
Stearylamine Liposome as an Anti-Parasitic Agent


{kind=link}
{kind=link}
Abstract
:1. Introduction
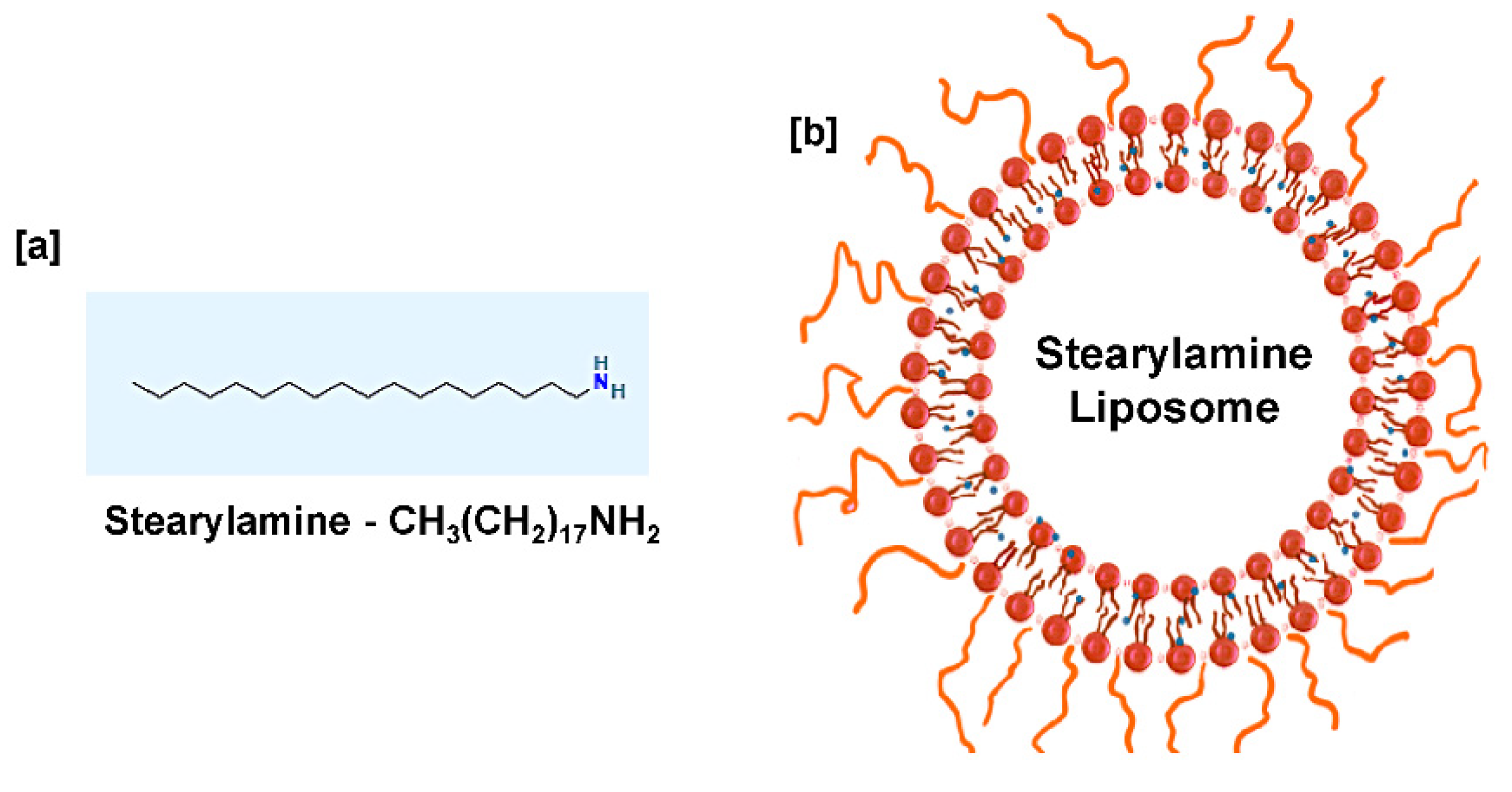
2. Stearylamine Liposome
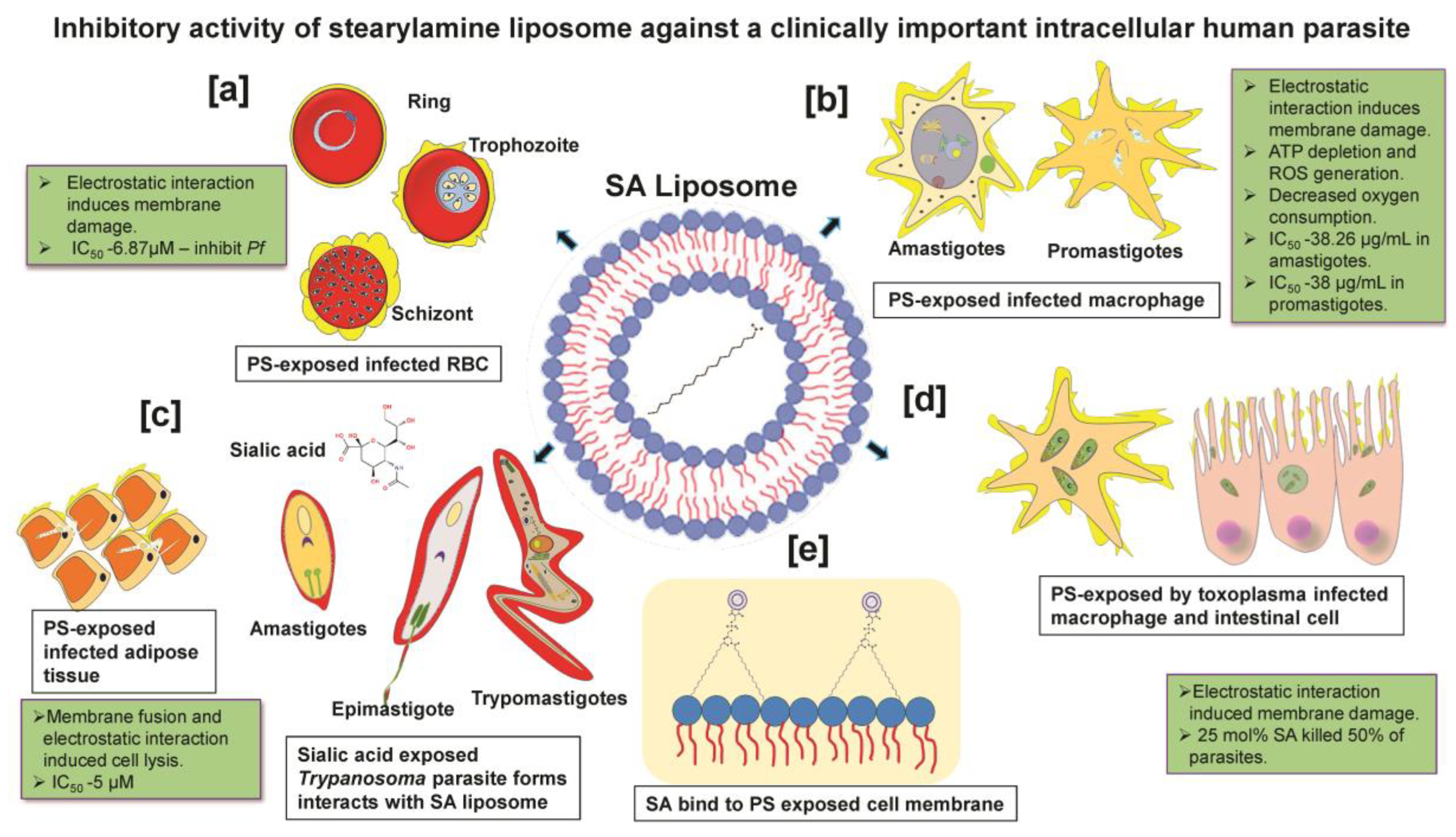
3. Stearylamine Liposome Activity on Plasmodium
4. Stearylamine Liposome Activity on Leishmania
5. Stearylamine Liposome Activity on Trypanosoma
6. Stearylamine Liposome Activity on Toxoplasma
7. Immunomodulatory Effect of SA Liposome
8. Safety and Biodistribution Profile of SA Liposome
9. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. A Global Brief on Vector-Borne Diseases; World Health Organization: Geneva, Switzerland, 2014.
- Gupta, S.; Gazendam, N.; Farina, J.M.; Saldarriaga, C.; Mendoza, I.; López-Santi, R.; Pérez, G.E.; Martínez-Sellés, M.; Baranchuk, A. Malaria and the heart: JACC state-of-the-art review. J. Am. Coll. Cardiol. 2021, 77, 1110–1121. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. World Malaria Report 2021; World Health Organization: Geneva, Switzerland, 2021.
- Bouyou-Akotet, M.K.; Ionete-Collard, D.E.; Mabika-Manfoumbi, M.; Kendjo, E.; Matsiegui, P.-B.; Mavoungou, E.; Kombila, M. Prevalence of Plasmodium falciparum infection in pregnant women in Gabon. Malar. J. 2003, 2, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salanti, A.; Staalsoe, T.; Lavstsen, T.; Jensen, A.T.; Sowa, M.K.; Arnot, D.E.; Hviid, L.; Theander, T.G. Selective upregulation of a single distinctly structured var gene in chondroitin sulphate A-adhering Plasmodium falciparum involved in pregnancy-associated malaria. Mol. Microbiol. 2003, 49, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Indari, O.; Chandramohanadas, R.; Jha, H.C. Epstein–Barr virus infection modulates blood–brain barrier cells and its co-infection with Plasmodium falciparum induces RBC adhesion. Pathog. Dis. 2021, 79, ftaa080. [Google Scholar] [CrossRef]
- Brooker, S.J.; Pullan, R.L.; Gitonga, C.W.; Ashton, R.A.; Kolaczinski, J.H.; Kabatereine, N.B.; Snow, R.W. Plasmodium–helminth coinfection and its sources of heterogeneity across east Africa. J. Infect. Dis. 2012, 205, 841–852. [Google Scholar] [CrossRef] [PubMed]
- Akinbo, F.; Olowookere, T.; Oriakhi, M.O. Co-infection of Plasmodium falciparum and HIV among pregnant women in Edo State, Nigeria. J. Afr. Assoc. Physiol. Sci. 2019, 7, 53–58. [Google Scholar]
- Kamhawi, S. Phlebotomine sand flies and Leishmania parasites: Friends or foes? Trends Parasitol. 2006, 22, 439–445. [Google Scholar] [CrossRef]
- WHO. 2022. Available online: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 15 July 2022).
- Agrawal, Y.; Sinha, A.; Upadhyaya, P.; Kafle, S.; Rijal, S.; Khanal, B. Hematological profile in visceral leishmaniasis. Int. J. Infect. Microbiol. 2013, 2, 39–44. [Google Scholar] [CrossRef]
- Rathnayake, D.; Ranawake, R.R.; Sirimanna, G.; Siriwardhane, Y.; Karunaweera, N.; De Silva, R. Co-infection of mucosal leishmaniasis and extra pulmonary tuberculosis in a patient with inherent immune deficiency. Int. J. Dermatol. 2010, 49, 549–551. [Google Scholar] [CrossRef]
- Gaifer, Z.; Boulassel, M.-R. Leishmania infantum and Epstein-Barr virus co-infection in a patient with hemophagocytosis. Infect. Dis. Rep. 2017, 8, 6545. [Google Scholar] [CrossRef] [Green Version]
- de Lourdes Higuchi, M.; De Brito, T.; Reis, M.M.; Barbosa, A.; Bellotti, G.; Pereira-Barreto, A.C.; Pileggi, F. Correlation between Trypanosoma cruzi parasitism and myocardial inflammatory infiltrate in human chronic chagasic myocarditis: Light microscopy and immunohistochemical findings. Cardiovasc. Pathol. 1993, 2, 101–106. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, R.B.; Troncon, L.E.A.; Dantas, R.O.; Meneghelli, U.G. Gastrointestinal manifestations of Chagas’ disease. Am. J. Gastroenterol. 1998, 93, 884–889. [Google Scholar] [CrossRef] [PubMed]
- WHO. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/trypanosomiasis-human-african-(sleeping-sickness) (accessed on 25 July 2022).
- Cordova, E.; Boschi, A.; Ambrosioni, J.; Cudos, C.; Corti, M. Reactivation of Chagas disease with central nervous system involvement in HIV-infected patients in Argentina, 1992–2007. Int. J. Infect. Dis. 2008, 12, 587–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartori, A.M.C.; Bezerra, R.C.; do S Guilherme, C.; Lopes, M.H.; Shikanai-Yasuda, M.A. Exacerbation of HIV viral load simultaneous with asymptomatic reactivation of chronic Chagas’ disease. Am. J. Trop. Med. Hyg. 2002, 67, 521–523. [Google Scholar] [CrossRef] [Green Version]
- WHO. 2015. Available online: https://www.euro.who.int/__data/assets/pdf_file/0011/294599/Factsheet-Toxoplasmosis (accessed on 10 August 2022).
- Alsammani, M.A.; Ahmed, S.R.; Alsheeha, M.A.; Saadia, Z.; Khairi, S.A.; Research, G. Co-infection with Toxoplasma gondii and Clostridium perfringens in a postpartum woman with uterine gas gangrene: A case report. J. Obstet. Gynaecol. Res. 2012, 38, 1024–1027. [Google Scholar] [CrossRef]
- Reynolds, M.G.; Oh, J.; Roos, D.S. In vitro generation of novel pyrimethamine resistance mutations in the Toxoplasma gondii dihydrofolate reductase. Antimicrob. Agents Chemother. 2001, 45, 1271–1277. [Google Scholar] [CrossRef] [Green Version]
- Mbengue, A.; Bhattacharjee, S.; Pandharkar, T.; Liu, H.; Estiu, G.; Stahelin, R.V.; Rizk, S.S.; Njimoh, D.L.; Ryan, Y.; Chotivanich, K.J.N. A molecular mechanism of artemisinin resistance in Plasmodium falciparum malaria. Nature 2015, 520, 683–687. [Google Scholar] [CrossRef] [Green Version]
- Chinappi, M.; Via, A.; Marcatili, P.; Tramontano, A. On the mechanism of chloroquine resistance in Plasmodium falciparum. PLoS ONE 2010, 5, e14064. [Google Scholar] [CrossRef] [Green Version]
- Purkait, B.; Kumar, A.; Nandi, N.; Sardar, A.H.; Das, S.; Kumar, S.; Pandey, K.; Ravidas, V.; Kumar, M.; De, T.; et al. Mechanism of amphotericin B resistance in clinical isolates of Leishmania donovani. Antimicrob. Agents Chemother. 2012, 56, 1031–1041. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Victoria, J.M.; Pérez-Victoria, F.J.; Parodi-Talice, A.; Jiménez, I.A.; Ravelo, A.G.; Castanys, S.; Gamarro, F. Alkyl-lysophospholipid resistance in multidrug-resistant Leishmania tropica and chemosensitization by a novel P-glycoprotein-like transporter modulator. Antimicrob. Agents Chemother. 2001, 45, 2468–2474. [Google Scholar] [CrossRef] [Green Version]
- Wyllie, S.; Foth, B.J.; Kelner, A.; Sokolova, A.Y.; Berriman, M.; Fairlamb, A.H. Nitroheterocyclic drug resistance mechanisms in Trypanosoma brucei. J. Antimicrob. Chemother. 2016, 71, 625–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, A.C.; Beverley, S.; Cotrim, P.C. Functional genetic identification of PRP1, an ABC transporter superfamily member conferring pentamidine resistance in Leishmania major. Mol. Biochem. Parasitol. 2003, 130, 83–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfefferkorn, E.R.; Borotz, S.E.; Nothnagel, R.F. Mutants of Toxoplasma gondii Resistant to Atovaquone (566C80) or Decoquinate. J. Parasitol. 1993, 79, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Sokolova, A.Y.; Wyllie, S.; Patterson, S.; Oza, S.; Read, K.D.; Fairlamb, A.H. Cross-Resistance to Nitro Drugs and Implications for Treatment of Human African Trypanosomiasis. Antimicrob. Agents Chemother. 2010, 54, 2893–2900. [Google Scholar] [CrossRef] [Green Version]
- Barrett, M.; Fairlamb, A. The Biochemical Basis of Arsenical–Diamidine Crossresistance in African Trypanosomes. Parasitol. Today 1999, 15, 136–140. [Google Scholar] [CrossRef]
- Cowman, A.F.; Galatis, D.; Thompson, J.K. Selection for mefloquine resistance in Plasmodium falciparum is linked to amplification of the pfmdr1 gene and cross-resistance to halofantrine and quinine. Proc. Natl. Acad. Sci. USA 1994, 91, 1143–1147. [Google Scholar] [CrossRef] [Green Version]
- Duraisingh, M.T.; Cowman, A.F. Contribution of the pfmdr1 gene to antimalarial drug-resistance. Acta Trop. 2005, 94, 181–190. [Google Scholar] [CrossRef]
- Aliee, M.; Castaño, S.; Davis, C.N.; Patel, S.; Miaka, E.M.; Spencer, S.E.F.; Keeling, M.J.; Chitnis, N.; Rock, K.S. Predicting the impact of COVID-19 interruptions on transmission of gambiense human African trypanosomiasis in two health zones of the Democratic Republic of Congo. Trans. R. Soc. Trop. Med. Hyg. 2021, 115, 245–252. [Google Scholar] [CrossRef]
- Sebastião, C.S.; Gaston, C.; Paixão, J.P.; Sacomboio, E.N.; Neto, Z.; de Vasconcelos, J.N.; Morais, J. Coinfection between SARS-CoV-2 and vector-borne diseases in Luanda, Angola. J. Med. Virol. 2022, 94, 366–371. [Google Scholar] [CrossRef]
- Mazaherifar, S.; Solhjoo, K.; Abdoli, A. Outbreak of cutaneous leishmaniasis before and during the COVID-19 pandemic in Jahrom, an endemic region in southwest of Iran: Cutaneous leishmaniasis and the COVID-19 pandemic. Emerg. Microbes Infect. 2022, 11, 2218–2221. [Google Scholar] [CrossRef]
- Teixeira, H.; Rosilio, V.; Laigle, A.; Lepault, J.; Erk, I.; Scherman, D.; Benita, S.; Couvreur, P.; Dubernet, C. Characterization of oligonucleotide/lipid interactions in submicron cationic emulsions: Influence of the cationic lipid structure and the presence of PEG-lipids. Biophys. Chem. 2001, 92, 169–181. [Google Scholar] [CrossRef]
- Manjunath, K.; Venkateswarlu, V. Pharmacokinetics, tissue distribution and bioavailability of clozapine solid lipid nanoparticles after intravenous and intraduodenal administration. J. Control. Release 2005, 107, 215–228. [Google Scholar] [CrossRef]
- Yoksan, R.; Chirachanchai, S. Amphiphilic chitosan nanosphere: Studies on formation, toxicity, and guest molecule incorporation. Bioorganic Med. Chem. 2008, 16, 2687–2696. [Google Scholar] [CrossRef] [PubMed]
- Elkholy, N.S.; Shafaa, M.W.; Mohammed, H.S. Biophysical characterization of lutein or beta carotene-loaded cationic liposomes. RSC Adv. 2020, 10, 32409–32422. [Google Scholar] [CrossRef] [PubMed]
- Rosing, J.; Speijer, H.; Zwaal, R.F.A. Prothrombin activation on phospholipid membranes with positive electrostatic potential. Biochemistry 1988, 27, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.; Vorauer-Uhl, K. Liposome technology for industrial purposes. J. Drug Deliv. 2011, 2011, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Lasic, D.D.; Walker, S.; Bode, C.J.; Paquet, K.-J. Kinetic and thermodynamic effects on the structure and formation of phosphatidylcholine vesicles. Hepatology 1991, 13, 1010–1013. [Google Scholar] [CrossRef]
- Ulrich, A.S. Biophysical aspects of using liposomes as delivery vehicles. Biosci. Rep. 2002, 22, 129–150. [Google Scholar] [CrossRef] [PubMed]
- Gogoi, M.; Jaiswal, M.K.; Sarma, H.D.; Bahadur, D.; Banerjee, R. Biocompatibility and therapeutic evaluation of magnetic liposomes designed for self-controlled cancer hyperthermia and chemotherapy. Integr. Biol. 2017, 9, 555–565. [Google Scholar] [CrossRef] [PubMed]
- Téllez, J.; Echeverry, M.C.; Romero, I.; Guatibonza, A.; Ramos, G.S.; De Oliveira, A.C.B.; Frézard, F.; Demicheli, C. Use of liposomal nanoformulations in antileishmania therapy: Challenges and perspectives. J. Liposome Res. 2020, 31, 169–176. [Google Scholar] [CrossRef]
- Yokoyama, S.; Inagaki, A.; Tsuchiya, K.; Sakai, H.; Imura, T.; Ohkubo, T.; Tsubaki, N.; Abe, M. Stearylamine Changes the Liposomal Shape from MLVs to LUVs. J. Oleo Sci. 2005, 54, 251–254. [Google Scholar] [CrossRef] [Green Version]
- Soni, A.; Jain, V.; Jain, S.K.; Khangar, P.K. Preparation and characterization of amphotericin B mannosylated liposomes for effective management of visceral leishmaniasis. J. Drug Deliv. Ther. 2021, 11, 113–118. [Google Scholar] [CrossRef]
- Moosavian, S.A.; Fallah, M.; Jaafari, M.R. The activity of encapsulated meglumine antimoniate in stearylamine-bearing liposomes against cutaneous leishmaniasis in BALB/c mice. Exp. Parasitol. 2019, 200, 30–35. [Google Scholar] [CrossRef] [PubMed]
- De, M.; Ghosh, S.; Sen, T.; Shadab, M.; Banerjee, I.; Basu, S.; Ali, N. A Novel Therapeutic Strategy for Cancer Using Phosphatidylserine Targeting Stearylamine-Bearing Cationic Liposomes. Mol. Ther. Nucleic Acids 2017, 10, 9–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkateswarlu, V.; Manjunath, K. Preparation, characterization and in vitro release kinetics of clozapine solid lipid nanoparticles. J. Control. Release 2004, 95, 627–638. [Google Scholar] [CrossRef]
- Nakanishi, T.; Kunisawa, J.; Hayashi, A.; Tsutsumi, Y.; Kubo, K.; Nakagawa, S.; Fujiwara, H.; Hamaoka, T.; Mayumi, T. Positively Charged Liposome Functions as an Efficient Immunoadjuvant in Inducing Immune Responses to Soluble Proteins. Biochem. Biophys. Res. Commun. 1997, 240, 793–797. [Google Scholar] [CrossRef]
- Lv, H.; Zhang, S.; Wang, B.; Cui, S.; Yan, J. Toxicity of cationic lipids and cationic polymers in gene delivery. J. Control. Release 2006, 114, 100–109. [Google Scholar] [CrossRef]
- Matos, A.P.D.S.; Lopes, D.C.D.X.P.; Peixoto, M.L.H.; Cardoso, V.D.S.; Vermelho, A.B.; Santos-Oliveira, R.; Viçosa, A.L.; Holandino, C.; Ricci-Júnior, E. Development, characterization, and anti-leishmanial activity of topical amphotericin B nanoemulsions. Drug Deliv. Transl. Res. 2020, 10, 1552–1570. [Google Scholar] [CrossRef]
- Yang, Z.; Tian, L.; Liu, J.; Huang, G. Construction and evaluation in vitro and in vivo of tedizolid phosphate loaded cationic liposomes. J. Liposome Res. 2017, 28, 322–330. [Google Scholar] [CrossRef]
- Webb, M.S.; Wheeler, J.J.; Bally, M.B.; Mayer, L.D. The cationic lipid stearylamine reduces the permeability of the cationic drugs verapamil and prochlorperazine to lipid bilayers: Implications for drug delivery. Biochim. Biophys. Acta (BBA) Biomembr. 1995, 1238, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Lotosh, N.Y.; Aliaseva, S.O.; Malashenkova, I.K.; Sorokoumova, G.M.; Vasilov, R.G.; Selischeva, A.A. Cationic Liposomes Cause ROS Generation and Release of Neutrophil Extracellular Traps. Biochem. Suppl. Ser. A Membr. Cell Biol. 2019, 13, 40–49. [Google Scholar] [CrossRef]
- Naderer, T.; Fulcher, M.C. Targeting apoptosis pathways in infections. J. Leukoc. Biol. 2018, 103, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Aramaki, Y.; Takano, S.; Arima, H.; Tsuchiya, S. Induction of apoptosis in WEHI 231 cells by cationic liposomes. Pharm. Res. 2000, 17, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Aramaki, Y.; Takano, S.; Tsuchiya, S. Induction of apoptosis in macrophages by cationic liposomes. FEBS Lett. 1999, 460, 472–476. [Google Scholar] [CrossRef] [Green Version]
- Takano, S.; Aramaki, Y.; Tsuchiya, S. Physicochemical Properties of Liposomes Affecting Apoptosis Induced by Cationic Liposomes in Macrophages. Pharm. Res. 2003, 20, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Takano, S.; Aramaki, Y.; Tsuchiya, S. Lipoxygenase May Be Involved in Cationic Liposome-Induced Macrophage Apoptosis. Biochem. Biophys. Res. Commun. 2001, 288, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Hasan, G.M.; Garg, N.; Dogra, E.; Surolia, R.; Ghosh, P.C. Inhibition of the growth of Plasmodium falciparum in culture by stearylamine-phosphatidylcholine liposomes. J. Parasitol. Res. 2011, 2011, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Dey, T.; Anam, K.; Afrin, F.; Ali, N. Antileishmanial Activities of Stearylamine-Bearing Liposomes. Antimicrob. Agents Chemother. 2000, 44, 1739–1742. [Google Scholar] [CrossRef] [Green Version]
- Tachibana, H.; Yoshihara, E.; Kaneda, Y.; Nakae, T. Protection of Toxoplasma gondii-Infected Mice by Stearylamine-Bearing Liposomes. J. Parasitol. 1990, 76, 352–355. [Google Scholar] [CrossRef]
- Yoshihara, E.; Tachibana, H.; Nakae, T. Trypanocidal activity of the stearylamine-bearing liposome invitro. Life Sci. 1987, 40, 2153–2159. [Google Scholar] [CrossRef]
- Sinha, R.; Roychoudhury, J.; Palit, P.; Ali, N. Cationic liposomal sodium stibogluconate (SSG), a potent therapeutic tool for treatment of infection by SSG-sensitive and-resistant Leishmania donovani. Antimicrob. Agents Chemother. 2015, 59, 344–355. [Google Scholar] [CrossRef] [Green Version]
- Antila, H.S.; Buslaev, P.; Favela-Rosales, F.; Ferreira, T.M.; Gushchin, I.; Javanainen, M.; Kav, B.; Madsen, J.J.; Melcr, J.; Miettinen, M.S.; et al. Headgroup Structure and Cation Binding in Phosphatidylserine Lipid Bilayers. J. Phys. Chem. B 2019, 123, 9066–9079. [Google Scholar] [CrossRef] [PubMed]
- Kay, J.G.; Grinstein, S. Sensing Phosphatidylserine in Cellular Membranes. Sensors 2011, 11, 1744–1755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariño, G.; Kroemer, G. Mechanisms of apoptotic phosphatidylserine exposure. Cell Res. 2013, 23, 1247–1248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadok, V.A.; de Cathelineau, A.; Daleke, D.L.; Henson, P.M.; Bratton, D.L. Loss of Phospholipid Asymmetry and Surface Exposure of Phosphatidylserine Is Required for Phagocytosis of Apoptotic Cells by Macrophages and Fibroblasts. J. Biol. Chem. 2001, 276, 1071–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeung, T.; Gilbert, G.E.; Shi, J.; Silvius, J.; Kapus, A.; Grinstein, S. Membrane Phosphatidylserine Regulates Surface Charge and Protein Localization. Science 2008, 319, 210–213. [Google Scholar] [CrossRef]
- Zhao, H.; Tuominen, E.K.J.; Kinnunen, P.K.J. Formation of Amyloid Fibers Triggered by Phosphatidylserine-Containing Membranes. Biochemistry 2004, 43, 10302–10307. [Google Scholar] [CrossRef]
- Yoshihara, E.; Nakae, T. Cytolytic activity of liposomes containing stearylamine. Biochim. Biophys. Acta (BBA)-Biomembr. 1986, 854, 93–101. [Google Scholar] [CrossRef]
- Banerjee, A.; De, M.; Ali, N. Complete Cure of Experimental Visceral Leishmaniasis with Amphotericin B in Stearylamine-Bearing Cationic Liposomes Involves Down-Regulation of IL-10 and Favorable T Cell Responses. J. Immunol. 2008, 181, 1386–1398. [Google Scholar] [CrossRef] [Green Version]
- Watarai, S.; Sasaki, Y. Evaluation of stearylamine-modified liposomes for the oral vaccine adjuvant. J. Infect. Dis. Ther. 2014, 2, 1–6. [Google Scholar]
- Sharma, S.; Rajendran, V.; Kulshreshtha, R.; Ghosh, P.C. Enhanced efficacy of anti-miR-191 delivery through stearylamine liposome formulation for the treatment of breast cancer cells. Int. J. Pharm. 2017, 530, 387–400. [Google Scholar] [CrossRef] [PubMed]
- Tahara, K.; Kobayashi, M.; Yoshida, S.; Onodera, R.; Inoue, N.; Takeuchi, H. Effects of cationic liposomes with stearylamine against virus infection. Int. J. Pharm. 2018, 543, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Antinori, S.; Galimberti, L.; Milazzo, L.; Corbellino, M. Biology of human malaria plasmodia including plasmodium knowlesi. Mediterr. J. Hematol. Infect. Dis. 2012, 4, e2012013. [Google Scholar] [CrossRef] [Green Version]
- Gilson, P.R.; Crabb, B.S. Morphology and kinetics of the three distinct phases of red blood cell invasion by Plasmodium falciparum merozoites. Int. J. Parasitol. 2009, 39, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, M.; Yamodo, I.; Ranjan, R.; Li, X.; Mines, G.; Marinkovic, M.; Hanada, T.; Oh, S.S.; Chishti, A.H. Human erythrocyte band 3 functions as a receptor for the sialic acid-independent invasion of Plasmodium falciparum. Role of the RhopH3–MSP1 complex. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2014, 1843, 2855–2870. [Google Scholar] [CrossRef] [Green Version]
- Orlandi, P.A.; Klotz, F.W.; Haynes, J.D. A malaria invasion receptor, the 175-kilodalton erythrocyte binding antigen of Plasmodium falciparum recognizes the terminal Neu5Ac (alpha 2-3) Gal-sequences of glycophorin A. J. Cell Biol. 1992, 116, 901–909. [Google Scholar] [CrossRef] [Green Version]
- Sim, B.K.L.; Chitnis, C.E.; Wasniowska, K.; Hadley, T.J.; Miller, L.H. Receptor and Ligand Domains for Invasion of Erythrocytes by Plasmodium falciparum. Science 1994, 264, 1941–1944. [Google Scholar] [CrossRef]
- Duraisingh, M.T.; Maier, A.G.; Triglia, T.; Cowman, A.F. Erythrocyte-binding antigen 175 mediates invasion in Plasmodium falciparum utilizing sialic acid-dependent and -independent pathways. Proc. Natl. Acad. Sci. USA 2003, 100, 4796–4801. [Google Scholar] [CrossRef] [Green Version]
- Gilberger, T.-W.; Thompson, J.K.; Triglia, T.; Good, R.T.; Duraisingh, M.T.; Cowman, A.F. A Novel Erythrocyte Binding Antigen-175 Paralogue fromPlasmodium falciparum Defines a New Trypsin-resistant Receptor on Human Erythrocytes. J. Biol. Chem. 2003, 278, 14480–14486. [Google Scholar] [CrossRef] [Green Version]
- Mayer, D.C.G.; Cofie, J.; Jiang, L.; Hartl, D.L.; Tracy, E.; Kabat, J.; Mendoza, L.H.; Miller, L.H. Glycophorin B is the erythrocyte receptor of Plasmodium falciparum erythrocyte-binding ligand, EBL-1. Proc. Natl. Acad. Sci. USA 2009, 106, 5348–5352. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Marinkovic, M.; Russo, C.; McKnight, C.J.; Coetzer, T.L.; Chishti, A.H. Identification of a specific region of Plasmodium falciparum EBL-1 that binds to host receptor glycophorin B and inhibits merozoite invasion in human red blood cells. Mol. Biochem. Parasitol. 2012, 183, 23–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobo, C.-A.; Rodriguez, M.; Reid, M.; Lustigman, S. Glycophorin C is the receptor for the Plasmodium falciparum erythrocyte binding ligand PfEBP-2 (baebl). Blood 2003, 101, 4628–4631. [Google Scholar] [CrossRef] [PubMed]
- Mayer, D.C.G.; Kaneko, O.; Hudson-Taylor, D.E.; Reid, M.E.; Miller, L.H. Characterization of a Plasmodium falciparum erythrocyte-binding protein paralogous to EBA-175. Proc. Natl. Acad. Sci. USA 2001, 98, 5222–5227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narum, D.L.; Fuhrmann, S.R.; Luu, T.; Sim, B.K.L. A novel Plasmodium falciparum erythrocyte binding protein-2 (EBP2/BAEBL) involved in erythrocyte receptor binding. Mol. Biochem. Parasitol. 2002, 119, 159–168. [Google Scholar] [CrossRef]
- Jaskiewicz, E.; Jodłowska, M.; Kaczmarek, R.; Zerka, A. Erythrocyte glycophorins as receptors for Plasmodium merozoites. Parasites Vectors 2019, 12, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, V.; Rohra, S.; Raza, M.; Hasan, G.M.; Dutt, S.; Ghosh, P.C. Stearylamine Liposomal Delivery of Monensin in Combination with Free Artemisinin Eliminates Blood Stages of Plasmodium falciparum in Culture and P. berghei Infection in Murine Malaria. Antimicrob. Agents Chemother. 2016, 60, 1304–1318. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Arias, C.; Rivera-Correa, J.; Gallego-Delgado, J.; Rudlaff, R.; Fernandez, C.; Roussel, C.; Götz, A.; Gonzalez, S.; Mohanty, A.; Mohanty, S.; et al. Anti-Self Phosphatidylserine Antibodies Recognize Uninfected Erythrocytes Promoting Malarial Anemia. Cell Host Microbe 2016, 19, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Eda, S.; Sherman, I.W. Cytoadherence of malaria-infected red blood cells involves exposure of phosphatidylserine. Cell. Physiol. Biochem. 2002, 12, 373–384. [Google Scholar] [CrossRef]
- Rajendran, V.; Singh, C.; Ghosh, P.C. Improved efficacy of doxycycline in liposomes against Plasmodium falciparum in culture and Plasmodium berghei infection in mice. Can. J. Physiol. Pharmacol. 2018, 96, 1145–1152. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, M.P.A.P.C.G.V.; Pachauri, M.; Ghosh, P.C. Combinatorial Effects of Monensin in Liposome Formulations with Antimalarial Drugs Against Blood Stages of Plasmodium falciparum in Culture and P. berghei Infection. Curr. Drug Ther. 2018, 13, 74–82. [Google Scholar] [CrossRef]
- Baruah, U.K.; Gowthamarajan, K.; Ravisankar, V.; Karri, V.V.S.R.; Simhadri, P.K.; Singh, V. Optimisation of chloroquine phosphate loaded nanostructured lipid carriers using Box–Behnken design and its antimalarial efficacy. J. Drug Target. 2017, 26, 576–591. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S.B.; Das, M.; Sudhandiran, G.; Shaha, C. Increase in Cytosolic Ca2+ Levels through the Activation of Non-selective Cation Channels Induced by Oxidative Stress Causes Mitochondrial Depolarization Leading to Apoptosis-like Death in Leishmania donovaniPromastigotes. J. Biol. Chem. 2002, 277, 24717–24727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, R.; Ghoshal, A.; Mandal, C.; Shaha, C. Leishmania cell surface prohibitin: Role in host–parasite interaction. Cell. Microbiol. 2010, 12, 432–452. [Google Scholar] [CrossRef]
- Rabhi, I.; Rabhi, S.; Ben-Othman, R.; Rasche, A.; Consortium, S.; Daskalaki, A.; Trentin, B.; Piquemal, D.; Regnault, B.; Descoteaux, A.; et al. Transcriptomic Signature of Leishmania Infected Mice Macrophages: A Metabolic Point of View. PLoS Negl. Trop. Dis. 2012, 6, e1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortéa, J.O.Y.; de La Llave, E.; Regnault, B.; Coppée, J.-Y.; Milon, G.; Lang, T.; Prina, E. Transcriptional signatures of BALB/c mouse macrophages housing multiplying Leishmania amazonensis amastigotes. BMC Genom. 2009, 10, 119. [Google Scholar] [CrossRef] [Green Version]
- Frame, M.J.; Mottram, J.C.; Coombs, G.H. Analysis of the roles of cysteine proteinases of Leishmania mexicana in the host–parasite interaction. Parasitology 2000, 121, 367–377. [Google Scholar] [CrossRef]
- Banerjee, A.; Roychoudhury, J.; Ali, N. Stearylamine-bearing cationic liposomes kill Leishmania parasites through surface exposed negatively charged phosphatidylserine. J. Antimicrob. Chemother. 2007, 61, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, A.; De, M.; Ali, N. Combination Therapy with Paromomycin-Associated Stearylamine-Bearing Liposomes Cures Experimental Visceral Leishmaniasis through Th1-Biased Immunomodulation. Antimicrob. Agents Chemother. 2011, 55, 1661–1670. [Google Scholar] [CrossRef] [Green Version]
- Pal, S.; Ravindran, R.; Ali, N. Combination Therapy Using Sodium Antimony Gluconate in Stearylamine-Bearing Liposomes against Established and Chronic Leishmania donovani Infection in BALB/c Mice. Antimicrob. Agents Chemother. 2004, 48, 3591–3593. [Google Scholar] [CrossRef] [Green Version]
- Afrin, F.; Dey, T.; Anam, K.; Ali, N. Leishmanicidal activity of stearylamine-bearing liposomes in vitro. J. Parasitol. 2001, 87, 188–193. [Google Scholar] [CrossRef]
- Roychoudhury, J.; Sinha, R.; Ali, N. Therapy with Sodium Stibogluconate in Stearylamine-Bearing Liposomes Confers Cure against SSG-Resistant Leishmania donovani in BALB/c Mice. PLoS ONE 2011, 6, e17376. [Google Scholar] [CrossRef] [Green Version]
- Rathore, A.; Jain, A.; Gulbake, A.; Shilpi, S.; Khare, P.; Jain, A.; Jain, S.K. Mannosylated liposomes bearing Amphotericin B for effective management of visceral Leishmaniasis. J. Liposome Res. 2011, 21, 333–340. [Google Scholar] [CrossRef]
- Das, A.; Kamran, M.; Ali, N. HO-3867 Induces ROS-Dependent Stress Response and Apoptotic Cell Death in Leishmania donovani. Front. Cell. Infect. Microbiol. 2021, 11, 1216. [Google Scholar] [CrossRef] [PubMed]
- Patere, S.N.; Pathak, P.O.; Shukla, A.K.; Singh, R.K.; Dubey, V.K.; Mehta, M.J.; Patil, A.G.; Gota, V.; Nagarsenker, M.S. Surface-Modified Liposomal Formulation of Amphotericin B: In vitro Evaluation of Potential Against Visceral Leishmaniasis. AAPS PharmSciTech 2016, 18, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Schwendener, R.; Lagocki, P.; Rahman, Y. The effects of charge and size on the interaction of unilamellar liposomes with macrophages. Biochim. Biophys. Acta (BBA) Biomembr. 1984, 772, 93–101. [Google Scholar] [CrossRef]
- Herman, M.; Pérez-Morga, D.; Schtickzelle, N.; Michels, P.A. Turnover of glycosomes during life-cycle differentiation ofTrypanosoma brucei. Autophagy 2008, 4, 294–308. [Google Scholar] [CrossRef] [Green Version]
- Castillo, C.; Carrillo, I.; Libisch, G.; Juiz, N.; Schijman, A.; Robello, C.; Kemmerling, U. Host-parasite interaction: Changes in human placental gene expression induced by Trypanosoma cruzi. Parasites Vectors 2018, 11, 1–13. [Google Scholar] [CrossRef]
- Souto-Padron, T.; De Carvalho, T.; Chiari, E.; De Souza, W. Further studies on the cell surface charge of Trypanosoma cruzi. Acta Trop. 1984, 41, 215–225. [Google Scholar]
- De Carvalho, T.U.; Souto-Padróon, T.; De Souza, W. Trypanosoma cruzi: Surface charge and freeze-fracture of amastigotes. Exp. Parasitol. 1985, 59, 12–23. [Google Scholar] [CrossRef]
- Souza, W.D.; Arguello, C.; Martinez-Palomo, A.; Trissl, D.; Gonzáles-Robles, A.; Chiari, E. Surface charge of Trypanosoma cruzi. Binding of cationized ferritin and measurement of cellular electrophoretic mobility. J. Protozool. 1977, 24, 411–415. [Google Scholar] [CrossRef]
- Tachibana, H.; Yoshihara, E.; Kaneda, Y.; Nakae, T. In vitro lysis of the bloodstream forms of Trypanosoma brucei gambiense by stearylamine-bearing liposomes. Antimicrob. Agents Chemother. 1988, 32, 966–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yongsheng, Y.; Yongchun, O.; Chengmai, R.; Yuanguo, C.; Fenqin, Z. Trypanocidal value of liposomal diminazene in experimental Trypanosoma brucei evansi infection in mice. Veter- Parasitol. 1996, 61, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.; Dubey, J. Toxoplasma gondii: Transmission, diagnosis and prevention. Clin. Microbiol. Infect. 2002, 8, 634–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, D. Use of molecular and ultrastructural markers to evaluate stage conversion of Toxoplasma gondii in both the intermediate and definitive host. Int. J. Parasitol. 2004, 34, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J. Advances in the life cycle of Toxoplasma gondii. Int. J. Parasitol. 1998, 28, 1019–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asthana, S.; Jaiswal, A.K.; Gupta, P.K.; Dube, A.; Chourasia, M.K. Th-1 biased immunomodulation and synergistic antileishmanial activity of stable cationic lipid–polymer hybrid nanoparticle: Biodistribution and toxicity assessment of encapsulated amphotericin B. Eur. J. Pharm. Biopharm. 2015, 89, 62–73. [Google Scholar] [CrossRef]
- De, M.; Ghosh, S.; Asad, M.; Banerjee, I.; Ali, N. Combining doxorubicin with stearylamine-bearing liposomes elicits Th1 cytokine responses and cures metastasis in a mouse model. Cancer Immunol. Immunother. 2020, 69, 1725–1735. [Google Scholar] [CrossRef]
- Pandita, D.; Ahuja, A.; Lather, V.; Dutta, T.; Velpandian, T.; Khar, R.K. Development, characterization and in vitro assessement of stearylamine-based lipid nanoparticles of paclitaxel. Die Pharm. 2011, 66, 171–177. [Google Scholar]
- Gregoriadis, G.; Neerunjun, D.E. Control of the rate of hepatic uptake and catabolism of liposome-entrapped proteins injected into rats. Possible therapeutic applications. Eur. J. Biochem. 1974, 47, 179–185. [Google Scholar] [CrossRef]
- Levchenko, T.S.; Rammohan, R.; Lukyanov, A.N.; Whiteman, K.R.; Torchilin, V.P. Liposome clearance in mice: The effect of a separate and combined presence of surface charge and polymer coating. Int. J. Pharm. 2002, 240, 95–102. [Google Scholar] [CrossRef]
- He, K.; Liu, J.; Gao, Y.; Hao, Y.; Yang, X.; Huang, G. Preparation and Evaluation of Stearylamine-Bearing Pemetrexed Disodium-Loaded Cationic Liposomes In Vitro and In Vivo. AAPS PharmSciTech 2020, 21, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kala, S.G.; Chinni, S. Bioavailability enhancement of vitamin E TPGS liposomes of nintedanib esylate: Formulation optimization, cytotoxicity and pharmacokinetic studies. Drug Deliv. Transl. Res. 2022, 12, 2856–2864. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vassoudevane, J.; Mariebernard, M.; Rajendran, V. Stearylamine Liposome as an Anti-Parasitic Agent. Drugs Drug Candidates 2023, 2, 95-108. https://doi.org/10.3390/ddc2010006
Vassoudevane J, Mariebernard M, Rajendran V. Stearylamine Liposome as an Anti-Parasitic Agent. Drugs and Drug Candidates. 2023; 2(1):95-108. https://doi.org/10.3390/ddc2010006
Chicago/Turabian StyleVassoudevane, Janani, Monika Mariebernard, and Vinoth Rajendran. 2023. "Stearylamine Liposome as an Anti-Parasitic Agent" Drugs and Drug Candidates 2, no. 1: 95-108. https://doi.org/10.3390/ddc2010006