Z. Irritability of Agglutinin from Pinellia ternata Breit, Pinellia pedtaisecta Schott and Typhonium giganteum Engl. Chin. J. Exp. Tradit. Med. Formulae 2011, 17, 244–247. [Google Scholar]
- Dhuna, V.; Bains, J.S.; Kamboj, S.S.; Singh, J.; Shanmugavel; Saxena, A.K. Purification and characterization of a lectin from Arisaema tortuosum schott having in-vitro anticancer activity against human cancer cell lines. J. Biochem. Mol. Biol. 2005, 38, 526–532. [Google Scholar] [CrossRef]
- Rubinstein, N.; Ilarregui, J.M.; Toscano, M.A.; Rabinovich, G.A. The role of galectins in the initiation, amplification and resolution of the inflammatory response. Tissue Antigens 2004, 64, 1–12. [Google Scholar] [CrossRef]
- Herre, J.; Willment, J.A.; Gordon, S.; Brown, G.D. The role of Dectin-1 in antifungal immunity. Crit. Rev. Immunol. 2004, 24, 193–203. [Google Scholar] [CrossRef]
- Balzarini, J.; Neyts, J.; Schols, D.; Hosoya, M.; van Damme, E.; Peumans, W.; de Clercq, E. The mannose-specific plant lectins from Cymbidium hybrid and Epipactis helleborine and the (N-acetylg1ucosamine)n-specific plant lectin from Urtica dioica are potent and selective inhibitors of human immunodeficiency virus and cytomegalovirus replication in vitro. Antivir. Res. 1992, 18, 191–207. [Google Scholar]
- Hilder, V.A.; Powell, K.S.; Gatehouse, A.M.R.; Gatehouse, J.A.; Gatehouse, L.N.; Shi, Y.; Hamilton, W.D.O.; Merryweathtrr, A.; Newell, C.; Timans, J.C.; et al. Expression of snowdrop lectin in transgenic tobacco plants results in added protection against aphids. Transgenic Res. 1995, 4, 18–25. [Google Scholar] [CrossRef]
- Alencar, N.M.N.; Assreuy, A.M.S.; Criddle, D.N.; Souza, E.P.; Soares, P.M.G.; Havt, A.; Aragao, K.S.; Bezerra, D.P.; Ribeiro, R.A.; Cavada, B.S. Vatairea Macrocarpa lectin induces paw edema with leukocyte infiltration. Protein Pept. Lett. 2004, 11, 195–200. [Google Scholar] [CrossRef]
- Alencar, N.M.N.; Assreuy, A.M.S.; Alencar, V.B.M.; Melo, S.C.; Ramos, M.V.; Cavada, B.S.; Cunha, F.Q.; Ribeiro, R.A. The galactose-binding lectin from Vatairea macrocarpa seeds induces in vivo neutrophil migration by indirect mechanism. Int. J. Biochem. Cell Biol. 2003, 35, 1674–1681. [Google Scholar] [CrossRef]
- Alencar, V.B.M.; Alencar, N.M.N.; Assreuy, A.M.S.; Mota, M.L.; Brito, G.A.C.; Aragao, K.S.; Bittencourt, F.S.; Pinto, V.P.T.; Debray, H.; Ribeiro, R.A.; et al. Pro-inflammatory effect of Arum maculatum lectin and role of resident cells. Int. J. Biochem. Cell Biol. 2005, 37, 1805–1814. [Google Scholar] [CrossRef]
- Alencar, V.B.M.; Assreuy, A.M.S.; Alencar, N.M.N.; Meireles, A.V.P.; Mota, M.R.L.; Aragao, K.S.; Cajazeiras, J.B.; Nagano, C.S.; Brito, G.A.C.; Silva, L.I.M.M.; et al. Lectin of Pisum arvense seeds induces in-vivo and in-vitro neutrophil migration. J. Pharm. Pharmacol. 2005, 57, 375–381. [Google Scholar]
- Lin, J.; Zhou, X.W.; Pang, Y.Z.; Gao, H.; Fei, J.; Shen, G.A.; Wang, J.; Li, X.S.; Sun, X.F.; Tang, K.X. Cloning and characterization of an agglutinin gene from Arisaema lobatum. Biosci. Rep. 2005, 25, 345–362. [Google Scholar] [CrossRef]
- Peng, H.; Lv, H.; Wang, Y.; Liu, Y.H.; Li, C.Y.; Meng, L.; Chen, F.; Bao, J.K. Clematis montana lectin, a novel mannose-binding lectin from traditional Chinese medicine with antiviral and apoptosis-inducing activities. Peptides 2009, 30, 1805–1815. [Google Scholar] [CrossRef]
- Bento, C.A.M.; Cavada, B.S.; Oliveira, J.T.A.; Moreira, R.A.; Barja-Fidalgo, C. Rat paw edema and leukocyte immigration induced by plant lectins. Agents Actions 1993, 38, 48–54. [Google Scholar] [CrossRef]
- Andrade, J.L.; Arruda, S.; Barbosa, T.; Paim, L.; Ramos, M.V.; Cavada, B.S.; Barral-Netto, M. Lectin-induced nitric oxide production. Cell. Immunol. 1999, 194, 98–102. [Google Scholar] [CrossRef]
- Higuchi, M.; Higashi, N.; Taki, H.; Osawa, T. Cytolytic mechanisms of activated macrophages, tumor necrosis factor and L-arginine-dependent mechanisms act synergistically as the major cytolytic mechanisms of activated macrophages. J. Immunol. 1990, 144, 1425–1431. [Google Scholar]
- Barral-Netto, M.; Barral, A. A simple method for assessing the binding of concanavalin A to mononuclear cell surfaces: NO interference of visceral leishmaniasis serum on this binding. Mem. Inst. Oswaldo Cruz 1986, 81, 343–345. [Google Scholar]
- Schultz, R.M.; Pavlidis, N.A.; Stylos, W.A.; Chirigos, M.A. Regulation of macrophage tumoricidal function: A role for prostaglandins of the E series. Science 1978, 202, 320–321. [Google Scholar]
- Locksley, R.M.; Killeen, N.; Lenardo, M.J. The TNF and TNF receptor superfamilies: Integrating mammalian biology. Cell 2001, 104, 487–501. [Google Scholar] [CrossRef]
- Ribeiro, R.A.; Flores, C.A.; Cunha, F.Q.; Ferreira, R.A. IL-8 in vivo neutrophil migration by a cell-dependent mechanism. Immunology 1991, 73, 472–477. [Google Scholar]
- Ribeiro, R.A.; Souza-Filho, M.V.P.; Souza, M.H.L.P.; Oliveira, S.H.P.; Costa, C.H.S.; Cunha, F.Q.; Ferreira, S.H. Role of resident mast cells and macrophages in the neutrophil migration induced by LTB4, fMLP and C5a des arg. Int. Arch. Allergy Immunol. 1997, 112, 27–35. [Google Scholar] [CrossRef]
- Rankin, J.A.; Sylvester, I.; Smith, S.; Yoshimura, T.; Leonard, E.J. Macrophages cultured in vitro release leukotriene B4 and neutrophil attractant activation protein (interleukin-8) sequentially in response to stimulation with lipopolysaccharide and zymosan. J. Clin. Invest. 1990, 80, 1554–1564. [Google Scholar]
- Whaley, K.; Fergusun, A. Molecular aspects of complement activation. Mol. Aspects Med. 1981, 4, 209–273. [Google Scholar] [CrossRef]
- Souza, G.E.P.; Ferreira, S.H. Blockade by anti-macrophages serum of the migration of PMN neutrophils into the inflamed peritoneal cavity. Agents Actions 1985, 17, 97–103. [Google Scholar] [CrossRef]
- Faccioli, L.H.; Souza, G.E.P.; Cunha, F.Q.; Poole, S.; Ferreira, S.H. Recombinant interleukin-1 and tumor necrosis factor induce neutrophil migration in vivo by indirect mechanisms. Agents Actions 1990, 30, 344–349. [Google Scholar] [CrossRef]
- Alencar, N.M.N.; Oliveira, R.S.B.; Figueiredo, J.G.; Cavalcante, I.J.M.; Matos, M.P.V.; Cunha, F.Q.; Nunes, J.V.S.; Bomfim, L.R.; Ramos, M.V. An anti-inflammatory lectin from Luetzelburgia auriculata seeds inhibits adhesion and rolling of leukocytes and modulates histamine and PGE2 action in acute inflammation models. Inflamm. Res. 2010, 59, 245–254. [Google Scholar] [CrossRef]
- Mota, M.R.L.; Criddle, D.N.; Alencar, N.M.N.; Gomes, R.C.; Meireles, A.V.P.; Santi-Gadekha, T.; Gadelha, C.A.A.; Oliveira, C.C.; Benevides, R.G.; Cavada, B.S.; et al. Modulation of acute inflammation by a chitin-binding lectin from Araucaria angustifolia seeds via mast cells. Naunyn Schmiedebergs Arch. Pharmakol. 2006, 374, 1–10. [Google Scholar] [CrossRef]
- Oda, Y.; Minami, K. Isolation and characterization of a lectin from tulip bulbs, Tulipa gesneriana. Eur. J. Biochem. 1986, 159, 239–245. [Google Scholar] [CrossRef]
- Wang, H.; Gao, J.; Ng, T.B. A new lectin with highly potent antihepatoma and antisarcoma activities from the oyster mushroom pleurotus ostreatus. Biochem. Biophys. Res. Commun. 2000, 275, 810–816. [Google Scholar] [CrossRef]
- Ferreira, S.H. A new method for measuring variations of rat paw volume. J. Pharm. Pharmacol. 1979, 31, 648. [Google Scholar] [CrossRef]
- Souza, G.E.P.; Cunha, F.Q.; Mello, C.R.; Ferreira, S.H. Neutrophil migration induced by inflammatory stimuli is reduced by macrophages depletion. Agents Actions 1988, 24, 377–380. [Google Scholar] [CrossRef]
- di Rosa, M.; Giround, J.P.; Willoughby, D.A. Studies of the mediators of the acute inflammatory response induced in rats in different sites by carrageenan and turpentine. J. Pathol. 1971, 104, 15–29. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are not available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, X.Q.; Wu, H.; Yu, H.L.; Zhao, T.F.; Pan, Y.Z.; Shi, R.J. Purification of a Lectin from Arisaema erubescens (Wall.) Schott and Its Pro-Inflammatory Effects. Molecules 2011, 16, 9480-9494. https://doi.org/10.3390/molecules16119480
Liu XQ, Wu H, Yu HL, Zhao TF, Pan YZ, Shi RJ. Purification of a Lectin from Arisaema erubescens (Wall.) Schott and Its Pro-Inflammatory Effects. Molecules. 2011; 16(11):9480-9494. https://doi.org/10.3390/molecules16119480
Chicago/Turabian StyleLiu, Xian Qiong, Hao Wu, Hong Li Yu, Teng Fei Zhao, Yao Zong Pan, and Run Jun Shi. 2011. "Purification of a Lectin from Arisaema erubescens (Wall.) Schott and Its Pro-Inflammatory Effects" Molecules 16, no. 11: 9480-9494. https://doi.org/10.3390/molecules16119480