1. Introduction
Scedosporium species are opportunistic filamentous fungi belonging to the
Microascaceae family. Among the ten species that have been so far identified in the
Scedosporium genus, only
S. apiospermum,
S. boydii,
S. aurantiacum, and
S. minutisporum are clinically relevant [
1]. These fungi are mesophilic, eutrophic, and environmental saprophytes frequently found in human-impacted areas, including industrial regions and wastewater [
2]. When introduced into the human body by inhalation and/or traumatic injury, conidia of these species are virulent enough to cause infections in both healthy and immunocompromised subjects, with attributable mortality of alarming rates in the case of disseminated infections [
3]. One example of a vulnerable population is patients with cystic fibrosis (CF), whose airways are often chronically colonized by
S. apiospermum,
S. boydii, and
S. aurantiacum. Colonized patients are usually not at risk of invasive pulmonary infections unless lung transplantation takes place [
4,
5]. In addition to their therapy-refractory nature, the treatment of infections caused by these species is impinged by their nocuous virulent features that lead to irreparable harm to invaded tissues. This includes the presence of cell wall components with immunomodulatory effects, the secretion of proteins and metabolites that promote the ability to establish infections, and the attitude of the fungus toward the stressful conditions imposed in the host [
6,
7,
8]. In this regard, and since the release of the first whole-genome sequencing of
S. apiospermum [
9], special attention has been paid to the antioxidant repertoire that permits the fungus to modulate host-imposed oxidative stress. Genomic and transcriptomic studies have elicited the key involvement of specific antioxidant enzymes in the tolerance of
S. apiospermum to chemically derived oxidants and phagocytes-mediated killing, such as the conidia-specific Cu,Zn-superoxide dismutase (SOD), the thioredoxin reductases (TrxR), and SaPrx2 (a peroxiredoxin) [
10,
11,
12]. Such evidence relative to the behavior of
Scedosporium in their natural niches and the human host infers that the ability of these fungi to adapt to stress is high. As a matter of fact, it is assumptive that the cellular response to any threats should have been operated by cellular regulators. As the stress-signaling pathways in
Scedosporium are still completely unexplored, deciphering the signaling pathways, especially that related to oxidative stress, is needed in order to pave the way for new therapeutic approaches.
The broad diversity of fungal niches has always been attractive for fundamental research, which provided a paradigm of adaptation to stress in the eukaryotic world. As a matter of fact, the ability of fungi to evoke adaptive responses to unforeseen conditions relies on sensor-effector complex networks that orchestrate global cellular reprogramming. One of the most conclusive pieces of evidence in this regard states that the high osmolarity glycerol (HOG) pathway constitutes the key molecular apparatus that accounts for fungal adaptation to myriad types of stress [
13,
14]. This is also reflected by the fact that Hog1, a member of the mitogen-activated protein kinase (MAPK) family and the key element in the HOG pathway, is highly conserved in the fungal kingdom regardless of the species niche (identity of 80 to 100%) [
15]. When activated under stress, the HOG pathway undergoes sequential reactions of phosphorylation of its three tiers: MAP kinase kinase kinase (MAPKKK), MAP kinase kinase (MAPKK), and Hog1 MAPK (
Figure 1) [
16]. The activation of the latter is represented by dual phosphorylation on its TGY (Threonine-Glycine-Tyrosine) motif, thus allowing the Hog1-mediated transcriptional and post-transcriptional regulation of cellular targets. The shuttling of cues to the HOG pathway is mainly facilitated by a two-component system (TCS), which is a multi-step histidine-aspartate phosphorelay system formed by a hybrid histidine kinase (HK), and a histidine-containing phosphotransfer protein (HPT) acting as a mediator between HK and a response regulator [
17]. Although formed by a limited number of components, the HOG pathway intriguingly constitutes a nexus regulator required to instigate fine-tuned responses to diverse stress, including osmotic, oxidative, antifungals, heat, acid, ethanol, and many others [
13,
14]. Interestingly, recent findings outlined that the HOG pathway plays a critical role in the adaptation of strains of
Aspergillus fumigatus to the lung microenvironment, including CF lungs [
18,
19].
In light of the nodal role played by the HOG pathway in fungal adaptive responses, we intended to probe the role of this pathway in the tolerance of S. apiospermum to stress, with a focus on oxidative stress. The established methodology was based on functional genomic approaches to produce mutants devoid of pathway components and to verify the phosphorylation of Hog1 in response to various stresses by Western blot. Here, we present the first record of the stress-induced phosphorylation of Hog1 in S. apiospermum (SaHog1). We report distinct responsiveness of SaHog1 to several types of stress, suggesting that the HOG pathway constitutes a stress Hub in S. apiospermum.
2. Results
2.1. SaHog1 Is Highly Conserved
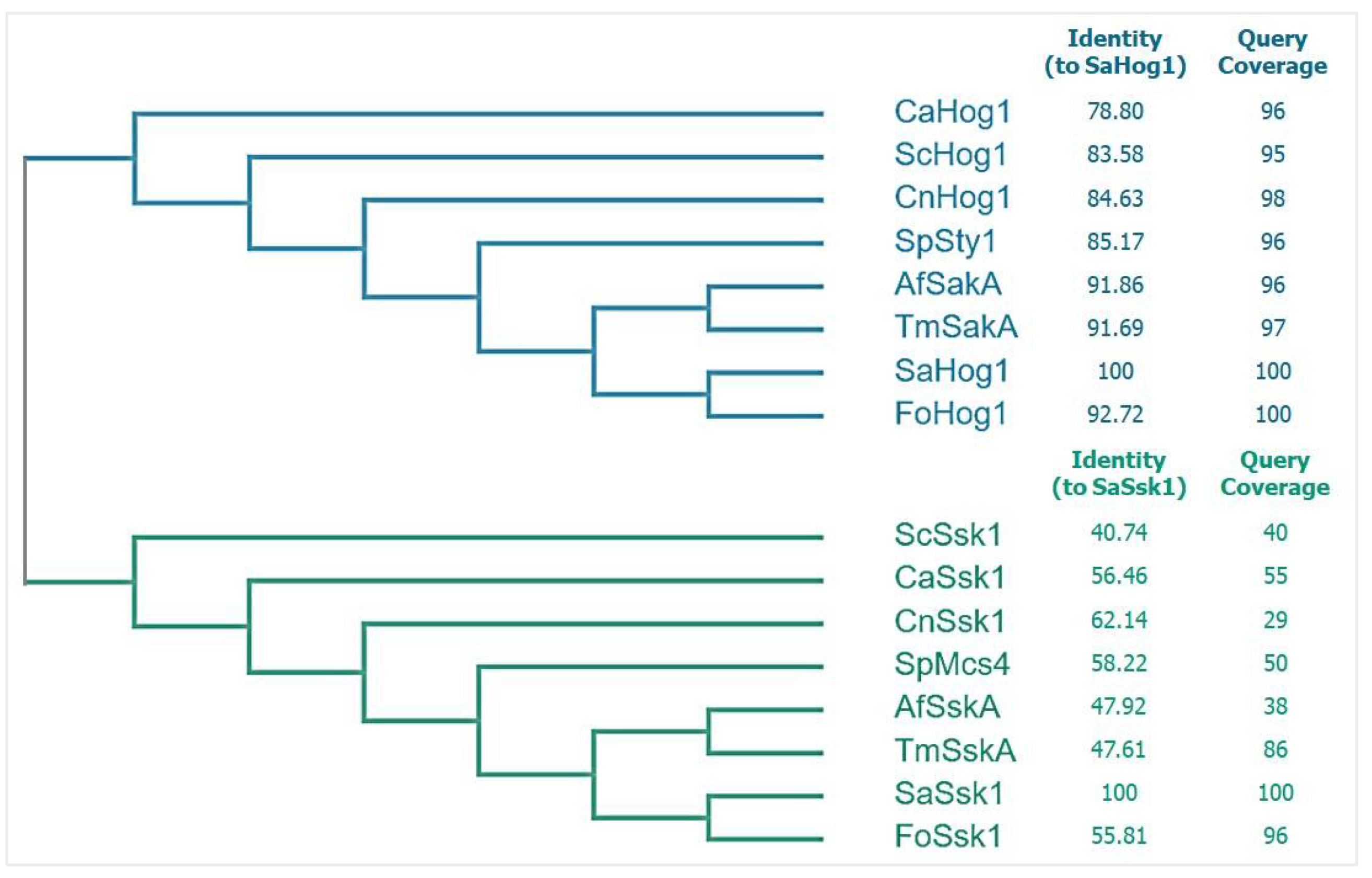
Bioinformatic analysis showed that Hog1 is encoded in the genome of S. apiospermum IHEM 14462, by the gene SAPIO_CDS7351. Unlike Aspergillus genomes which harbor two paralogs of Hog1 [
20,
21], the genome of S. apiospermum IHEM 14462 comprises only one copy. The cladogram analysis of fungal Hog1 shown in
Figure 2 reveals that SaHog1 (XP_016641060.1) belongs to the Hog1 cluster that is different from the Ssk1 response regulator cluster. Identity analysis shows that fungal Hog1 is more conserved than fungal Ssk1 response regulator, supporting findings brought by Nikolaou et al. [
15]. SaHog1 (XP_016641060.1) is most and least identical to that of Fusarium oxysporum (92.72%) and Candida albicans (78.50%), supporting that it is highly conserved regardless of the phylogenetic distance. SaHog1 is of 357 amino acids in length, and the estimated molecular mass is 41.9 kDa.
2.2. The Phosphorylation of SaHog1 Is Triggered by Many Stresses
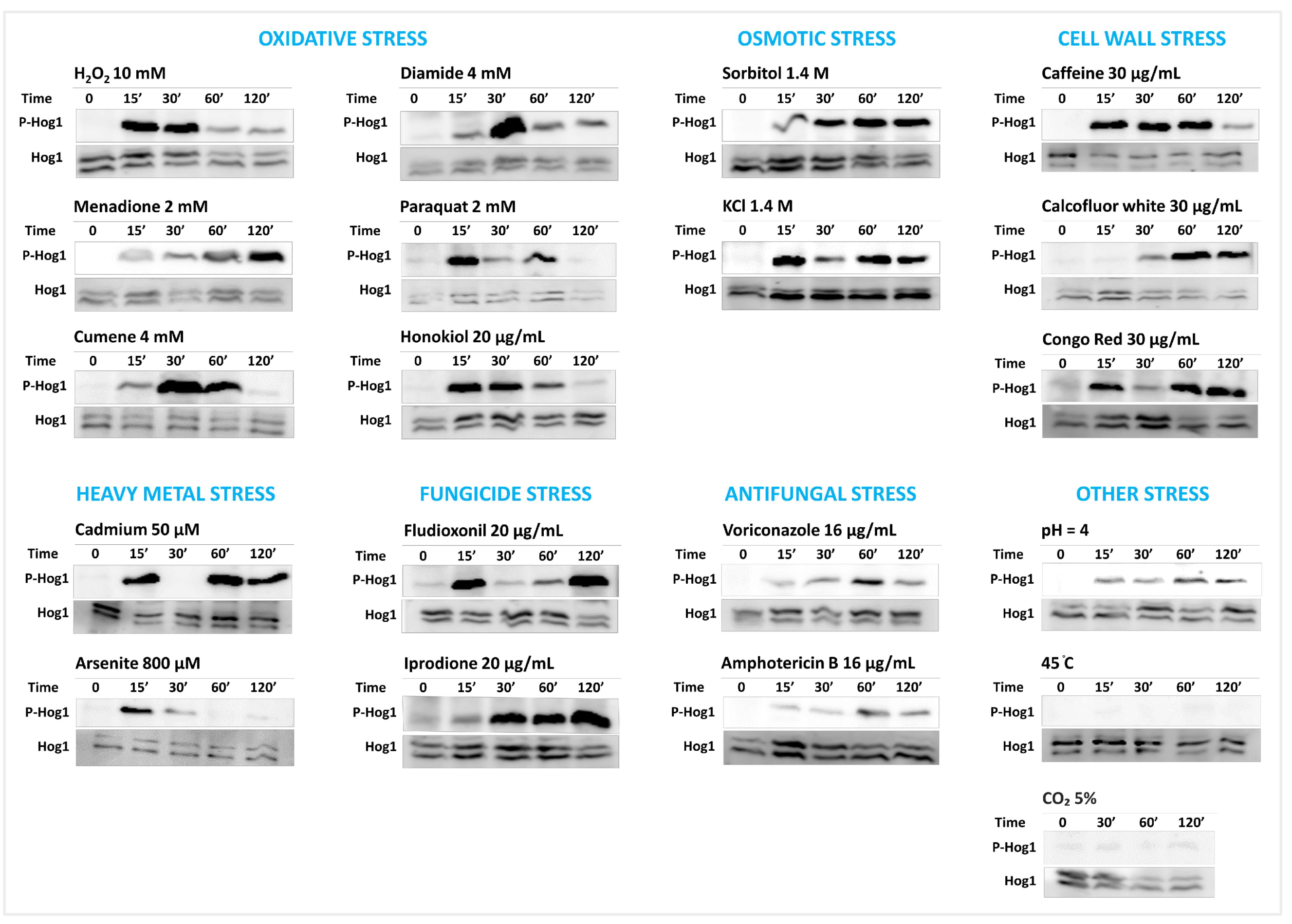
The Western blot analysis showed that the phosphorylation of SaHog1 is induced in 18 out of the 20 tested conditions in comparison to unstressed conditions (
Figure 3). The two conditions that had no discernable effect on the phosphorylation of SaHog1 are the thermal stress (45 °C) and the hypercapnia (5% CO
2). Interestingly, the kinetics of phosphorylation were diverse even within the same group of conditions. The kinetics for each group of stresses are described below.
Oxidative stress: All oxidative stress conditions (10 mM H2O2, 2 mM menadione, 4 mM cumene hydroperoxide, 4 mM diamide, 4 mM paraquat, and 20 μg/mL honokiol) triggered the robust phosphorylation of SaHog1. However, the kinetics of phosphorylation were not always the same. The stress imposed by H2O2, honokiol, or paraquat-induced the rapid activation of SaHog1, with a maximum induction since the first 15 min, followed by an incremental decrease in intensity with time, so that the phosphorylation was barely detected after 120 min of incubation. The cumene- and diamide-induced phosphorylation of SaHog1 stress occurred at a slower pace; phosphorylation, weakly detected at 15 min, reached its maximum at 30 min, and then decreased with time. Treatment with menadione induced the very slow activation of SaHog1, with the maximum intensity attained at the latest tested time point (120 min).
Osmotic stress: Sorbitol and KCl (both at 1.4 M) induced the strong phosphorylation of SaHog1 with comparable patterns. The intensity of phosphorylation was sustained regardless of the time.
Cell wall stress: The cell wall-imposing agents like caffeine, calcofluor white, and Congo Red (all at 30 μg/mL) promoted the phosphorylation of SaHog1 in distinct kinetics. The caffeine-induced activation of SaHog1 was rapid and transient, while that induced by calcofluor white was slow and prolonged. Using Congo Red, a “discrete-activation effect” was observed, meaning that SaHog1 phosphorylation declined between two increases; at 30 min of stress, Congo Red-induced phosphorylated SaHog1 returned to nearly its basal level of phosphorylation before it arose again. This effect was reproducible even when different experimental runs were used.
Heavy metal stress: The activation of SaHog1 by cadmium [Cd (NO3)2, 50 μM] and arsenite (As2O3, 800 μM) was kinetically different. Cadmium also exhibited a discrete-activation effect; SaHog1 underwent intense activation within 15 min of exposure, then returned to a normal level 30 min before it was prompted again by 60 min. On the other hand, arsenite triggered the rapid and temporal activation of SaHog1.
Fungicide stress: The phenylpyrrole fludioxonil (20 μg/mL) also induced a discontinued activation pattern, with a marked activation of SaHog1 at 15 min followed by a decreased phosphorylation and its re-activation at 120 min. By contrast, exposure to the dicarboximide iprodione (20 μg/mL) resulted in a delayed and prolonged induction of SaHog1 phosphorylation.
Other stress: Voriconazole and amphotericin B (both at 16 μg/mL), as well as acid pH (pH 4), stimulated the time-independent and moderate phosphorylation of SaHog1, while the basal level of phosphorylated SaHog1 remained intact when challenged with thermal and hypercapnia stress.
Overall, these results infer that the role of the HOG pathway as a general stress-activated pathway is conserved in S. apiospermum and that SaHog1 responds to stress in an agent-dependent fashion.
3. Discussion
Scedosporium species are typical opportunistic fungi. Infections caused by these fungi are difficult to treat and are associated with high mortality rates when they take place in immunocompromised patients, such as lung transplant recipients [
4]. These fungi are also renowned for being the second most frequent fungal colonizers of the CF airways, after
A. fumigatus [
4]. One questionable aspect that could be raised from these observations is the ability to tolerate immune defenses, especially oxidative-mediated killing, which is a prerequisite to persist in the CF airways. Previous studies in
S. apiospermum showed that the fungus harbor a set of 33 genes encoding a battery of putative antioxidant enzymes, of which the expression was distinctly induced by exposure to oxidative stress derived from chemicals or phagocytes [
10,
11,
12,
22]. These enzymes arguably constitute one side of commander-effector networks of signaling pathways. On the other side, the natural occurrence of these fungi in polluted regions [
2] also denotes the existence of signaling pathways orchestrating protective responses against unpredictable events. These pathways have never been addressed in
Scedosporium. Knowledge gathered on fungal signaling pathways functioning during stress prioritized starting the investigations with the HOG pathway since it is the most cardinal general stress regulator in fungi [
13]. We intended, therefore, to identify the role of the HOG pathway in the tolerance of
S. apiospermum to stresses that are likely to be encountered in hostile environments, with particular attention to oxidative stress. Probing the phosphorylation of SaHog1 in conditions of stress using Western blot appeared to be a reliable way to verify its involvement. A bioinformatic analysis in the
S. apiospermum IHEM 14462 genome was required to ensure the dosage of gene-encoding SaHog1 and its conservation degree. We showed that SaHog1 is highly identical to fungal Hog1 regardless of the harboring species, which agrees with the high conservation degrees of Hog1 in fungi [
15]. Western blot experiments demonstrated that SaHog1 is a multi-stress-responding MAPK kinase as it boosts its activity upon a variety of stressful conditions, including oxidants (H
2O
2, menadione, cumene hydroperoxide, diamide, paraquat, and honokiol), osmotic agents (sorbitol and KCl), cell-wall stress agents (caffeine, calcofluor white, and Congo Red), heavy metals (cadmium and arsenite), fungicides (fludioxonil and iprodione), antifungals (voriconazole and amphotericin B), and acid stress (pH 4). This is consistent with the fact that many of these conditions have already been identified as activators of Hog1 in other fungal species, which were recently reviewed elsewhere [
13,
14]. To the best of our knowledge, we report for the first time that voriconazole is an activator of the HOG pathway in fungi.
The other interesting concept we exposed here is the diverse kinetics of stress-induced phosphorylation of SaHog1. This occurred even within the same group of stress, such as in the case of oxidative, parietal, heavy metal, and fungicide stress. For each of these groups, the agents tested display their proper mode of action, which reasonably drives stress-specific responses, and perhaps the stress-specific activation kinetics of SaHog1. Taking oxidative stress as an example, genetic screening studies performed in
S. cerevisiae helped to unveil the mechanisms required to respond to such stress. The inorganic peroxide H
2O
2 leads to the formation of reactive hydroxyl radicals OH
• via Fenton and Haber–Weiss reactions. Therefore, a protective response heavily depends on genes with mitochondrial respiratory functions. Likewise, honokiol is believed to induce mitochondrial dysfunction, which leads to the increased intracellular accumulation of superoxide anion O
2•− and the activation of H
2O
2-related transcriptional response in fungi [
23,
24,
25,
26,
27]. As for cumene, it is an organic hydroperoxide inducing lipid oxidation; therefore, yeast cells require genes with functions in transcriptional regulation and vacuolar acidification [
28]. In contrast, menadione and paraquat are redox-cycling compounds that transfer electrons from electron carriers to the oxygen molecule to produce O
2•−. Surviving their odd effect requires a functional pentose phosphate pathway able to maintain NADPH flux [
28]. Diamide depletes the pool of reduced glutathione by oxidizing thiol groups. In response, yeast cells employ endoplasmic reticulum protein sorting and vacuolar protein targeting and translocation [
28]. Hence, we postulate that different kinetics observed here reflect that SaHog1 plays distinct regulatory roles. Moreover, sorbitol and KCl, which are believed to act osmotically the same, generated comparable kinetics. In fact, although osmotic stress agents are chemically different, their main effect on cells is the loss of turgor pressure. It was shown that the nuclear enrichment of
S. cerevisiae Hog1 occurs within tens of seconds post-osmotic shock and gradually decreases as soon as turgor pressure begins to restore, and this adaptation was not specific to the chemical nature of the stimulus [
29].
Another hypothesis that explains the distinct kinetics of phosphorylation of SaHog1 is that some stress types prompt negative and/or positive interplays between the HOG pathway and other regulatory pathways, which would affect the activities of each pathway throughout the time of exposure to stress. Reports in this regard highlight mutualistic relationships between the fungal HOG pathway and the Cell-Wall Integrity (CWI) pathway, thus allowing a cross-protection against cell-wall disturbing agents but also cadmium and heat upshift [
13,
30]. Taking this into account, one cannot disregard that the discrete activation effect seen for Congo Red and cadmium stress may reflect opposing interplays between the SaHOG and SaCWI pathways, in which the activity of SaHog1 is weakened when the other pathway is activated.
The activation of SaHog1 to fungicides like fludioxonil was expected. From a mechanistic point of view, the drug is toxic to fungal cells due to the hyperactivation of Hog1 mediated by HK group III that is characterized by HAMP domains (histidine kinases/adenylyl cyclases/methyl-accepting receptors/phosphatases) [
13,
31]. Unpublished work revealed that among the eight genes encoding HKs in the genome of
S. apiospermum IHEM 14462, one encodes a group III HK, which justifies the activation of SaHog1 to the drug. Antifungals like fluconazole, caspofungin, and amphotericin B have already been reported as activators of fungal Hog1, but the involved mechanisms remain overlooked [
13]. It is worth mentioning that the activation of SaHog1 to amphotericin B and voriconazole may be due to drug-associated secondary effects rather than to the mode of action. For instance, amphotericin B is known to promote nascent intracellular reactive oxygen species (ROS), although its fungicidal activity is only due to interaction with ergosterol [
32]. Fluconazole was also shown to induce ROS [
32]. Whether the same applies to voriconazole-induced activation of SaHog1 needs further investigation.
With regard to the responsiveness of SaHog1 to stress, we may postulate that the SaHOG pathway participates in the occurrence of these fungi in nature by rendering them recalcitrant to toxic events. The pathway may also contribute to fungal persistence in CF lungs since both oxidative and acid stress, which are characteristic of CF airways, induced the phosphorylation of SaHog1. Nevertheless, based on current findings, it is still premature to draw any definitive conclusion. This work should be ensued with functional genomic experiments to allow the identification of the downstream effectors of the SaHOG pathway, such as transcription factors and regulated genes. In addition, the identification of the upstream regulators of the HOG pathway will also help dissect whether stress signals are always relayed via TCS or non-canonical routes.
4. Materials and Methods
4.1. Identification of HOG1 Gene
The protein sequence of the Hog1 of the budding yeast S. cerevisiae S288C was used as a query input against S. apiospermum IHEM 14462 in BLAST (taxid: 563466). Once the SaHOG1 gene was identified, the relative protein sequence was then blasted against the harboring genome to check the presence of paralogs. Cladogram analysis of fungal Hog1 was performed using Clustal Omega.
4.2. Culture and Stress of Fungi
The strain S. apiospermum IHEM 14462 was cultured for 9 days at 37 °C on yeast extract-peptone-dextrose agar (YPDA, containing in g/L: yeast extract, 5; pancreatic peptone, 10; glucose, 20; bacteriological agar, 20; and chloramphenicol, 0.5). The conidia were sterilely harvested in distilled water and filtered using Miracloth membranes. Fifteen microliters of the conidial suspension (2.5 × 102 conidia/mL) were spread on sterile cellophane membranes (autoclaved in water) laid on the surface of YPDA plates. The plates were then incubated for 7 days at 37 °C in the dark (to sidestep the potential light-induced phosphorylation of Hog1). Cellophane membranes, on which fungal colonies were grown, were then delicately removed from the original plates and placed on freshly prepared YPDA plates supplemented or not with the stressful agents. Tested stress included osmotic, oxidative, parietal, heavy metals, acid, and antifungals. The plates were reversed and left at room temperature (25 °C) and in the dark for 15, 30, 60, or 120 min. In the case of stress with high temperature (45 °C), colonies-containing cellophane membranes were laid on new YPDA plates that had already been incubated at 45 °C for a short time (30 min), and then the plates were left lidless and re-incubated at 45 °C, in the dark, for the indicated times. The plates were also left lidless in case of hypercapnia stress (5% CO2). Afterwards, the cellophane membranes were removed, covered with aluminum foils, and put in liquid nitrogen. The samples were then lyophilized for 48 h and stored at −80 °C until use. Lyophilized colonies were put in 1.5 mL microtubes (100 μg of fungal biomass per tube) and proceeded to total protein extraction.
4.3. Protein Extraction and Electrophoresis
One milliliter of ice-cold, freshly prepared extraction Laemmli buffer (62.5 mM Tris, 2% SDS, 10% glycerol, and 2.5% β-mercaptoethanol) supplemented with 1 mM phenylmethyl sulfonyl fluoride (PMSF) was added to sample tubes. After the addition of glass beads, tubes were vigorously shaken in a Mini-bead beater (RetschTM MM 200) 4 times for 30 s with 30 s of incubation on ice in between. Tubes were then centrifuged at 15,000 rpm for 15 min at 4 °C, and the supernatants were transferred to a new 1.5 mL microtube tube. Protein samples were stored at −20 °C until use. The protein concentration was determined using Pierce™ Microplate BCA Protein Assay Kit—Reducing Agent Compatible (Thermo Fisher Scientific). For protein separation using SDS-polyacrylamide gel electrophoresis, samples were diluted in loading buffer and heated in boiling water for 2 min, and equal amounts of proteins (75 μg) were deposited into the wells of 10% SDS-polyacrylamide gels. The ladder used was PageRuler™ Plus Prestained Protein Ladder (Thermo Fisher Scientific). Gels were then electro-transferred to an Immuno-blot PVDF membrane (Bio-Rad) and blocked with 5% bovine serum albumin (BSA) prepared in phosphate-buffered saline (PBS). The membranes were left overnight at 4 °C in the blocking buffer and then washed 3 times (PBS with 0.05% Tween-20) for 5 min before the performance of immunoblotting.
4.4. Western Blot
The anti-Hog1 (y-215) polyclonal antibody (sc-9079, Santa Cruz Biotechnology), kindly given by Pr. Thomas Guillemette (IRHS, Angers, France), was used to detect total SaHog1 at a 1:1000 dilution in PBS containing 0.3% BSA and 0.05% Tween-20. Given that the probing of total Hog1 serves as a loading control, we also ensured the validity of the anti-Hog1 (y-215) by probing in parallel the actin amount using as the primary antibody a rabbit anti-β-actin polyclonal antibody (GTX109639, GeneTex) diluted to 1:5000 in the same dilution buffer. To detect phosphorylated SaHog1, the phospho-p38 MAP kinase polyclonal antibody (9211S, Cell-signaling Technology) was used as the primary antibody at a 1:1000 dilution. The membranes were incubated with the primary antibodies for 1 h at room temperature under gentle agitation. After washing 3 times with the washing solution (PBS containing 0.05% Tween-20), the membranes were incubated for 1 h at room temperature with horseradish peroxidase-conjugated anti-rabbit IgG antibodies (7074S, Cell signaling Technology) used at a 1:1000 dilution. Signals were detected by chemiluminescence on LAS4000 (GE Healthcare) after a 5 min incubation of the membranes with Clarity™ Western ECL Substrate (Bio-Rad).


 ,
, {kind=link}
{kind=link}
{kind=link}


