
Aquatic and Semiaquatic Heteroptera (Hemiptera: Insecta) Distribution in Streams on the Cerrado–Amazon Ecotone in Headwaters of Xingu River

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
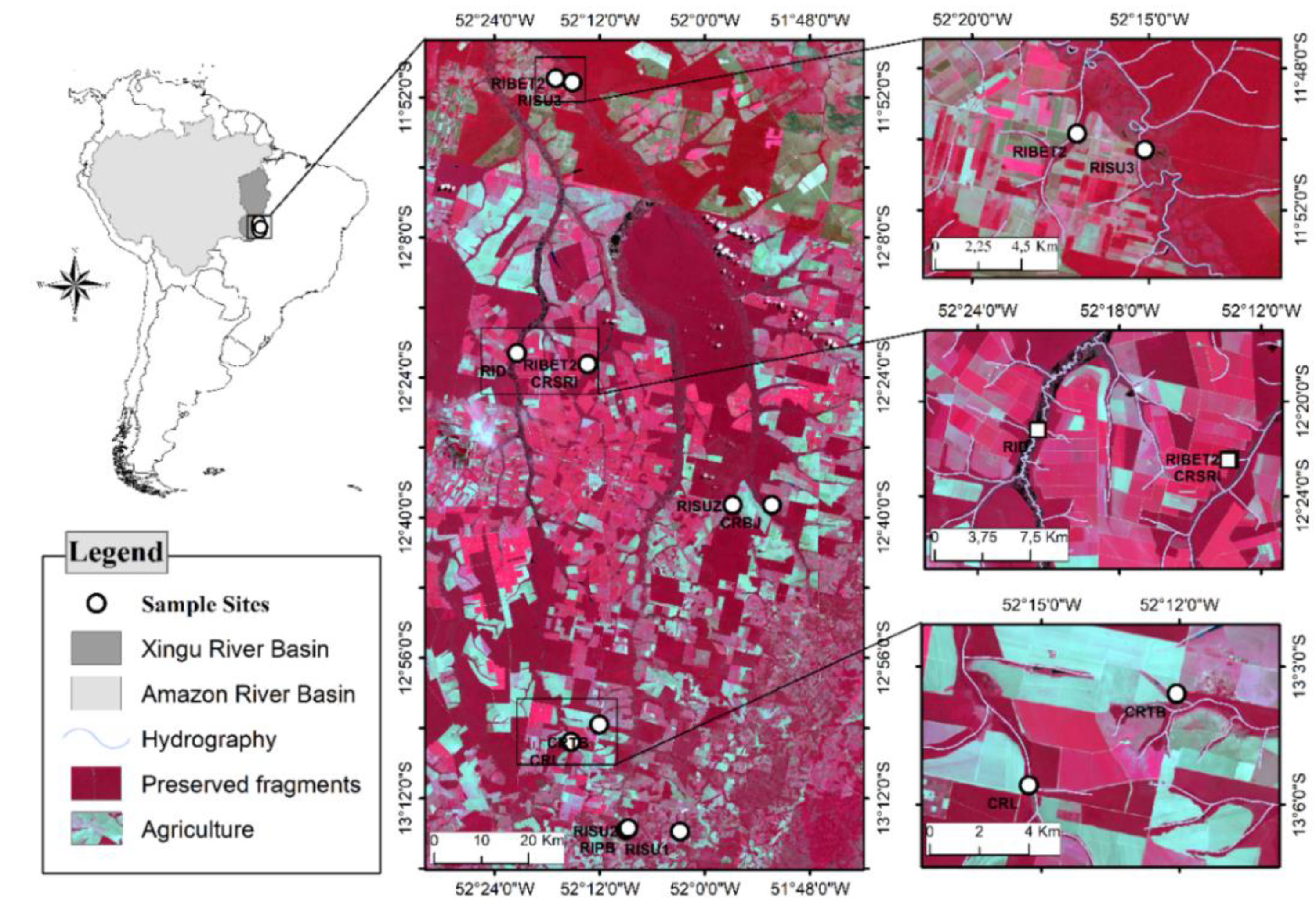
2.1. Study Area
2.2. Data Collection
2.2.1. Biological Sampling
2.2.2. Environmental Variables
2.2.3. Habitat Integrity Sample
2.2.4. Sample of Vegetation Index
- = Reflection of the top of the atmosphere;
- = Band-specific multiplicative scaling factor;
- = Band-specific additive scaling factor;
- = Calibrated standard product pixel;
- = Local elevation angle of the sun.
- NIR = Spectral band of near infrared;
- Red = Red spectral band.
- G = 2.5;
- C1 e C2 = Adjustment coefficients for the effect of aerosols in the atmosphere, being C1 = 6, C2 = 7.5;
- NIR = Spectral band of near infrared;
- Red = Red spectral band;
- Blue = Blue spectral band.
2.3. Data Analysis
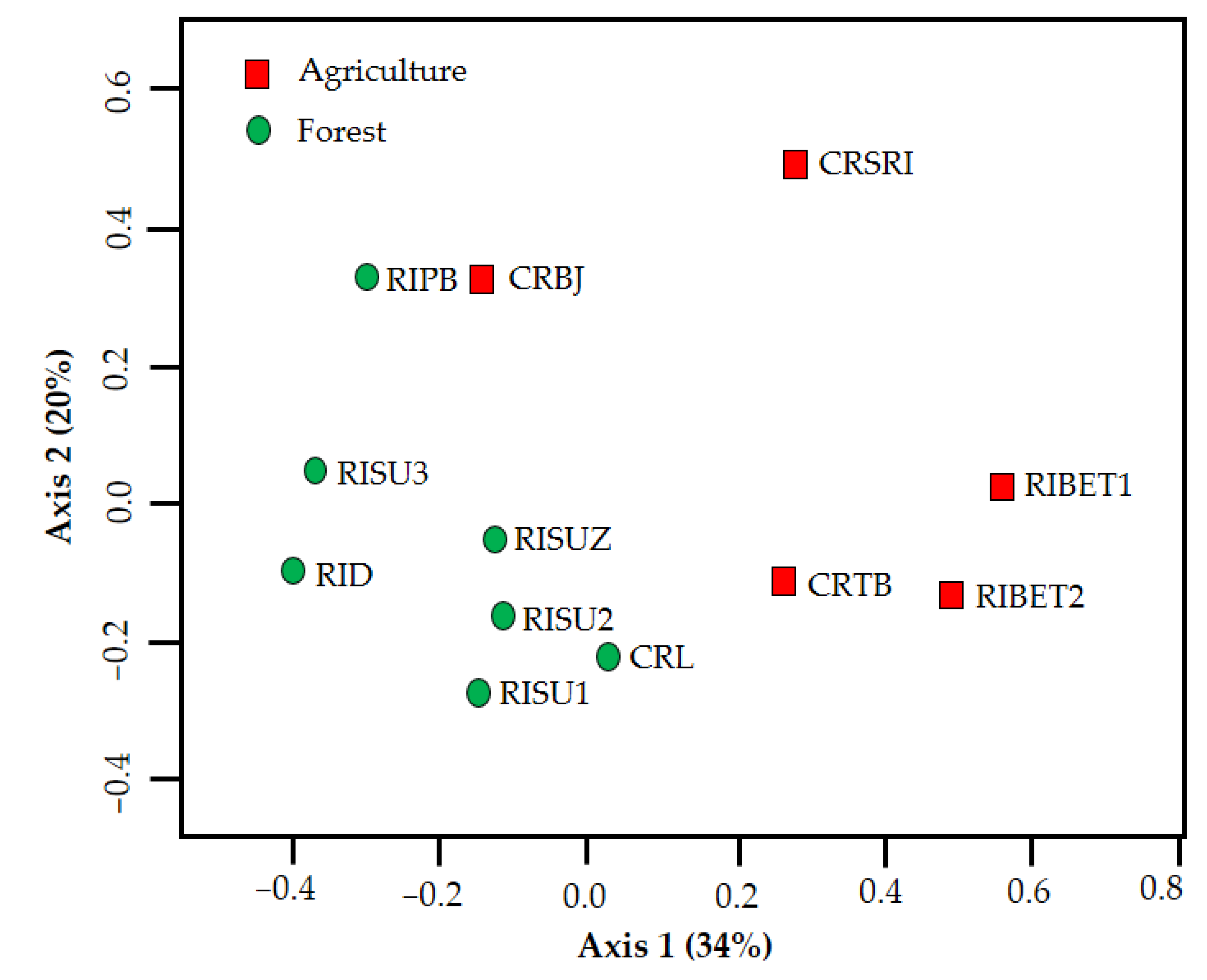
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Allen, J.D. Landscapes and riverscapes: The influence of land use on stream ecosystems. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 257–284. [Google Scholar] [CrossRef]
- Juen, L.; Cunha, E.J.; Carvalho, F.G.; Ferreira, M.C.; Begot, T.O.; Luiza-Andrade, A.; Shimano, Y.; Leão, H.; Pompeu, P.S.; Montag, L.F.A. Effects of oil palm plantations on the habitat structure and biota of streams in Eastern Amazon. River Res. Appl. 2016, 32, 2081–2094. [Google Scholar] [CrossRef]
- Allan, J.D.; Castilho, M.M. Stream Ecology: Structure and Function of Running Waters, 2nd ed.; Springer: Amsterdam, The Netherlands, 2007; 436p. [Google Scholar]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Thorp, J.H.; Thoms, M.C.; Delong, M.D. The riverine ecosystem synthesis: Biocomplexity in river networks across space and time. River Res. Appl. 2006, 22, 123–147. [Google Scholar] [CrossRef]
- Brito, J.G.; Roque, F.O.; Martins, R.T.; Nessimian, J.L.; Oliveira, V.C.; Hughes, R.M.; De Paula, F.R.; Silvio, F.B.F.; Hamada, N. Small forest losses degrade stream macroinvertebrate assemblages in the eastern Brazilian Amazon. Biol. Conserv. 2020, 241, 108263. [Google Scholar] [CrossRef]
- Dala-Corte, R.B.; Melo, A.S.; Siqueira, T.; Bini, L.M.; Martins, R.T.; Cunico, A.M.; Pes, A.M.; Magalhães, A.L.B.; Godoy, B.S.; Leal, C.G.; et al. Thresholds of freshwater biodiversity in response to riparian vegetation loss in the Neotropical region. J. Appl. Ecol. 2020, 57, 1391–1402. [Google Scholar] [CrossRef]
- Bacellar, L.A.P. O papel das florestas no regime hidrológico de bacias hidrográficas. Geo. br 2005, 1, 1–39. [Google Scholar]
- Bispo, P.C.; Oliveira, L.G.; Bini, L.M.; Sousa, K.G. Ephemeroptera, Plecoptera and Trichoptera assemblages from riffles in mountain streams of Central Brazil: Environmental factors influencing the distribution and abundance of immatures. Braz. J. Biol. 2006, 66, 611–622. [Google Scholar] [CrossRef]
- Martins, R.T.; Couceiro, S.R.M.; Melo, A.S.; Moreira, M.P.; Hamada, N. Effects of urbanization on stream benthic invertebrate communities in Central Amazon. Ecol. Indic. 2017, 73, 480–491. [Google Scholar] [CrossRef]
- Bonada, N.; Prat, N.; Resh, V.H.; Statzner, B. Developments in aquatic insect biomonitoring: A comparative analysis of recent approaches. Annu. Rev. Entomol. 2006, 5, 495–523. [Google Scholar] [CrossRef]
- Lammert, M.; Allan, J.D. Assessing biotic integrity of streams: Effects of scale in measuring the influence of land use/cover and habitat structure on fish and macroinvertebrates. Environ. Manag. 1999, 23, 257–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brando, P.M.; Coe, M.; Defries, R.; Azevedo, A.A. Ecology, economy and management of an agroindustrial frontier landscape in the southeast Amazon. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120152. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.; Martins, I.S.; Kastner, T.; Plutzar, C.; Theurl, M.C.; Eisenmenger, N.; Huijbregts, M.A.J.; Wood, R.; Stadler, K.; Bruckner, M.; et al. Increasing impacts of land use on biodiversity and carbon sequestration driven by population and economic growth. Nat. Ecol. Evol. 2019, 3, 628–637. [Google Scholar] [CrossRef] [PubMed]
- Montag, L.F.; Winemiller, K.O.; Keppeler, F.W.; Leão, H.; Benone, N.L.; Torres, N.R.; Prudente, B.S.; BeGot, T.O.; Bower, L.M.; Saenz, D.E.; et al. Land cover, riparian zones and instream habitat influence stream fish assemblages in the eastern Amazon. Ecol. Freshw. Fish 2019, 28, 317–329. [Google Scholar] [CrossRef]
- Nogueira, D.S.; Cabette, H.S.R.; Juen, L. Estrutura e composição da comunidade de Trichoptera (Insecta) de rios e áreas alagadas da bacia do rio Suiá-Miçú, Mato Grosso, Brasil. Iheringia Sér. Zool. 2011, 101, 173–180. [Google Scholar] [CrossRef]
- Shimano, Y.; Cabette, H.S.R.; Salles, F.F.; Juen, L. Composição e distribuição da fauna de Ephemeroptera (Insecta) em área de transição Cerrado-Amazônia, Brasil. Iheringia Sér. Zool. 2010, 100, 301–308. [Google Scholar] [CrossRef]
- Fischer, J.; Lindenmayer, D.B. Landscape modification and habitat fragmentation: A synthesis. Glob. Ecol. Biogeogr. 2007, 16, 265–280. [Google Scholar] [CrossRef]
- Rosenberg, D.M.; Resh, V.H. Freshwater Biomonitoring and Benthic Macroinvertebrates; Chapman & Hall: New York, NY, USA, 1993; 488p. [Google Scholar]
- Giehl, N.F.; Dias-Silva, K.; Juen, L.; Batista, J.D.; Cabette, H.S.R. Taxonomic and numerical resolutions of Nepomorpha (Insecta: Heteroptera) in cerrado streams. PLoS One 2014, 9, e103623. [Google Scholar] [CrossRef]
- Dias-Silva, K.; Brasil, L.S.; Veloso, G.K.O.; Cabette, H.S.R.; Juen, L. Land use change causes environmental homogeneity and low beta-diversity in Heteroptera of streams. Ann. Limnol. Int. J. Lim. 2020, 56, 9. [Google Scholar] [CrossRef]
- Moy, K.M.; Brasil, L.S.; Oliveira-Junior, J.M.B.; Juen, L.; Vieira, T.B.; Dias-Silva, K. Effects of Environmental Changes on Gerromorpha (Heteroptera: Hemiptera) Communities from Amazonian Streams. Hydrobiology 2022, 1, 8. [Google Scholar] [CrossRef]
- Cordeiro, I.R.; Moreira, F.F. New distributional data on aquatic and semiaquatic bugs (Hemiptera: Heterop-tera: Gerromorpha & Nepomorpha) from South America. Biodivers. Data J. 2015, 3, e4913. [Google Scholar] [CrossRef] [Green Version]
- Cunha, E.J.; Juen, L. Environmental drivers of the metacommunity structure of insects on the surface of tropical streams of the Amazon. Austral Ecol. 2020, 45, 586–595. [Google Scholar] [CrossRef]
- Guterres, A.P.; Cunha, E.J.; Godoy, B.S.; Silva, R.R.; Juen, L. Co-occurrence patterns and morphological similarity of semiaquatic insects (Hemiptera: Gerromorpha) in streams of Eastern Amazonia. Ecol. Entomol. 2020, 45, 155–166. [Google Scholar] [CrossRef]
- Cabette, H.S.R.; Giehl, N.F.S.; Dias-Silva, K.; Juen, L.; Batista, J.B. Distribuição de Nepomorpha e Gerro-morpha (Insecta: Heteroptera) da Bacia Hidrográfica do Rio Suiá- Miçu, MT: Riqueza relacionada à qualidade da água e do hábitat. In Gestão e Educação Ambiental: Água, Biodiversidade e Cultura; Santos, J.E., Galbiati, C., Moschini, L.E., Eds.; RiMa: São Carlos, Brazil, 2010; Volume 2, pp. 113–137. [Google Scholar]
- Strahler, A.N. Quantitative analysis of watershed geomorphology. Eos. Trans. Am. Geophys. Un. 1957, 38, 913–920. [Google Scholar] [CrossRef]
- Skorupa, L.A.; Costa, U.M.P.; Cunha, P.; Rossete, A.N.; Fidalgo, E.C.C.; Simões, M.; Choueri, R. Monitoramento da Qualidade da Água na Bacia Hidrográfica do Rio Suiá-Miçu. Embrapa Meio Ambiente-Documentos; IN-FOTECA-E: Brasília, Brazil, 2017. [Google Scholar]
- Köppen, W. Grundriss der Klimakunde; Gruyter: Berlin, Germany, 1931; 338p. [Google Scholar]
- Ratter, J.A.; Askew, G.P.; Montgomery, R.F.; Gifford, D.R. Observations on the vegetation of northeastern Mato Grosso II. Forest and Soils of the Rio Suiá-Miçú area. Proc. R. Soci. Lond. 1978, 203, 191–208. [Google Scholar] [CrossRef]
- Domingues, M.S.; Bermann, C. O arco de desflorestamento na Amazônia: Da pecuária à soja. Ambient. Soc. 2012, 15, 1–22. [Google Scholar] [CrossRef]
- Ferreira-Peruquetti, P.; De Marco, P., Jr. Efeito da alteração ambiental sobre comunidades de Odonata em riachos de Mata Atlântica de Minas Gerais, Brasil. Rev. Bras. Zool. 2002, 19, 317–327. [Google Scholar] [CrossRef]
- Dias-Silva, K.; Juen, L.; Cabette, H.S.R.; De Marco, P., Jr. The influence of habitat integrity and physical-chemical water variables on the structure of aquatic and semi-aquatic Heteroptera. Zoologia 2010, 27, 918–930. [Google Scholar] [CrossRef]
- Nieser, N.; Melo, A.L. Os Heterópteros Aquáticos de Minas Gerais: Guia Introdutório com Chave de Identificação Para as Espécies de Nepomorpha e Gerromorpha; Universidade Federal de Minas Gerais: Belo Horizonte, Brazil, 1997; 180p. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association; American Water Works Association and Water Environmental Federation: Washington DC, USA, 1998. [Google Scholar]
- Nessimian, J.L.; Venticinque, E.M.; Zuanon, J.; De Marco, P., Jr.; Gordo, M.; Fidelis, L.; Batista, J.D.; Juen, L. Land use, habitat integrity, and aquatic insect assemblages in Central Amazonian streams. Hydrobiologia 2008, 614, 117–131. [Google Scholar] [CrossRef]
- Esri. ArcGIS Desktop: Version 10.1. Environmental Systems Research Institute: Redlands, CA, USA, 2014. [Google Scholar]
- Prince, S.D.; Becker-Reshef, I.; Rishmawi, K. Detection and mapping of long-term land degradation using local net production scaling: Application to Zimbabwe. Remote Sens. Environ. 2009, 113, 1046–1057. [Google Scholar] [CrossRef]
- Prates-Clark, C.D.C.; Saatchi, S.S.; Agosti, D. Predicting geographical distribution models of high-value timber trees in the Amazon Basin using remotely sensed data. Ecol. Modell. 2008, 211, 309–323. [Google Scholar] [CrossRef]
- Neigh, C.S.; Tucker, C.J.; Townshend, J.R. North American vegetation dynamics observed with multi-resolution satellite data. Remote Sens. Environ. 2008, 112, 1749–1772. [Google Scholar] [CrossRef]
- Justice, C.O.; Vermote, E.; Townshend, J.R.; Defries, R.; Roy, D.P.; Hall, D.K.; Salomonson, V.V.; Privette, J.L.; Riggs, G.; Strahler, A.; et al. The Moderate Resolution Imaging Spectroradiometer (MODIS): Land remote sensing for global change research. IEEE Trans. Geosci. Remote Sens. 1998, 36, 1228–1249. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Henry, M.; Stevens, W.H. Vegan: Community Ecology Package. R package version, 1.17-4. 2010. Available online: http://CRAN.R-project.org/package=vegan (accessed on 20 April 2020).
- Clarke, K.R.; Ainsworth, M. A method of linking multivariate community structure to environmental variables. Mar. Ecol. Prog. Ser. 1993, 92, 205–219. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Oxford, UK, 2012. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2019. Available online: https://www.R-project.org/ (accessed on 20 April 2020).
- Ilha, P.; Rosso, S.; Schiesari, L. Effects of deforestation on headwater stream fish assemblages in the Upper Xingu River Basin, Southeastern Amazonia. Neotrop. Ichthyol. 2019, 17. [Google Scholar] [CrossRef]
- De Faria, A.P.J.; Ligeiro, R.; Callisto, M.; Juen, L. Response of aquatic insect assemblages to the activities of traditional populations in eastern Amazonia. Hydrobiologia 2017, 802, 39–51. [Google Scholar] [CrossRef]
- Mendes, T.P.; Cabette, H.S.R.; Juen, L. Setting boundaries: Environmental and spatial effects on Odonata larvae distribution (Insecta). An. Acad. Bras. Cienc. 2015, 87, 239–248. [Google Scholar] [CrossRef]
- Brasil, L.S.; Batista, J.D.; Giehl, N.F.S.; Valadão, M.B.X.; Santos, J.O.D.; Dias-Silva, K. Environmental integrity and damselfly species composition in Amazonian streams at the “arc of deforestation” region, Mato Grosso, Brazil. Acta Limnol. Bras. 2014, 26, 278–287. [Google Scholar] [CrossRef]
- Juen, L.; Oliveira-Junior, J.M.B.; Shimano, Y.; Mendes, T.P.; Cabette, H.S.R. Composição e riqueza de Odonata (Insecta) em riachos com diferentes níveis de conservação em um ecótone Cerrado-Floresta Amazônica. Acta Amaz. 2014, 44, 223–233. [Google Scholar] [CrossRef]
- Voelz, N.J.; Mcarthur, J.V. An exploration of factors influencing lotic insect species richness. Biodivers. Conserv. 2000, 9, 1543–1570. [Google Scholar] [CrossRef]
- Leal, C.G.; Lennox, G.D.; Ferraz, S.F.; Ferreira, J.; Gardner, T.A.; Thomson, J.R.; Berenguer, E.; Lees, A.C.; Hughes, R.M.; Nally, R.M.; et al. Integrated terrestrial-freshwater planning doubles conservation of tropical aquatic species. Science 2020, 370, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Dias-Silva, K.; Juen, L.; Brasil, L.S.; Freitas, P.V.; De Marco, P., Jr. Influence of local variables and landscape metrics on Gerromorpha (Insecta: Heteroptera) assemblages in Savanna streams, Brazil. Neotropic. Entomol. 2020, 49, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Giehl, N.F.S.; Cabette, H.S.R.; Dias-Silva, K.; Juen, L.; Moreira, F.F.F.; Castro, L.A.; Ferreira, V.R.S.; Batista, J.D. Variation in the diversity of semiaquatic bugs (Insecta: Heteroptera: Gerromorpha) in altered and preserved veredas. Hydrobiologia 2020, 847, 3497–3510. [Google Scholar] [CrossRef]
- Liess, S.; Snyder, P.K.; Harding, K.J. The effects of boreal forest expansion on the summer Arctic frontal zone. Clim. Dyn. 2012, 38, 1805–1827. [Google Scholar] [CrossRef]
- Lock, K.; Adriaens, T.; Van De Meutter, F.; Goethals, P. Effect of water quality on waterbugs (Hemiptera: Gerromorpha & Nepomorpha) in Flanders (Belgium): Results from a large-scale field survey. Ann. Limnol. Int. J. Lim. 2013, 49, 121–128. [Google Scholar] [CrossRef]
- Bastos, R.C.; Brasil, L.S.; Oliveira-Junior, J.M.B.; Carvalho, F.G.; Lennox, G.D.; Barlow, J.; Juen, L. Morphological and phylogenetic factors structure the distribution of damselfly and dragonfly species (Odonata) along an environmental gradient in Amazonian streams. Ecol. Indic. 2021, 122, 107257. [Google Scholar] [CrossRef]
- Cummins, K.W.; Wilzbach, M.A.; Gates, D.M.; Perry, J.B.; Taliaferro, W.B. Shredders and riparian vegetation. BioScience 1989, 39, 24–30. [Google Scholar] [CrossRef]
- Giehl, N.F.; Resende, B.O.; Roges, P.F.; Dias-Silva, K.; Nogueira, D.S.; Cabette, H.S.R. Diversidade de presas e predadores (Insecta) em mesohabitats de córregos de Cerrado. Iheringia Sér. Zool. 2018, 108, e2018042. [Google Scholar] [CrossRef]
- Castro, D.M.P.; Dolédec, S.; Callisto, M. Land cover disturbance homogenizes aquatic insect functional structure in neotropical savanna streams. Ecol. Indic. 2018, 84, 573–582. [Google Scholar] [CrossRef]
- Martins, R.T.; Brito, J.; Dias-Silva, K.; Leal, C.G.; Leitão, R.P.; Oliveira, V.C.; Oliveira-Junior, J.M.B.; Ferraz, S.F.B.; De Paula, F.R.; Roque, F.O.; et al. Low forest-loss thresholds threaten Amazonian fish and macroinvertebrate assemblage integrity. Ecol. Indic. 2021, 127, 107773. [Google Scholar] [CrossRef]
- Pimentel, C.S.; Ramos, M.E.P.L.; Dias-Silva, K.; Costa, U.M.P. Avaliação de algumas variáveis limnológicas de três córregos em Nova Xavantina-MT. Enciclopédia Biosfera 2014, 10, 2023–2035. [Google Scholar]
- Brasil, L.S.; Luiza-Andrade, A.; Calvão, L.B.; Dias-Silva, K.; Faria, A.P.J.; Shimano, Y.; Oliveira-Junior, J.M.B.; Cardoso, M.N.; Juen, L. Aquatic insects and their environmental predictors: A scientometric study focused on environmental monitoring in lotic environmental. Environ. Monit. Assess. 2020, 192, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Peeters, E.T.; Gylstra, R.; Vos, J.H. Benthic macroinvertebrate community structure in relation to food and environmental variables. Hydrobiologia 2004, 519, 103–115. [Google Scholar] [CrossRef]
- Godoy, B.S.; Faria, A.P.J.; Juen, L.; Lodi, S.; Oliveira, L.G. Taxonomic sufficiency and effects of environ-mental and spatial drivers on aquatic insect community. Ecol. Indic. 2019, 107, 105624. [Google Scholar] [CrossRef]
- Goulart, M.; Melo, A.L.; Callisto, M. Qual a relação entre variáveis ambientais e diversidade de heterópteros aquáticos em nascentes de altitude? BIOS Cad. Do Dep. De Ciências Biológicas Da PUC Minas 2002, 10, 63–76. [Google Scholar]
- Leitão, R.P.; Zuanon, J.; Mouillot, D.; Leal, C.G.; Hughes, R.M.; Kaufmann, P.R.; Villéger, S.; Pompeu, P.S.; Kasper, D.; de Paula, F.R.; et al. Disentangling the pathways of land use impacts on the functional structure of fish assemblages in Amazon streams. Ecography 2018, 41, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Couceiro, S.R.M.; Hamada, N.; Luz, S.L.; Forsberg, B.R.; Pimentel, T.P. Deforestation and sewage effects on aquatic macroinvertebrates in urban streams in Manaus, Amazonas, Brazil. Hydrobiologia 2007, 575, 271–284. [Google Scholar] [CrossRef]
- Song, Y.; Song, X.; Shao, G.; Hu, T. Effects of Land Use on Stream Water Quality in the Rapidly Urbanized Areas: A Multiscale Analysis. Water 2020, 12, 1123. [Google Scholar] [CrossRef]
- Richter, B.D.; Braun, D.P.; Mendelson, M.A.; Master, L.L. Threats to imperiled freshwater fauna: Amena-zas a la fauna dulceacuicola en riesgo. Conserv. Biol. 1997, 11, 1081–1093. [Google Scholar] [CrossRef]
- Newcombe, C.P.; Jensen, J.O. Channel suspended sediment and fisheries: A synthesis for quantitative assessment of risk and impact. N. Am. J. Fish. Manag. 1996, 16, 693–727. [Google Scholar] [CrossRef]
- Calvão, L.B.; Nogueira, D.S.; Montag, L.F.A.; Lopes, M.A.; Juen, L. Are Odonata communities impacted by conventional or reduced impact logging? For. Ecol. Manag. 2016, 382, 143–150. [Google Scholar] [CrossRef]
- Herlihy, A.T.; Sifneos, J.C.; Hughes, R.M.; Peck, D.V.; Mitchell, R.M. The relation of lotic fish and benthic macroinvertebrate condition indices to environmental factors across the conterminous USA. Ecol. Indic. 2020, 112, 105958. [Google Scholar] [CrossRef] [PubMed]
- Ongaratto, R.; Loureiro, R.; Restello, R.; Hepp, L.U. Effects of land use and limnological variables on the dissimilarity of common and rare aquatic insects in Atlantic Forest streams. Int. J. Trop. Biol. 2018, 66, 1223–1231. [Google Scholar] [CrossRef] [Green Version]
- Guterres, A.P.M.; Cunha, E.J.; Juen, L. Tolerant semiaquatic bugs species (Heteroptera: Gerromorpha) are associated to pasture and conventional logging in the Eastern Amazon. J. Insect Conserv. 2021, 25, 555–567. [Google Scholar] [CrossRef]
- Vieira, T.B.; Dias-Silva, K.; Pacífico, E.S. Effects of riparian vegetation integrity on fish and heteroptera communities. Appl. Ecol. Environ. Res. 2015, 13, 53–65. [Google Scholar] [CrossRef]
- Benstead, J.P.; Douglas, M.M.; Pringle, C.M. Relationships of stream invertebrate communities to deforestation in eastern Madagascar. Ecol. Applic. 2003, 13, 1473–1490. [Google Scholar] [CrossRef]
- Casatti, L.; Teresa, F.B.; Gonçalves-Souza, T.; Bessa, E.; Manzotti, A.R.; Gonçalves, C.D.S.; Zeni, J.D.O. From forests to cattail: How does the riparian zone influence stream fish? Neotrop. Ichthyol. 2012, 10, 205–214. [Google Scholar] [CrossRef]
- Tanaka, M.O.; De Souza, A.L.T.; Moschini, L.E.; De Oliveira, A.K. Influence of watershed land use and riparian characteristics on biological indicators of stream water quality in southeastern Brazil. Agric. Ecosyst. Environ. 2016, 216, 333–339. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Infraorder/Family/Genus | Total | Forest | Agriculture |
|---|---|---|---|
| Gerromorpha | |||
| Gerridae | |||
| Brachymetra Mayr, 1865 | 10 | × | |
| Cylindrostethus Mayr, 1865 | 13 | × | × |
| Limnogonus Stål, 1868 | 1 | × | |
| Neogerris Matsumura, 1913 | 16 | × | × |
| Hydrometridae | |||
| Hydrometra Latreille, 1796 | 1 | × | |
| Veliidae | |||
| Euvelia Drake, 1957 | 9 | × | |
| Rhagovelia Mayr, 1865 | 32 | × | × |
| Stridulivelia Hungerford, 1929 | 2 | × | × |
| Nepomorpha | |||
| Belostomatidae | |||
| Belostoma Latreille, 1807 | 3 | × | × |
| Corixidae | |||
| Tenagobia Bergroth, 1899 | 7 | × | × |
| Naucoridae | |||
| Ambrysus Stål, 1862 | 13 | × | × |
| Ctenipocoris Montandon, 1897 | 1 | × | |
| Limnocoris Stål, 1860 | 121 | × | × |
| Pelocoris Stål, 1876 | 1 | × | |
| Placomerus La Rivers, 1956 | 2 | × | |
| Nepidae | |||
| Curicta Stål, 1862 | 1 | × | |
| Ranatra Fabricius, 1790 | 3 | × | |
| Notonectidae | |||
| Buenoa Kirkaldy, 1904 | 6 | × | |
| Martarega White, 1879 | 20 | × | × |
| Potomocorinae | 1 | × | |
| Total | 263 |
| Combinations of Environmental Variables | Correlation |
|---|---|
| Dep | 0.409 |
| Cond + Dep | 0.470 |
| Cond+ Dep + Pho | 0.511 |
| NDVI + Cond + Dep + Pho | 0.522 |
| HII + NDVI + Cond + Turb + Dep | 0.524 |
| HII+ EVI + Cond + Dep + Nitr + Pho | 0.522 |
| HII+ EVI + Cond + AT + Dep + Nitr + Pho | 0.515 |
| HII+ EVI + Cond + Turb + AT + Dep + Nitr + Pho | 0.511 |
| HII+ EVI + NDVI + Cond + Turb + AT + Dep + Nitr + Pho | 0.490 |
| HII+ EVI + NDVI + pH + Cond + Turb + AT + Dep + Nitr + Pho | 0.463 |
| HII + EVI + NDVI + pH + Cond + Turb + AT + WT+ Dep+ Nitr + Pho | 0.436 |
| HII + EVI + NDVI + pH + Cond + Turb + AT + WT + Prof + Wid + Nitr + Pho | 0.413 |
| HII + EVI + NDVI + pH + Cond + Turb + AT + WT + DO + Dep + Wid + Nitr + Pho | 0.379 |
| HII+ EVI + NDVI + pH + Cond +Turb + AT + WT + DO + Dep + Wid + Nitr + Pho + Mg | 0.345 |
| HII + EVI + NDVI + pH + Cond + Turb + AT + WT + DO + Dep + Wid + Nitr + Pho + TH + Mg | 0.231 |
| HII + EVI + NDVI + pH + Cond + Turb + AT + WT + DO + Dep + Wid + Nitr + Pho + TH + Ca + Mg | 0.088 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva-Costa, I.; Giehl, N.F.S.; Pozzobom, U.M.; Carvalho-Soares, A.A.; Oliveira-Junior, J.M.B.; Cabette, H.S.R.; Dias-Silva, K. Aquatic and Semiaquatic Heteroptera (Hemiptera: Insecta) Distribution in Streams on the Cerrado–Amazon Ecotone in Headwaters of Xingu River. Arthropoda 2023, 1, 13-24. https://doi.org/10.3390/arthropoda1010004
Silva-Costa I, Giehl NFS, Pozzobom UM, Carvalho-Soares AA, Oliveira-Junior JMB, Cabette HSR, Dias-Silva K. Aquatic and Semiaquatic Heteroptera (Hemiptera: Insecta) Distribution in Streams on the Cerrado–Amazon Ecotone in Headwaters of Xingu River. Arthropoda. 2023; 1(1):13-24. https://doi.org/10.3390/arthropoda1010004
Chicago/Turabian StyleSilva-Costa, Iluany, Nubia França Silva Giehl, Ully Mattilde Pozzobom, Anderson André Carvalho-Soares, José Max Barbosa Oliveira-Junior, Helena Soares Ramos Cabette, and Karina Dias-Silva. 2023. "Aquatic and Semiaquatic Heteroptera (Hemiptera: Insecta) Distribution in Streams on the Cerrado–Amazon Ecotone in Headwaters of Xingu River" Arthropoda 1, no. 1: 13-24. https://doi.org/10.3390/arthropoda1010004