

Fungicide Scent Pollution Disrupts Floral Search-and-Selection in the Bumblebee Bombus impatiens


Abstract
:1. Introduction
1.1. Bumblebees Serve as Vital Insect Pollinators in Our Ecosystem
1.2. Agrochemicals Are a Contributing Stressor in Pollinator Declines
1.3. Agrochemicals Indirectly Disrupt Bumblebee Learning and Foraging Behavior
2. Materials and Methods
2.1. Animals
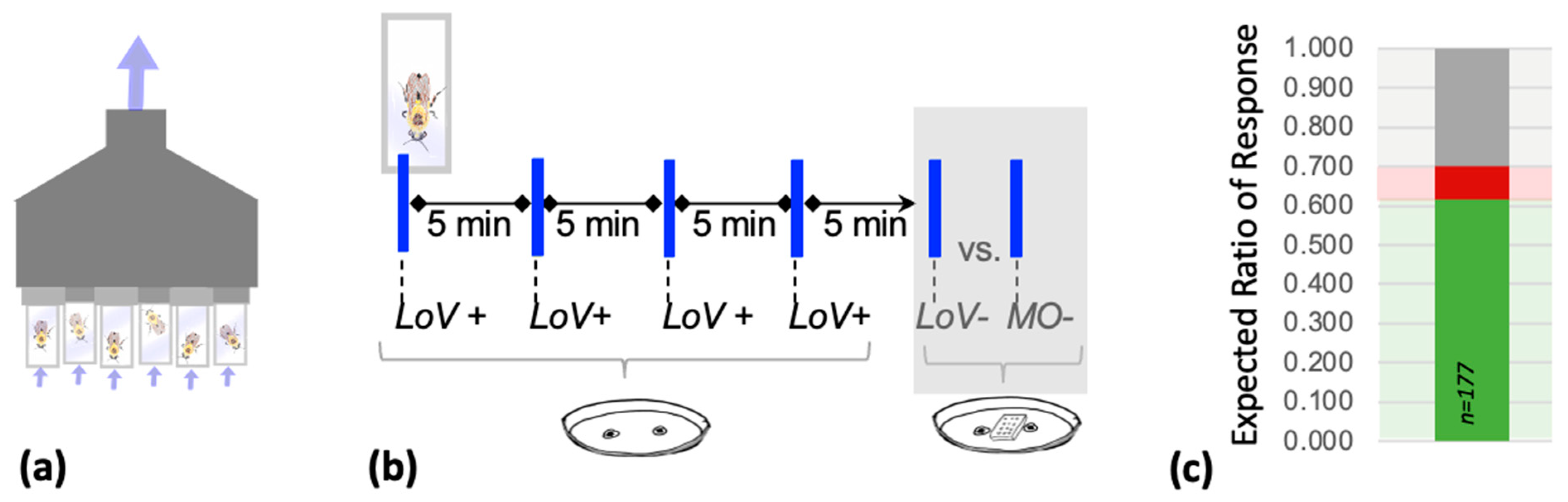
2.2. Free-Moving Proboscis Extension Reflex (FMPER) Protocol
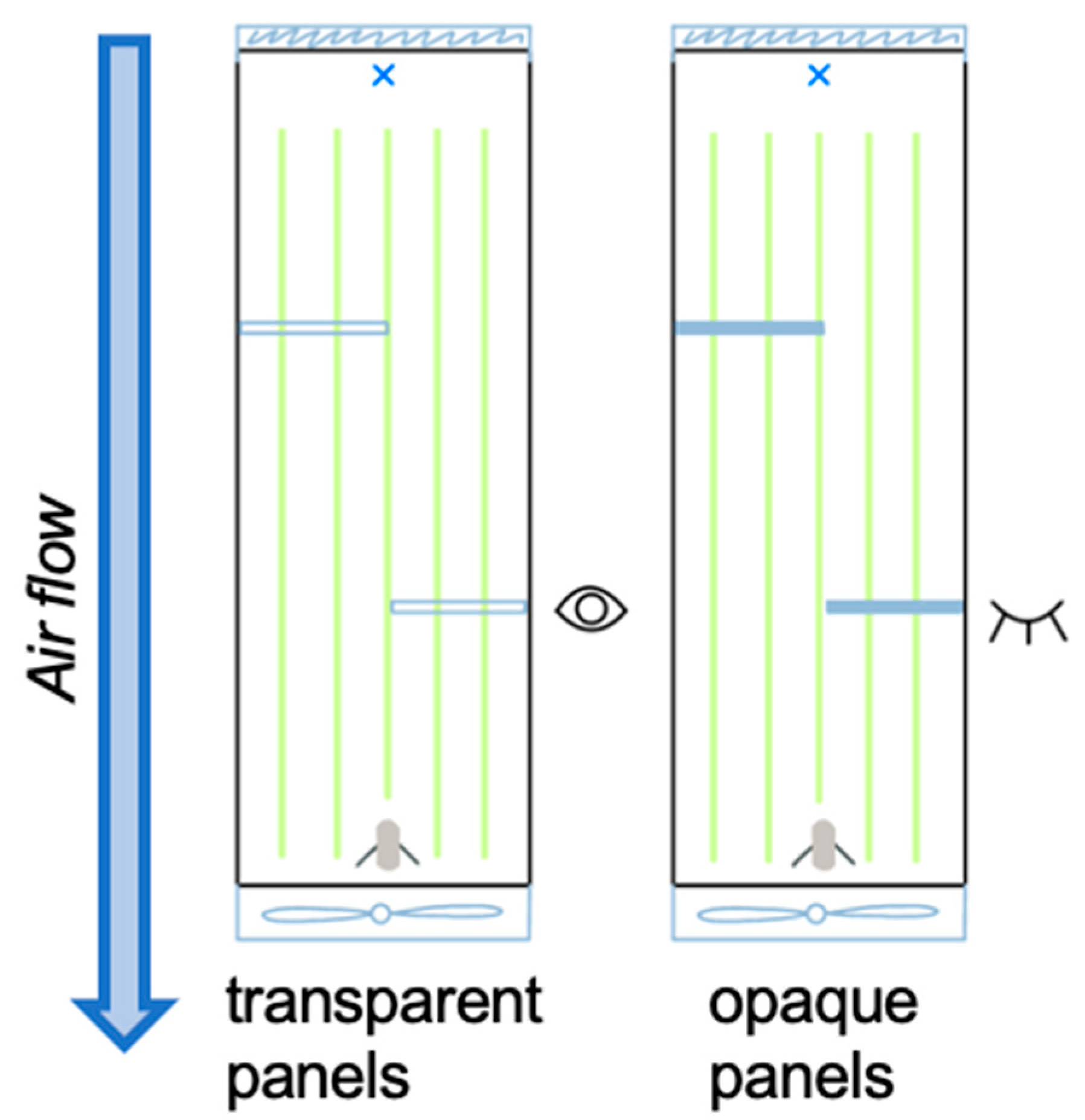
2.3. Wind Tunnel Experiments
2.4. Experimental Conditions
2.5. Statistical Analysis
3. Results
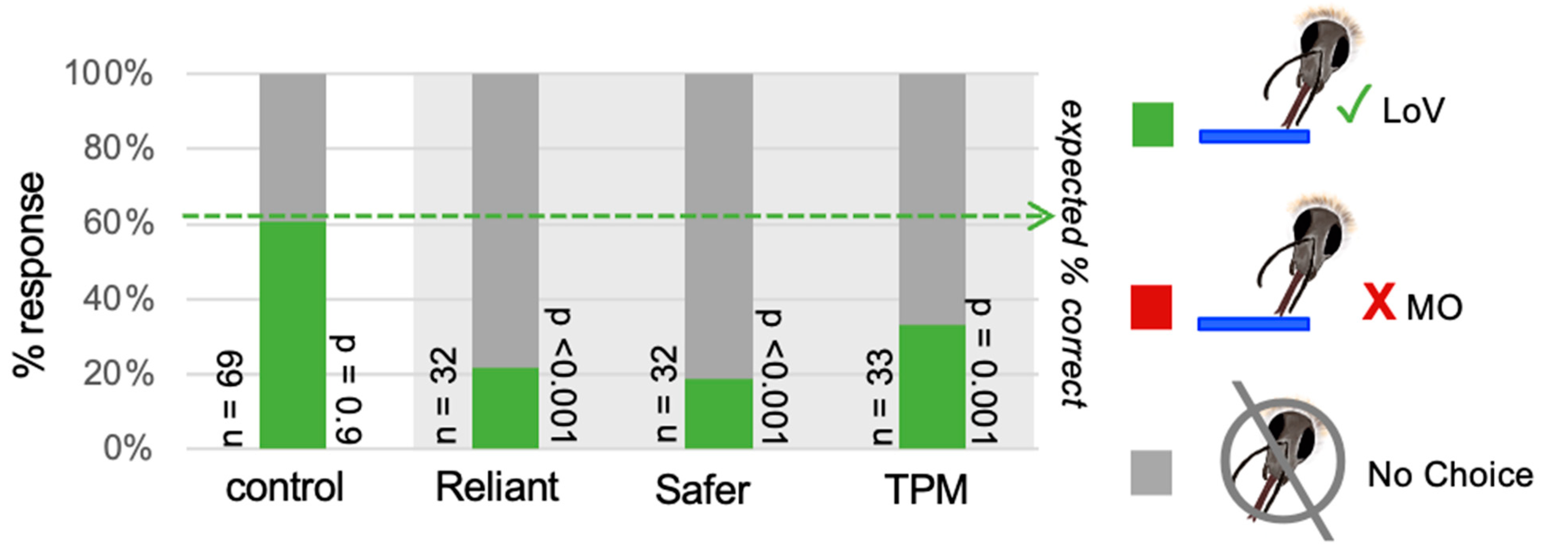
3.1. Fungicide Odor Pollution Reduces Bumblebee Responses to a Learned Floral Odor, Lily of the Valley, in an Associative Odor Learning Paradigm
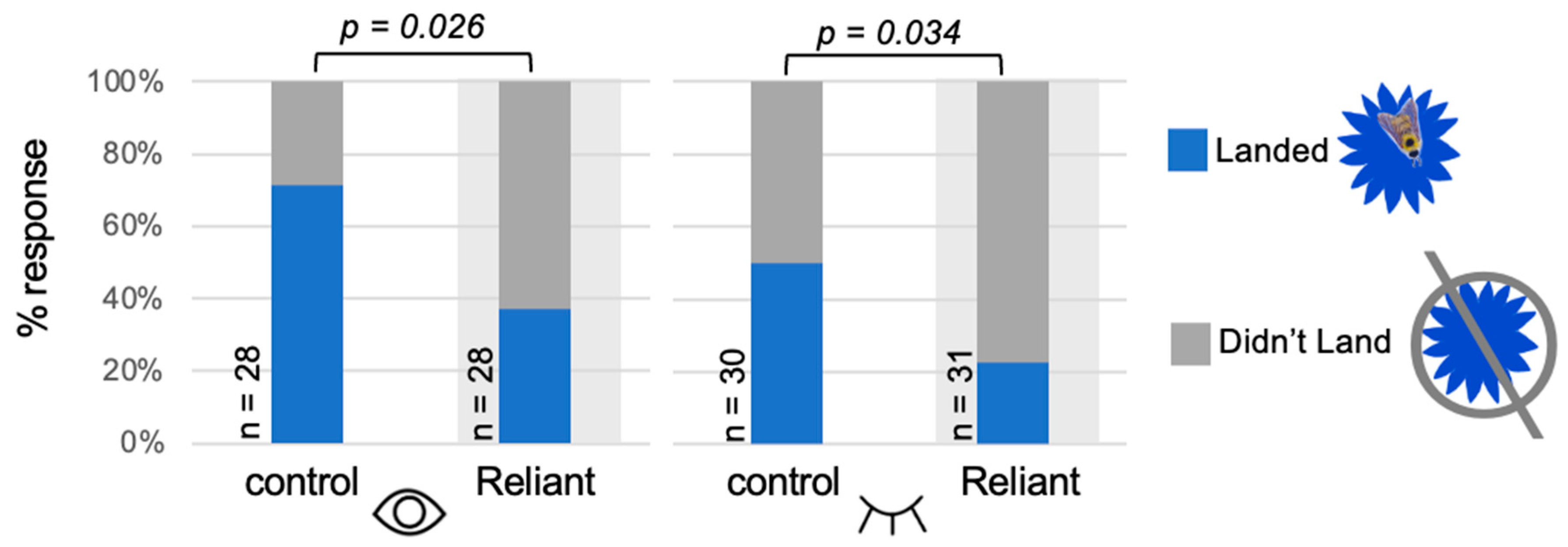
3.2. Fungicide Odor Pollution Reduces Bumblebee Landing Frequencies in Selection-and-Search Paradigms
4. Discussion
4.1. Foraging Behavior and Conditioned Odor Recognition Are Hindered by Fungicide Odor Pollution
4.2. Limitations and Implications
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The Importance of Pollinators; United States Department of Agriculture (USDA): Washington, DC, USA, 2022.
- Khalifa, S.A.M.; Elshafiey, E.H.; Shetaia, A.A.; El-Wahed, A.A.A.; Algethami, A.F.; Musharraf, S.G.; AlAjmi, M.F.; Zhao, C.; Masry, S.H.D.; Abdel-Daim, M.M.; et al. Overview of Bee Pollination and Its Economic Value for Crop Production. Insects 2021, 12, 688. [Google Scholar] [CrossRef]
- Cameron, S.A.; Sadd, B.M. Global Trends in Bumble Bee Health. Annu. Rev. Entomol. 2019, 65, 209–232. [Google Scholar] [CrossRef]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and Conservation of Bumble Bees. Annu. Rev. Èntomol. 2008, 53, 191–208. [Google Scholar] [CrossRef]
- Szabo, N.D.; Colla, S.R.; Wagner, D.L.; Gall, L.F.; Kerr, J.T. Do Pathogen Spillover, Pesticide Use, or Habitat Loss Explain Recent North American Bumblebee Declines? Conserv. Lett. 2012, 5, 232–239. [Google Scholar] [CrossRef]
- Becher, M.A.; Twiston-Davies, G.; Penny, T.D.; Goulson, D.; Rotheray, E.L.; Osborne, J.L. Bumble-BEEHAVE: A Systems Model for Exploring Multifactorial Causes of Bumblebee Decline at Individual, Colony, Population and Community Level. J. Appl. Ecol. 2018, 55, 2790–2801. [Google Scholar] [CrossRef]
- Soroye, P.; Newbold, T.; Kerr, J. Climate Change Contributes to Widespread Declines among Bumble Bees across Continents. Science 2020, 367, 685–688. [Google Scholar] [CrossRef]
- Vercelli, M.; Novelli, S.; Ferrazzi, P.; Lentini, G.; Ferracini, C. A Qualitative Analysis of Beekeepers’ Perceptions and Farm Management Adaptations to the Impact of Climate Change on Honey Bees. Insects 2021, 12, 228. [Google Scholar] [CrossRef]
- Carvell, C.; Bourke, A.F.G.; Dreier, S.; Freeman, S.N.; Hulmes, S.; Jordan, W.C.; Redhead, J.W.; Sumner, S.; Wang, J.; Heard, M.S. Bumblebee Family Lineage Survival Is Enhanced in High-Quality Landscapes. Nature 2017, 543, 547–549. [Google Scholar] [CrossRef]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee Declines Driven by Combined Stress from Parasites, Pesticides, and Lack of Flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Girling, R.D.; Lusebrink, I.; Farthing, E.; Newman, T.A.; Poppy, G.M. Diesel Exhaust Rapidly Degrades Floral Odours Used by Honeybees. Sci. Rep. 2013, 3, 2779. [Google Scholar] [CrossRef]
- Botías, C.; Jones, J.C.; Pamminger, T.; Bartomeus, I.; Hughes, W.O.H.; Goulson, D. Multiple Stressors Interact to Impair the Performance of Bumblebee Bombus terrestris Colonies. J. Anim. Ecol. 2021, 90, 415–431. [Google Scholar] [CrossRef]
- Cameron, S.A.; Lozier, J.D.; Strange, J.P.; Koch, J.B.; Cordes, N.; Solter, L.F.; Griswold, T.L. Patterns of Widespread Decline in North American Bumble Bees. Proc. National. Acad. Sci. USA 2011, 108, 662–667. [Google Scholar] [CrossRef]
- Rothman, J.A.; Russell, K.A.; Leger, L.; McFrederick, Q.S.; Graystock, P. The Direct and Indirect Effects of Environmental Toxicants on the Health of Bumblebees and Their Microbiomes. Proc. R. Soc. B 2020, 287, 20200980. [Google Scholar] [CrossRef]
- Siviter, H.; Richman, S.K.; Muth, F. Field-realistic Neonicotinoid Exposure Has Sub-lethal Effects on Non-Apis Bees: A Meta-analysis. Ecol. Lett. 2021, 24, 2586–2597. [Google Scholar] [CrossRef]
- Siviter, H.; Brown, M.J.F.; Leadbeater, E. Sulfoxaflor Exposure Reduces Bumblebee Reproductive Success. Nature 2018, 561, 109–112. [Google Scholar] [CrossRef]
- Siviter, H.; Pardee, G.L.; Baert, N.; McArt, S.; Jha, S.; Muth, F. Wild Bees Are Exposed to Low Levels of Pesticides in Urban Grasslands and Community Gardens. Sci. Total Environ. 2022, 858, 159839. [Google Scholar] [CrossRef]
- Siviter, H.; Johnson, A.K.; Muth, F. Bumblebees Exposed to a Neonicotinoid Pesticide Make Suboptimal Foraging Decisions. Environ. Entomol. 2021, 50, 1299–1303. [Google Scholar] [CrossRef]
- Siviter, H.; Bailes, E.J.; Martin, C.D.; Oliver, T.R.; Koricheva, J.; Leadbeater, E.; Brown, M.J.F. Agrochemicals Interact Synergistically to Increase Bee Mortality. Nature 2021, 596, 389–392. [Google Scholar] [CrossRef]
- USEPA. Insecticides; United States Environmental Protection Agency (EPA): Washington, DC, USA, 2022.
- Botías, C.; David, A.; Hill, E.M.; Goulson, D. Quantifying Exposure of Wild Bumblebees to Mixtures of Agrochemicals in Agricultural and Urban Landscapes. Environ. Pollut. 2017, 222, 73–82. [Google Scholar] [CrossRef]
- Muth, F.; Leonard, A.S. A Neonicotinoid Pesticide Impairs Foraging, but Not Learning, in Free-Flying Bumblebees. Sci. Rep. 2019, 9, 4764. [Google Scholar] [CrossRef]
- Phelps, J.D.; Strang, C.G.; Sherry, D.F. Imidacloprid Impairs Performance on a Model Flower Handling Task in Bumblebees (Bombus impatiens). Ecotoxicology 2020, 29, 359–374. [Google Scholar] [CrossRef]
- Lämsä, J.; Kuusela, E.; Tuomi, J.; Juntunen, S.; Watts, P.C. Low Dose of Neonicotinoid Insecticide Reduces Foraging Motivation of Bumblebees. Proc. R. Soc. B Biol. Sci. 2018, 285, 20180506. [Google Scholar] [CrossRef]
- Tamburini, G.; Pereira-Peixoto, M.-H.; Borth, J.; Lotz, S.; Wintermantel, D.; Allan, M.J.; Dean, R.; Schwarz, J.M.; Knauer, A.; Albrecht, M.; et al. Fungicide and Insecticide Exposure Adversely Impacts Bumblebees and Pollination Services under Semi-Field Conditions. Environ. Int. 2021, 157, 106813. [Google Scholar] [CrossRef]
- Scholer, J.; Krischik, V. Chronic Exposure of Imidacloprid and Clothianidin Reduce Queen Survival, Foraging, and Nectar Storing in Colonies of Bombus impatiens. PLoS ONE 2014, 9, e91573. [Google Scholar] [CrossRef]
- McFrederick, Q.S.; Kathilankal, J.C.; Fuentes, J.D. Air Pollution Modifies Floral Scent Trails. Atmos. Environ. 2008, 42, 2336–2348. [Google Scholar] [CrossRef]
- Fuentes, J.D.; Chamecki, M.; Roulston, T.; Chen, B.; Pratt, K.R. Air Pollutants Degrade Floral Scents and Increase Insect Foraging Times. Atmos. Environ. 2016, 141, 361–374. [Google Scholar] [CrossRef]
- Ryalls, J.M.W.; Langford, B.; Mullinger, N.J.; Bromfield, L.M.; Nemitz, E.; Pfrang, C.; Girling, R.D. Anthropogenic Air Pollutants Reduce Insect-Mediated Pollination Services. Environ. Pollut. 2022, 297, 118847. [Google Scholar] [CrossRef]
- Sprayberry, J.D.H.; Ritter, K.A.; Riffell, J.A. The Effect of Olfactory Exposure to Non-Insecticidal Agrochemicals on Bumblebee Foraging Behavior. PLoS ONE 2013, 8, e76273. [Google Scholar] [CrossRef]
- David, N.F.; Henry, T.J.; Sprayberry, J.D.H. Odor-Pollution From Fungicides Disrupts Learning and Recognition of a Common Floral Scent in Bumblebees (Bombus impatiens). Front. Ecol. Evol. 2022, 10, 765388. [Google Scholar] [CrossRef]
- Sprayberry, J.D.H. Compounds without Borders: A Mechanism for Quantifying Complex Odors and Responses to Scent-Pollution in Bumblebees. PLoS Comput. Biol. 2020, 16, e1007765. [Google Scholar] [CrossRef]
- Sommer, J.; Rao, V.; Sprayberry, J. Deconstructing and Contextualizing Foraging Behavior in Bumble Bees and Other Central Place Foragers. Apidologie 2022, 53, 32. [Google Scholar] [CrossRef]
- Muth, F.; Carvalheiro, L.; Cooper, T.R.; Bonilla, R.F.; Leonard, A.S. A Novel Protocol for Studying Bee Cognition in the Wild. Methods Ecol. Evol. 2017, 9, 78–87. [Google Scholar] [CrossRef]
- Baird, E.; Boeddeker, N.; Srinivasan, M.V. The Effect of Optic Flow Cues on Honeybee Flight Control in Wind. Proc. R. Soc. B 2021, 288, 20203051. [Google Scholar] [CrossRef] [PubMed]
- Sprayberry, J.D.H. The Prevalence of Olfactory- versus Visual-Signal Encounter by Searching Bumblebees. Sci. Rep. 2018, 8, 14590. [Google Scholar] [CrossRef] [PubMed]
- Amrhein, V.; Greenland, S.; McShane, B. Scientists Rise up against Statistical Significance. Nature 2019, 567, 305–307. [Google Scholar] [CrossRef] [PubMed]
- Ciarlo, T.J.; Mullin, C.A.; Frazier, J.L.; Schmehl, D.R. Learning Impairment in Honey Bees Caused by Agricultural Spray Adjuvants. PLoS ONE 2012, 7, e40848. [Google Scholar] [CrossRef]
- Gill, R.J.; Ramos-Rodriguez, O.; Raine, N.E. Combined Pesticide Exposure Severely Affects Individual- and Colony-Level Traits in Bees. Nature 2013, 490, 105–108. [Google Scholar] [CrossRef]
- De Bivort, B.L.; van Swinderen, B. Evidence for Selective Attention in the Insect Brain. Curr. Opin. Insect. Sci. 2016, 15, 9–15. [Google Scholar] [CrossRef]
- Vinauger, C.; Breugel, F.V.; Locke, L.T.; Tobin, K.K.S.; Dickinson, M.H.; Fairhall, A.L.; Akbari, O.S.; Riffell, J.A. Visual-Olfactory Integration in the Human Disease Vector Mosquito Aedes Aegypti. Curr. Biol. 2019, 29, 2509–2516.e5. [Google Scholar] [CrossRef]
- Riffell, J.A. The Neuroecology of Insect-Plant Interactions: The Importance of Physiological State and Sensory Integration. Curr. Opin. Insect. Sci. 2020, 42, 118–124. [Google Scholar] [CrossRef]
- Reitmayer, C.M.; Ryalls, J.M.W.; Farthing, E.; Jackson, C.W.; Girling, R.D.; Newman, T.A. Acute Exposure to Diesel Exhaust Induces Central Nervous System Stress and Altered Learning and Memory in Honey Bees. Sci. Rep. 2019, 9, 5793. [Google Scholar] [CrossRef] [PubMed]
- Lusebrink, I.; Girling, R.D.; Farthing, E.; Newman, T.A.; Jackson, C.W.; Poppy, G.M. The Effects of Diesel Exhaust Pollution on Floral Volatiles and the Consequences for Honey Bee Olfaction. J. Chem. Ecol. 2015, 41, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Démares, F.; Gibert, L.; Creusot, P.; Lapeyre, B.; Proffit, M. Acute Ozone Exposure Impairs Detection of Floral Odor, Learning, and Memory of Honey Bees, through Olfactory Generalization. Sci. Total Environ. 2022, 827, 154342. [Google Scholar] [CrossRef] [PubMed]
- Saunier, A.; Grof-Tisza, P.; Blande, J.D. Effect of Ozone Exposure on the Foraging Behaviour of Bombus terrestris. Environ. Pollut. 2022, 316, 120573. [Google Scholar] [CrossRef]
- Lihoreau, M.; Collett, T.; Raine, N.E.; Reynolds, A.M.; Stelzer, R.J.; Lim, K.S.; Smith, A.D.; Osborne, J.L.; Chittka, L. Radar Tracking and Motion-Sensitive Cameras on Flowers Reveal the Development of Pollinator Multi-Destination Routes over Large Spatial Scales. PLoS Biol. 2012, 10, e1001392. [Google Scholar] [CrossRef]
- Lihoreau, M.; Chittka, L.; Raine, N.E. Travel Optimization by Foraging Bumblebees through Readjustments of Traplines after Discovery of New Feeding Locations. Am. Nat. 2010, 176, 744–757. [Google Scholar] [CrossRef]
- Ohashi, K.; Thomson, J.D.; D’Souza, D. Trapline Foraging by Bumble Bees: IV. Optimization of Route Geometry in the Absence of Competition. Behav. Ecol. 2007, 18, 1–11. [Google Scholar] [CrossRef]
- Saleh, N.; Chittka, L. Traplining in Bumblebees (Bombus impatiens): A Foraging Strategy’s Ontogeny and the Importance of Spatial Reference Memory in Short-Range Foraging. Oecologia 2006, 151, 719–730. [Google Scholar] [CrossRef]
- Woodgate, J.L.; Makinson, J.C.; Lim, K.S.; Reynolds, A.M.; Chittka, L. Continuous Radar Tracking Illustrates the Development of Multi-Destination Routes of Bumblebees. Sci. Rep. 2017, 7, 17323. [Google Scholar] [CrossRef]
- Alignier, A.; Lenestour, N.; Jeavons, E.; van Baaren, J.; Aviron, S.; Uroy, L.; Ricono, C.; Lann, C.L. Floral Resource Maps: A Tool to Explain Flower-Visiting Insect Abundance at Multiple Spatial Scales. Landsc. Ecol. 2023, 1–15. [Google Scholar] [CrossRef]
- Osborne, J.L.; Clark, S.J.; Morris, R.J.; Williams, I.H.; Riley, J.R.; Smith, A.D.; Reynolds, D.R.; Edwards, A.S. A Landscape-scale Study of Bumble Bee Foraging Range and Constancy, Using Harmonic Radar. J. Appl. Ecol. 1999, 36, 519–533. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungicide | %C/I/NR | Sample Size | p-Value (Compared to Expected) | Post Hoc: Correct p-Value | Post Hoc: Incorrect p-Value | Post Hoc: No Choice p-Value |
|---|---|---|---|---|---|---|
| Control (no fungicide) | 61/0/39 | 75 | 0.002 | 0.9 | 0.004 | 0.11 |
| Reliant | 22/0/78 | 32 | <0.001 | <0.001 | 0.11 | <0.001 |
| Safer | 19/0/81 | 35 | <0.001 | <0.001 | 0.11 | <0.001 |
| Scotts | 33/0/67 | 33 | <0.001 | 0.001 | 0.11 | <0.001 |
| Wind Tunnel Configuration | Treatment | Average Time to Flower (min) | n (Bees That Landed) | p Value (t-Test) |
|---|---|---|---|---|
| Transparent | Control | 3.11 | 15 | 0.75 |
| Reliant | 3.49 | 6 | ||
| Opaque | Control | 2.6 | 15 | 0.55 |
| Reliant | 3.42 | 7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yousry, N.; Henderson, P.; Sprayberry, J. Fungicide Scent Pollution Disrupts Floral Search-and-Selection in the Bumblebee Bombus impatiens. Agrochemicals 2023, 2, 181-192. https://doi.org/10.3390/agrochemicals2020013
Yousry N, Henderson P, Sprayberry J. Fungicide Scent Pollution Disrupts Floral Search-and-Selection in the Bumblebee Bombus impatiens. Agrochemicals. 2023; 2(2):181-192. https://doi.org/10.3390/agrochemicals2020013
Chicago/Turabian StyleYousry, Nour, Paige Henderson, and Jordanna Sprayberry. 2023. "Fungicide Scent Pollution Disrupts Floral Search-and-Selection in the Bumblebee Bombus impatiens" Agrochemicals 2, no. 2: 181-192. https://doi.org/10.3390/agrochemicals2020013