1. Introduction
California currently leads the world in hybrid sunflower (
Helianthus annuus) seed production [
1]. Neonicotinoid seed treatments, typically thiamethoxam, are used on California sunflower crops to control for soil pests including variegated cutworms (
Peridroma saucia), wireworms (
Limonius spp.), and seedcorn maggot (
Delia platura), each of which can cause serious damage to seedlings and significantly affect crop growth [
1]. However, the ecological impact of neonicotinoid seed treatments and their effectiveness on crop yield are not well documented for many crops, including sunflowers [
2,
3,
4]. Furthermore, neonicotinoids are known to have deleterious effects on many beneficial insects, particularly pollinators [
5,
6], and as such, neonicotinoids could impact crop yield in pollinator dependent crops.
Wild bees and honey bees are crucial pollinators for many food crops [
7]. In fact, while honey bees are generally used to pollinate hybrid sunflowers, wild bees are known to enhance the sunflower pollination efficiency by 5-fold [
8]. However, wild bees are in decline largely due to agricultural intensification and land use change, which alter the availability of floral and nesting resources and increase exposure to pathogens and pesticides [
9,
10]. While potentially not as toxic to bees as imidacloprid [
11], thiamethoxam has received less attention than other commonly used neonicotinoids (i.e., imidacloprid and clothianidin). Despite this, thiamethoxam is known to be highly toxic to honey bees and bumble bees [
6]. A number of sublethal effects have been reported for these taxa, including evidence that thiamethoxam negatively impacts bee learning, memory, reproduction, immune response, and colony fitness, even when individuals are only exposed as larvae [
12,
13,
14,
15]. Thiamethoxam is also known to affect honey bees through synergistic interactions with parasitic infections (e.g.,
Nosema ceranae) and pathogens (e.g., deformed wing virus) [
16,
17]. The literature on the deleterious effects to honey bees and bumble bees has documented effects ranging from the impairment of flight, phototaxis, and thermoregulation [
18,
19,
20,
21] to the impairment of crop pollination services [
22]. In comparison, few studies, and much less field studies, have been conducted on the exposure and toxicity of thiamethoxam to other wild bees due to the challenges of using wild bees as a model species [
23,
24,
25] (see [
26]), although there is some evidence that wild bees may be more severely impacted than honey bees [
27].
Aside from honey bees, the sunflower specialist bee,
Melissodes agilis, is the most common sunflower visitor in the western United States [
28], although a number of other generalist and specialist species also visit sunflowers. Among these,
Anthophora urbana,
Diadasia spp.,
Melissodes spp., and
Svastra obliqua expurgata provide the highest levels of pollination efficiency in sunflower crops in California [
8]. Wild bees nest in the soil of both fields and field margins of sunflower crops and some sunflower specialists, including
M. agilis, have been found to nest only in field soils [
29]. Therefore, many wild bees may be exposed to thiamethoxam when used as a seed treatment in sunflowers if it is translocated to the pollen and nectar of the sunflowers and/or is retained in the soil where they nest.
The objectives of this study were to (1) assess the exposure of honey bees and wild bees to a thiamethoxam-treated sunflower seed along with other pesticides that may be used on a field or nearby; (2) to quantify the effects of thiamethoxam treated seed on wild bee abundance and biodiversity; and (3) to determine the effects of this neonicotinoid pesticide on the efficiency of the pollination services provided by bees in sunflower crops. We conducted a field study using a matched pairs design to test for differences in pesticide detection and pesticide concentration in soil, pollen, nectar, honey bees, and
M. agilis, and to estimate wild bee biodiversity and crop yield in hybrid sunflower fields sown with and without a thiamethoxam seed treatment. Although the focus of this study was on thiamethoxam, we also analyzed field samples for the residues of a wide range of other pesticides to gain a more complete picture of the potential exposure of bees found in sunflower crops to pesticides applied to other crops in the relatively diverse agricultural landscapes of California. While previous studies have examined the effects of imidacloprid treatment of sunflower seeds on bee pollinators [
30,
31,
32], to our knowledge, no studies have analyzed whether thiamethoxam seed treatments in sunflower remain detectable in field soils where bees may nest [
29], whether they are translocated to the pollen and nectar from which bees forage (see [
33] for similar work in a greenhouse), and whether treatment affects bee abundance and biodiversity in the field. The results from our study have the potential both to inform pest management practices with regard to their effects on wild bee conservation and to inform farmers and seed companies of the potential of thiamethoxam-treated sunflower seed to impact seed yield.
2. Materials and Methods
2.1. Study Sites and Sampling Design
Using a matched pairs design for a field study conducted near Sacramento, CA, USA, we collected samples to investigate pesticide exposure for bees foraging in sunflowers (H. annuus) sown with and without a neonicotinoid seed treatment. One side of each field was sown with thiamethoxam-treated seed (referred to as thiamethoxam treatment) and the other side sown with seed that was not treated with thiamethoxam (referred to as control treatment). During peak bloom (late June to early July 2016), M. agilis, pollen foraging honey bees, nectar foraging honey bees, sunflower pollen, sunflower nectar, and soil samples were collected from both sides of four fields.
Sunflower crops are grown such that pollen-producing parental lines (hereafter, “P-type”) and non-pollen-producing parental lines from which the seed is harvested (hereafter, “S-type”) are sown in separate rows. Although P-type and S-type sunflower plants have historically been referred to as “male” and “female” plants, respectively [
1], here we use gender neutral terms to describe parental lineages [
34]. Bees foraging for pollen and nectar must move pollen from P-type flowers to S-type flowers for pollination to occur. After pollination, the P-type plants are mowed down and the seed is harvested from the S-type plants. Honey bee hives are commonly rented from bee keepers and set out on field edges as P-type plants begin to flower and before S-type plants bloom. Honey bees tend to have distinct subsets of workers dedicated to pollen- versus nectar-foraging. Since P-type (pollen and nectar) and S-type (nectar only) plants do not co-occur in the same rows in hybrid sunflower monoculture, this means that individual honey bees tend to forage on only one type of plant during a foraging trip, which is not conducive to pollination. Wild bees forage for both pollen and nectar and their presence in the system indirectly increases the pollination of sunflowers because wild bees impact honey bee behavior, causing them to move back and forth between P-type and S-type rows, thereby doubling the pollination services of honey bees [
8].
Multiple companies grow sunflower seeds in central California, and we partnered with one small seed company for this study. The seed company received seeds from the European Union (EU) region, which had already been coated with two fungicides: Apron XL 350ES (Syngenta; 0.3 L metalaxyl-M/100 kg seed) and Maxim 025 FS (Syngenta; 0.5 L fludioxonil/100 kg seed) (note that metalaxyl-M and mefenoxam are synonymous terms.) Due to the 2013 ban on the use of certain neonicotinoids in the EU, some of the seed was additionally treated in California with the neonicotinoid Cruiser 5FS (Syngenta; 0.51 L thiamethoxam/100 kg seed). This made it possible for the company to withhold the neonicotinoid seed treatment of some seeds for the purpose of this study. In an effort to minimize crop damage from the lack of insecticide treatment for one half of each field, the seed company chose four field sites that were anecdotally known to have less activity from soil-dwelling insect pests. Each field (4 fields; 2 treatments per field for a total of n = 8 treatments) was sown such that all seeds were coated with the two fungicides (n = 8), but only half the field had seeds coated with thiamethoxam (n = 4). Each field was sown in mid-April 2016 with a different cultivar (the cultivars were reall, RGT cllif, Mooglli CL, and RGT absolute); and the cultivars were identical between matched pairs. Three of the fields ranged from 20 to 23 ha in size, while the fourth field was smaller at 11 ha. The P-type seed density was 49,420–56,830 seeds/ha and the S-type seed density was 46,950–59,310 seeds/ha. Sunflower seeds were sown such that there were 6–10 rows of S-type, interrupted by two rows of P-type, as is typical for hybrid seed production. Fields were not treated with any other pesticides after planting and crops grown in these fields during the previous two years were a mix of wheat, tomatoes, safflower, and squash.
2.2. Sampling for Pesticide Residue Analysis
For each field, pollen, nectar, and bee samples were collected along two transects in each treatment, located at least 20 rows (~18 m) away from the center (i.e., where the two paired seed treatments met). The two transects were established between S-type and P-type rows of sunflower plants and were at least three sets of P-type rows apart. Sampling began at least 15 m from all field edges. Soil samples were collected at five random locations within each treatment at each field site. Nectar foraging honey bees were collected with bug vacuums (Backyard Safari, Alex Brands, Fairfield, NJ, USA) and placed on ice in coolers in the field prior to nectar harvesting procedures, but then stored at −20 °C until pesticide analysis. All other samples were placed in dry ice coolers in the field and then stored at −20 °C until pesticide analysis.
2.2.1. Soil
Pesticides can be present in soils for a variety of reasons including persistence from previous applications, aerial drift, leaching from other applications, and residue from seed treatments. Many wild and native bees nest in soil; sunflower specialists are thought to sometimes nest in soils growing the crops on which they forage [
30]. To assess pesticide exposure to crop plants and nesting bees, a composite soil sample (5 soil cores; avoiding sunflower plants and roots) to a depth of 7 cm was collected using a 2.5 cm diameter soil probe in each treatment at each field site and stored in Ziplock bags. Soil samples were collected after one-month post seeding (to check for the detection of seed treatments in the soil) and again during bloom (to coincide with the timing of bee and floral samples, 2.5 months post seeding).
2.2.2. Pollen
Pesticides can be present in pollen due to either systemic uptake or spray residue. To assess pesticide exposure to bees due to pollen, samples were directly collected from blooming P-type sunflowers. Anther dehiscence in sunflowers occurs early in the morning, just as the flowers become exposed to direct sunlight, and is immediately followed by bees foraging for pollen. Thus, pollen samples were immediately collected upon dehiscence, which is apparent from the anthers turning from a dull yellow brown color to a bright yellow as the pollen is exposed. Pollen was scraped from the anthers into glass jars using small plastic rulers and plastic funnels. An pollen sample of approximately 50 mg was collected from each transect and then pooled for a single 100 mg pollen sample from each treatment at each field site. Small arthropods and other debris were removed before pesticide analysis. Pollen and nectar sampling was performed the same morning that bee pesticide residue collection began (see
Section 2.2.4).
2.2.3. Nectar
Pesticides can also be present in nectar due to either systemic uptake or spray residues. To assess pesticide exposure to bees via nectar, samples were collected from nectar-foraging honey bees. Honey bees observed foraging from blooming S-type sunflowers for >30 s were collected with bug vacuums (Backyard Safari, Alex Brands, Fairfield, NJ, USA), transferred individually to microcentrifuge tubes, and placed on ice in a cooler in the field to immobilize them. At least 6 nectar foraging honey bees were collected from each transect. Within 24 h, the honey bees were removed from the cooler and squeezed laterally between the thumb and the forefinger to regurgitate their nectar, which was collected into 20 μL capillary tubes [
35]. Capillary tubes were then sealed, wrapped in bubble wrap, and placed in a freezer at −20 °C. At least 50 μL nectar was collected from each treatment for each field site. The actual volume of nectar collected varied between field sites and treatments but was harvested from the same number of bees for both paired treatments at each field site (12 bees from each treatment at two field sites and 14 bees from each treatment at the other two field sites); the exact volume collected from each field site and treatment was measured prior to pesticide residue analysis.
2.2.4. Bees
There are many routes by which bees may be exposed to pesticides, including the pollen and nectar of crop plants. Three types of bee samples were collected to assess differential exposure to pesticides: M. agilis (an abundant sunflower specialist), pollen-foraging honey bees, and nectar-foraging honey bees. As M. agilis cannot be fully identified by eye in the field, 20 similar wild bees observed to be foraging on P-type flowers were collected and later identified, after which 10 M. agilis were randomly selected and pooled as a composite sample for pesticide residue analysis for each treatment from each field site. Similarly, at least 12 honey bees observed to be foraging on P-type flowers, with pollen loads in their corbiculae as evidence of pollen foraging, were collected, as were 12–14 nectar-foraging honey bees collected for the regurgitation of nectar. For both foraging groups, 10 of the 12–14 individuals collected were randomly selected and pooled for each treatment from each field site. Small brushes and a solidified mixture of 2:6:7 gelatin–glycerin–water were used to remove most of the pollen from all bees in an effort to analyze the pesticide load of the bee and not the pollen it was carrying. Nectar-foraging honey bees, despite not having pollen loads, were subjected to the same pollen removal protocol to control for the potential effects of the cleaning process. The bees for pesticide residue analysis were collected over multiple days (up to 3 days for an individual field), and all the sampling was performed over a 5-day window (29 June–3 July).
2.3. Sampling for Wild Bee Biodiversity
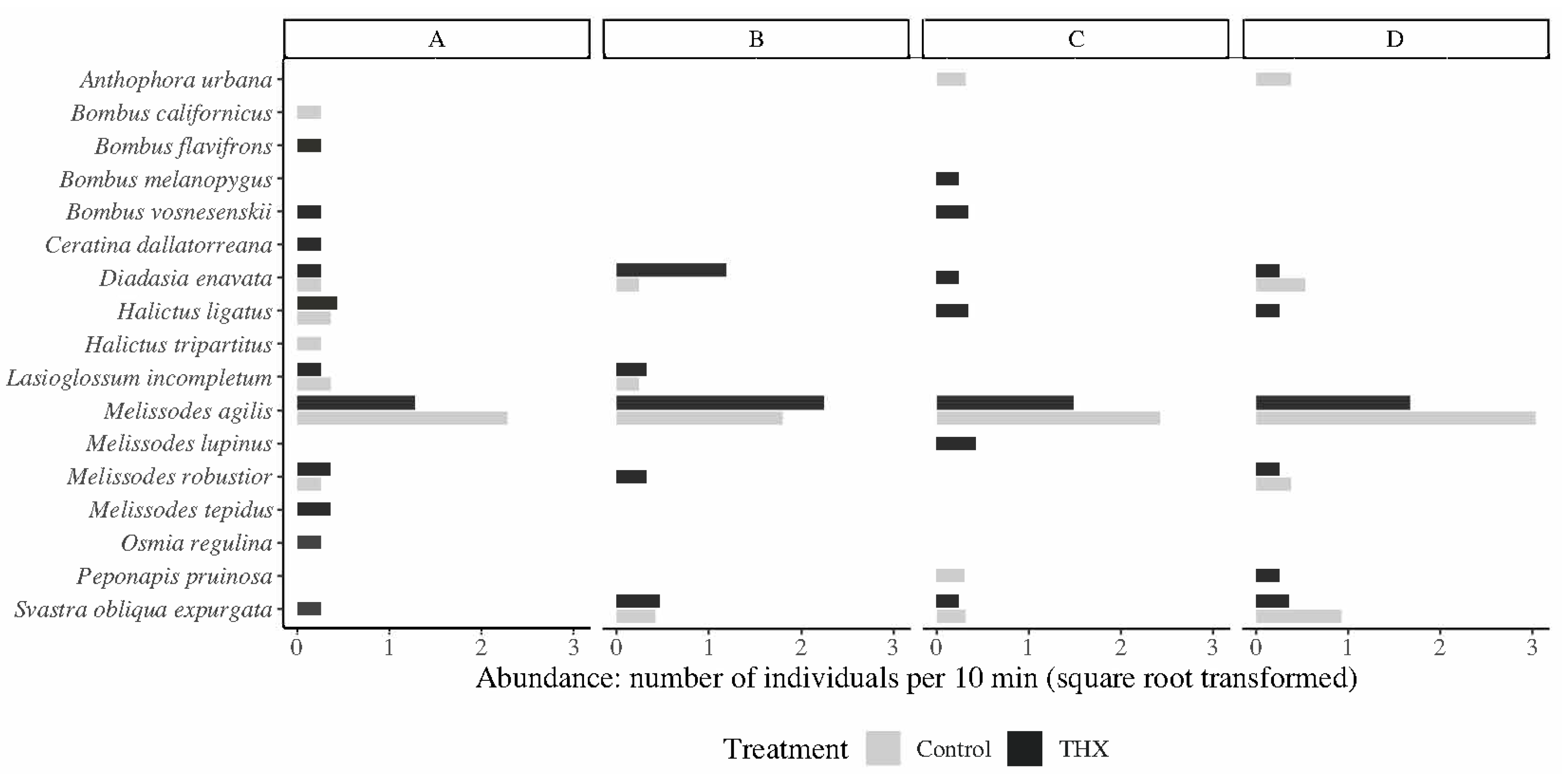
Wild bee sampling was conducted during the bloom season between 07:00 and 17:00 h on days when weather conditions were favorable for bee flight and foraging activity (little to no cloud cover, temperatures > 15.5 °C, and average wind speeds < 2.5 m/s). Wild bee sampling was performed over one day (morning and afternoon) for each site, over a 4-day window (4–7 July) for all sites. Random walks were used to collect wild bees foraging on sunflowers; bees were collected with sweep nets and bug vacuums, with the goal of sampling at least 40 individuals from each treatment for each field site. The average time for abundance sampling was 132 min (range of 69–168 min). Both sides of each field (control and treatment) were sampled simultaneously, with one person on each side of the field. Wild bee abundance was expressed as the number of wild bees collected per 10 min. Since we expected
M. agilis to be a dominant species, we also calculated the abundance of
M. agilis separately as the number of individuals collected per 10 min, as standardized by the total time spent collecting bees within each treatment at each field site. Bees were transferred into microcentrifuge tubes, placed on dry ice in the field, and later identified in the laboratory. Once identified, wild bee biodiversity was estimated as asymptotic species richness using the Chao1 estimator and as species diversity using the exponential Shannon entropy estimator from the SpadeR package in R version 3.6.1 [
36,
37]. Due to the expected dominance of the wild bee assemblage by one sunflower specialist (
M. agilis), species richness was also estimated without this species.
2.4. Sampling for Floral Resources and Crop Yield
To determine whether thiamethoxam seed treatment affected the floral resources present at our field sites, flower production and growth were estimated from five random S-type and five random P-type plants located at least 15 m into each transect. S-type plants typically have just a single flower head that blooms, whereas P-type plants can have multiple flower heads that bloom. Consequently, the diameter of the entire flower head and the distance from the external edge of the flower head to the inner most open floret were measured for the single head on S-type plants and for the largest head on P-type plants. These measurements were used to estimate the flower head area, flower head bloom area, and bloom area as a proportion of the area blooming to the full head area. We also counted the number of flower heads present on each P-type plant, the number of flower heads blooming, and then estimated the proportion of heads blooming. Floral resource sampling was performed the same morning that bee pesticide residue collection began (see
Section 2.2.4).
To examine whether treatment influenced the crop yield, yield measurements were estimated from the photographs of the flower heads at harvest. Just before seeds were harvested by the seed company, 60 photographs per treatment per field site were taken haphazardly from selected S-type heads, with a ruler placed against the flower head for scale. For each photograph, the “ruler tool” from Adobe Photoshop was used to measure 1 cm from the ruler and allow a grid of 2.25 cm² quadrats to be overlaid on the image of the flower head. Quadrats were numerically labeled in sequence and a random number generator was used to select six that fully overlapped with the sunflower head for counts of the number of viable seeds (≥0.25 cm in width) present. For quadrats where no seeds were visible (i.e., either absent or undeveloped), a seed count of 0.0001 was recorded and a mean viable seed count per 2.25 cm² quadrat was calculated for each flower head. The total area of each flower head was measured using ImageJ [
38] after adjusting for image scale and these values were used to estimate the total seed count for each flower head.
2.5. Analytical Methods for Pesticide Detection
Pesticides were extracted from soil, pollen, and bees using pressurized solvent extraction with dispersive solid-phase extraction (SPE) clean-up [
39,
40,
41]. Nectar samples were extracted using a modified QuEChERS method of acetonitrile, hexane, water, and citrate salts; with dispersive SPE clean-up the same as for the other sample types examined [
42]. Samples were analyzed for 168 pesticides and their degradates using both gas and liquid chromatography–tandem mass spectrometry. The complete list of compounds and analytical limits of detection (LOD) for each can be found elsewhere [
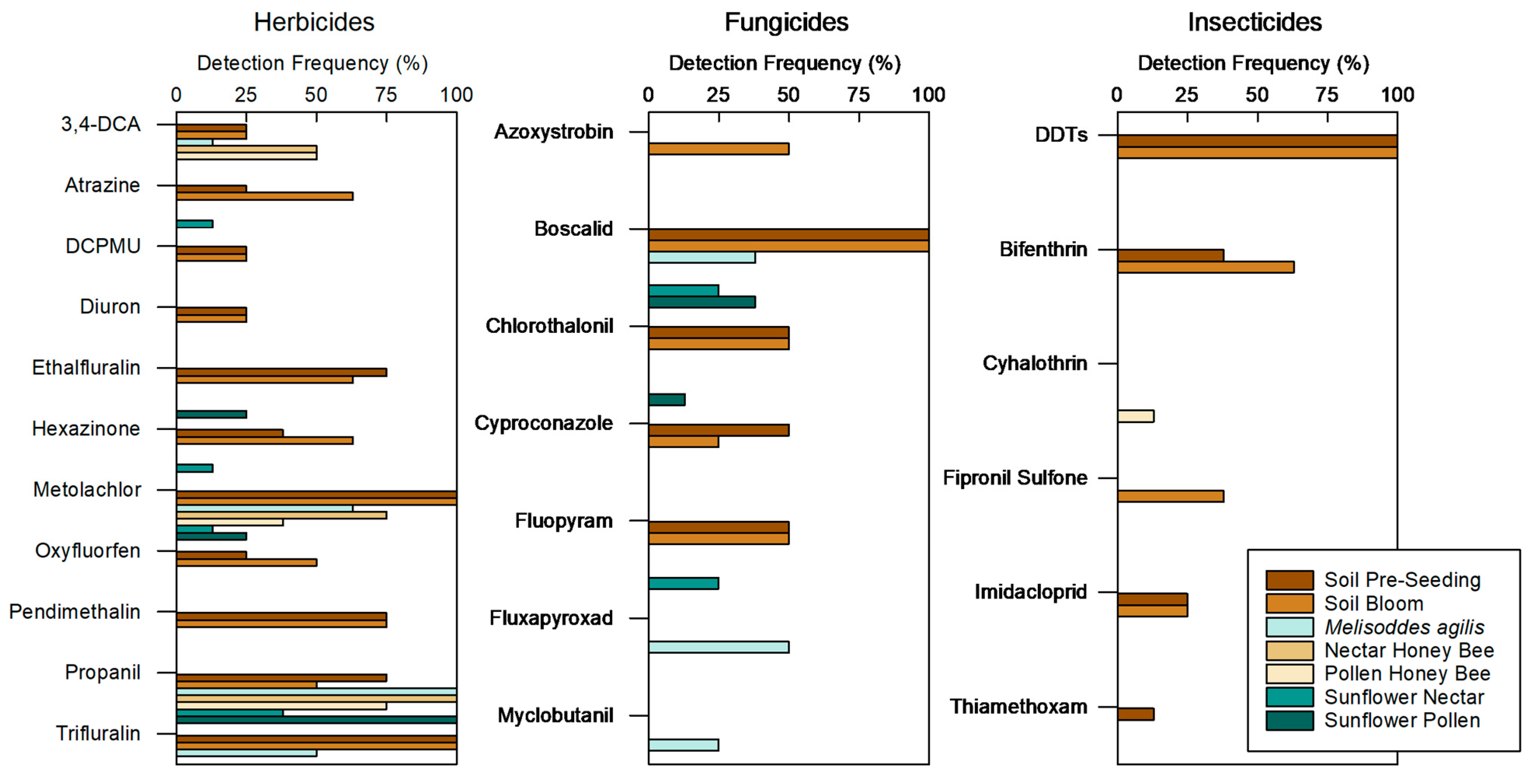
43]. Thiamethoxam, clothianidin (thiamethoxam’s primary metabolite) and the two fungicides (fludioxonil and mefenoxam/metalaxyl) applied by the seed company were among the analyzed pesticides.
2.6. Statistical Analysis
We used linear mixed models to test the effect of sunflower seed treatment (thiamethoxam versus control) on the pesticide number (i.e., the number of pesticides), pesticide concentration (ng/g), floral resource availability (proportion of head area in bloom and proportion of heads blooming), crop yield (seeds per quadrat, seed head area, and seeds per flower head), and bee community composition (richness, diversity, and abundance). Models were constructed using the lme4 package in R version 3.6.1 [
44,
45]. For the pesticide analyses (pesticide number and concentration), the sample type and pesticide category (insecticide, herbicide, fungicide) were included in the models alongside seed treatment as fixed effects. In models in which just one pesticide was analyzed, the pesticide category was removed from the model. In addition, for all pesticide analyses, soil samples from one-month post planting and during bloom were pooled in the overall analysis if they were not found to be significantly different when analyzed independently. For continuous response variables, a Gaussian error distribution was used and, in some cases, were log transformed to ensure linearity. For response variables that were counts (e.g., number of pesticides, species richness, seed counts), we used a Poisson error distribution. The field site was included as a random factor in all models to account for the matched pairs design. Type II Wald chi-square tests were used to determine the significance of fixed effects in the models using the ANOVA function from the car package in R [
46]. The post hoc comparisons of marginal means among sample types and pesticide categories (within significant fixed effects) were made using the emmeans function from the emmeans package in R [
47].
4. Discussion
We demonstrate that honey bee and wild bee pesticide exposure in agroecosystems may operate at the landscape level and that neonicotinoid seed treatment can drive ecological impacts even when the neonicotinoid treatment is nearly undetected in environmental samples. Neonicotinoid seed treatments can be effective against soil dwelling pests in sunflower fields in California [
1], but these insecticides are known to have sublethal effects on honey bees and wild bees [
5,
6]. As sunflowers rely on honey bees and wild bees for efficient pollination, the use of neonicotinoid seed treatment on sunflower is a potential concern for bee health [
1,
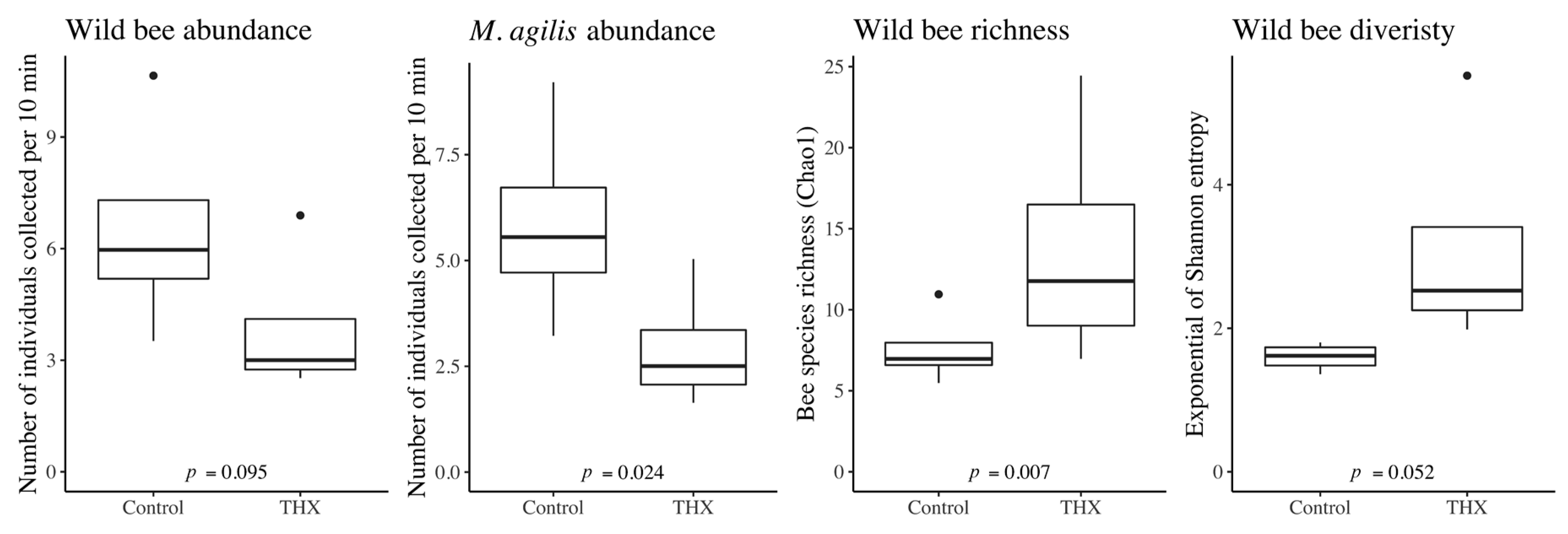
8]. This study was designed to better assess pesticide exposure for bees that forage in thiamethoxam seed-treated sunflowers in comparison to sunflowers sown without thiamethoxam treatments, and to test the effect of thiamethoxam seed treatment on bee abundance and biodiversity and crop yield. Although a number of pesticides were detected in the various samples collected as part of this study, the pesticides used for seed treatment were largely undetected, with thiamethoxam detected in only one soil sample from a control treatment (and its primary metabolite, clothianidin, never detected). Despite the absence of thiamethoxam residues from our samples, the results from our study suggest that the thiamethoxam seed treatment reduced the overall wild bee abundance (though not significantly) for all wild bees (but also significantly reduced the
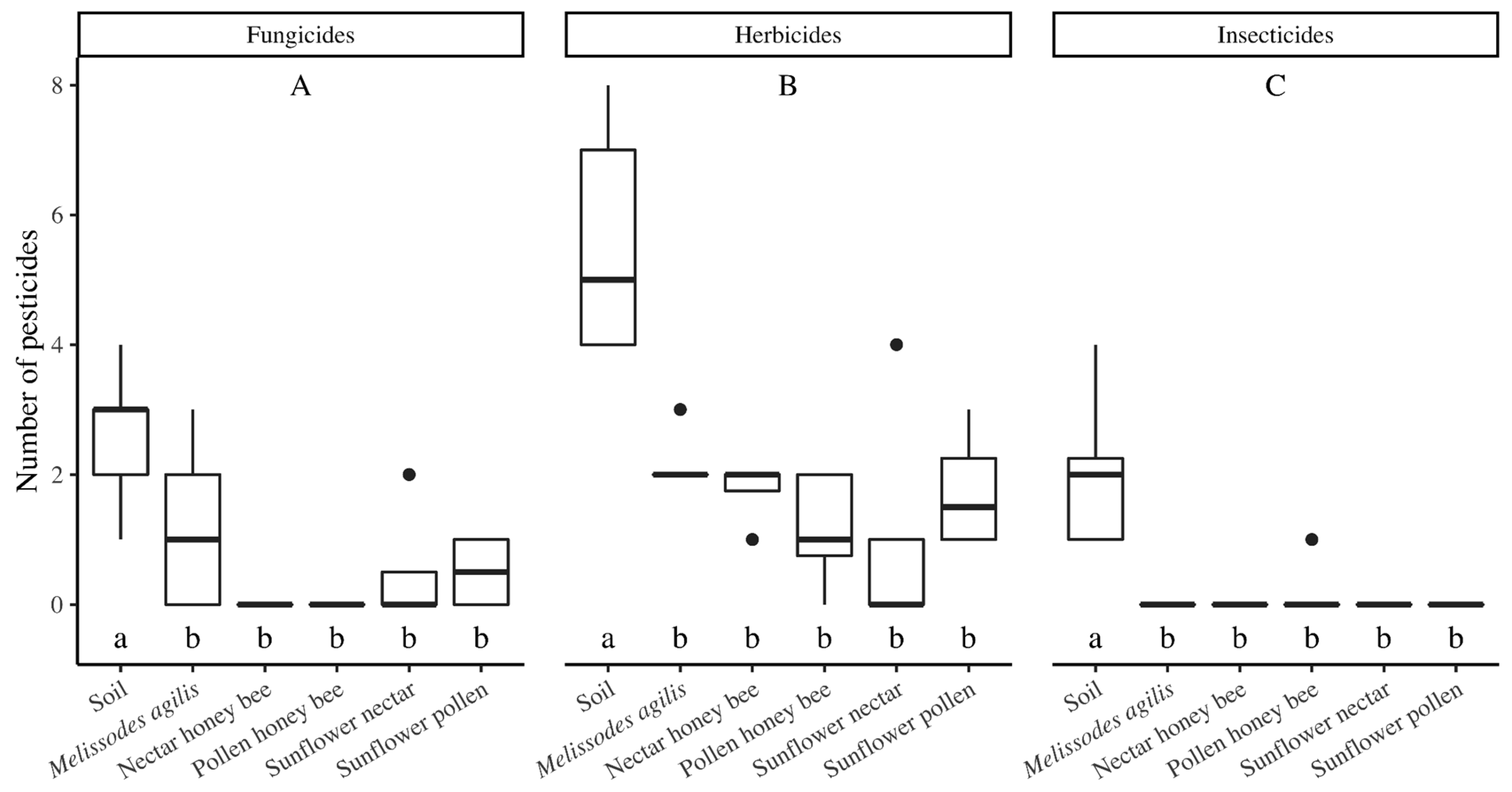
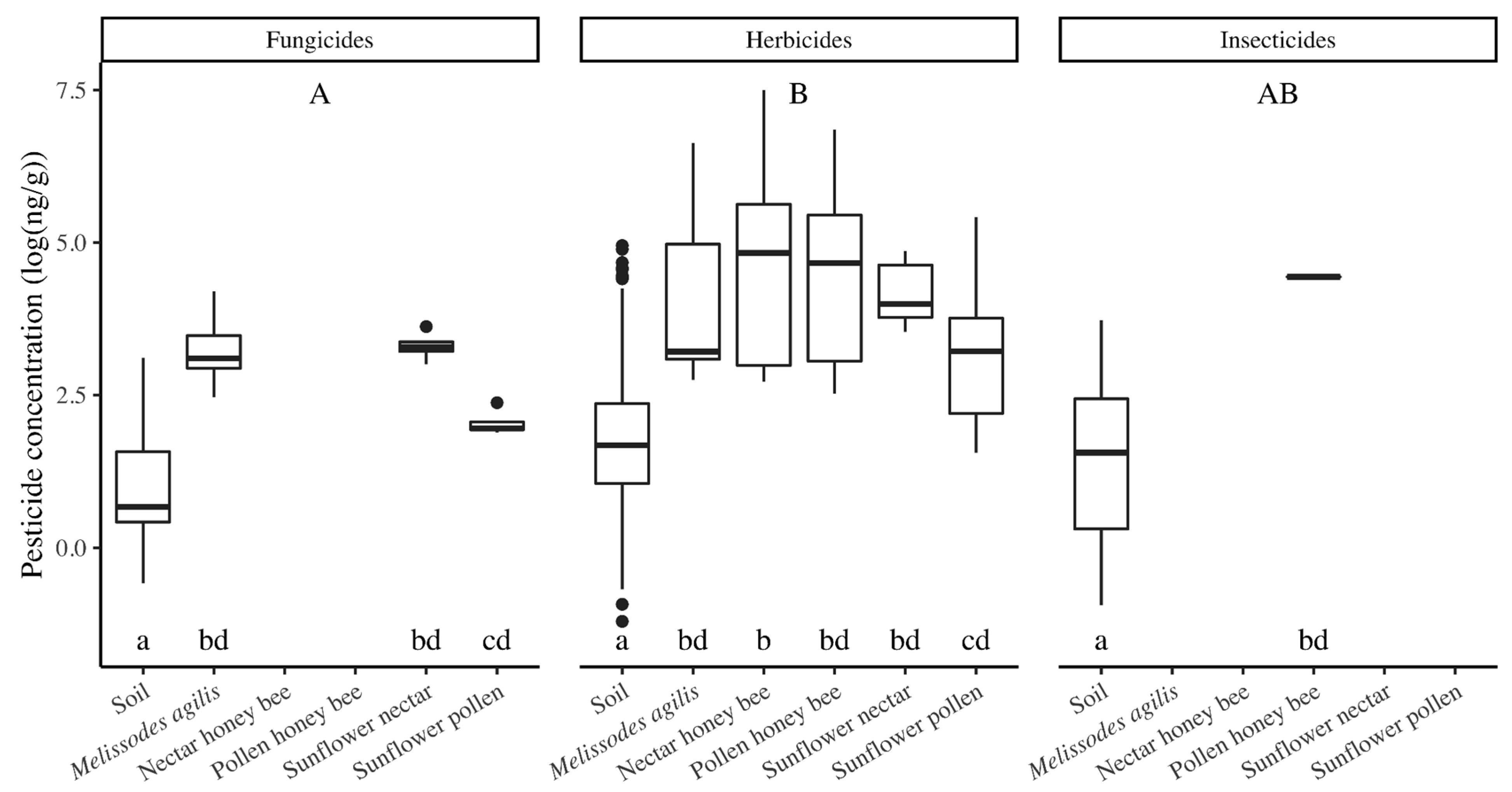
M. agilis abundance) and decreased the seed yield, but increased wild bee species richness and diversity. Although the seed-treatment pesticides were rarely detected, a number of other pesticides were detected, with no difference in the number of pesticides detected nor the concentration of pesticides detected between treatments. The greatest number of pesticides were detected in soils, though pesticide concentration was lowest in soils. Pesticide concentrations were similar among bees and nectar, with a marginal trend showing a higher pesticide concentration in bees than in pollen, indicating that bees were either exposed to pesticides by visiting other plants and/or that they were accumulating pesticides from the exposure in the sunflower fields.
Three main themes arise from these results. First, thiamethoxam seed treatment in sunflowers appeared to affect the bee abundance and biodiversity despite remaining nearly undetectable in our field samples. Second, thiamethoxam seed treatment in sunflowers decreased the seed yield, perhaps due to the decreased wild bee abundance, particularly for the sunflower specialist M. agilis. Third, pesticide exposure for bees foraging in sunflower fields extended beyond pesticides applied within the crop itself.
4.1. Effect of Thiamethoxam Seed Treatment on Bee Abundance and Biodiversity
Although thiamethoxam was only detected in one soil sample in our study, we still observed the differences in bee abundance and biodiversity between thiamethoxam and control treatments. Wild bee abundance was greater in the control treatment, which was primarily driven by increased
M. agilis abundance. Greater plant visitation rates for wild bees in control sunflower fields compared to thiamethoxam treated fields have been observed previously [
33]. The greater wild bee abundance found in the control treatment of our study could have been due to a direct deleterious effect of the thiamethoxam seed treatment itself. However, as residues were mostly undetected in our samples, these results suggest that the thiamethoxam treatment may have negatively impacted bee abundance at levels below the detection limit or indirectly through the availability and/or quality of floral resources.
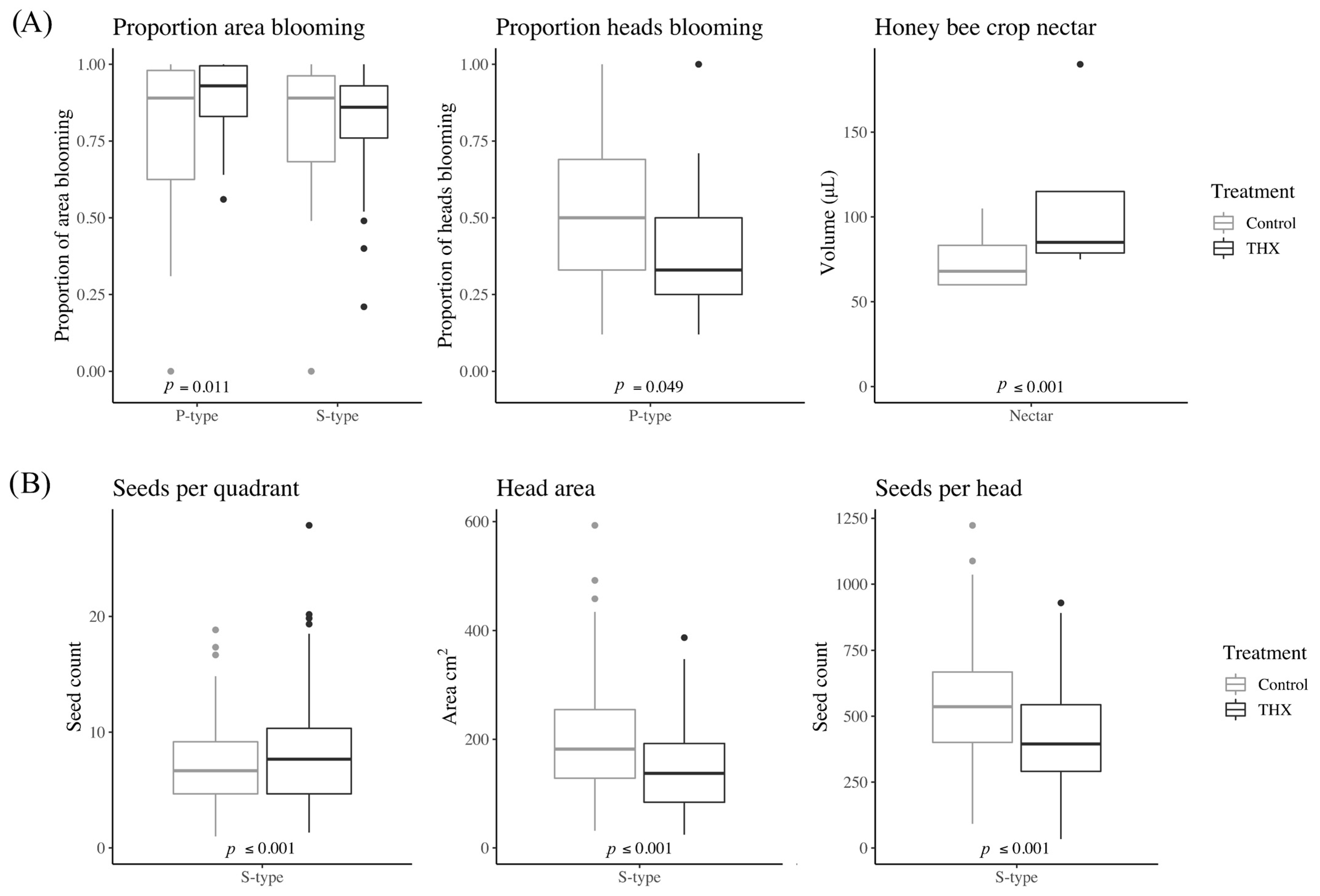
The abundance of flowers and/or pollen can impact bee abundance and species richness [
48]. In our study, the greater proportion of P-type heads blooming in the control versus thiamethoxam treatment may have positively influenced the abundance of
M. Agilis. Interestingly, the proportion of bloom area for P-type heads (i.e., the size of the sunflower landing pads while in bloom) was greater in the thiamethoxam than control treatment. This indicates that the proportion of sunflower heads in bloom is a more important factor influencing wild bee abundance in sunflower fields than the proportion of flower heads in bloom. Similarly, a greater nectar quality and quantity can positively impact bee abundance [
49]. However, the volume of nectar regurgitated by honey bees foraging from S-type plants was significantly greater in the thiamethoxam treatment, which had a lower wild bee abundance. Honey bees have been shown to consume more sucrose solution when it is laced with thiamethoxam [
50], suggesting an effect of this pesticide on foraging behavior. As we did not detect thiamethoxam in the nectar extracted from honey bees in our study, there is a possibility that the increased nectar consumption in the thiamethoxam treatment resulted from reduced competition with wild bees or from greater nectar production per flower head. It is also possible that the thiamethoxam seed treatment changed the chemistry or microbiology of the sunflower plants in ways that might cause honey bees to consume more nectar during foraging trips. For example, microbial activity is known to affect the quality and quantity of both nectar and pollen, and alter the floral scent as well as the floral rewards for pollinators [
51,
52]. Future studies could investigate the potential effects of neonicotinoid insecticides on floral microbes as well as whether and how these effects impact floral attraction and rewards for pollinators.
The greater richness and diversity of wild bee species in the thiamethoxam treatment may have been driven by the greater proportion of bloom area in the thiamethoxam treatment for P-type flowers; however, it may also have been driven by interspecific competition. Resource partitioning through interspecific competition is known to shape bee assemblage composition [
53]. Given the proximity of the paired treatments, the dominance of
M. agilis in the control treatments may have driven other bee species to forage in the thiamethoxam treatments. Similarly, this may indicate a preference for nesting in soils without a thiamethoxam treatment. Consequently, the greater species richness of less abundant wild bee species in the thiamethoxam treatments may have been because of the dominance of
M. agilis in the control treatments.
4.2. Effect of Thiamethoxam Seed Treatment on Seed Yield per Flower Head
Although previous research suggested that thiamethoxam seed treatment had no effect on sunflower crop yield [
2], the sunflower head area at harvest and the estimated seed yield per flower head were both greater in the control treatment in our study. On average, while there was only approximately one less seed per quadrat in the control treatment, because the head area was 60 cm
2 greater in the control treatment, we found over 110 more seeds per sunflower head in the control treatment. The greater seed yield may have been mediated by the greater abundance of
M. agilis in these plots. While honey bees are the primary pollinators of sunflowers in California, interactions between wild bees and honey bees have been shown to effectively double the pollination services provided by honey bees in sunflower fields [
8]. When present alone, honey bees tend to forage for either nectar or pollen during a foraging trip and thus they have no reason to move pollen from the pollen producing (P-type) plants to the seed producing (S-type) plants. However, in the presence of wild bees, honey bees can more readily switch to moving between plant types [
8]. Thus, the greater seed yields per flower head observed in the control treatment could have resulted at least partially from the effect of the greater wild bee abundance on the efficiency of the pollination services provided by honey bees. It remains unknown whether honey bees were also more abundant in the control treatment, as this was not quantified as part of our study. Future studies should also consider assessing the abundance of wild bees in relation to honey bees. However, other studies found no difference in plant visitation rates by honey bees in sunflower fields with and without thiamethoxam seed treatments [
33]. The lower seed count per quadrat in control treatment could also indicate that pollination was not maximized; that is, despite the abundance of
M. agilis in the control treatment, other factors could have caused the greater overall seed count per head. Future studies could include more extensive pollination and plant physiology measurements for a more holistic assessment of the factors that mediate maximal pollination and crop yield in sunflowers. As thiamethoxam seed treatments are an additional cost for sunflower production without showing any discernable benefit to seed yield, the greater overall seed count per head suggests that there may be some economic advantages for farmers and seed companies that abstain from the prophylactic use of thiamethoxam seed treatment in sunflowers in areas with lower soil-dwelling insect pest activity.
4.3. Pesticide Exposure for Bees Extends beyond Materials Applied within the Sunflower Crop
While the motivation for this study was focused on use of thiamethoxam seed treatment in sunflowers, a number of pesticides not used in this crop were detected in our samples, reinforcing the complexity of pesticide movement and bee exposure throughout the landscape. Although this agricultural region of California primarily consists of conventionally managed row crops and orchards, the crops are relatively diverse compared to agricultural regions in other parts of the country. We detected the greatest number of pesticides in soil samples, although the pesticide concentrations were the lowest in these samples, a finding that is consistent with those of other studies [
54]. In addition, pesticide concentrations were consistently higher in all bee groups (
M. agilis, pollen-foraging honey bees, and nectar-foraging honey bees) than in sunflower pollen, while the concentrations in pollen and nectar did not differ. In contrast, previous research showed that pesticides were detected in bees at lower concentrations than those found in pollen and nectar due to bees’ metabolism [
54,
55], and many studies have found higher concentrations of pesticides in pollen than in nectar [
56]. While the detection of residues in soil and plant samples likely stem from pesticide drift, those in bees might be indicative of their foraging in other parts of the landscape. Bees are known to forage from both crop and non-crop plants in agricultural landscapes [
57,
58], and bees foraging from such plants may acquire multiple pesticides which were not applied to the crop being studied [
54,
59,
60,
61].
The identity of the other pesticides detected in our study and the resulting potential mixtures of compounds raises concerns for bee health. Myclobutanil, a fungicide known to increase the toxicity of bifenthrin and imidacloprid in bumble bees [
62] and to decrease metabolism in honey bees [
63], was found in
M. agilis samples (and only
M. agilis samples). While bifenthrin and imidacloprid were not detected in bee samples, they were detected in soil samples, suggesting the potential for
M. agilis to be exposed to these insecticides as adults and/or as larvae while nesting in the soil. Chlorothalonil was detected in sunflower pollen and soil samples. This fungicide is known to result in fewer workers and smaller queens in bumble bees [
64] and to predict the occurrence of the bumble bee pathogen (
Nosema bombi). Bumble bees do visit sunflowers and were among the species observed at our field sites, and as such, may be transporting pollen with chlorothalonil back to their nests. Collectively, these pesticide detections show how wild bees foraging and nesting in crops are exposed to pesticides not applied to the crop but rather applied in the greater landscape.
The detection of pesticide residues also varied among bee species. Although the mean number and mean concentration of pesticides did not differ among the three categories of bees in our study, pesticides detected in
M. agilis were not always detected in honey bees and vice versa. For example, cyhalothrin was the only insecticide detected in bees and it was detected in only one pollen-foraging honey bee sample. The detection level of cyhalothrin in our bee sample was 1.9 ng/bee, and as the lethal dose to kill 50% of honey bees after 48 h of contact with cyhalothrin (LD50) is 6.1 ng/bee [
65], this represents a 0.31 toxicity unit (TU: concentration/LD50). Some research indicates that TU > 0.20 may contribute to acute toxicity and that is without accounting for potential sublethal effects, chronic effects and/or synergism between compounds that increase toxicity [
66]. All other pesticides detected in bee tissues had TU < 0.08, and were not considered for further TU analysis. In contrast to the cyhalothrin example, some pesticides were detected in wild bees but not honey bees. For example, fungicides were detected in
M. agilis but not honey bees. Specifically, the fungicide, boscalid, which is known to synergistically increase the toxicity of thiamethoxam [
67], was detected in
M. agilis and in the nectar collected by honey bees but not in the honey bees themselves. These exposure profiles exhibit a differential exposure and/or metabolism among bee species, indicating that honey bees are not a good model for the prediction of pesticide risk among wild bee species.
5. Conclusions
Although this study was limited to one year, the findings suggest that thiamethoxam seed treatment negatively impacted the M. agilis abundance and the number of seeds per flower head in hybrid sunflowers despite this compound rarely being detected in soil, and not detected in nectar, pollen, or bees. Our findings also reveal that targeted pesticide residue analysis does not always capture the full impact of field realistic pesticide exposure. The inclusion of other metrics in pesticide risk analysis for wild bees, such as abundance and diversity, broadens the objective assessment of risk. In addition, our results showed that bees were exposed to mixtures of many other pesticides that were not applied in the sunflower crop. The detection of these mixtures suggests that pesticide exposure for bees needs to be considered at the landscape level, as bees foraging within sunflower crops were exposed to pesticides not applied to those fields. Based on the increase in wild bee abundance and sunflower seed yield in plots without thiamethoxam seed treatment, we suggested that sunflower seed companies and farmers further study benefits from such treatment when the activity of soil-dwelling insect pests is low. Overall, our results indicate that pesticide exposure for honey bees and wild bees in agroecosystems can be detrimental to ecosystem services, including crop production, and furthermore, that adequate detection of pesticides requires the inclusion of multiple metrics at both the field and landscape scale.



 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





