

Multiple Pesticide Resistance in Rust-Red Flour Beetle (Tribolium castaneum, Herbst 1797) from Northern Nigeria Is Probably Driven by Metabolic Mechanisms



Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Rearing of Beetles
2.2. Morphological Identification of the Beetles
2.3. Insecticides Susceptibility Filter Paper Bioassays
2.4. Investigation of the Role of Metabolic Resistance Using Synergist Bioassay
2.5. Investigation of the Role of Target-Site Pyrethroid/DDT Insensitivity Resistance Mutations
2.6. Molecular Identification of the Beetles to Species Level
2.7. Data Analysis
3. Results
3.1. Morphological and Molecular Identification of Beetles to Species Level
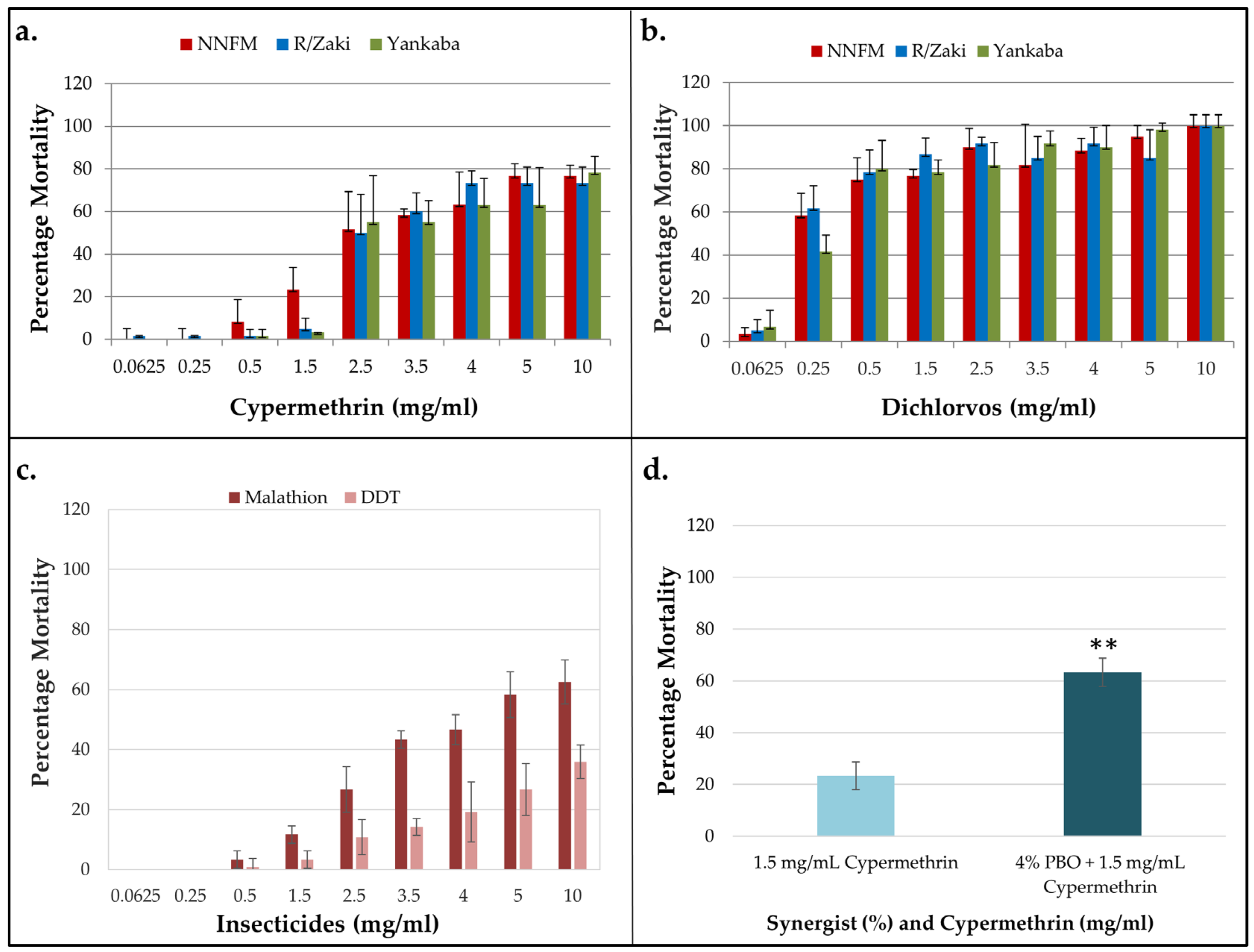
3.2. Insecticides Resistance Profile of the T. castaneum Population
3.3. Assessment of the Role of Metabolic Resistance in Cypermethrin Resistance
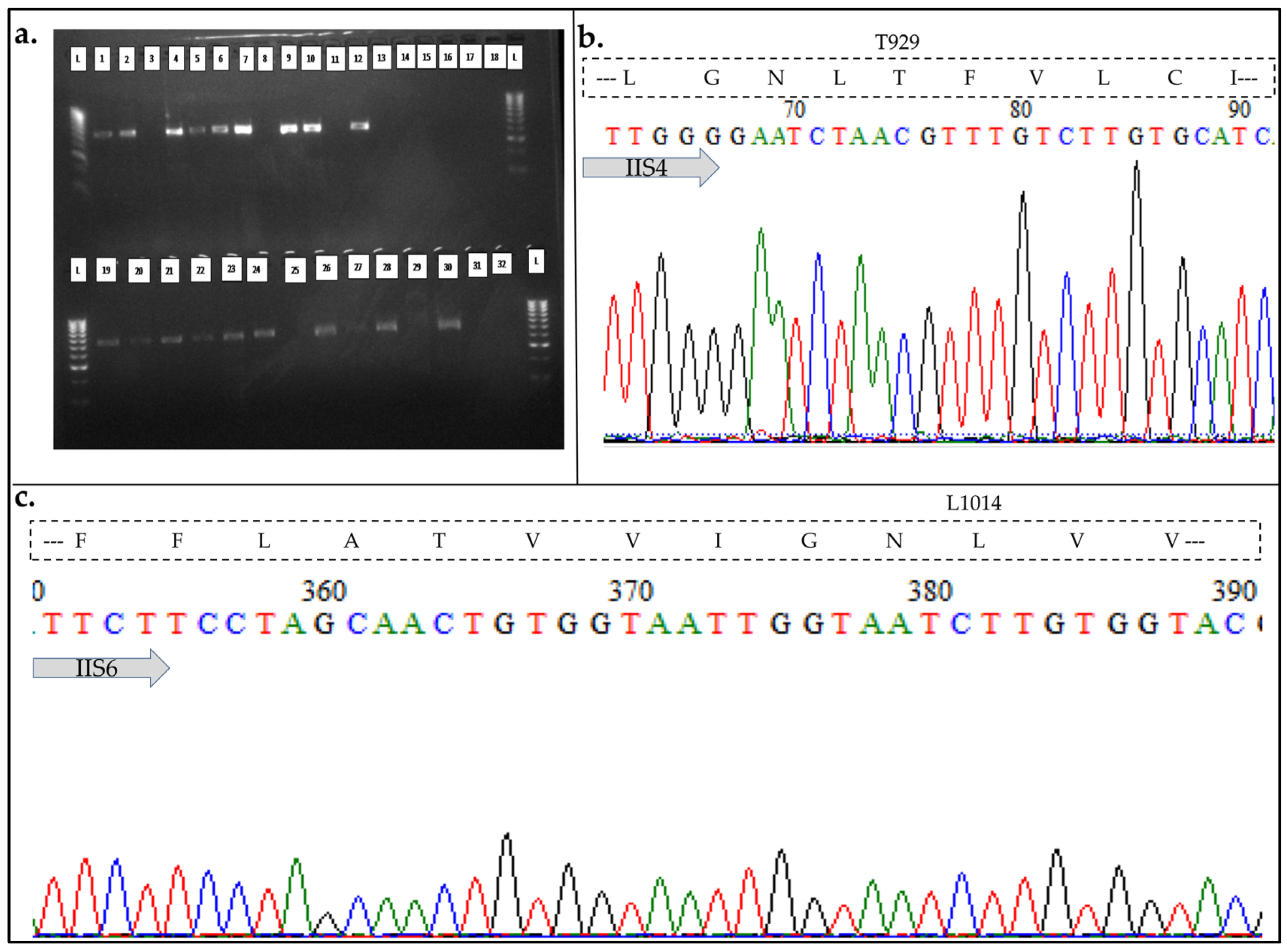
3.4. Assessment of the Role of VGSC Target-Site Insensitivity kdr Mutations in Cypermethrin Resistance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lyddon, C. Focus on Nigeria; Sosland Publishing Company: Kansas, MO, USA, 2022; Volume 4, pp. 88–100. Available online: https://world-grain.com/ (accessed on 22 January 2022).
- Reidy, J. Nigeria Wheat Initiative Reaches First-Year Goal; Sosland Publishing Company: Kansas, MO, USA, 2022; Available online: https://www.world-grain.com/articles/17449-nigeria-wheat-initiative-reaches-first-year-goal# (accessed on 9 December 2022).
- Turaki, J.M.; Sastawa, B.M.; Kabir, B.G.J.; Lale, N.E.S. Susceptibility of flours derived from various cereal grains to infestation by the rust-red flour beetle (Tribolium castaneum Herbst) (Coleoptera: Tenebrionidae) in different seasons. J. Plant Prot. Res. 2007, 47, 279–288. Available online: http://www.plantprotection.pl/Susceptibility-of-flours-derived-from-various-cereal-grains-to-infestation-by-the,91119,0,2.html (accessed on 9 December 2022).
- Mailafiya, D.M.; Maina, Y.T.; Degri, Y.M.; Gadzama, U.U. Traders’ perception of food grain storage and pest management in Dalwa market, Borno State, Nigeria. J. Agric. Crop Res. 2014, 2, 62–70. [Google Scholar]
- Jagadeesan, R.; Collins, P.J.; Daglish, G.J.; Ebert, P.R.; Schlipalius, D.I. Phosphine resistance in the rust red flour beetle, Tribolium castaneum (Coleoptera: Tenebrionidae): Inheritance, gene interactions and fitness costs. PLoS ONE 2012, 7, 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, R.; Hassan, S.; Ahmad, S.; Nighat, S.; Devi, Y.K.; Javeed, K.; Usmani, S.; Ansari, M.J.; Erturk, S.; Alkan, M.; et al. Stored Grain Pests and Current Advances for Their Management. In Postharvest Technology—Recent Advances, New Perspectives and Applications; IntechOpen: London, UK, 2022. [Google Scholar] [CrossRef]
- Baldwin, R.; Fasulo, T.R. Confused Flour Beetle, Tribolium confusum Jacquelin du Val and Red Flour Beetle, Tribolium castaneum (Herbst) (Insecta: Coleoptera: Tenebrionidae). Edis 2020, 2005, 1–5. [Google Scholar] [CrossRef]
- Pires, E.M.; Souza, E.Q.; Nogueira, R.M.; Soares, M.A.; Dias, T.K.R.; Oliveira, M.A. Damage caused by Tribolium castaneum (Coleoptera: Tenebrionidae) in Stored Brazil nut. Sci. Electron. Arch. 2017, 10, 1–5. [Google Scholar] [CrossRef]
- Campbell, J.F.; Runnion, C. Patch exploitation by female red flour beetles, Tribolium castaneum. J. Insect Sci. 2003, 3, 20. [Google Scholar] [CrossRef] [Green Version]
- Nafiu, M.I.; Lawal, N.; Aminu, A.M.; Muohammed, A.S.; Abdullahi, R.I.; Bello, T.A. Productivity of Tribolium Species (Coleoptera; Tenebrionidae) within Flours Derived from Edible Tubers and Effects on Proximate Composition. Int. J. Sci. Glob. Sustain. 2020, 6, 12. Available online: https://fugus-ijsgs.com.ng/index.php/ijsgs/article/view/68 (accessed on 12 September 2022).
- Ajayi, F.A.; Rahman, S.A. Susceptibility of some staple processed meals to red flour beetle, Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae). Pak. J. Biol. Sci. 2006, 9, 1744–1748. [Google Scholar] [CrossRef] [Green Version]
- Ehisianya, C.N.; Stephen, A.G.; Onunka, B.N. Development of Tribolium castaneum Herbst (Coleoptera: Tenebrionidae) and Damage to Selected Flours in Storage. Nig. Agric. J. 2022, 53, 193–198. Available online: https://www.ajol.info/index.php/naj/article/view/232673 (accessed on 12 September 2022).
- Vojoudi, S.; Saber, M.; Mahdavi, V.; Golshan, H.; Abedi, Z. Efficacy of some Insecticides Against Red Flour Beetle, Tribolium castaneum Herbst (Coleoptera: Tenebrionidae) Adults Exposed on Glass, Ceramic Tile, Plastic and Paper Disc Surfaces. J. Lif. Sci. 2012, 6, 405–410. [Google Scholar]
- Pieterse, A.H.; Schulten, G.G.M.; Kuyken, W. A study on insecticide resistance in Tribolium castaneum (Herbst) (Coleoptera, Tenebrionidae) in Malawi (Central Africa). J. Stored Prod. Res. 1972, 8, 183–191. [Google Scholar] [CrossRef]
- Rauf, A.; Wilkins, R.M. Malathion-resistant Tribolium castaneum has enhanced response to oxidative stress, immunity, and fitness. Pest. Biochem. Physiol. 2022, 184, 105128. [Google Scholar] [CrossRef]
- Shamjana, U.; Grace, T. Review of Insecticide Resistance and Its Underlying Mechanisms in Tribolium castaneum,” Insecticides—Impact and Benefits of Its Use for Humanity. In Review of Insecticide Resistance and Its Underlying Mechanisms in Tribolium castaneum; IntechOpen: London, UK, 2022. [Google Scholar] [CrossRef]
- Dyte, C.E.; Blackman, D.G. The spread of insecticide resistance in Tribolium castaneum (Herbst) (Coleoptera, Tenebrionidae). J. Stored Prod. Res. 1970, 6, 255–261. [Google Scholar] [CrossRef]
- Attia, M.A.; Wahba, T.F.; Shaarawy, N.; Moustafa, F.I.; Guedes, R.N.C.; Dewer, Y. Stored grain pest prevalence and insecticide resistance in Egyptian populations of the red flour beetle Tribolium castaneum (Herbst) and the rice weevil Sitophilus oryzae (L.). J. Stored Prod. Res. 2020, 87, 101611. [Google Scholar] [CrossRef]
- Ayo, F.K.; Hezekiah, A.S.; Temitope, A.A. Comparative Pesticidal Activities of Essential Oils Extracted from Indigenous Plants Against Tribolium castaneum Herbst (Coleoptera: Tenebrionidae). Asian J. Sci. Res. 2019, 12, 502–507. [Google Scholar] [CrossRef] [Green Version]
- Ukeh, D.A.; Umoetoka, S.B.A. Repellent effects of five monoterpenoid odours against Tribolium castaneum (Herbst) and Rhyzopertha dominica (F.) in Calabar, Nigeria. Crop Prot. 2011, 30, 1351–1355. [Google Scholar] [CrossRef]
- Gbaye, O.A.; Oyeniyi, E.A.; Ogunleye, E.I.; Aloba, O.V. The impact of cassava and wheat flour substrates on the biological parameters and insecticide response in Tribolium castaneum (Herbst). Biocatal. Agric. Biotechnol. 2021, 38, 102194. [Google Scholar] [CrossRef]
- Beeman, R.W.; Haas, S.; Friesen, K. An Introduction to the Care and Handling of Tribolium castaneum; USDA Tribolium Stock Maintenance: Washington, DC, USA, 2022. Available online: https://www.ars.usda.gov/plains-area/mhk/cgahr/spieru/docs/tribolium-stock-maintenance/ (accessed on 12 November 2022).
- Dia, C.A.K.M.; Sarr, A.G.R.J.; Kafom, A.; Ngom, D.; Diome, T.; Thiaw, C.; Ndiaye, S.; Sembene, M. Morphological Identification of Trophic Tribolium Castaneum Populations Herbst (Coleoptera, Tenebrionidae) in West Africa. Int. J. Adv. Res. 2018, 6, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Parthasarathy, R.; Tan, A.; Bai, H.; Palli, S.R. Transcription factor broad suppresses precocious development of adult structures during larval–pupal metamorphosis in the red flour beetle, Tribolium castaneum R. Mech. Dev. 2008, 125, 299–313. [Google Scholar] [CrossRef] [Green Version]
- IRAC. IRAC Test Methods Series. Test No. 006. Insecticides Resistance Action Committee Susceptibility. 2009. Version 3. Available online: https://irac-online.org/content/uploads/2009/09/Method_006_v3_june09.pdf (accessed on 2 February 2022).
- Bala, I.; Mukhtar, M.M.; Saka, H.K.; Abdullahi, N.; Ibrahim, S.S. Determination of insecticide susceptibility of field populations of tomato leaf miner (Tuta absoluta) in northern Nigeria. Agriculture 2019, 9, 7. [Google Scholar] [CrossRef] [Green Version]
- Haddi, K.; Valbon, W.R.; Jumbo, L.O.V.; de Oliveira, L.O.; Guedes, R.N.C.; Oliveira, E.E. Diversity and convergence of mechanisms involved in pyrethroid resistance in the stored grain weevils, Sitophilus spp. Sci. Rep. 2018, 8, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araújo, R.A.; Williamson, M.S.; Bass, C.; Field, L.M.; Duce, I.R. Pyrethroid resistance in Sitophilus zeamais is associated with a mutation (T929I) in the voltage-gated sodium channel. Insect Mol. Biol. 2011, 20, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J. Organization and mapping of a sequence on the Drosophila melanogaster X and Y chromosomes that is transcribed during spermatogenesis. Genetics 1984, 107, 611–634. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, Y.-J.; Guo, W.; Luo, D.; Wu, Y.; Kučerová, Z.; Stejskal, V.; Opit, G.; Cao, Y.; Li, F.-J.; et al. DNA barcoding, species-specific PCR and real-time PCR techniques for the identification of six Tribolium pests of stored products. Sci. Rep. 2016, 6, 28494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Èntomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Hodges, R.J.; Buzby, J.C.; Bennett, B. Postharvest losses and waste in developed and less developed countries: Opportunities to improve resource use. J. Agric. Sci. 2011, 149, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Ming, Q.; Wang, A.; Cheng, C. Molecular identification of Tribolium castaneum and T.confusum (Coleoptera: Tenebrionidae) using PCR-RFLP analysis. J. Genet. 2014, 93, e17–e21. Available online: https://pubmed.ncbi.nlm.nih.gov/24823304/ (accessed on 5 January 2023).
- Kayode, O.Y.; Adedire, C.O.; Akinkurolere, R.O. Influence of four cereal flours on the growth of Tribolium castaneum Herbst (Coleoptera: Tenebrionidae). Ife J. Sci. 2014, 16, 505–516. [Google Scholar]
- Rossi, E.; Cosimi, S.; Loni, A. Insecticide resistance in Italian populations of Tribolium flour beetles. Bull. Insectology 2010, 63, 251–258. [Google Scholar]
- Abo, O.; Ja, E. An evaluation of infestation of insect pests of flours in Benin City, Edo State, Nigeria. J. Appl. Sci. Environ. Manag. 2014, 18, 487–494. Available online: https://www.ajol.info/index.php/jasem/article/view/109917 (accessed on 5 January 2023).
- McFarlane, J.A. Guidelines for Pest Management to Reduce Stored Food Losses Caused by Insects and Mites; Greenwich Academic Literature Archive (GALA): London, UK, 1989; p. 62. Available online: https://gala.gre.ac.uk/id/eprint/10727/1/Doc-0115.pdf (accessed on 5 January 2023).
- Abouelkassem, S.; Salem, A.A.; Arab, A.R.B. Toxicity and development of resistance in Tribolium castaneum and Sitophilus oryzae oryzae to certain selected insecticides. Egypt. J. Plant Prot. Res. Inst. 2018, 1, 188–198. Available online: http://www.ejppri.eg.net/pdf/v1n2/11.pdf (accessed on 5 January 2023).
- Naeem, M.; Anjum, S.I.; Ismail, M.; Khan, J.; Bibi, S. Laboratory assessment of different botanical extracts and cypermethrin against insect pests. J. Entomol. Zool. Stud. 2015, 3, 84–88. Available online: https://www.entomoljournal.com/archives/2015/vol3issue5/PartB/3-5-36.pdf (accessed on 5 January 2023).
- Khalequzzaman, M.; Khanom, M. Effects of cypermethrin alone and in combination with leaf and seed extracts of neem against adult Tribolium castaneum (Herbst). Univ. J. Zool. Rajshahi Univ. 2006, 25, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Andrić, G.; Kljajić, P.; Perić, I.; Golić, M.P. Susceptibility of red flour beetle Tribolium castaneum (Herbst) populations from Serbia to contact insecticides. In Proceedings of the 10th International Working Conference on Stored Product Protection, Estoril, Portugal, 27 June–2 July 2010; Volume 425, pp. 869–873. [Google Scholar] [CrossRef]
- Reddy, D.; Srivastava, C. Persistent Toxicity of Malathion and Dichlorvos on Jute Surface Against Tribolium castaneum (Herbst). Ann. Plant Prot. Sci. 2004, 12, 41–44. Available online: https://www.indianjournals.com/ijor.aspx?target=ijor:apps&volume=12&issue=1&article=011 (accessed on 6 January 2023).
- Parkin, E.A.; Scott, E.I.C.; Forster, R. Increased resistance of stored-product insects to insecticides. The resistance of field strains of beetles. Pest Infest. Res. 1961, 34–35, 1962. [Google Scholar]
- Champ, B.R.; Campbell-Brown, M.J. Insecticide resistance in Australian Tribolium castaneum (Herbst) (Coleoptera, Tenebrionidae)—II. Malathion resistance in eastern Australia. J. Stored Prod. Res. 1970, 6, 111–131. [Google Scholar] [CrossRef]
- Greenin, H.G. Malathion Resistance in the Red Flour Beetle. Aust. J. Entomol. 1970, 9, 160–162. [Google Scholar] [CrossRef]
- Dyte, C.E.; Blackman, D.G. Selection of a DDT-resistant strain of Tribolium castaneum (Herbst) (Coleoptera, Tenebrionidae). J. Stored Prod. Res. 1967, 2, 211–228. [Google Scholar] [CrossRef]
- Feyereisen, R. Insect P450 inhibitors and insecticides: Challenges and opportunities. Pest Manag. Sci. 2015, 71, 793–800. [Google Scholar] [CrossRef]
- Ibrahim, S.S.; Mukhtar, M.M.; Irving, H.; Riveron, J.M.; Fadel, A.N.; Tchapga, W.; Hearn, J.; Muhammad, A.; Sarkinfada, F.; Wondji, C.S. Exploring the mechanisms of multiple insecticide resistance in a highly plasmodium-infected malaria vector Anopheles funestus sensu stricto from Sahel of northern Nigeria. Genes 2020, 11, 454. [Google Scholar] [CrossRef] [Green Version]
- Mukhtar, M.M.; Ibrahim, S.S. Temporal Evaluation of Insecticide Resistance in Populations of the Major Arboviral Vector Aedes Aegypti from Northern Nigeria. Insects 2022, 13, 187. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Parthasarathy, R.; Bai, H.; Woithe, K.; Kaussmann, M.; Nauen, R.; Harrison, D.A.; Palli, S.R. A brain-specific cytochrome P450 responsible for the majority of deltamethrin resistance in the QTC279 strain of Tribolium castaneum. Proc. Natl. Acad. Sci. USA. 2010, 107, 8557–8562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmer, C.T.; Nauen, R. Cytochrome P450 mediated pyrethroid resistance in European populations of Meligethes aeneus (Coleoptera: Nitidulidae). Pestic. Biochem. Physiol. 2011, 100, 264–272. [Google Scholar] [CrossRef]
- Rösner, J.; Tietmeyer, J.; Merzendorfer, H. Functional analysis of ABCG and ABCH transporters from the red flour beetle, Tribolium castaneum. Pest Manag. Sci. 2021, 77, 2955–2963. [Google Scholar] [CrossRef]
- Dong, K.; Du, Y.; Rinkevich, F.; Nomura, Y.; Xu, P.; Wang, L.; Silver, K.; Zhorov, B.S. Molecular biology of insect sodium channels and pyrethroid resistance. Insect Biochem. Mol. Biol. 2014, 50, 1–17. [Google Scholar] [CrossRef] [Green Version]
- El Halim, H.M.A.; Alshukri, B.M.H.; Ahmad, M.S.; Nakasu, E.Y.T.; Awwad, M.H.; Salama, E.M.; Gatehouse, A.M.R.; Edwards, M.G. RNAi-mediated knockdown of the voltage gated sodium ion channel TcNav causes mortality in Tribolium castaneum. Sci. Rep. 2016, 6, 29301. [Google Scholar] [CrossRef]
- Rösner, J.; Wellmeyer, B.; Merzendorfer, H. Tribolium castaneum: A Model for Investigating the Mode of Action of Insecticides and Mechanisms of Resistance. Curr. Pharm. Des. 2020, 26, 3554–3568. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Insecticide | Population | LC50 (mg/mL) | 95% CI | LC90 (mg/m) | 95% CI | Slope (SE) | χ2 | df | p |
|---|---|---|---|---|---|---|---|---|---|
| Cypermethrin | NNFM | 4.35 | 3.68–5.01 | 12.74 | 7.90–20.87 | 2.27 (0.18) | 6.48 | 7 | 0.37 |
| R/Zaki | 4.45 | 3.83–5.02 | 13.45 | 6.99–26.25 | 2.02 (0.14) | 14.21 | 7 | 0.50 | |
| Yankaba | 5.46 | 4.64–6.27 | 12.81 | 7.35–20.06 | 1.76 (0.12) | 41.45 | 7 | 0.40 | |
| Dichlorvos | NNFM | 0.28 | 0.07–0.63 | 4.94 | 1.62–9.12 | 1.33 (0.08) | 50.26 | 7 | 0.1 |
| R/Zaki | 0.17 | 0.03–0.46 | 3.83 | 1.39–10.76 | 1.26 (0.28) | 60.86 | 7 | 0.09 | |
| Yankaba | 0.35 | 0.06–0.63 | 2.67 | 1.59–5.71 | 1.47 (0.89) | 36.22 | 7 | 0.15 | |
| Malathion | NNFM | 3.71 | 1.72–5.92 | 15.41 | 11.46–23.85 | 2.313 (0.24) | 3.56 | 7 | 0.73 |
| DDT | NNFM | 15.32 | 5.09–20.33 | 47.63 | 23.82–192.19 | 2.001 (0.35) | 1.41 | 7 | 0.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mukhtar, M.M.; Mustapha, M.A.; Aliyu, M.; Ibrahim, S.S. Multiple Pesticide Resistance in Rust-Red Flour Beetle (Tribolium castaneum, Herbst 1797) from Northern Nigeria Is Probably Driven by Metabolic Mechanisms. Agrochemicals 2023, 2, 170-180. https://doi.org/10.3390/agrochemicals2020012
Mukhtar MM, Mustapha MA, Aliyu M, Ibrahim SS. Multiple Pesticide Resistance in Rust-Red Flour Beetle (Tribolium castaneum, Herbst 1797) from Northern Nigeria Is Probably Driven by Metabolic Mechanisms. Agrochemicals. 2023; 2(2):170-180. https://doi.org/10.3390/agrochemicals2020012
Chicago/Turabian StyleMukhtar, Muhammad M., Muhammad A. Mustapha, Mubarak Aliyu, and Sulaiman S. Ibrahim. 2023. "Multiple Pesticide Resistance in Rust-Red Flour Beetle (Tribolium castaneum, Herbst 1797) from Northern Nigeria Is Probably Driven by Metabolic Mechanisms" Agrochemicals 2, no. 2: 170-180. https://doi.org/10.3390/agrochemicals2020012