
Population Dynamics of Digitaria sanguinalis and Effects on Soybean Crop under Different Glyphosate Application Timings


Abstract
:1. Introduction
2. Materials and Methods
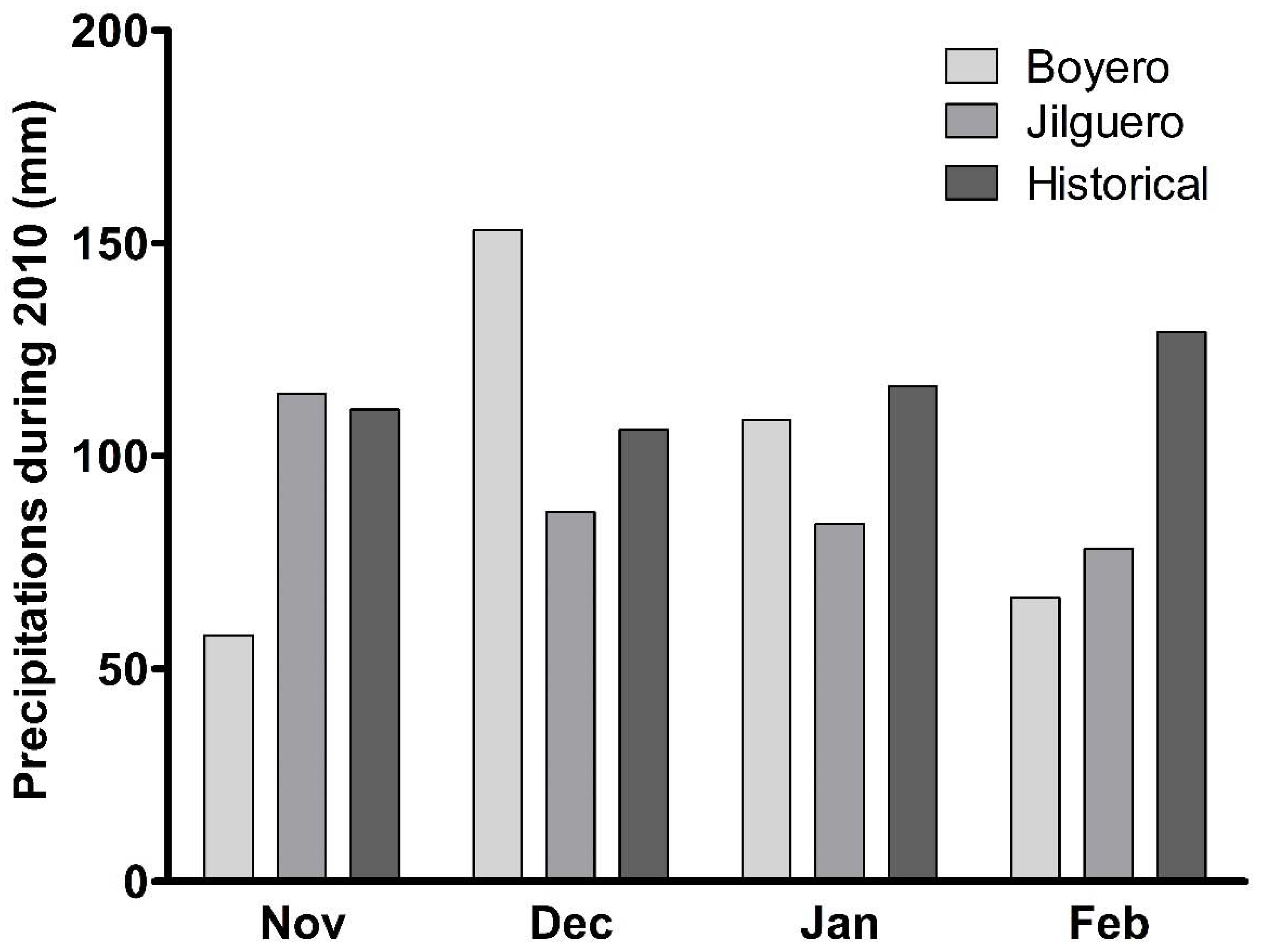
2.1. Experimental Site
2.2. Experiments
2.3. Measurements
2.4. Data Analysis
3. Results
3.1. Digitaria Sanguinalis Population Dynamics
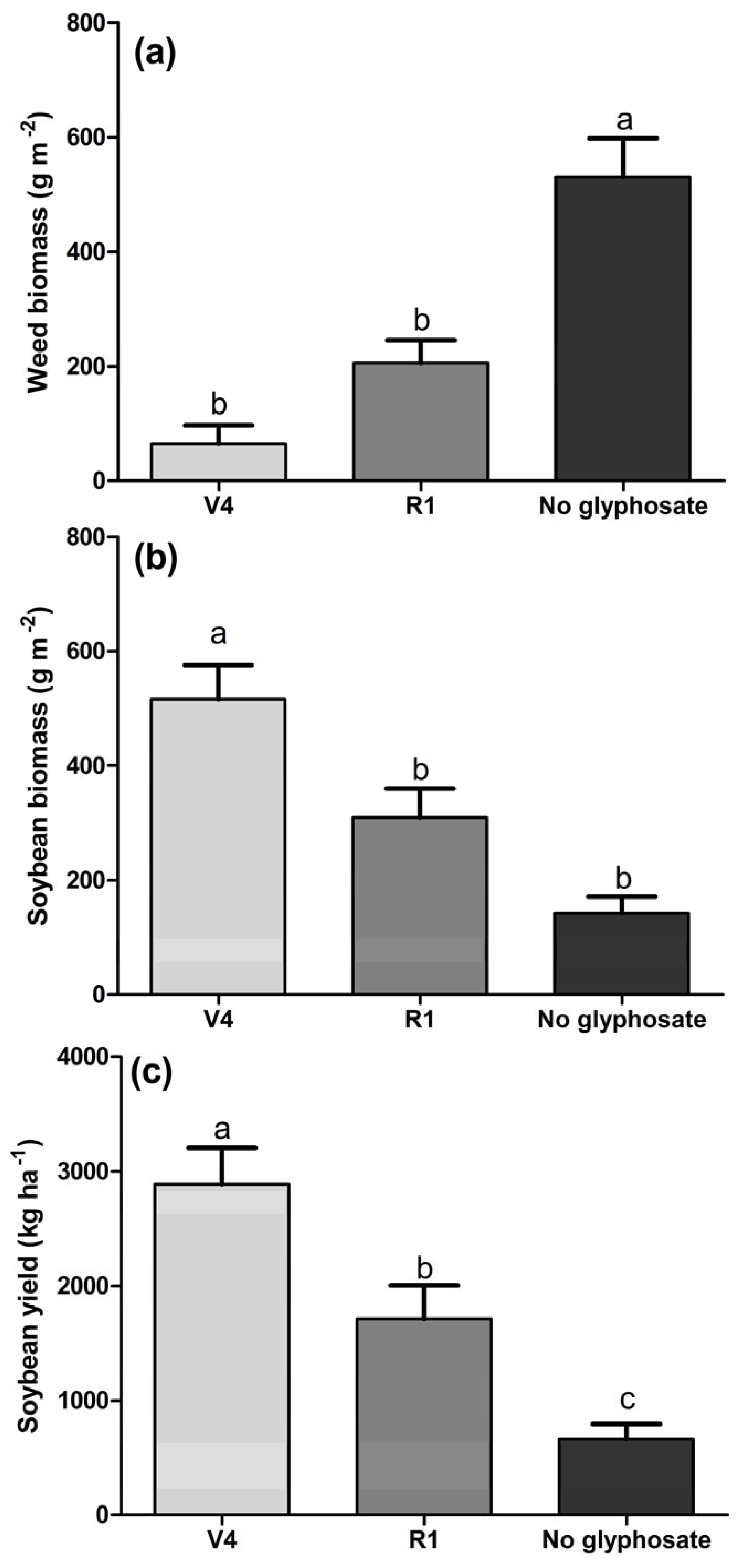
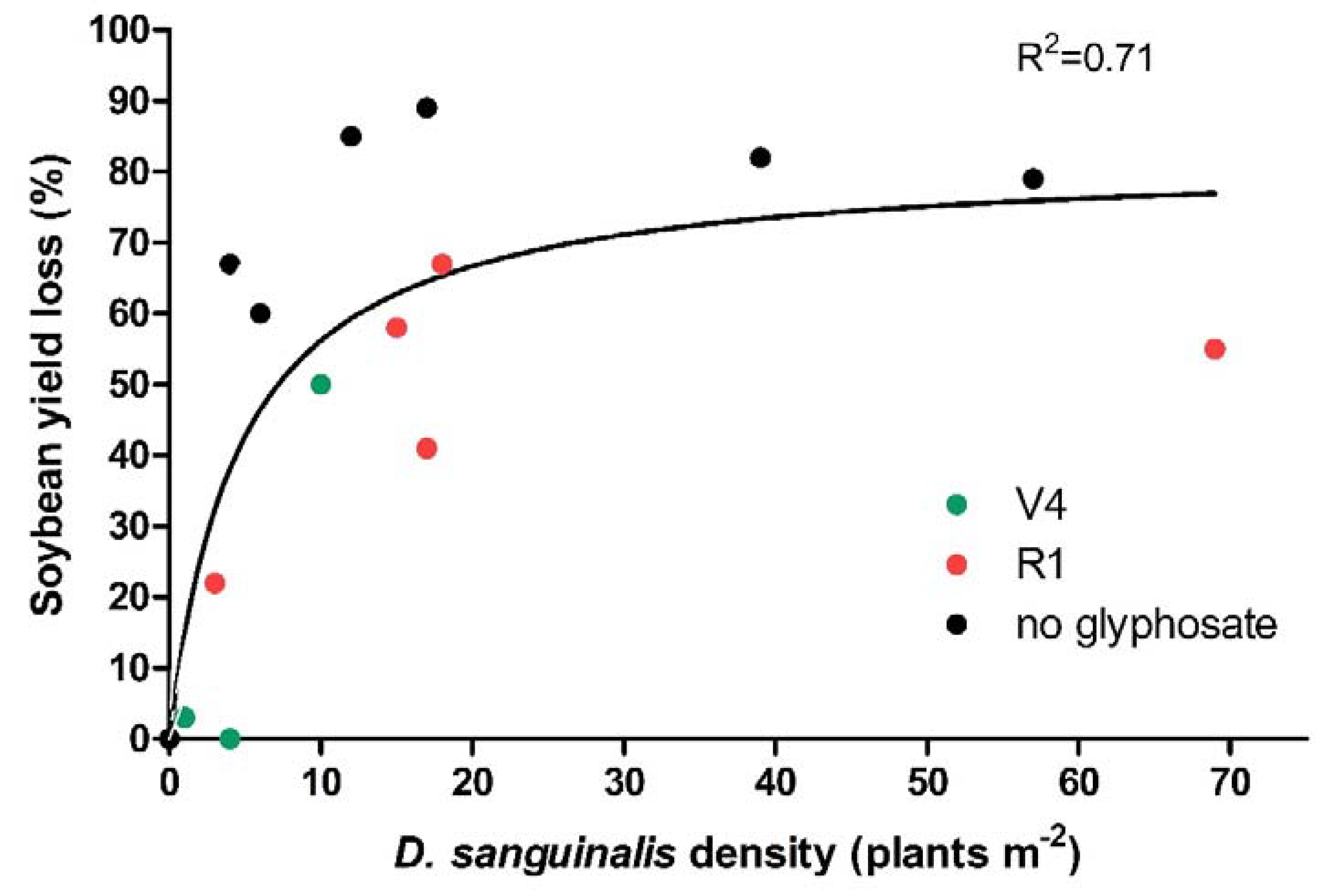
3.2. Effects on Soybean
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jones, E.A.; Contreras, D.J.; Everman, W.J. Digitaria ciliaris, Digitaria ischaemum, and Digitaria sanguinalis. In Biology and Management of Problematic Crop Weed Species; Chauhan, B.S., Ed.; Academic Press: San Diego, CA, USA, 2021; pp. 173–195. [Google Scholar]
- Scursoni, J.A.; Vera, A.C.D.; Oreja, F.H.; Kruk, B.C.; de la Fuente, E.B. Weed management practices in Argentina crops. Weed Technol. 2019, 33, 459–463. [Google Scholar] [CrossRef]
- Gianessi, L.P. The increasing importance of herbicides in worldwide crop production. Pest Manag. Sci. 2013, 69, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Culpepper, A.S.; Gimenez, A.E.; York, A.C.; Batts, R.B.; Willcut, J.W. Morningglory (Ipomoea spp.) and Large Crabgrass (Digitaria sanguinalis) control with glyphosate and 2,4-DB mixtures in glyphosate-resistant soybean (Glycine max). Weed Technol. 2001, 15, 56–61. [Google Scholar] [CrossRef]
- Van Gessel, M.J.; Ayeni, A.O.; Majek, B.A. Glyphosate in full-season in no-till glyphosate-resistant soybean: Role of pre-plant applications and residual herbicides. Weed Technol. 2001, 15, 714–724. [Google Scholar] [CrossRef]
- Puricelli, E.; Tuesca, D. Weed density and diversity under glyphosate-resistant crop sequences. Crop Prot. 2005, 24, 533–542. [Google Scholar] [CrossRef]
- De la Fuente, E.B.; Suárez, S.A.; Ghersa, C.M. Soybean weed community composition and richness between 1995 and 2003 in the Rolling Pampas (Argentina). Agric. Ecosyst. Environ. 2006, 115, 229–236. [Google Scholar] [CrossRef]
- Scursoni, J.A.; Satorre, E.H. Glyphosate management strategies, weed diversity and soybean yield in Argentina. Crop Prot. 2010, 29, 957–962. [Google Scholar] [CrossRef]
- De la Fuente, E.B.; Oreja, F.H.; Lenardis, A.E.; Torcat Fuentes, M.; Agosti, M.B.; Barrio, A.; Barberis, S.; Robredo, J.; Gil, A.; Marzetti, M.; et al. Intensification of crop rotation affecting weed communities and the use of herbicides in the rolling Pampa. Heliyon 2021, 7, e06089. [Google Scholar] [CrossRef]
- Yanniccari, M.; Vázquez-García, J.G.; Gigón, R.; Palma-Bautista, C.; Vila-Aiub, M.; Prado, R.D. A novel EPSPS Pro-106-His mutation confers the first case of glyphosate resistance in Digitaria sanguinalis. Pest Manag. Sci. 2022, 78, 3135–3143. [Google Scholar] [CrossRef]
- Norris, R.F. Weed fecundity: Current status and future needs. Crop Prot. 2007, 26, 182–188. [Google Scholar] [CrossRef]
- Harrison, S.K.; Regnier, E.E.; Schmoll, J.T. Postdispersal predation of giant ragweed (Ambrosia trifida) seed in no-tillage corn. Weed Sci. 2003, 51, 955–964. [Google Scholar] [CrossRef]
- Gallart, M.; Mas, M.T.; Verdu, A.M.C. Demography of Digitaria sanguinalis: Effect of the emergence time on survival, reproduction and biomass. Weed Biol. Manag. 2010, 10, 132–140. [Google Scholar] [CrossRef]
- Cardina, J.; Herms, C.P.; Herms, D.A. Phenological indicators for emergence of Large and Smooth Crabgrass (Digitaria sanguinalis and D. ischaemun). Weed Technol. 2011, 25, 141–150. [Google Scholar] [CrossRef]
- Everman, W.J.; Clewis, S.B.; York, A.C.; Wilcut, J.W. Weed control and yield with flumioxazin, fomesafen, and S-metolachlor systems for glufosinate-resistant cotton residual weed management. Weed Technol. 2009, 23, 391–397. [Google Scholar] [CrossRef]
- Cahoon, C.W.; York, A.C.; Jordan, D.L.; Everman, W.J.; Seagroves, R.W.; Braswell, L.R.; Jennings, K.M. Weed control in cotton by combinations of microencapsulated acetochlor and various residual herbicides applied preemergence. Weed Technol. 2015, 29, 740–750. [Google Scholar] [CrossRef]
- Stewart, C.L.; Nurse, R.E.; Hamill, A.S.; Sikkema, P.H. Environment and soil conditions influence pre-and postemergence herbicide efficacy in soybean. Weed Technol. 2010, 24, 234–243. [Google Scholar] [CrossRef]
- Farha, W.; AM, A.E.A.; Rahman, M.D.; Shin, H.C.; Shim, J.H. An overview on common aspects influencing the dissipation pattern of pesticides: A review. Environ. Monit. Assess. 2016, 188, 693. [Google Scholar] [CrossRef] [PubMed]
- Oreja, F.H.; Batlla, D.; de la Fuente, E.B. Digitaria sanguinalis seed dormancy release and seedling emergence are affected by crop canopy and stubble. Weed Res. 2020, 60, 111–120. [Google Scholar] [CrossRef]
- Swanton, C.J.; Mahoney, K.J.; Chandler, K.; Gulden, R.H. Integrated weed management: Knowledge-based weed management systems. Weed Sci. 2008, 56, 168–172. [Google Scholar] [CrossRef]
- Van Acker, C.R.; Swanton, C.J.; Weise, S.E. The critical period of weed control in soybean [Glycine max (L.) Merr.]. Weed Sci. 1993, 41, 194–200. [Google Scholar] [CrossRef]
- Bagavathiannan, M.V.; Norsworthy, J.K. Late-season seed production in arable weed communities: Management implications. Weed Sci. 2012, 60, 325–334. [Google Scholar] [CrossRef]
- Soltani, N.; Stewart, C.L.; Nurse, R.E.; Van Eerd, L.L.; Vyn, R.J.; Sikkema, P.H. Weed control, environmental impact and profitability of weed management strategies in glyphosate-resistant corn. Am. J. Plant Sci. 2012, 3, 1594. [Google Scholar] [CrossRef]
- Beckie, H.J. Herbicide-resistant weed management: Focus on glyphosate. Pest Manag. Sci. 2011, 67, 1037–1048. [Google Scholar] [CrossRef] [PubMed]
- Oreja, F.H.; Inman, M.D.; Jordan, D.L.; Leon, R.G. Population growth rates of weed species in response to herbicide programme intensity and their impact on weed community. Weed Res. 2021, 61, 509–518. [Google Scholar] [CrossRef]
- Davis, A.S.; Liebman, M. Cropping system effects on giant foxtail (Setaria faberi) demography, I: Green manure and tillage timing. Weed Sci. 2003, 51, 919–929. [Google Scholar] [CrossRef]
- Caswell, H. Matrix Population Models: Construction, Analysis and Interpretation; Sinauer: Sunderland, MA, USA, 2001; pp. 133–174. [Google Scholar]
- González-Andújar, J.L.; Fernández-Quintanilla, C. Modelling the population dynamics of annual ryegrass (Lolium rigidum) under various weed management systems. Crop Prot. 2004, 23, 723–729. [Google Scholar] [CrossRef]
- Bussan, A.J.; Boerboom, C.M.; Stoltenberg, D.E. Response of Setaria faberi demographic processes to herbicide rates. Weed Sci. 2000, 48, 445–453. [Google Scholar] [CrossRef]
- De la Fuente, E.B.; Suárez, S.A.; Lenardis, A.E.; Oreja, F.H.; Torcat Fuentes, M. Cambios en las comunidades de malezas en los cultivos de maíz de la pampa ondulada (Argentina) entre 1960 y 2019. Agron. Y Ambiente Rev. Fac. Agron. Univ. Buenos Aires 2021, 41, 169–178. [Google Scholar]
- Fernández-Quintanilla, C. Studying the population dynamics of weeds. Weed Res. 1988, 28, 443–447. [Google Scholar] [CrossRef]
- Sagar, G.R. An approach to the study of the population dynamics of plants with special reference to weeds. Ann. Appl. Biol. 1976, 1, 1–47. [Google Scholar]
- Cousens, R. An empirical model relating crop yield to weed and crop density and a statistical comparison with other models. J. Agric. Sci. 1985, 105, 513–521. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: https://www.R-project.org/ (accessed on 10 July 2022).
- González-Andújar, J.L.; Fernández-Quintanilla, C. Modelling the population dynamics of Avena sterilis under dry-land cereal cropping systems. J. Appl. Ecol. 1991, 28, 16–27. [Google Scholar] [CrossRef]
- Oreja, F.H.; de la Fuente, E.B.; Batlla, D. Role of seed environment and covering structures on large crabgrass germination. S. Afr. J. Bot. 2017, 111, 170–175. [Google Scholar] [CrossRef]
- Mohler, C.L.; Callaway, M.B. Effects of tillage and mulch on weed seed production and seed banks in sweet corn. J. Appl. Ecol. 1995, 32, 627–639. [Google Scholar] [CrossRef]
- Scursoni, J.; Gastaldi, E. Demografía de Pasto cuaresma (Digitaria sanguinalis) en cultivos de soja de segunda, sembrados en sistema de siembra directa. In Proceedings of the XIII Congreso Latinoamericano de Malezas, Buenos Aires, Argentina, 17–19 September 1997. [Google Scholar]
- Norsworthy, J.K. Soybean canopy formation effects on pitted morningglory (Ipomoea lacunosa), common cocklebur (Xanthium strumarium), and sicklepod (Senna obtusifolia) emergence. Weed Sci. 2004, 52, 954–960. [Google Scholar] [CrossRef]
- Oreja, F.H.; Batlla, D.; de la Fuente, E.B. Effect of soybean crop structure on large crabgrass (Digitaria sanguinalis) growth and seed dormancy. Weed Sci. 2021, 69, 372–378. [Google Scholar] [CrossRef]
- Knezevic, S.Z.; Weise, S.F.; Swanton, C.L. Interference of redroot pigweed (Amaranthus retroflexus) in corn (Zea mays). Weed Sci. 1994, 42, 568–573. [Google Scholar] [CrossRef]
- Steckel, L.E.; Sprague, C.L. Late-season common waterhemp (Amaranthus rudis) interference in narrow- and wide-row soybean. Weed Technol. 2004, 18, 947–952. [Google Scholar] [CrossRef]
- Norsworthy, J.K.; Jha, P.; Bridges, W., Jr. Sicklepod survival and fecundity in wide- and narrow-row glyphosate-resistant soybean (Glycine max). Weed Sci. 2007, 55, 252–259. [Google Scholar] [CrossRef]
- Hoss, N.E.; Al-Khatib, K.; Peterson, D.E.; Loughin, T.M. Efficacy of glyphosate, glufosinate, and imazethapyr on selected weed species. Weed Sci. 2003, 51, 110–117. [Google Scholar] [CrossRef]
- DeGreeff, R.D.; Varanasi, A.V.; Dille, J.A.; Peterson, D.E.; Jugulam, M. Influence of plant growth stage and temperature on glyphosate efficacy in common lambsquarters (Chenopodium album). Weed Technol. 2018, 32, 448–453. [Google Scholar] [CrossRef]
- Mulugeta, D.; Boerboom, C.M. Critical time of weed removal in glyphosate-resistant Glycine max. Weed Sci. 2000, 48, 35–42. [Google Scholar] [CrossRef]
- Jha, P.; Norsworthy, J.K.; Bridges, W.; Riley, M.B. Influence of glyphosate timing and row width on Palmer amaranth (Amaranthus palmeri) and pusley (Richardia spp.) demographics in glyphosate-resistant soybean. Weed Sci. 2008, 56, 408–415. [Google Scholar] [CrossRef]
- Weiner, J. Allocation, plasticity and allometry in plants. Perspect. Plant Ecol. Evol. Syst. 2004, 6, 207–215. [Google Scholar] [CrossRef]
- Williams, M.M.; Boydston, R.A.; Davis, A.S. Crop competitive ability contributes to herbicide performance in sweet corn. Weed Res. 2008, 48, 58–67. [Google Scholar] [CrossRef]
- Green-Tracewicz, E.; Page, E.R.; Swanton, C.J. Shade avoidance in soybean reduces branching and increases plant-to-plant variability in biomass and yield per plant. Weed Sci. 2011, 59, 43–49. [Google Scholar] [CrossRef]
- Board, J.E.; Harville, B.G. Soybean yield component responses to a light interception gradient during the reproductive period. Crop Sci. 1993, 33, 772–777. [Google Scholar] [CrossRef]
- Knezevic, S.Z.; Evans, S.P.; Blankenship, E.E.; Van Acker, R.C.; Lindquist, J.L. Critical period for weed control: The concept and data analysis. Weed Sci. 2002, 50, 773–786. [Google Scholar] [CrossRef]
- Halford, C.; Hamill, A.S.; Zhang, J.; Doucet, C. Critical period of weed control in no-till soybean (Glycine max) and corn (Zea mays). Weed Technol. 2001, 15, 737–744. [Google Scholar] [CrossRef]
- Eyherabide, J.J.; Cendoya, M.G. Critical periods of weed control in soybean for full field and in-furrow interference. Weed Sci. 2002, 50, 162–166. [Google Scholar] [CrossRef]
- Keramati, S.; Pirdashti, H.; Esmaili, M.A.; Abbasian, A.; Habibi, M. The Critical Period of Weed Control in Soybean (Glycine max (L.) Merr.). Pak. J. Biol. Sci. 2008, 11, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, D.E.; Landa, K. Competitive effect and response: Hierarchies and correlated traits in the early stages of competition. J. Ecol. 1991, 79, 1013–1030. [Google Scholar] [CrossRef] [Green Version]
- Aguyoh, J.N.; Masiunas, J.B. Interference of large crabgrass (Digitaria sanguinalis) with snap beans. Weed Sci. 2003, 51, 171–176. [Google Scholar] [CrossRef]
- Fu, R.; Ashley, R.A. Interference of large crabgrass (Digitaria sanguinalis), redroot pigweed (Amaranthus retroflexus), and hairy galinsoga (Galinsoga ciliata) with bell pepper. Weed Sci. 2006, 54, 364–372. [Google Scholar] [CrossRef]
- Basinger, N.T.; Jennings, K.M.; Monks, D.W.; Jordan, D.L.; Everman, W.J.; Hestir, E.L.; Waldschmidt, M.D.; Smith, S.C.; Brownie, C. Interspecific and intraspecific interference of Palmer amaranth (Amaranthus palmeri) and large crabgrass (Digitaria sanguinalis) in sweetpotato. Weed Sci. 2019, 67, 426–432. [Google Scholar] [CrossRef]
- Sartorato, I.; Berti, A.; Zanin, G.; Dunan, C.M. Modeling of glyphosate application timing in glyphosate-resistant soybean. Weed Sci. 2011, 59, 390–397. [Google Scholar] [CrossRef]
- Storkey, J.; Neve, P. What good is weed diversity? Weed Res. 2018, 58, 239–243. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| No Glyphosate | Glyphosate at V4 | Glyphosate at R1 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Initial seedbank | 7037A | 5961A | 6893A | ||||||
| Er | 0.17A | 0.11A | 0.15A | ||||||
| Cohort | 1st | 2nd | 3rd | 1st | 2nd | 3rd | 1st | 2nd | 3rd |
| Seedlings | 1175Aa | 167Aab | 0Ab | 621Aa | 17Ab | 25Ab | 721Aa | 200Aab | 88Ab |
| Sr | 0.42Aa | 0.22Aa | 0Aa | 0Ba | 0Aa | 0.5Aa | 0.19Aba | 0.42Aa | 0.40Aa |
| Adult plants | 388Aa | 29Ab | 0Ab | 0Ba | 0Aa | 17Aa | 150Aba | 113Aa | 63Aa |
| Fr | 1556.2Aa | 0.5Ab | 0Ab | 0Ba | 0Aa | 1.7Aa | 3.4B | 0.25A | 0.5A |
| Boyero | Treatments | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| No Glyphosate | Glyphosate at V4 | Glyphosate at R1 | |||||||
| Cohort | 1st | 2nd | 3rd | 1st | 2nd | 3rd | 1st | 2nd | 3rd |
| Tillers per plant | 7.03b | 0.5a | 0a | 0a | 0a | 0.67a | 1.58ab | 0.5a | 0.67a |
| Racemes/tiller | 4.06b | 0.5a | 0a | 0a | 0a | 1a | 1ab | 0.42a | 0.5a |
| Spikelets/raceme | 30.89b | 3a | 0a | 0a | 0a | 5a | 4.25a | 2.42a | 3a |
| Jilguero | Treatments | ||||||||
| No Glyphosate | Glyphosate in V4 | Glyphosate in R1 | |||||||
| Cohort | 1st | 2nd | 3rd | 1st | 2nd | 3rd | 1st | 2nd | 3rd |
| Tillers/plant | 14.25b | 2.67b | 2.67a | 0a | 0a | 0a | 0a | 0a | 0a |
| Racemes/tiller | 4.75b | 2.67a | 1.33a | 0a | 0a | 0a | 0a | 0a | 0a |
| Spikelets/raceme | 33.25b | 14.67a | 9.33a | 0a | 0a | 0a | 0a | 0a | 0a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oreja, F.H.; Stempels, M.; de la Fuente, E.B. Population Dynamics of Digitaria sanguinalis and Effects on Soybean Crop under Different Glyphosate Application Timings. Grasses 2023, 2, 12-22. https://doi.org/10.3390/grasses2010002
Oreja FH, Stempels M, de la Fuente EB. Population Dynamics of Digitaria sanguinalis and Effects on Soybean Crop under Different Glyphosate Application Timings. Grasses. 2023; 2(1):12-22. https://doi.org/10.3390/grasses2010002
Chicago/Turabian StyleOreja, Fernando H., Mateo Stempels, and Elba B. de la Fuente. 2023. "Population Dynamics of Digitaria sanguinalis and Effects on Soybean Crop under Different Glyphosate Application Timings" Grasses 2, no. 1: 12-22. https://doi.org/10.3390/grasses2010002