
Lyn Kinase Structure, Regulation, and Involvement in Neurodegenerative Diseases: A Mini Review


Division of Clinical Pharmacology, School of Medicine, Indiana University, Indianapolis, IN 46202, USA
*
Author to whom correspondence should be addressed.
Kinases Phosphatases 2023, 1(1), 23-38; https://doi.org/10.3390/kinasesphosphatases1010004
Submission received: 20 December 2022
/
Revised: 9 January 2023
/
Accepted: 11 January 2023
/
Published: 23 January 2023
(This article belongs to the Special Issue Human Protein Kinases: Development of Small-Molecule Therapies)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:LYN proto-oncogene, Src family tyrosine kinase (Lyn) is a tyrosine kinase that belongs to the Src family (SFK). It is expressed as two isoforms in humans, LynA and LynB. Like other SFKs, Lyn consists of five protein domains, an N-terminal SH4 domain followed by a unique domain, the SH3 and SH2 domains, and a catalytic SH1 domain. The autophosphorylation of Tyr397 activates the protein, while the phosphorylation of the C-terminal inhibitory Tyr508 by C-terminal Src kinase (Csk) or Csk homologous kinase (Chk) inhibits the catalytic activity. The interaction of the SH2 domain with the phosphorylated Tyr508 stabilizes a compact, self-inhibited state. The interaction of the SH3 domain with a linker between the SH2 and catalytic domains further stabilizes this inactive conformation. The two critical structural features of the catalytic domain are a conserved DFG moiety and the αC helix, which can adopt in or out conformations. In the active state, both the DFG moiety and αC helix adopt in conformations, while in the inactive state, they adopt out conformations. Lyn has well-established functions in various hematopoietic cell types and more recent studies have revealed its roles in non-hematopoietic cells. At the molecular level, these functions are mainly exerted by phosphorylating specific tyrosine residues in immunoreceptor tyrosine-based inhibitory motifs (ITIMs) and immunoreceptor tyrosine-based activator motifs (ITAMs) associated with cell surface receptors. The phosphorylation of ITAMs by Lyn can initiate either activating or inhibitory (ITAMi) cell signaling depending on the receptor, targeting mode (crosslinking or monovalent targeting), and the cellular context. The phosphorylation of ITIMs by Lyn initiates inhibitory cell signaling via the recruitment of phosphatases to the ITIM-bearing receptor. The role of Lyn in cancer and autoimmune diseases has been extensively discussed in the literature. The involvement of Lyn in neurodegenerative diseases has been described more recently and, as such, it is now an emerging target for the treatment of neurodegenerative diseases.
Keywords:
Lyn; unique domain; SH3 domain; SH2 domain; ITAM; ITIM; neurodegenerative disease; Alzheimer’s disease1. Introduction
LYN proto-oncogene, Src family tyrosine kinase (Lyn) is a non-receptor protein tyrosine kinase belonging to the Src family of kinases (SFKs), which includes nine members: Src, Lck, Yes, Hck, Fyn, Fgr, Blk, and Yrk [1]. Lyn exists as two isoforms, LynA (p56) and LynB (p53), differing by 21 amino acids at the N-terminal end, which arise from the alternate splicing of exon-2 [2]. A functional difference between the two isoforms has not been reported. Lyn is expressed in the hematopoietic cells of both the myeloid and lymphoid lineages, except for T cells [2,3]. Lyn is also expressed in the brain, suggesting common signaling mechanisms in the immune and central nervous systems [4]. Lyn phosphorylates the specific tyrosine residues of many cytosolic and membrane-bound effector proteins that initiate activating and inhibitory cell signaling [5]. This dual activity is attributed to the phosphorylation of immunoreceptor tyrosine-based activator motifs (ITAMs) and the immunoreceptor tyrosine-based inhibitor motifs (ITIMs) of immune receptors and adaptor proteins. The role of Lyn in cancer and autoimmune diseases is well-established and has been extensively reviewed in the literature [6,7,8], while studies of its involvement in neurodegenerative diseases are more recent. Therefore, in this review, we present a general review of Lyn, focusing on its structure, regulation, and function in terms of the ITAM/ITIM phosphorylation, experimentally validated binding partners, and its involvement in neurodegenerative diseases.
2. Lyn Kinase Structure
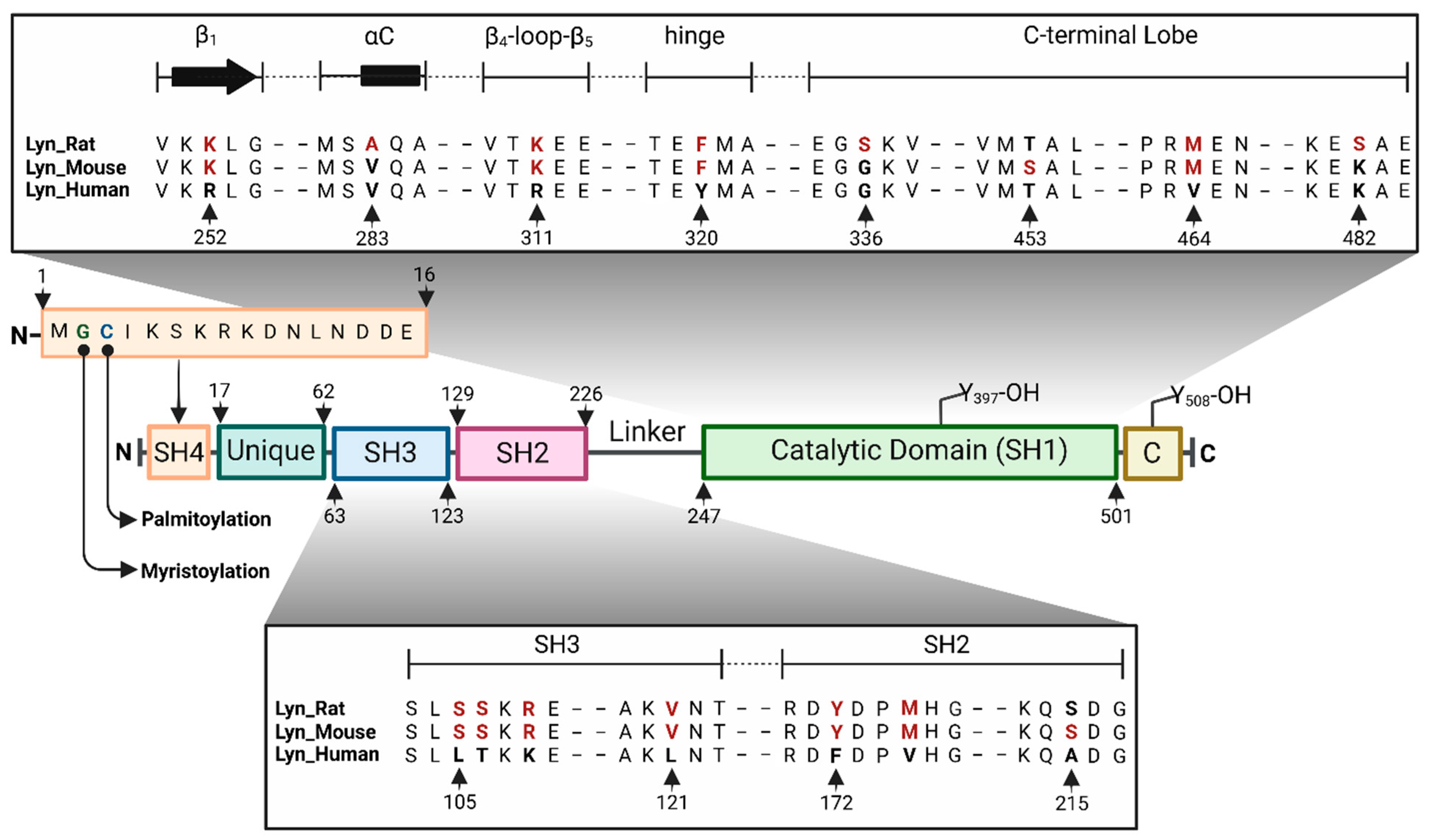
The human Lyn gene (LYN) encodes a 512 amino acid protein, consisting of 5 domains, an N-terminal SH4 domain followed by a unique domain, the SH3 and SH2 domains, and a catalytic (SH1) domain with an architecture similar to other SFKs (Figure 1) [9]. The beginning of the N-terminal SH4 domain has a specific amino acid sequence that can act as a membrane-anchoring site upon N-myristoylation and palmitoylation. The C terminus consists of a regulatory sequence, on which the phosphorylation of Tyr508 autoinhibits the kinase activity via interactions with the SH2 domain. The mouse and rat genes for Lyn each encode a 512 amino acid protein with a 96% identity to the human isoform, with most of the variations occurring outside the catalytic domain in regions of the SH4 and unique domains. The catalytic domain of mouse and rat kinase shows a 98% and 97.2% similarity to human Lyn, and the variations occur in residues outside the active site, except for the hinge region. In human Lyn, the position 320 of the hinge region is occupied by a tyrosine, whereas in the mouse and rat proteins, this residue is Phe (Figure 1). Therefore, the inhibitors that interact strongly with the human-Tyr320 via hydrogen bonding may be inactive when advanced to rodent models.
2.1. SH4 and Unique Domains
The N-terminal SH4 domain is a short 16-residue stretch of amino acids that consists of a specific Met-Gly-Cys sequence at the beginning, known as the myristate plus palmitate two-signal motif. Upon a dual acylation with myristate and palmitate, the SH4 domain can act as a membrane-anchoring site (Figure 1) [10,11]. During myristoylation, the initiating methionine is cotranslationally cleaved by methionine amino-peptidase, and the resulting glycine is coupled to myristate via an amide bond which is catalyzed by N-myristoyl transferase [12]. The myristoylation of glycine facilitates the palmitoylation of the adjacent cysteine, which is catalyzed by palmitoyl acyl transferase [13,14]. A mutational study of Lyn at the myristoylation and palmitoylation sites has revealed that N-terminal myristoylation is essential for anchoring to the plasma membrane, despite the palmitoylation state [15]. The same study found that N-myristoylation is a necessary prerequisite for palmitoylation, and the dual acylation of the SH4 domain is essential for its association with lipid rafts. Therefore, the SH4 domain plays a key role in membrane association and initiating or inhibiting immunoreceptor signaling via the phosphorylation of various membrane receptors or receptor subunits. The unique domain is an intrinsically disordered region that connects the SH3 domain to the membrane-anchoring SH4 domain. Due to its highly disordered nature, the unique domain is often missing from x-ray crystal structures. As the name implies, there is considerable variation in the sequence and length of this domain within the SFKs. The unique domain of LynB is 21 amino acids shorter than LynA, and this segment contains Tyr32, which is known to be phosphorylated by the Epidermal Growth Factor Receptor (EGFR) and potentiate the MCM7-mediated DNA replication [16].
2.2. SH3 Domain
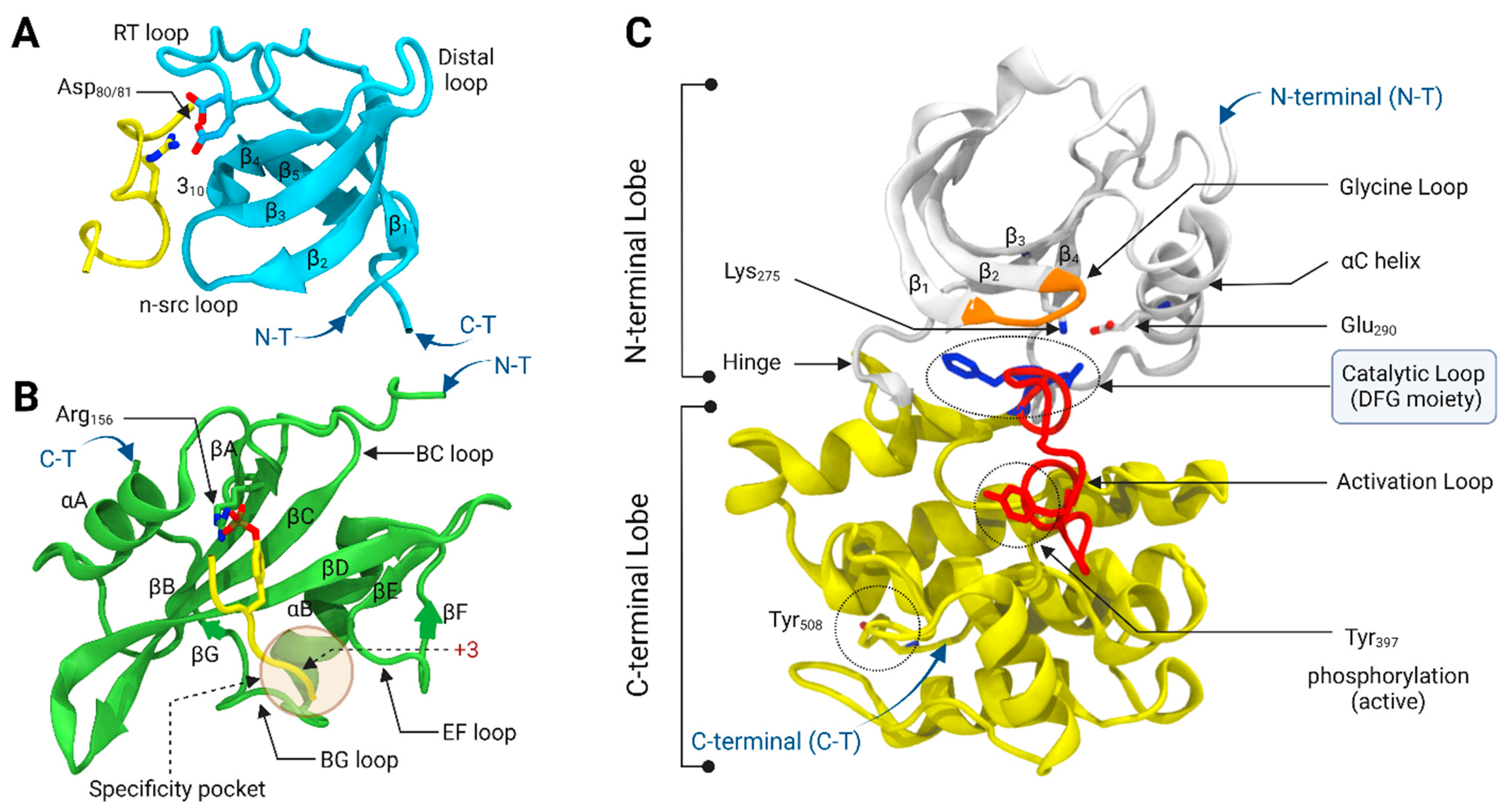
The Lyn SH3 domain is a 61 amino acid module that interacts with the proline-rich motifs of binding partners, such as Ras-GAP and the PI3K kinases [17,18,19]. It recognizes the PxxP motifs that form polyproline type II helixes. The structure of the Lyn SH3 domain consists of two β-sheets, one with two β-strands (β1,β5), and a second β-sheet with three β-strands (β2–β4) (Figure 2A). There are four main loops, namely RT, n-src, distal, and 310 helical loops, that connect the β1–β2, β2–β3, β3–β4, and β4–β5 strands, respectively. The polyproline motif binding site consists of two hydrophobic pockets and a negative charge specificity pocket, which resides between the RT and n-src loops [20]. The negative charge specificity pocket is made of two aspartate residues (Asp80 and Asp81) that facilitates the binding of the flanking basic residue of the ligand. One of the main functions of the SH3 domain is to maintain the inactive kinase conformation by intramolecularly binding to the polyproline type II sequence within the linker between the SH2 and the kinase domains [21,22]. In the active conformation, the SH3 domain interacts with the binding partners, leaving the kinase domain in an active conformation where it can phosphorylate substrates [23,24]. Eighteen cancer-associated mutations in the SH3 domain have been identified [25,26,27]. Some of these mutations, such as D18N and W99L, directly involve the binding site’s hydrophobic and negative charge pockets and are likely transformative by increasing the kinase activity through the elimination of self-inhibition.
2.3. SH2 Domain
The SH2 domain is a 98-residue modular protein structure that binds phosphotyrosine-containing peptide segments, which include the Tyr508 phosphorylated C-terminal regulatory sequence of Lyn itself [28,29]. The SH2 domain core structure mainly consists of a central five-standard antiparallel β-sheet flanked by two α-helices (Figure 2B) [30,31,32]. There are two three-residue irregular β-strands outside the five central β-strands, namely C-terminal βA and N-terminal βG strands, located before the N-terminal α-helix (αA) and after the C-terminal α-helix (αB), respectively. These two irregular β-strands align parallel to the first β-strand (βB) of the core antiparallel β-sheet and form a parallel β-sheet, which brings the N- and C-termini into close proximity. The central antiparallel β-sheet consists of five β-sheets (βB-βF) and several loops that connect the β-strands and α-helices (BC, BG, and EF loops). The conserved residues from three core β-strands (βB, βC, and βD), αA helix, and the phosphate-binding loop (BC loop) form the phosphotyrosine binding pocket. The Arg156 residue on the βC strand forms direct interactions with the phosphate group of the phosphotyrosine residue. The specificity of the phosphopeptide to the SH2 domain is determined by the three-residues C-terminal to the phosphorylated tyrosine residue. The C-terminal pY + 3 residue of the phosphotyrosine peptide segment occupies a deep specificity pocket that consists of the residues from the BG and EF loops. The SH2 domain regulates the kinase activity by binding to the phosphorylated Tyr508 of the C-terminal regulatory sequence, which maintains the kinase in an inactive conformation. Recently, it has been revealed that the phosphorylation of a conserved Tyr194 residue in the SH2 domain impacts the binding affinity and specificity of the SH2 domain with phosphotyrosine peptides [32].
2.4. Catalytic (SH1) Domain
The Lyn catalytic (kinase) domain is composed of 255 amino acid residues with a bilobal protein–tyrosine kinase fold that catalyzes the phosphorylation of the specific tyrosine residues of various immune receptors and effector proteins. The small N-terminal lobe consists of a five-stranded antiparallel β-sheet (β1–β5) and a large α-helix (αC) (Figure 2C). The first two α-helices (αA and αB) that are found in some protein kinases are absent in Lyn, similar to other SFKs. The large C-terminal lobe is predominantly helical with two-stranded antiparallel β-sheets. The ATP-binding pocket is located in a deep cleft between the two lobes, connected by a loop that forms the hinge region. The adenine ring of ATP forms hydrogen bonding interactions with the hinge region. The loop that connects the β1 and β2 strands is known as the Glycine loop (or phosphate binding P-loop) that interacts with the non-transferable ATP α/β-phosphates and facilitates the transfer of the γ-phosphoryl group to the substrates [33]. The activation loop in the C-terminal lobe regulates the catalytic activity via the autophosphorylation of the specific Tyr397 residue, which correlates with movement in the loop and the αC helix. In addition, the phosphorylation of Tyr508 at the C-terminal end regulates the catalytic activity.
The two key structural features that define the active and inactive conformations of the kinase domain are a conserved Asp-Phe-Gly (DFG) moiety (Asp385, Phe386, Gly387 in Lyn) located at the beginning of the activation loop and the αC helix, where both can take either “in” or “out” orientations (DFG-in/DFG-out and αC-in/αC-out). The active kinase domain adopts the DFG-in/αC-in conformation and is targeted by ATP-competitive type I inhibitors. In the αC-in conformation, there is a specific salt bridge interaction between the conserved Lys275 residue of the β3 strand and Glu290 of the αC helix, which is essential for the catalytic activity of the enzyme and contributes to the coordination of the ATP phosphate groups (Figure 3A). The Asp residue of the DFG moiety also coordinates the catalytically important Mg2+ ion in the DFG-in conformation. The inactive kinase domain can either be in the DFG-in/αC-out or DFG-out/αC-in conformations, which are targeted by type I1/2 and type II inhibitors, respectively. In the inactive αC-out conformation, the αC helix moves outwards and rotates a conserved Glu290 away from the ATP-binding site, which causes the specific salt bridge to be broken. This opens up a binding pocket close to the αC helix that can be occupied by the tail end of type I1/2 inhibitors, which helps to achieve specificity (Figure 3B). Similarly, the outwards movement of the DFG moiety creates a new allosteric pocket that can be occupied by the tail group of type II inhibitors, allowing type II inhibitors to achieve a better selectivity than type I inhibitors (Figure 3C).
As of 2022, only six x-ray crystal structures of the Lyn catalytic domain are available in the protein data bank, and all of them are in the DFG-in/αC-in active conformation: four mouse-derived (2ZVA, 2ZV9, 2ZV8, and 2ZV7) and two human-derived (3A4O and 5XY1). The lack of available Lyn kinase X-ray crystal structures in inactive conformations (DFG-in/αC-out and DFG-out/αC-in) presents a challenge for the structure-based design of novel type I1/2 and II specific inhibitors. Therefore, there is a pressing need to resolve Lyn catalytic domain X-ray crystal structures with known type I1/2 and II inhibitors.
3. Regulation of the Lyn Kinase Activity
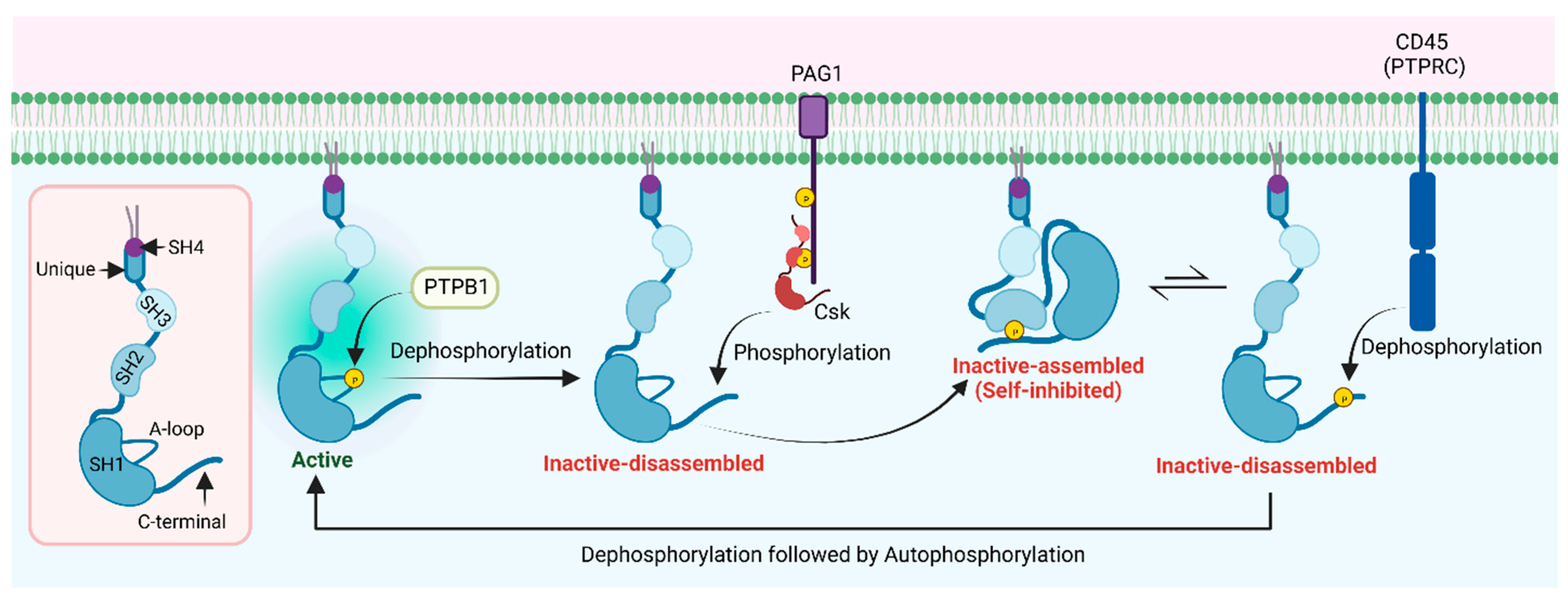
The regulation of the Lyn kinase activity is similar to other SFKs. The phosphorylation states of both the activation loop Tyr397 (the positive regulatory tyrosine) and the C-terminal Tyr508 (the negative regulatory tyrosine) regulate the kinase activity of Lyn. The activation loop Tyr397 undergoes an autophosphorylation, while the C-terminal Tyr508 is phosphorylated by the C-terminal Src kinase (Csk) or Csk homologous kinase (Chk) [34]. The phosphorylation of Tyr397 stabilizes the activation loop and increases the catalytic activity, while phosphorylated Tyr508 interacts with the phosphotyrosine binding site of the SH2 domain, which stabilizes the auto-inhibited inactive conformation of the catalytic domain (Figure 4). In fact, a Y508F mutation of Lyn has been reported in pediatric patients presenting with an early onset autoinflammatory phenotype, suggesting that an over-activation is causative [35]. As described previously, the inactive conformation is further stabilized by the interactions of the SH3 domain with the linker region that connects the SH2 and catalytic domains. These interactions promote an αC-out conformation of the kinase domain, which disrupts a salt bridge interaction between the conserved Lys275 (β3 strand) and Glu290 (αC), which is essential for the proper orientation of ATP inside the binding pocket. It has been shown that the activation loop phosphorylation of SFKs blocks their inactivation by Csk. The same study demonstrated that Csk can still phosphorylate the activation loop of autophosphorylated SFKs at the C-terminal tyrosine (doubly phosphorylated SFK) [36]. However, the phosphorylated activation loop prevents interactions between the SH2 domain and the phosphorylated C-terminal tyrosine residue, which helps maintain an active kinase conformation despite the phosphorylation of Tyr508. Therefore, the dephosphorylation of the activation loop by tyrosine phosphatases, such as protein tyrosine phosphatase 1B (PTP1B), allows SFKs to be inactivated by Csk.
The structure of Csk consists of SH3 and SH2 and catalytic domains similar to SFKs. However, it does not possess an SH4 domain for an acylation and membrane association, thus it is preferentially located in the cytosol [37]. Membrane-anchoring proteins such as Cbp/PAG1 (Csk binding protein/phosphoprotein associated with glycosphingolipid-enriched membrane) facilitate the translocation of Csk to the plasma membrane, where it can phosphorylate membrane-bound Lyn, promoting the inactive assembled conformation (Figure 4). The inactive assembled conformation is in equilibrium with the inactive disassembled (phosphorylated Tyr508 free from the SH2 domain), where the inactive assembled form is more favorable. This inactive disassembled conformation facilitates the dephosphorylation of the Tyr508 by transmembrane phosphatases such as CD45, which promote the active conformation of the catalytic domain after the autophosphorylation of Tyr397 at the activation loop [38].
4. Lyn Kinase Function-Itim/Itam Phosphorylation
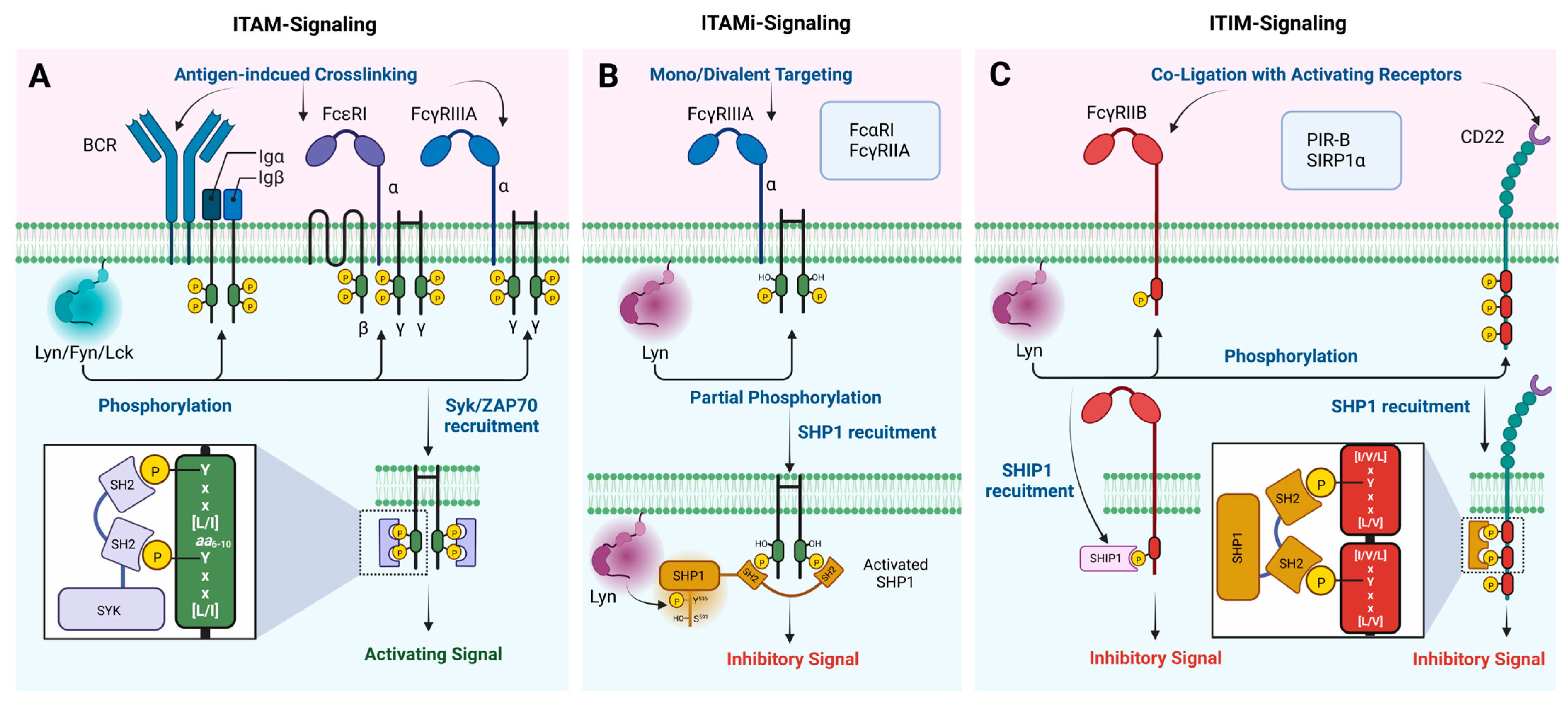
Lyn has well-established functions in various hematopoietic cell types and non-hematopoietic cells in different areas such as the brain, colon, breast, and prostate [39,40,41,42,43,44]. At the molecular level, these functions are mainly exerted by phosphorylating specific tyrosine residues in the ITIMs and ITAMs associated with various immune receptors and non-immune receptors such as B-cell receptors (BCR), T-cell receptors (TCR), Fc receptors (FcR), Sialic acid-binding immunoglobulin-type lectins (Siglecs/CD) receptors, paired immunoglobulin (Ig)-like receptor B (PIR-B), and the signal regulatory protein-1α (SIRP1α) (Figure 5). Lyn is the first known SFK member with a dual role acting as both a positive and negative regulator of cell signaling [9,45]. The negative regulation of the cell signaling occurs when Lyn phosphorylates specific tyrosine residues within the ITIMs present in inhibitory receptors. In contrast, a positive regulation occurs when Lyn phosphorylates ITAMs in activating receptors. Therefore, this section will mainly discuss the involvement of Lyn in the phosphorylation of ITIM- and ITAM-bearing immune receptors and the unique role played by Lyn in the negative regulation of cell signaling via both ITIM and ITAM phosphorylation.
4.1. Phosphorylation of ITIM Motifs
Several inhibitory plasma membrane receptors with ITIMs are phosphorylated by Lyn, including FcγRIIB, CD22, PIR-B, and SIRPα in various cell types. A single consensus sequence [I/V/L]xYxx[L/V] defines the ITIM motif, where x is any amino acid and is generally present in the receptor’s cytoplasmic tail or associated subunits [46]. The inhibitory effect of these ITIM-bearing receptors requires a co-ligation with their heterologous activating receptors by immune complexes, which facilitates the phosphorylation of ITIMs by Lyn (Figure 5C) [47]. Therefore, a negative regulation is context dependent only when cells are activated, and it does not occur in resting cells. The phosphorylated ITIMs act as docking sites for phosphatases that bear SH2 domains, such as Src homology-2 domain-containing phosphatases (SHP-1 and SHP-2) and the inositol polyphosphate 5-phosphatases (SHIP-1 and SHIP-2). The recruitment of these phosphatases to the plasma membrane dephosphorylates various effector proteins and phosphatidylinositol substrates, respectively, which modulates cell signaling.
FcγRIIB is an inhibitory receptor expressed on the surface of immune cells that binds the Fc region of the IgG antibodies. It is the only Fcγ receptor expressed by B cells and negatively regulates the activation via a co-ligation with BCR [48]. Other Fcγ receptors, such as high-affinity FcγRI and low-affinity FcγRIIA, FcγRIIC, FcγRIIIA, and FcγRIIIB, are activating. Upon a co-ligation with BCR, FcγRIIB is mono-phosphorylated by Lyn, which creates a docking site for SHIP-1 and SHIP-2 phosphatases, which contain a single SH2 domain (Figure 6) [49]. FcγRIIB is the only reported ITIM-containing receptor that recruits SHIP-1 and SHIP-2 upon an ITIM phosphorylation, and this unique affinity depends on a specific amino acid residue at the Y + 2 position in the ITIM [50]. Once recruited and brought close to the membrane, SHIP1 hydrolyzes the 5-phosphate of PI(3,4,5)P3 to give PI(3,4)P2 [51]. This prevents the PI(3,4,5)P3-dependent recruitment of pleckstrin homology (PH) domain-containing kinases and phospholipases to the inner leaflet of the plasma membrane, such as the protein kinases Atk and Btk and the phospholipases PLCγ1 and PLCγ2, which in turn promotes their phosphorylation and activation (Figure 6). The inactivation of PLCγ inhibits the generation of inositol 1,4,5-trisphosphate (IP3) and DAG from PI(4,5)P2 via lipid hydrolysis, which results in the inhibition of calcium mobilization and B cell activation (Figure 6).
CD22 (siglec 2) is another inhibitory transmembrane glycoprotein receptor exclusively expressed by B cells that inhibits the B cell activation upon a co-ligation with activated BCR [52,53]. It has three ITIMs and six tyrosine residues in the cytoplasmic domain. A co-ligation with BCR leads to the phosphorylation of these ITIMs by Lyn [54,55]. The phosphorylated ITIMs recruit SHP-1, a tyrosine phosphatase that dephosphorylates various effector proteins, which leads to the inhibition of the calcium mobilization and B cell activation [56].
PIR-B is one of the inhibitory receptors identified on murine B cells and myeloid-lineage cells (not on T cells and natural killer (NK) cells), with three ITIMs in the cytoplasmic domain [57]. PIR-B is an orthologue of human leucocyte immunoglobulin-like receptor B (LILR-B), an inhibitory receptor expressed in human B cells and myeloid cells [57,58]. PIR-A is the activating co-receptor and is expressed in a pair-wise fashion with PIR-B. The phosphorylation of the ITIMs of PIR-B by Lyn recruits SHP-1 and SHP-2 tyrosine phosphatases, leading to the inhibition of the B cell activation [59]. Similar to PIR-B, the human orthologue LILR-B is also phosphorylated by Lyn [60].
In addition to the above inhibitory receptors, Lyn is known to phosphorylate the ITIMs of SIRP1α, a glycoprotein receptor expressed by myeloid and neuronal cells [61,62]. The receptor has four ITIMs, and the phosphorylation recruits SHP1/2, inhibiting the cell activation via the dephosphorylation of downstream effectors. In macrophages, this prevents phagocytosis towards healthy host cells [63].
4.2. Phosphorylation of ITAM Motifs
Lyn also phosphorylates the ITAMs associated with activating immune receptors, and similar to ITIMs, an ITAM is defined by two consecutive Yxx[L/I] sequences separated by 6 to 10 amino acids [64]. In Fcγ receptors, such as FcγRI and FcγRIIIA, an ITAM can be found in the γ subunit [65]. In some cases, ITAMs can be found in the adaptor molecules associated with activating receptors, such as CD3 (T-cell receptor), Igα/Igβ (BCR), and DAP12 [66]. The phosphorylation of ITAMs will recruit SH2 domain-bearing tyrosine kinases Syk or ZAP70, which have two SH2 domains and phosphorylate multiple stimulatory effector proteins, ultimately leading to an intracellular Ca2+ mobilization and cell activation (Figure 5A).
In B-cells, Lyn is known to phosphorylate the ITAMs of the Igα/β subunits of BCR upon the antigen-induced crosslinking of the receptor. The phosphorylated ITAMs act as docking sites for tyrosine kinase Syk, which phosphorylates downstream protein kinases such as PI3K and Btk, leading to the activation of the PLCγ and intracellular Ca2+ release (Figure 6). Interestingly, the role of Lyn in initiating BCR activation signaling is non-essential and can be replaced by other SFKs, such as Fyn and Lck [67].
Lyn is reported to associate with tetrameric FcεRI receptors in mast cells, which consist of α, β, and homodimeric γ subunits. Upon the antigen-induced crosslinking of the FcεRI receptors, Lyn phosphorylates the ITAMs associated with the β and γ subunits (three ITAMs), providing docking sites for Syk, which initiates a mast cell activation. In addition, FcγRIIIA with an ITAM-bearing γ subunit similar to FcεRI is known to phosphorylate by Lyn in mast cells [68].
Recently, it has been shown that upon the mono or divalent targeting of low-avidity activating ITAM-bearing FcRs, such as FcαRI, FcγRIIA, and FcγRIIIA, ITAM-mediated inhibitory (ITAMi) signaling can be induced without co-ligating to an inhibitory receptor [69]. The bi-functional activity of these immune receptors modulates susceptibility to autoimmune and inflammatory diseases [70,71]. Other immune receptors, such as BCR and TCR, can also initiate ITAMi signaling [72,73]. Lyn plays an essential role in ITAMi signaling via a partial phosphorylation (single Tyr phosphorylation of ITAM) of the ITAM and the simultaneous phosphorylation of SHP-1 at Tyr536, which activates and recruits SHP1 to ITAM instead of Syk or ZAP70 (Figure 5B) [5]. The same receptors can induce activating ITAM signaling upon the multivalent crosslinking double phosphorylation of the ITAM by Fyn and the recruitment of Syk. Interestingly, Fyn silencing by siRNA reversed the activating ITAM to an ITAMi signaling, despite the receptors’ multivalent crosslinking, demonstrating the importance of Fyn and Lyn in ITAM switching [5]. Overall, these findings suggest the vital role played by Lyn in inducing negative signaling via the phosphorylation of both ITIM and ITAM motifs, while the role of Lyn in positive signaling via the phosphorylation of the ITAM is redundant by other Src kinases.
5. Lyn Kinase Binding Partners
To date, several binding partners of Lyn have been identified and experimentally validated. Most of these binding partners are scaffold, anchoring, or adaptor proteins with no enzyme activity. They recruit Lyn into multiprotein signaling networks or places Lyn close to its substrates. Lyn is mainly associated with these binding partners via SH3-phosphotyrosine or SH3-proline interactions. Src homology and collagen (Sch) proteins are adaptor proteins known to directly interact with phosphorylated Lyn in stimulated neutrophils [4]. The Shc proteins consist of the N-terminal phosphotyrosine binding (PTB) domain, a linker collagen homology 1 (CH1) domain, and a carboxy-terminal SH2 domain [74]. In stimulated neutrophils, the SH2 domain of the Shc protein interacts with the phosphorylated Lyn, ultimately leading to the phosphorylation of the Shc protein by Lyn [75]. The active Lyn is also known to interact with RING-type E3 ubiquitin-protein ligase c-Cbl and Cbl-b in rat basophilic leukemia RBL-2H3 cells upon the stimulation of FcεRI [76]. The association is mainly via the SH3 domain of Lyn and the proline-rich regions of c-Cbl, which leads to the phosphorylation of c-Cbl by Lyn. The phosphorylated c-Cbl mediates ubiquitination and the subsequent degradation of Lyn in the proteasome, causing the down-regulation of Lyn. In addition to the above adaptor proteins, felic and β-arrestins were also reported in the literature as the direct binding partners of Lyn. Felic is a novel scaffolding protein identified in a yeast 2-hybrid screen, which directly interacts with the SH3 domain of Lyn via polyproline motifs, and is known to be important in the phagocytosis of macrophages [77]. β-arrestins, traditionally known to play a crucial role in terminating GPCR-mediated signals, have been shown to interact directly with Lyn, in response to the CCR5s activation in human primary macrophages, revealing its novel function as a scaffold protein [78]. Apart from scaffolding proteins, Lyn is reported to have enzymes as binding partners. Cyclic AMP (cAMP)-specific phosphodiesterase-4 (PDE4) is one such enzyme that directly interacts with the SH3 domain of Lyn via its multifunctional docking site on the catalytic unit [79]. In addition, the SH3 domain of Lyn is also known to directly interact with SHP-1 (PTPN6) and phosphorylates its C-terminal Tyr564 [80].
6. Lyn Kinase in Neurodegenerative Diseases
Even though Lyn is expressed in the brain, information regarding its function in the CNS is limited [4]. In cortical neurons, Lyn is known to phosphorylate the α3 subunit of Na+/K+ ATPase and upregulates its activity [81]. Lyn was also shown to participate in the AMPA receptor-mediated upregulation of the brain-derived neurotrophic factor (BDNF) via the activation of the mitogen-activated protein kinase pathway [82]. In addition, a negative regulatory role of Lyn has also been proposed related to the NMDA receptor’s activity [40].
Because of the critical function exerted by Lyn in numerous cell types, it is involved in various disease conditions. The role of Lyn in diverse types of cancers, such as acute myeloid leukemia, colorectal, breast, renal, prostate, ovarian, and liver cancer, are reported and have been reviewed [7,25,26,27,83,84,85,86,87]. The role of Lyn in autoimmune diseases has also been thoroughly reviewed [7]. The involvement of Lyn in neurodegenerative diseases has been reported more recently with studies that focus on the molecular mechanisms underlying the involvement of Lyn in neurodegeneration.
Alzheimer’s disease (AD) is one of the main neurodegenerative diseases where the role of Lyn has been evaluated. Microglia are the resident phagocytes of the central nervous system [88]. Genome-wide association studies (GWAS), whole genome sequencing, and gene-expression network analyses have identified numerous AD risks and protective variants in genes such as TREM2, CD33, APOE, ABCA7, PLCG2, and INPP5D, which are essential to the microglia function [89]. Comprehensive single-cell RNA sequencing (RNA-seq) and the analysis of AD and the aging brain have identified a rare subset, disease-associated microglia (DAMs), which are conserved in mice and humans [90]. Lyn is highly enriched in the microglia from mouse models upon an exposure to amyloid beta (Aβ) [91]. The deposition of Aβ is a major pathological hallmark of AD [92,93]. The expression of Lyn is high in human and mouse microglia compared to other SFKs, as revealed by RNA sequencing databases [94]. Additionally, an increase in the Lyn expression was observed in microglia isolated from the post-mortem brains of AD patients [95]. Interestingly, Combs et al. reported that the stimulation of primary microglia and THP-1 cells with Aβ-fibrils does not induce the activation of Lyn, only the activation of c-Src. The inhibitor PPI, which targets Lyn, blocked the Aβ-stimulated production of neurotoxins and promoted neuronal survival [95]. PPI is a broad-spectrum Src kinase inhibitor with inhibitory potencies IC50 values of 6 and 5 nM for Fyn and Lck, respectively. The inhibition of Fyn would have a significant effect on the microglia’s activation [96]. Several other independent studies have reported an increased level of Lyn activation in mouse models microglia upon an exposure to Aβ oligomers, similar to the microglia isolated from the post-mortem brains of AD patients [97,98,99].
A comprehensive study conducted by Gwon et al. focused on a mode of action of Lyn in AD that has revealed an important role played for Lyn in Aβ-triggered neurotoxicity and tau hyperphosphorylation via FcγRIIb2 phosphorylation [100]. The transcriptional variation 2 of FcγRIIb (FcγRIIb2), which is mostly expressed in neurons, plays a critical role in the internalization of oligomeric Aβ1–42 [101]. This study also reported an increase in the Lyn activity as measured by autophosphorylation at the Tyr397 residue upon an exposure to oligomeric Aβ1–42 in mouse neuronal cells. The activation of Lyn was rapid (as early as 15 min), reaching ten-fold after 1 h of oligomeric Aβ1–42 stimulation, which lasted longer than Fyn. It was also found that the activation of Lyn was blocked in FcγRIIb2 knockout neurons, indicating that the Lyn’s activation was mediated by FcγRIIb2-Aβ1–42 interactions. Furthermore, the analysis of AD patients’ brain tissue samples also revealed a three-fold increase in the autophosphorylation of Lyn at Tyr397 in the hippocampus among AD patients compared to non-AD controls. Additional experiments in SH-SY5Y cells confirmed the direct phosphorylation of the FcγRIIb2 ITIM at Tyr273 by Lyn upon an exposure to oligomeric Aβ1–42. Interestingly, the knockdown of the expression of Lyn in B103 rat neuroblastoma and HT22 mouse hippocampal cell lines significantly suppressed the oligomeric Aβ1–42-induced cell death, revealing the importance of Lyn in attenuating Aβ1–42-induced neurotoxicity. Additional experiments revealed the recruitment of SHIP2 to FcγRIIb2 upon a phosphorylation by Lyn, similar to B cells (the mechanism is explained in detail in a previous section) which ultimately led to tau-hyperphosphorylation. Finally, using a novel, potent Lyn inhibitor (KICG2576), the researchers demonstrated that the inhibition of the Lyn’s activity can attenuate the Aβ-FcγRIIb2-mediated neuronal cell death and rescue the Aβ-triggered memory impairment in mice. However, the study also highlighted the potential broad spectrum Src inhibitor activity associated with KICG2576 and the importance of developing novel, potent Lyn selective kinase inhibitors to further evaluate the role of Lyn in AD.
Another study conducted by Peikert et al. describes the association of Lyn with Chorea-Acanthocytosis (ChAc), an ultra-rare neurodegenerative disease with 1000–5000 cases worldwide [102]. The disease is caused by loss-of-function mutations in the vacuolar protein sorting 13 homolog A (VPS13A) gene. The study has identified the accumulation of activated Lyn as a key pathophysiological event in ChAc. Furthermore, the inhibition of Lyn by Nilotinib in a Vsp13a−/− mouse model improved autophagy and prevented neuroinflammation. These literature findings suggest Lyn as a potential target for treating various neurodegenerative diseases.
7. Concluding Remarks
Lyn has been shown to have the unique ability to phosphorylate the ITIM and ITAMs associated with different immune receptors and adaptor proteins, distinguishing it from other Src kinases. This makes Lyn a key SFK in both activating and inhibitory cell signaling pathways. There is a tight regulation of the Lyn activity in cells, where dysregulation is implicated in cancer and autoimmune diseases. Recent studies have described the role for Lyn in neurodegenerative diseases, especially in AD. However, a further evaluation of its distinct mechanism is limited by the lack of Lyn-specific potent kinase inhibitors. The finite number of X-ray crystal structures of Lyn available in the protein data bank that is limited to an active conformation exerts a major challenge for structure-based drug discovery campaigns that focused on delivering Lyn-specific kinase inhibitors.
Author Contributions
Conceptualization, literature review and writing, P.M.W.; review and editing, T.I.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the National Institute of Aging via U54AG065181.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Brown, M.T.; Cooper, J.A. Regulation, Substrates and Functions of Src. Biochim. Biophys. Acta Rev. Cancer 1996, 1287, 121–149. [Google Scholar] [CrossRef] [PubMed]
- Yi, T.L.; Bolen, J.B.; Ihle, J.N. Hematopoietic Cells Express Two Forms of Lyn Kinase Differing by 21 Amino Acids in the Amino Terminus. Mol. Cell. Biol. 1991, 11, 2391–2398. [Google Scholar] [CrossRef]
- Yamanashi, Y.; Mori, S.; Yoshida, M.; Kishimoto, T.; Inoue, K.; Yamamoto, T.; Toyoshima, K. Selective Expression of a Protein-Tyrosine Kinase, P56lyn, in Hematopoietic Cells and Association with Production of Human T-Cell Lymphotropic Virus Type I. Proc. Natl. Acad. Sci. USA 1989, 86, 6538–6542. [Google Scholar] [CrossRef] [Green Version]
- Umemori, H.; Wanaka, A.; Kato, H.; Takeuchi, M.; Tohyama, M.; Yamamoto, T. Specific Expressions of Fyn and Lyn, Lymphocyte Antigen Receptor-Associated Tyrosine Kinases, in the Central Nervous System. Mol. Brain Res. 1992, 16, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Mkaddem, S.B.; Murua, A.; Flament, H.; Titeca-Beauport, D.; Bounaix, C.; Danelli, L.; Launay, P.; Benhamou, M.; Blank, U.; Daugas, E.; et al. Lyn and Fyn Function as Molecular Switches That Control Immunoreceptors to Direct Homeostasis or Inflammation. Nat. Commun. 2017, 8, 246. [Google Scholar] [CrossRef] [Green Version]
- Brodie, E.J.; Infantino, S.; Low, M.S.Y.; Tarlinton, D.M. Lyn, Lupus, and (B) Lymphocytes, a Lesson on the Critical Balance of Kinase Signaling in Immunity. Front. Immunol. 2018, 9, 401. [Google Scholar] [CrossRef] [Green Version]
- Ingley, E. Functions of the Lyn Tyrosine Kinase in Health and Disease. Cell Commun. Signal. 2012, 10, 21. [Google Scholar] [CrossRef] [Green Version]
- Su, R.; Zhang, J. Oncogenic Role of LYN in Human Gastric Cancer via the Wnt/Β-catenin and AKT/MTOR Pathways. Exp. Ther. Med. 2020, 20, 646–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowell, C. Src-Family Kinases: Rheostats of Immune Cell Signaling. Mol. Immunol. 2004, 41, 631–643. [Google Scholar] [CrossRef]
- Resh, M.D. Regulation of Cellular Signalling by Fatty Acid Acylation and Prenylation of Signal Transduction Proteins. Cell Signal. 1996, 8, 403–412. [Google Scholar] [CrossRef]
- Rawat, A.; Harishchandran, A.; Nagaraj, R. Fatty Acyl Chain-Dependent but Charge-Independent Association of the SH4 Domain of Lck with Lipid Membranes. J. Biosci. 2013, 38, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Rudnick, D.A.; McWherter, C.A.; Adams, S.P.; Ropson, I.J.; Duronio, R.J.; Gordon, J.I. Structural and Functional Studies of Saccharomyces Cerevisiae Myristoyl-CoA:Protein N-Myristoyltransferase Produced in Escherichia Coli. Evidence for an Acyl-Enzyme Intermediate. J. Biol. Chem. 1990, 265, 13370–13378. [Google Scholar] [CrossRef] [PubMed]
- Berthiaume, L.; Resh, M.D. Biochemical Characterization of a Palmitoyl Acyltransferase Activity That Palmitoylates Myristoylated Proteins. J. Biol. Chem. 1995, 270, 22399–22405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunphy, J.T.; Greentree, W.K.; Manahan, C.L.; Linder, M.E. G-Protein Palmitoyltransferase Activity Is Enriched in Plasma Membranes. J. Biol. Chem. 1996, 271, 7154–7159. [Google Scholar] [CrossRef] [Green Version]
- Kovářová, M.; Tolar, P.; Arudchandran, R.; Dráberová, L.; Rivera, J.; Dráber, P. Structure-Function Analysis of Lyn Kinase Association with Lipid Rafts and Initiation of Early Signaling Events after Fcɛ Receptor I Aggregation. Mol. Cell. Biol. 2001, 21, 8318–8328. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.-H.; Huo, L.; Wang, Y.-N.; Xia, W.; Wei, Y.; Chang, S.-S.; Chang, W.-C.; Fang, Y.-F.; Chen, C.-T.; Lang, J.-Y.; et al. Epidermal Growth Factor Receptor Potentiates MCM7-Mediated DNA Replication through Tyrosine Phosphorylation of Lyn Kinase in Human Cancers. Cancer Cell 2013, 23, 796–810. [Google Scholar] [CrossRef] [Green Version]
- Toubiana, J.; Rossi, A.-L.; Belaidouni, N.; Grimaldi, D.; Pene, F.; Chafey, P.; Comba, B.; Camoin, L.; Bismuth, G.; Claessens, Y.-E.; et al. Src-Family-Tyrosine Kinase Lyn Is Critical for TLR2-Mediated NF-ΚB Activation through the PI 3-Kinase Signaling Pathway. Innate Immun. 2015, 21, 685–697. [Google Scholar] [CrossRef] [Green Version]
- Briggs, S.D.; Bryant, S.S.; Jove, R.; Sanderson, S.D.; Smithgall, T.E. The Ras GTPase-Activating Protein (GAP) Is an SH3 Domain-Binding Protein and Substrate for the Src-Related Tyrosine Kinase, Hck. J. Biol. Chem. 1995, 270, 14718–14724. [Google Scholar] [CrossRef] [Green Version]
- Songyang, Z. Recognition and Regulation of Primary-Sequence Motifs by Signaling Modular Domains. Prog. Biophys. Mol. Biol. 1999, 71, 359–372. [Google Scholar] [CrossRef]
- Lim, W.A.; Richards, F.M.; Fox, R.O. Structural Determinants of Peptide-Binding Orientation and of Sequence Specificity in SH3 Domains. Nature 1994, 372, 375–379. [Google Scholar] [CrossRef]
- Sicheri, F.; Kuriyan, J. Structures of Src-Family Tyrosine Kinases. Curr. Opin. Struct. Biol. 1997, 7, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.K. The Y’s That Bind: Negative Regulators of Src Family Kinase Activity in Platelets. J. Thromb. Haemost. 2009, 7, 195–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, M.A.; Gonfloni, S.; Superti-Furga, G.; Roux, B.; Kuriyan, J. Dynamic Coupling between the SH2 and SH3 Domains of C-Src and Hck Underlies Their Inactivation by C-Terminal Tyrosine Phosphorylation. Cell 2001, 105, 115–126. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.S.; Miller, W.T. Cooperative Activation of Src Family Kinases by SH3 and SH2 Ligands. Cancer Lett. 2007, 257, 116–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.-J.; Shamsaddini, A.; Pan, Y.; Smith, K.; Crichton, D.J.; Simonyan, V.; Mazumder, R. A Framework for Organizing Cancer-Related Variations from Existing Databases, Publications and NGS Data Using a High-Performance Integrated Virtual Environment (HIVE). Database 2014, 2014, bau022. [Google Scholar] [CrossRef] [PubMed]
- Dingerdissen, H.M.; Torcivia-Rodriguez, J.; Hu, Y.; Chang, T.-C.; Mazumder, R.; Kahsay, R. BioMuta and BioXpress: Mutation and Expression Knowledgebases for Cancer Biomarker Discovery. Nucleic Acids Res. 2018, 46, D1128–D1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.; Karagiannis, K.; Zhang, H.; Dingerdissen, H.; Shamsaddini, A.; Wan, Q.; Simonyan, V.; Mazumder, R. Human Germline and Pan-Cancer Variomes and Their Distinct Functional Profiles. Nucleic Acids Res. 2014, 42, 11570–11588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malek, S.N.; Desiderio, S. SH2 Domains of the Protein-Tyrosine Kinases Blk, Lyn, and Fyn(T) Bind Distinct Sets of Phosphoproteins from B Lymphocytes. J. Biol. Chem. 1993, 268, 22557–22565. [Google Scholar] [CrossRef]
- Songyang, Z.; Shoelson, S.E.; McGlade, J.; Olivier, P.; Pawson, T.; Bustelo, X.R.; Barbacid, M.; Sabe, H.; Hanafusa, H.; Yi, T. Specific Motifs Recognized by the SH2 Domains of Csk, 3BP2, Fps/Fes, GRB-2, HCP, SHC, Syk, and Vav. Mol. Cell. Biol. 1994, 14, 2777–2785. [Google Scholar] [CrossRef]
- Waksman, G.; Kominos, D.; Robertson, S.C.; Pant, N.; Baltimore, D.; Birge, R.B.; Cowburn, D.; Hanafusa, H.; Mayer, B.J.; Overduin, M.; et al. Crystal Structure of the Phosphotyrosine Recognition Domain SH2 of V-Src Complexed with Tyrosine-Phosphorylated Peptides. Nature 1992, 358, 646–653. [Google Scholar] [CrossRef]
- Jaber Chehayeb, R.; Boggon, T.J. SH2 Domain Binding: Diverse FLVRs of Partnership. Front. Endocrinol. 2020, 11, 575220. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.L.; Wybenga-Groot, L.E.; Tong, J.; Taylor, P.; Minden, M.D.; Trudel, S.; McGlade, C.J.; Moran, M.F. Tyrosine Phosphorylation of the Lyn Src Homology 2 (SH2) Domain Modulates Its Binding Affinity and Specificity*[S]. Mol. Cell. Proteom. 2015, 14, 695–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barouch-Bentov, R.; Che, J.; Lee, C.C.; Yang, Y.; Herman, A.; Jia, Y.; Velentza, A.; Watson, J.; Sternberg, L.; Kim, S.; et al. A Conserved Salt Bridge in the G Loop of Multiple Protein Kinases Is Important for Catalysis and for In Vivo Lyn Function. Mol. Cell 2009, 33, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, D.J.; Rivnay, B.; Avraham, H. CHK Down-Regulates SCF/KL-Activated Lyn Kinase Activity in Mo7e Megakaryocytic Cells. Biochem. Biophys. Res. Commun. 1999, 259, 611–616. [Google Scholar] [CrossRef] [PubMed]
- De Jesus, A.A.; Montealegre, G.; Liu, Y.; Marrero, B.; Kuehn, H.; Calvo, K.; Rosenzweig, S.; Fleisher, T.; Lee, R.C.-C.; Brundidge, A.; et al. A de Novo Nonsense Mutation in the Tyrosine Kinase Lyn in a Patient with an Early Onset Autoinflammatory Phenotype. Pediatr. Rheumatol. 2014, 12, O25. [Google Scholar] [CrossRef] [Green Version]
- Sun, G.; Sharma, A.K.; Budde, R.J. Autophosphorylation of Src and Yes Blocks Their Inactivation by Csk Phosphorylation. Oncogene 1998, 17, 1587–1595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, M. Regulation of the Src Family Kinases by Csk. Int. J. Biol. Sci. 2012, 8, 1385–1397. [Google Scholar] [CrossRef] [Green Version]
- Yanagi, S.; Sugawara, H.; Kurosaki, M.; Sabe, H.; Yamamura, H.; Kurosaki, T. CD45 Modulates Phosphorylation of Both Autophosphorylation and Negative Regulatory Tyrosines of Lyn in B Cells. J. Biol. Chem. 1996, 271, 30487–30492. [Google Scholar] [CrossRef] [Green Version]
- Hochgräfe, F.; Zhang, L.; O’Toole, S.A.; Browne, B.C.; Pinese, M.; Porta Cubas, A.; Lehrbach, G.M.; Croucher, D.R.; Rickwood, D.; Boulghourjian, A.; et al. Tyrosine Phosphorylation Profiling Reveals the Signaling Network Characteristics of Basal Breast Cancer Cells. Cancer Res. 2010, 70, 9391–9401. [Google Scholar] [CrossRef] [Green Version]
- Umemori, H.; Ogura, H.; Tozawa, N.; Mikoshiba, K.; Nishizumi, H.; Yamamoto, T. Impairment of N-Methyl-d-Aspartate Receptor-Controlled Motor Activity in LYN-Deficient Mice. Neuroscience 2003, 118, 709–713. [Google Scholar] [CrossRef]
- Goldenberg-Furmanov, M.; Stein, I.; Pikarsky, E.; Rubin, H.; Kasem, S.; Wygoda, M.; Weinstein, I.; Reuveni, H.; Ben-Sasson, S.A. Lyn Is a Target Gene for Prostate Cancer. Cancer Res. 2004, 64, 1058–1066. [Google Scholar] [CrossRef] [Green Version]
- Ingley, E.; McCarthy, D.J.; Pore, J.R.; Sarna, M.K.; Adenan, A.S.; Wright, M.J.; Erber, W.; Tilbrook, P.A.; Klinken, S.P. Lyn Deficiency Reduces GATA-1, EKLF and STAT5, and Induces Extramedullary Stress Erythropoiesis. Oncogene 2005, 24, 336–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stettner, M.R.; Wang, W.; Nabors, L.B.; Bharara, S.; Flynn, D.C.; Grammer, J.R.; Gillespie, G.Y.; Gladson, C.L. Lyn Kinase Activity Is the Predominant Cellular Src Kinase Activity in Glioblastoma Tumor Cells. Cancer Res. 2005, 65, 5535–5543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, R.C.; Edwards, N.S.; Burns, G.F.; Fisher, D.E. A CD44 Survival Pathway Triggers Chemoresistance via Lyn Kinase and Phosphoinositide 3-Kinase/Akt in Colon Carcinoma Cells. Cancer Res. 2001, 61, 5275–5283. [Google Scholar] [PubMed]
- Berton, G.; Mócsai, A.; Lowell, C.A. Src and Syk Kinases: Key Regulators of Phagocytic Cell Activation. Trends Immunol. 2005, 26, 208–214. [Google Scholar] [CrossRef]
- Vivier, E.; Daëron, M. Immunoreceptor Tyrosine-Based Inhibition Motifs. Immunol. Today 1997, 18, 286–291. [Google Scholar] [CrossRef]
- Malbec, O.; Fong, D.C.; Turner, M.; Tybulewicz, V.L.; Cambier, J.C.; Fridman, W.H.; Daëron, M. Fc Epsilon Receptor I-Associated Lyn-Dependent Phosphorylation of Fc Gamma Receptor IIB during Negative Regulation of Mast Cell Activation. J. Immunol. 1998, 160, 1647–1658. [Google Scholar] [CrossRef]
- Amigorena, S.; Bonnerot, C.; Choquet, D.; Fridman, W.H.; Teillaud, J.L. Fc Gamma RII Expression in Resting and Activated B Lymphocytes. Eur. J. Immunol. 1989, 19, 1379–1385. [Google Scholar] [CrossRef]
- Chan, V.W.F.; Meng, F.; Soriano, P.; DeFranco, A.L.; Lowell, C.A. Characterization of the B Lymphocyte Populations in Lyn-Deficient Mice and the Role of Lyn in Signal Initiation and Down-Regulation. Immunity 1997, 7, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Daëron, M.; Jaeger, S.; Du Pasquier, L.; Vivier, E. Immunoreceptor Tyrosine-Based Inhibition Motifs: A Quest in the Past and Future. Immunol. Rev. 2008, 224, 11–43. [Google Scholar] [CrossRef]
- Damen, J.E.; Liu, L.; Rosten, P.; Humphries, R.K.; Jefferson, A.B.; Majerus, P.W.; Krystal, G. The 145-KDa Protein Induced to Associate with Shc by Multiple Cytokines Is an Inositol Tetraphosphate and Phosphatidylinositol 3,4,5-Triphosphate 5-Phosphatase. Proc. Natl. Acad. Sci. USA 1996, 93, 1689–1693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jellusova, J.; Nitschke, L. Regulation of B Cell Functions by the Sialic Acid-Binding Receptors Siglec-G and CD22. Front. Immunol. 2012, 2, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tedder, T.F.; Tuscano, J.; Sato, S.; Kehrl, J.H. CD22, A B Lymphocyte–Specific Adhesion Molecule That Regulates Antigen Receptor Signaling. Annu. Rev. Immunol. 1997, 15, 481–504. [Google Scholar] [CrossRef] [PubMed]
- Cambier, J.C. Inhibitory Receptors Abound? Proc. Natl. Acad. Sci. USA 1997, 94, 5993–5995. [Google Scholar] [CrossRef] [Green Version]
- Nitschke, L. The Role of CD22 and Other Inhibitory Co-Receptors in B-Cell Activation. Curr. Opin. Immunol. 2005, 17, 290–297. [Google Scholar] [CrossRef]
- Doody, G.M.; Justement, L.B.; Delibrias, C.C.; Matthews, R.J.; Lin, J.; Thomas, M.L.; Fearon, D.T. A Role in B Cell Activation for CD22 and the Protein Tyrosine Phosphatase SHP. Science 1995, 269, 242–244. [Google Scholar] [CrossRef] [PubMed]
- Kubagawa, H.; Burrows, P.D.; Cooper, M.D. A Novel Pair of Immunoglobulin-like Receptors Expressed by B Cells and Myeloid Cells. Proc. Natl. Acad. Sci. USA 1997, 94, 5261–5266. [Google Scholar] [CrossRef] [Green Version]
- Hayami, K.; Fukuta, D.; Nishikawa, Y.; Yamashita, Y.; Inui, M.; Ohyama, Y.; Hikida, M.; Ohmori, H.; Takai, T. Molecular Cloning of a Novel Murine Cell-Surface Glycoprotein Homologous to Killer Cell Inhibitory Receptors. J. Biol. Chem. 1997, 272, 7320–7327. [Google Scholar] [CrossRef] [Green Version]
- Maeda, A.; Kurosaki, M.; Ono, M.; Takai, T.; Kurosaki, T. Requirement of SH2-Containing Protein Tyrosine Phosphatases SHP-1 and SHP-2 for Paired Immunoglobulin-like Receptor B (PIR-B)–Mediated Inhibitory Signal. J. Exp. Med. 1998, 187, 1355–1360. [Google Scholar] [CrossRef]
- Van der Touw, W.; Chen, H.-M.; Pan, P.-Y.; Chen, S.-H. LILRB Receptor-Mediated Regulation of Myeloid Cell Maturation and Function. Cancer Immunol. Immunother. 2017, 66, 1079–1087. [Google Scholar] [CrossRef]
- Adams, S.; van der Laan, L.J.; Vernon-Wilson, E.; Renardel de Lavalette, C.; Döpp, E.A.; Dijkstra, C.D.; Simmons, D.L.; van den Berg, T.K. Signal-Regulatory Protein Is Selectively Expressed by Myeloid and Neuronal Cells. J. Immunol. 1998, 161, 1853–1859. [Google Scholar] [CrossRef] [PubMed]
- Harder, K.W.; Parsons, L.M.; Armes, J.; Evans, N.; Kountouri, N.; Clark, R.; Quilici, C.; Grail, D.; Hodgson, G.S.; Dunn, A.R.; et al. Gain- and Loss-of-Function Lyn Mutant Mice Define a Critical Inhibitory Role for Lyn in the Myeloid Lineage. Immunity 2001, 15, 603–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.-N.; Yan, H.-X.; Chen, L.; Dong, L.-W.; Yang, W.; Liu, Q.; Yu, L.-X.; Huang, D.-D.; Liu, S.-Q.; Liu, H.; et al. LPS-Induced Down-Regulation of Signal Regulatory Protein {alpha} Contributes to Innate Immune Activation in Macrophages. J. Exp. Med. 2007, 204, 2719–2731. [Google Scholar] [CrossRef] [PubMed]
- Bezbradica, J.S.; Medzhitov, R. Role of ITAM Signaling Module in Signal Integration. Curr. Opin. Immunol. 2012, 24, 58–66. [Google Scholar] [CrossRef]
- Hogarth, P.M. Fc Receptors Are Major Mediators of Antibody Based Inflammation in Autoimmunity. Curr. Opin. Immunol. 2002, 14, 798–802. [Google Scholar] [CrossRef]
- Humphrey, M.B.; Lanier, L.L.; Nakamura, M.C. Role of ITAM-Containing Adapter Proteins and Their Receptors in the Immune System and Bone. Immunol. Rev. 2005, 208, 50–65. [Google Scholar] [CrossRef]
- Takata, M.; Sabe, H.; Hata, A.; Inazu, T.; Homma, Y.; Nukada, T.; Yamamura, H.; Kurosaki, T. Tyrosine Kinases Lyn and Syk Regulate B Cell Receptor-Coupled Ca2+ Mobilization through Distinct Pathways. EMBO J. 1994, 13, 1341–1349. [Google Scholar] [CrossRef]
- Pullen, N.A.; Falanga, Y.T.; Morales, J.K.; Ryan, J.J. The Fyn-STAT5 Pathway: A New Frontier in IgE- and IgG-Mediated Mast Cell Signaling. Front. Immunol. 2012, 3, 117. [Google Scholar] [CrossRef] [Green Version]
- Ben Mkaddem, S.; Hayem, G.; Jönsson, F.; Rossato, E.; Boedec, E.; Boussetta, T.; El Benna, J.; Launay, P.; Goujon, J.-M.; Benhamou, M.; et al. Shifting FcγRIIA-ITAM from Activation to Inhibitory Configuration Ameliorates Arthritis. J. Clin. Investig. 2014, 124, 3945–3959. [Google Scholar] [CrossRef] [Green Version]
- Kanamaru, Y.; Pfirsch, S.; Aloulou, M.; Vrtovsnik, F.; Essig, M.; Loirat, C.; Deschênes, G.; Guérin-Marchand, C.; Blank, U.; Monteiro, R.C. Inhibitory ITAM Signaling by FcαRI-FcRγ Chain Controls Multiple Activating Responses and Prevents Renal Inflammation. J. Immunol. 2008, 180, 2669–2678. [Google Scholar] [CrossRef]
- Pasquier, B.; Launay, P.; Kanamaru, Y.; Moura, I.C.; Pfirsch, S.; Ruffié, C.; Hénin, D.; Benhamou, M.; Pretolani, M.; Blank, U.; et al. Identification of FcαRI as an Inhibitory Receptor That Controls Inflammation. Immunity 2005, 22, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Štefanová, I.; Hemmer, B.; Vergelli, M.; Martin, R.; Biddison, W.E.; Germain, R.N. TCR Ligand Discrimination Is Enforced by Competing ERK Positive and SHP-1 Negative Feedback Pathways. Nat. Immunol. 2003, 4, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Getahun, A.; Beavers, N.A.; Larson, S.R.; Shlomchik, M.J.; Cambier, J.C. Continuous Inhibitory Signaling by Both SHP-1 and SHIP-1 Pathways Is Required to Maintain Unresponsiveness of Anergic B Cells. J. Exp. Med. 2016, 213, 751–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.B.M.; Prigent, S.A. Insights into the Shc Family of Adaptor Proteins. J. Mol. Signal. 2017, 12, 2. [Google Scholar] [CrossRef] [Green Version]
- Ptasznik, A.; Traynor-Kaplan, A.; Bokoch, G.M. G Protein-Coupled Chemoattractant Receptors Regulate Lyn Tyrosine Kinase·Shc Adapter Protein Signaling Complexes. J. Biol. Chem. 1995, 270, 19969–19973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyo, S.; Sada, K.; Qu, X.; Maeno, K.; Miah, S.M.S.; Kawauchi-Kamata, K.; Yamamura, H. Negative Regulation of Lyn Protein-Tyrosine Kinase by c-Cbl Ubiquitin-Protein Ligase in FcɛRI-Mediated Mast Cell Activation. Genes Cells 2003, 8, 825–836. [Google Scholar] [CrossRef]
- Dombrosky-Ferlan, P.; Grishin, A.; Botelho, R.J.; Sampson, M.; Wang, L.; Rudert, W.A.; Grinstein, S.; Corey, S.J. Felic (CIP4b), a Novel Binding Partner with the Src Kinase Lyn and Cdc42, Localizes to the Phagocytic Cup. Blood 2003, 101, 2804–2809. [Google Scholar] [CrossRef]
- Cheung, R.; Malik, M.; Ravyn, V.; Tomkowicz, B.; Ptasznik, A.; Collman, R.G. An Arrestin-Dependent Multi-Kinase Signaling Complex Mediates MIP-1β/CCL4 Signaling and Chemotaxis of Primary Human Macrophages. J. Leukoc. Biol. 2009, 86, 833–845. [Google Scholar] [CrossRef] [Green Version]
- Houslay, K.F.; Christian, F.; MacLeod, R.; Adams, D.R.; Houslay, M.D.; Baillie, G.S. Identification of a Multifunctional Docking Site on the Catalytic Unit of Phosphodiesterase-4 (PDE4) That Is Utilised by Multiple Interaction Partners. Biochem. J. 2017, 474, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, K.; Kharbanda, S.; Kufe, D. Functional Interaction between SHPTP1 and the Lyn Tyrosine Kinase in the Apoptotic Response to DNA Damage. J. Biol. Chem. 1999, 274, 34663–34668. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Q.; Yu, S.P. Novel Regulation of Na+, K+-ATPase by Src Tyrosine Kinases in Cortical Neurons. J. Neurochem. 2005, 93, 1515–1523. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Umemori, H.; Mishina, M.; Yamamoto, T. The AMPA Receptor Interacts with and Signals through the Protein Tyrosine Kinase Lyn. Nature 1999, 397, 72–76. [Google Scholar] [CrossRef]
- Roseweir, A.K.; Qayyum, T.; Lim, Z.; Hammond, R.; MacDonald, A.I.; Fraser, S.; Oades, G.M.; Aitchison, M.; Jones, R.J.; Edwards, J. Nuclear Expression of Lyn, a Src Family Kinase Member, Is Associated with Poor Prognosis in Renal Cancer Patients. BMC Cancer 2016, 16, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsberger, B.; Fullerton, R.; Zino, S.; Jordan, F.; Mitchell, T.J.; Brunton, V.G.; Mallon, E.A.; Shiels, P.G.; Edwards, J. Breast Cancer Patients’ Clinical Outcome Measures Are Associated with Src Kinase Family Member Expression. Br. J. Cancer 2010, 103, 899–909. [Google Scholar] [CrossRef] [PubMed]
- Croucher, D.R.; Hochgräfe, F.; Zhang, L.; Liu, L.; Lyons, R.J.; Rickwood, D.; Tactacan, C.M.; Browne, B.C.; Ali, N.; Chan, H.; et al. Involvement of Lyn and the Atypical Kinase SgK269/PEAK1 in a Basal Breast Cancer Signaling Pathway. Cancer Res. 2013, 73, 1969–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, G.; Breen, E.J.; Ranganathan, S. Identification of Ovarian Cancer Associated Genes Using an Integrated Approach in a Boolean Framework. BMC Syst. Biol. 2013, 7, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, C.; Demur, C.; Bardet, V.; Prade-Houdellier, N.; Payrastre, B.; Récher, C. A Critical Role for Lyn in Acute Myeloid Leukemia. Blood 2008, 111, 2269–2279. [Google Scholar] [CrossRef]
- Michell-Robinson, M.A.; Touil, H.; Healy, L.M.; Owen, D.R.; Durafourt, B.A.; Bar-Or, A.; Antel, J.P.; Moore, C.S. Roles of Microglia in Brain Development, Tissue Maintenance and Repair. Brain 2015, 138, 1138–1159. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.; Parikh, I.; Vasquez, J.B.; Smith, C.; Tai, L.; Bu, G.; LaDu, M.J.; Fardo, D.W.; Rebeck, G.W.; Estus, S. Genetics Ignite Focus on Microglial Inflammation in Alzheimer’s Disease. Mol. Neurodegener. 2015, 10, 52. [Google Scholar] [CrossRef] [Green Version]
- Keren-Shaul, H.; Spinrad, A.; Weiner, A.; Matcovitch-Natan, O.; Dvir-Szternfeld, R.; Ulland, T.K.; David, E.; Baruch, K.; Lara-Astaiso, D.; Toth, B.; et al. A Unique Microglia Type Associated with Restricting Development of Alzheimer’s Disease. Cell 2017, 169, 1276–1290.e17. [Google Scholar] [CrossRef]
- Sierksma, A.; Lu, A.; Mancuso, R.; Fattorelli, N.; Thrupp, N.; Salta, E.; Zoco, J.; Blum, D.; Buée, L.; De Strooper, B.; et al. Novel Alzheimer Risk Genes Determine the Microglia Response to Amyloid-β but Not to TAU Pathology. EMBO Mol. Med. 2020, 12, e10606. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, A.; Ekavali. A Review on Alzheimer’s Disease Pathophysiology and Its Management: An Update. Pharmacol. Reports 2015, 67, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J. Alzheimer’s Disease: The Amyloid Cascade Hypothesis: An Update and Reappraisal. J. Alzheimer’s Dis. 2006, 9, 151–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portugal, C.C.; Almeida, T.O.; Socodato, R.; Relvas, J.B. Src Family Kinases (SFKs): Critical Regulators of Microglial Homeostatic Functions and Neurodegeneration in Parkinson’s and Alzheimer’s Diseases. FEBS J. 2021, 289, 7760–7775. [Google Scholar] [CrossRef]
- Combs, C.K.; Johnson, D.E.; Cannady, S.B.; Lehman, T.M.; Landreth, G.E. Identification of Microglial Signal Transduction Pathways Mediating a Neurotoxic Response to Amyloidogenic Fragments of β-Amyloid and Prion Proteins. J. Neurosci. 1999, 19, 928–939. [Google Scholar] [CrossRef] [Green Version]
- Hanke, J.H.; Gardner, J.P.; Dow, R.L.; Changelian, P.S.; Brissette, W.H.; Weringer, E.J.; Pollok, B.A.; Connelly, P.A. Discovery of a Novel, Potent, and Src Family-Selective Tyrosine Kinase Inhibitor. J. Biol. Chem. 1996, 271, 695–701. [Google Scholar] [CrossRef] [Green Version]
- Sondag, C.M.; Dhawan, G.; Combs, C.K. Beta Amyloid Oligomers and Fibrils Stimulate Differential Activation of Primary Microglia. J. Neuroinflamm. 2009, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Dhawan, G.; Floden, A.M.; Combs, C.K. Amyloid-β Oligomers Stimulate Microglia through a Tyrosine Kinase Dependent Mechanism. Neurobiol. Aging 2012, 33, 2247–2261. [Google Scholar] [CrossRef] [Green Version]
- Manocha, G.D.; Puig, K.L.; Austin, S.A.; Seyb, K.; Glicksman, M.A.; Combs, C.K. Characterization of Novel Src Family Kinase Inhibitors to Attenuate Microgliosis. PLoS ONE 2015, 10, e0132604. [Google Scholar] [CrossRef]
- Gwon, Y.; Kim, S.-H.; Kim, H.T.; Kam, T.-I.; Park, J.; Lim, B.; Cha, H.; Chang, H.-J.; Hong, Y.R.; Jung, Y.-K. Amelioration of Amyloid Β-FcγRIIb Neurotoxicity and Tau Pathologies by Targeting LYN. FASEB J. 2019, 33, 4300–4313. [Google Scholar] [CrossRef]
- Gwon, Y.; Kam, T.-I.; Kim, S.-H.; Song, S.; Park, H.; Lim, B.; Lee, H.; Lee, W.; Jo, D.-G.; Jung, Y.-K. TOM1 Regulates Neuronal Accumulation of Amyloid-β Oligomers by FcγRIIb2 Variant in Alzheimer’s Disease. J. Neurosci. 2018, 38, 9001–9018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peikert, K.; Federti, E.; Matte, A.; Constantin, G.; Pietronigro, E.C.; Fabene, P.F.; Defilippi, P.; Turco, E.; Del Gallo, F.; Pucci, P.; et al. Therapeutic Targeting of Lyn Kinase to Treat Chorea-Acanthocytosis. Acta Neuropathol. Commun. 2021, 9, 81. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Organization of human Lyn domains and key residue differences between the human (P07948), mouse (P25911), and rat (Q07014) proteins.
Figure 1.
Organization of human Lyn domains and key residue differences between the human (P07948), mouse (P25911), and rat (Q07014) proteins.

Figure 2.
The secondary structure of Lyn (A) SH3 domain bound proline-rich peptide ligand (Figure was constructed by superimposing and combining the human Lyn SH3 domain (PDB ID: 6NMW) with c-Src SH3 domain bound peptide ligand (PDB ID: 1QWE) and (B) SH2 domain bound phosphotyrosine peptide (Figure was constructed by superimposing and combining the human Lyn SH2 domain (PDB ID: 4TZI) with Src SH2 domain bound phosphotyrosine ligand (PDB ID 1SPR) (C) SH1 (catalytic) domain in DFG-out/αC-in conformation (Figure was constructed by homology modeling based on Lck (2PL0). N-Terminal (N-T) end. C-Terminal (C-T) end.
Figure 2.
The secondary structure of Lyn (A) SH3 domain bound proline-rich peptide ligand (Figure was constructed by superimposing and combining the human Lyn SH3 domain (PDB ID: 6NMW) with c-Src SH3 domain bound peptide ligand (PDB ID: 1QWE) and (B) SH2 domain bound phosphotyrosine peptide (Figure was constructed by superimposing and combining the human Lyn SH2 domain (PDB ID: 4TZI) with Src SH2 domain bound phosphotyrosine ligand (PDB ID 1SPR) (C) SH1 (catalytic) domain in DFG-out/αC-in conformation (Figure was constructed by homology modeling based on Lck (2PL0). N-Terminal (N-T) end. C-Terminal (C-T) end.

Figure 3.
The structure of Lyn kinase catalytic domain in different conformations (A) DFG-in/αC-in (DFG-Asp-in) active conformation bound ATP analog (ATP-CHF) and Mg2+ (Figure was constructed by superimposing and combining c-Src-ATP-CHF complex (PDB ID:5XP5) and active mouse Lyn (PDB ID:2ZV8) (B) DFG-in/αC-out (DFG-Asp-in) inactive conformation bound type I1/2 inhibitor (Figure was constructed by homology modeling of Lyn DFG-in/αC-out based on human Hck (PDB ID: 5H0B) (C) DFG-out/αC-in (DFG-Asp-out) inactive conformation bound type II inhibitor (Imatinib) (Figure was constructed by homology modeling of Lyn DFG-out/αC-in based on human Lck (PDB: 2PL0).
Figure 3.
The structure of Lyn kinase catalytic domain in different conformations (A) DFG-in/αC-in (DFG-Asp-in) active conformation bound ATP analog (ATP-CHF) and Mg2+ (Figure was constructed by superimposing and combining c-Src-ATP-CHF complex (PDB ID:5XP5) and active mouse Lyn (PDB ID:2ZV8) (B) DFG-in/αC-out (DFG-Asp-in) inactive conformation bound type I1/2 inhibitor (Figure was constructed by homology modeling of Lyn DFG-in/αC-out based on human Hck (PDB ID: 5H0B) (C) DFG-out/αC-in (DFG-Asp-out) inactive conformation bound type II inhibitor (Imatinib) (Figure was constructed by homology modeling of Lyn DFG-out/αC-in based on human Lck (PDB: 2PL0).

Figure 4.
Schematic representation of Lyn kinase regulation (dephosphorylation of the activation loop (Tyr397) by phosphatases such as PTPB1 followed by phosphorylation of C-terminal (Tyr508) by Csk leads to inactive-assembled (self-inhibited) state. Dephosphorylation of the C-terminal by transmembrane phosphatases such as CD45, followed by autophosphorylation of the activation loop, leads to the active state).
Figure 4.
Schematic representation of Lyn kinase regulation (dephosphorylation of the activation loop (Tyr397) by phosphatases such as PTPB1 followed by phosphorylation of C-terminal (Tyr508) by Csk leads to inactive-assembled (self-inhibited) state. Dephosphorylation of the C-terminal by transmembrane phosphatases such as CD45, followed by autophosphorylation of the activation loop, leads to the active state).

Figure 5.
Summary of the ITAM/ITIM bearing receptors and adaptor proteins known to be phosphorylated by Lyn and the general mechanism of the activating/inhibitory signaling. (A) Phosphorylation of ITAM by Lyn and initiation of activating signaling (ITAM-signaling) (B) Partial phosphorylation of ITAM by Lyn and initiation of inhibitory signaling (ITAMi-signaling) (C) Phosphorylation of ITIM by Lyn and initiation of inhibitory signaling (ITIM-signaling). (Differential targeting of ITAM-bearing receptors can activate either activating or inhibitory signaling where Lyn is required for both processes. (B) was constructed based on the mechanism suggested in [5]).
Figure 5.
Summary of the ITAM/ITIM bearing receptors and adaptor proteins known to be phosphorylated by Lyn and the general mechanism of the activating/inhibitory signaling. (A) Phosphorylation of ITAM by Lyn and initiation of activating signaling (ITAM-signaling) (B) Partial phosphorylation of ITAM by Lyn and initiation of inhibitory signaling (ITAMi-signaling) (C) Phosphorylation of ITIM by Lyn and initiation of inhibitory signaling (ITIM-signaling). (Differential targeting of ITAM-bearing receptors can activate either activating or inhibitory signaling where Lyn is required for both processes. (B) was constructed based on the mechanism suggested in [5]).

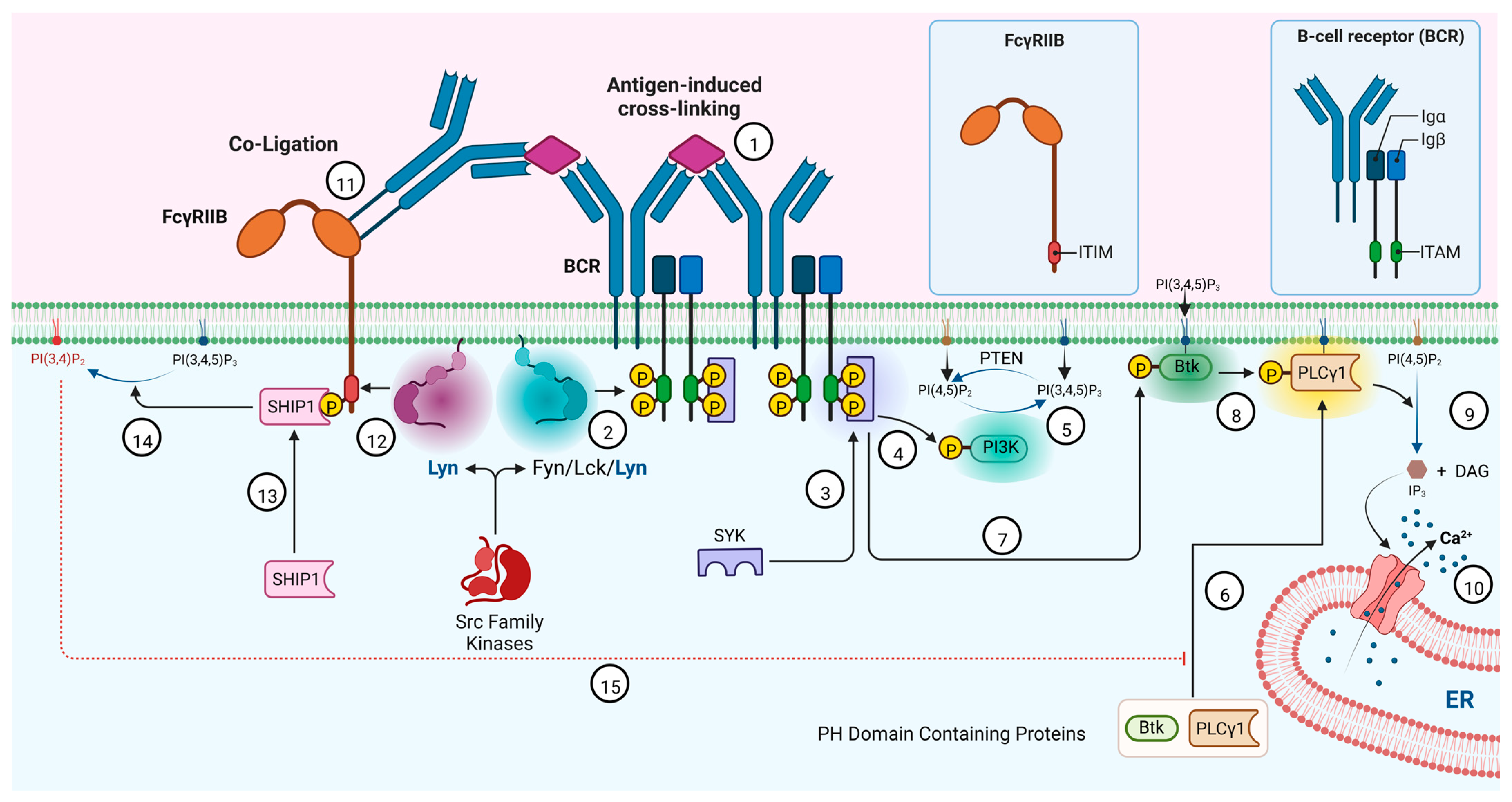
Figure 6.
Lyn involvement in B cell activation/inhibition mechanism. Activation pathway (steps 1–10): (1) antigen-induced crosslinking of BCR activates and translocates Src kinases such as Lyn and Fyn to the inner leaflet of the membrane and enhances the association with lipid raft; (2) Lyn/Fyn phosphorylates ITAMs associated with BCR; (3) phosphorylated ITAMs recruit Syk and activate it; (4) activated Syk phosphorylate PI3K; (5) activated PI3K phosphorylates PI(4,5)P2 and generate PI(3,4,5)P3; (6) PI(3,4,5)P3 act as membrane docking sites for PH domain-containing proteins such as Btk and PLCγ1; (7) membrane-recruited Btk is phosphorylated by activated Syk (proximity); (8) activated Btk phosphorylates PLCγ1; (9) activated PLCγ1 converts PI(4,5)P2 to IP3; and (10) increased IP3 level enhances the calcium mobilization leading to B-cell activation. Inhibition pathway (steps 11–15): (11) co-ligation of FcγRII enhances the association of Lyn with ITIM of the receptor; (12) activated Lyn phosphorylates ITIM of the FcγRII; (13) phosphorylated ITIM recruits and activates SHIP1; (14) SHIP1 converts PI(3,4,5)P3 to PI(3,4)P2 and impairs the recruitment of PH domain containing Btk and PLCγ1 to the cell membrane and their activation, causing a decrease in IP3 level. This leads to impaired Ca2+ mobilization causing B-cell inactivation.
Figure 6.
Lyn involvement in B cell activation/inhibition mechanism. Activation pathway (steps 1–10): (1) antigen-induced crosslinking of BCR activates and translocates Src kinases such as Lyn and Fyn to the inner leaflet of the membrane and enhances the association with lipid raft; (2) Lyn/Fyn phosphorylates ITAMs associated with BCR; (3) phosphorylated ITAMs recruit Syk and activate it; (4) activated Syk phosphorylate PI3K; (5) activated PI3K phosphorylates PI(4,5)P2 and generate PI(3,4,5)P3; (6) PI(3,4,5)P3 act as membrane docking sites for PH domain-containing proteins such as Btk and PLCγ1; (7) membrane-recruited Btk is phosphorylated by activated Syk (proximity); (8) activated Btk phosphorylates PLCγ1; (9) activated PLCγ1 converts PI(4,5)P2 to IP3; and (10) increased IP3 level enhances the calcium mobilization leading to B-cell activation. Inhibition pathway (steps 11–15): (11) co-ligation of FcγRII enhances the association of Lyn with ITIM of the receptor; (12) activated Lyn phosphorylates ITIM of the FcγRII; (13) phosphorylated ITIM recruits and activates SHIP1; (14) SHIP1 converts PI(3,4,5)P3 to PI(3,4)P2 and impairs the recruitment of PH domain containing Btk and PLCγ1 to the cell membrane and their activation, causing a decrease in IP3 level. This leads to impaired Ca2+ mobilization causing B-cell inactivation.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Weerawarna, P.M.; Richardson, T.I. Lyn Kinase Structure, Regulation, and Involvement in Neurodegenerative Diseases: A Mini Review. Kinases Phosphatases 2023, 1, 23-38. https://doi.org/10.3390/kinasesphosphatases1010004
AMA Style
Weerawarna PM, Richardson TI. Lyn Kinase Structure, Regulation, and Involvement in Neurodegenerative Diseases: A Mini Review. Kinases and Phosphatases. 2023; 1(1):23-38. https://doi.org/10.3390/kinasesphosphatases1010004
Chicago/Turabian StyleWeerawarna, Pathum M., and Timothy I. Richardson. 2023. "Lyn Kinase Structure, Regulation, and Involvement in Neurodegenerative Diseases: A Mini Review" Kinases and Phosphatases 1, no. 1: 23-38. https://doi.org/10.3390/kinasesphosphatases1010004