
Crossiella, a Rare Actinomycetota Genus, Abundant in the Environment

 , , , and
, , , and
Abstract
:1. Introduction
2. The Genus Crossiella in Caves
3. Crossiella in the Environment
4. Crossiella Isolates
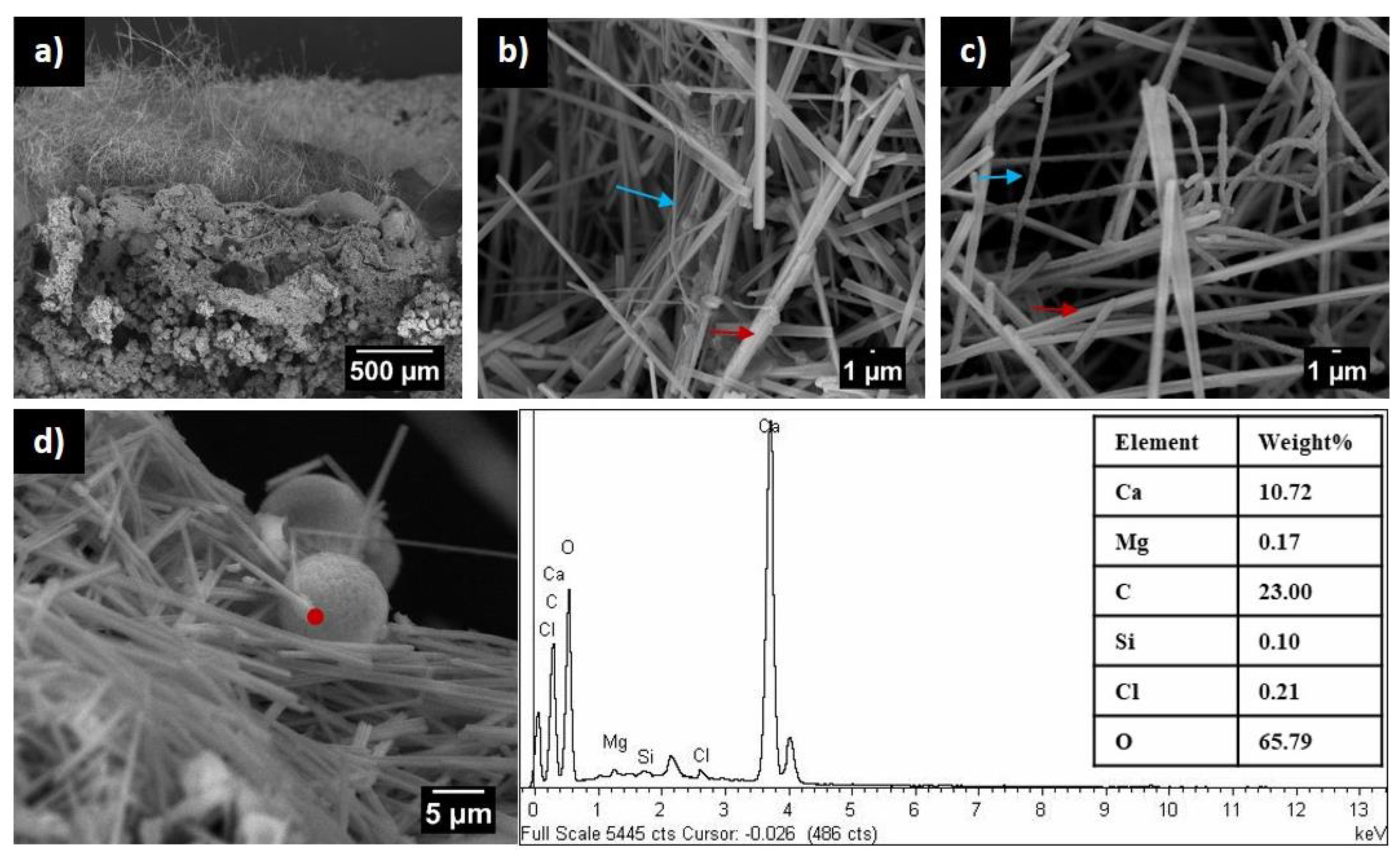
5. Biomineralization in Caves Induced by Crossiella
6. Moonmilk Formation
7. Is Moonmilk a Source of Bioactive Compounds?
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takahashi, A.; Hotta, K.; Saito, N.; Morioka, M.; Okami, Y.; Umezawa, H. Production of novel antibiotic, dopsisamine, by a new subspecies of Nocardiopsis mutabilis with multiple antibiotic resistance. J. Antibiot. 1986, 39, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Labeda, D.P.; Lechevalier, M.P. Amendment of the genus Saccharothrix Labeda et al. 1984 and descriptions of Saccharothrix espanaensis sp. nov., Saccharothrix cryophilis sp. nov., and Saccharothrix mutabilis comb. nov. Int. J. Syst. Bacteriol. 1989, 39, 420–423. [Google Scholar] [CrossRef]
- Labeda, D.P. Crossiella gen. nov., a new genus related to Streptoalloteichus. Int. J. Syst. Evol. Microbiol. 2001, 51, 575–579. [Google Scholar] [CrossRef]
- Donahue, J.M.; Williams, N.M.; Sells, S.F.; Labeda, D.P. Crossiella equi sp. nov., isolated from equine placentas. Int. J. Syst. Evol. Microbiol. 2002, 52, 2169–2173. [Google Scholar]
- Sánchez-Moral, S. Estudio Integral del Estado de Conservación de la Cueva de Altamira y su Arte Paleolítico (2007–2009). Perspectivas Futuras de Conservación; Monografías N° 24; Museo Nacional y Centro de Investigación de Altamira: Santillana del Mar, Spain, 2014. [Google Scholar]
- Adeyemo, O.M.; Onilude, A.A. Antimicrobial potential of a rare actinomycete isolated from soil: Crossiella sp.-EK18. J. Adv. Microbiol. 2018, 11, 1–15. [Google Scholar] [CrossRef]
- Cimermanova, M.; Pristas, P.; Piknova, M. Biodiversity of actinomycetes from heavy metal contaminated technosols. Microorganisms 2021, 9, 1635. [Google Scholar] [CrossRef]
- González-Riancho Fernández, C. Análisis Descriptivo y Funcional de Las Colonias Microbianas Visibles Que Crecen en la Cueva de Altamira, Enfocado al Diseño de Medidas de Control. Ph.D. Thesis, Universidad de Cantabria, Santander, Spain, 2021. [Google Scholar]
- Gonzalez-Pimentel, J.L.; Dominguez-Moñino, I.; Jurado, V.; Laiz, L.; Caldeira, A.T.; Saiz-Jimenez, C. The rare actinobacterium Crossiella sp. is a potential source of new bioactive compounds with activity against bacteria and fungi. Microorganisms 2022, 10, 1575. [Google Scholar] [CrossRef]
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol. 2021, 71, 005056. [Google Scholar] [CrossRef] [PubMed]
- Berdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef]
- Subramani, R.; Aalbersberg, W. Culturable rare Actinomycetes: Diversity, isolation and marine natural product discovery. Appl. Microbiol. Biotechnol. 2013, 97, 9291–9321. [Google Scholar] [CrossRef]
- Tiwari, K.; Gupta, R.K. Rare actinomycetes: A potential storehouse for novel antibiotics. Crit. Rev. Biotechnol. 2012, 32, 108–132. [Google Scholar] [CrossRef]
- Tiwari, K.; Gupta, R.K. Diversity and isolation of rare actinomycetes: An overview. Crit. Rev. Microbiol. 2013, 39, 256–294. [Google Scholar] [CrossRef] [PubMed]
- Seong, C.N.; Choi, J.H.; Baik, K.-S. An improved selective isolation of rare actinomycetes from forest soil. J. Microbiol. 2001, 39, 17–23. [Google Scholar]
- Bredholdt, H.; Galatenko, O.A.; Engelhardt, K.; Fjærvik, E.; Terekhova, L.P.; Zotchev, S.B. Rare actinomycete bacteria from the shallow water sediments of the Trondheim Fjord, Norway: Isolation, diversity and biological activity. Environ. Microbiol. 2007, 9, 2756–2764. [Google Scholar] [CrossRef]
- Fang, B.-Z.; Salam, N.; Han, M.-X.; Jiao, J.-Y.; Cheng, J.; Wei, D.-Q.; Xiao, M.; Li, W.-J. Insights on the effects of heat pretreatment, pH, and calcium salts on isolation of rare actinobacteria from karstic caves. Front. Microbiol. 2017, 8, 1535. [Google Scholar] [CrossRef] [PubMed]
- Goodfellow, M.; Nouioui, I.; Sanderson, R.; Xie, F.; Bull, A.T. Rare taxa and dark microbial matter: Novel bioactive actinobacteria abound in Atacama Desert soils. Anton. Leeuw. 2018, 111, 1315–1332. [Google Scholar] [CrossRef]
- Benhadj, M.; Gacemi-Kirane, D.; Menasria, T.; Guebla, K.; Ahmane, Z. Screening of rare actinomycetes isolated from natural wetland ecosystem (Fetzara Lake, northeastern Algeria) for hydrolytic enzymes and antimicrobial activities. J. King Saud Univ. Sci. 2019, 31, 706–712. [Google Scholar] [CrossRef]
- Zamora-Quintero, A.Y.; Torres-Beltrán, M.; Guillén Matus, D.G.; Oroz-Parra, I.; Millán-Aguiñaga, N. Rare actinobacteria isolated from the hypersaline Ojo de Liebre Lagoon as a source of novel bioactive compounds with biotechnological potential. Microbiology 2022, 168, 001144. [Google Scholar] [CrossRef]
- Tiwari, K.; Upadhyay, D.J.; Mösker, E.; Süssmuth, R.; Gupta, R.K. Culturable bioactive actinomycetes from the Great Indian Thar Desert. Ann. Microbiol. 2015, 65, 1901–1914. [Google Scholar] [CrossRef]
- Mohammadipanah, F.; Wink, J. Actinobacteria from arid and desert habitats: Diversity and biological activity. Front. Microbiol. 2016, 6, 1541. [Google Scholar] [CrossRef]
- Gacem, M.A.; Ould-El-Hadj-Kheli, A.; Abd-Elsalam, K.A.; Wink, J. Actinobacteria in the Algerian Sahara: Diversity, adaptation mechanism and special unexploited biotopes for the isolation of novel rare taxa. Biologia 2001, 76, 3787–3799. [Google Scholar] [CrossRef]
- Hui, M.L.-Y.; Tan, L.T.-H.; Letchumanan, V.; He, Y.-W.; Fang, C.-M.; Chan, K.-G.; Law, J.W.-F.; Lee, L.-H. The extremophilic Actinobacteria: From microbes to medicine. Antibiotics 2021, 10, 682. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Pimentel, J.L.; Martin-Pozas, T.; Jurado, V.; Miller, A.Z.; Caldeira, A.T.; Fernandez-Lorenzo, O.; Sanchez-Moral, S.; Saiz-Jimenez, C. Prokaryotic communities from a lava tube cave in La Palma Island (Spain) are involved in the biogeochemical cycle of major elements. PeerJ 2021, 9, e11386. [Google Scholar] [CrossRef] [PubMed]
- Martin-Pozas, T.; Cuezva, S.; Fernandez-Cortes, A.; Cañaveras, J.C.; Benavente, D.; Jurado, V.; Saiz-Jimenez, C.; Janssens, I.; Seijas, N.; Sanchez-Moral, S. Role of subterranean microbiota in the carbon cycle and greenhouse gas dynamics. Sci. Total Environ. 2022, 831, 154921. [Google Scholar] [CrossRef]
- Cuezva, S.; Martin-Pozas, T.; Fernandez-Cortes, A.; Cañaveras, J.C.; Janssens, I.; Sanchez-Moral, S. On the role of cave-soil in the carbon cycle. A first approach. EGU Gen. Assem. 2020. Available online: https://presentations.copernicus.org/EGU2020/EGU2020-21793_presentation.pdf. (accessed on 22 April 2022).
- Martin-Pozas, T.; Fernandez-Cortes, A.; Cuezva, S.; Cañaveras, J.C.; Benavente, D.; Duarte, E.; Saiz-Jimenez, C.; Sanchez-Moral, S. New insights into the structure, microbial diversity and ecology of yellow biofilms in a Paleolithic rock art cave (Pindal Cave, Asturias, Spain). Sci. Total Environ. 2023, 882. in press. [Google Scholar]
- González-Pimentel, J.L. Microorganismos de las Cuevas Volcánicas de La Palma (Islas Canarias). Diversidad y Potencial Uso Biotecnológico. Ph.D. Thesis, Universidad Pablo Olavide, Sevilla, Spain, 2019. [Google Scholar]
- Jurado, V.; del Rosal, Y.; Gonzalez-Pimentel, J.L.; Hermosin, B.; Saiz-Jimenez, C. Biological control of phototrophic biofilms in a show cave: The case of Nerja Cave. Appl. Sci. 2020, 10, 3448. [Google Scholar] [CrossRef]
- Martin-Pozas, T. Papel de los Microorganismos en Procesos de Captación y Emisión de Gases de Efecto Invernadero en Ambientes Subterráneos. Ph.D. Thesis, Universidad Complutense de Madrid, Madrid, Spain, 2023. [Google Scholar]
- Sánchez-Moral, S.; Martín-Pozas, T.; Seijas Morales, N.; Fernández-Cortés, A.; Benavente García, D.; Cañaveras Jiménez, J.C.; Cuezva, S. Instalación de Sensores en Sótano Arqueológico del Museo Carmen Thyssen de Málaga Para la Toma de Datos, el Análisis y Adopción de Medidas Correctoras del Deterioro Del Recinto; Unpublished Report; MNCN: Madrid, Spain, 2021. [Google Scholar]
- Stomeo, F.; Portillo, M.C.; Gonzalez, J.M.; Laiz, L.; Saiz-Jimenez, C. Pseudonocardia in white colonizations in two caves with Paleolithic paintings. Int. Biodeter. Biodegr. 2008, 62, 483–486. [Google Scholar] [CrossRef]
- Portillo, M.C.; Gonzalez, J.M. Microbial community diversity and the complexity of preserving cultural heritage. In Biocolonization of Stone: Control and Preventive Methods; Charola, A.E., McNamara, C., Koestler, R.J., Eds.; Smithsonian Institution, Scholarly Press: Washington, DC, USA, 2011; pp. 19–28. [Google Scholar]
- Yun, Y.; Wang, H.; Man, B.; Xiang, X.; Zhou, J.; Qiu, X.; Duan, Y.; Engel, A.S. The relationship between ph and bacterial communities in a single karst ecosystem and its implication for soil acidification. Front. Microbiol. 2016, 7, 1955. [Google Scholar] [CrossRef] [PubMed]
- Lepinay, C.; Mihajlovski, A.; Seyer, D.; Touron, S.; Bousta, F.; Di Martino, P. Biofilm communities survey at the areas of salt crystallization on the walls of a decorated shelter listed at UNESCO World cultural Heritage. Int. Biodeter. Biodegr. 2017, 122, 116–127. [Google Scholar] [CrossRef]
- Lepinay, C.; Mihajlovski, A.; Touron, S.; Seyer, D.; Bousta, F.; Di Martino, P. Bacterial diversity associated with saline efflorescences damaging the walls of a French decorated prehistoric cave registered as a World Cultural Heritage Site. Int. Biodeter. Biodegr. 2018, 130, 55–64. [Google Scholar] [CrossRef]
- Alonso, L.; Pommier, T.; Kaufmann, B.; Dubost, A.; Chapulliot, D.; Doré, J.; Douady, C.J.; Moënne-Loccoz, Y. Anthropization level of Lascaux Cave microbiome shown by regional-scale comparisons of pristine and anthropized caves. Mol. Ecol. 2019, 28, 3383–3394. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Fang, C.; Kawasaki, S.; Huang, M.; Achal, V. Bio-consolidation of cracks in masonry cement mortars by Acinetobacter sp. SC4 isolated from a karst cave. Int. Biodeter. Biodegr. 2019, 141, 94–100. [Google Scholar] [CrossRef]
- Long, Y.; Jiang, J.; Hu, X.; Zhou, J.; Hu, J.; Zhou, S. Actinobacterial community in Shuanghe Cave using culture-dependent and -independent approaches. World J. Microbiol. Biotechnol. 2019, 35, 153. [Google Scholar] [CrossRef]
- Wiseschart, A.; Mhuantong, W.; Tangphatsornruang, S.; Chantasingh, D.; Pootanakit, K. Shotgun metagenomic sequencing from Manao-Pee cave, Thailand, reveals insight into the microbial community structure and its metabolic potential. BMC Microbiol. 2019, 19, 144. [Google Scholar] [CrossRef] [PubMed]
- Frazier, V.E. Carbon Metabolism in Cave Subaerial Biofilms. Master’s Thesis, University of Tennessee, Knoxville, TN, USA, 2020. [Google Scholar]
- He, D.; Wu, F.; Ma, W.; Zhang, Y.; Gu, J.-D.; Duan, Y.; Xu, R.; Feng, H.; Wang, W.; Li, S.-W. Insights into the bacterial and fungal communities and microbiome that causes a microbe outbreak on ancient wall paintings in the Maijishan Grottoes. Int. Biodeter. Biodegr. 2021, 163, 105250. [Google Scholar] [CrossRef]
- Ma, L.; Huang, X.; Wang, H.; Yun, Y.; Cheng, X.; Liu, D.; Lu, X.; Qiu, X. Microbial interactions drive distinct taxonomic and potential metabolic responses to habitats in karst cave ecosystem. Microbiol. Spect. 2021, 9, e01152-21. [Google Scholar] [CrossRef]
- Zada, S.; Xie, J.; Yang, M.; Yang, X.; Sajjad, W.; Rafiq, M.; Hasan, F.; Hu, Z.; Wang, H. Composition and functional profiles of microbial communities in two geochemically and mineralogically different caves. Appl. Microbiol. Biotechnol. 2021, 105, 8921–8936. [Google Scholar] [CrossRef]
- Ai, J.; Guo, J.; Li, Y.; Zhong, X.; Lv, Y.; Li, J.; Yang, A. The diversity of microbes and prediction of their functions in karst caves under the influence of human tourism activities—A case study of Zhijin Cave in Southwest China. Environ. Sci. Pollut. Res. 2022, 29, 25858–25868. [Google Scholar] [CrossRef]
- Buresova-Faitova, A.; Kopecky, J.; Sagova-Mareckova, M.; Alonso, L.; Vautrin, F.; Moënne-Loccoz, Y.; Rodriguez-Nava, V. Comparison of Actinobacteria communities from human-impacted and pristine karst caves. MicrobiologyOpen 2022, 11, e1276. [Google Scholar] [CrossRef]
- Djebaili, R.; Mignini, A.; Vaccarelli, I.; Pellegrini, M.; Spera, D.M.; Del Gallo, M.; D’Alessandro, A.M. Polyhydroxybutyrate-producing cyanobacteria from lampenflora: The case study of the “Stiffe” caves in Italy. Front. Microbiol. 2022, 13, 933398. [Google Scholar] [CrossRef]
- Cheng, X.; Xiang, X.; Yun, Y.; Wang, W.; Wang, H.; Bodelier, P.L.E. Archaea and their interactions with bacteria in a karst ecosystem. Front. Microbiol. 2023, 14, 1068595. [Google Scholar] [CrossRef] [PubMed]
- Dimkic, I.; Copic, M.; Petrovic, M.; Stupar, M.; Savkovic, Ž.; Kneževic, A.; Simic, G.S.; Grbic, M.L.; Unkovic, N. Bacteriobiota of the cave church of Sts. Peter and Paul in Serbia—Culturable and non-culturable communities’ assessment in the bioconservation potential of a peculiar fresco painting. Int. J. Mol. Sci. 2023, 24, 1016. [Google Scholar] [CrossRef]
- Riquelme, C.; Rigal, F.; Hathaway, J.J.M.; Northup, D.E.; Spilde, M.N.; Borges, P.A.V.; Gabriel, R.; Amorin, I.R.; Dapkevicius, M.L.N.E. Cave microbial community composition in oceanic islands: Disentangling the effect of different colored mats in diversity patterns of Azorean lava caves. FEMS Microbiol. Ecol. 2015, 91, fiv141. [Google Scholar] [CrossRef] [PubMed]
- Spilde, M.N.; Northup, D.E.; Caimi, N.A.; Boston, P.J.; Stone, F.D.; Smith, S. Microbial mat communities in Hawaiian lava caves. Int. Symp. Vulcanospeleol. 2016. Available online: https://www.cavepics.com/IVS17/SPILDE.pdf. (accessed on 29 October 2022).
- Lavoie, K.H.; Winter, A.S.; Read, K.J.H.; Hughes, E.M.; Spilde, M.N.; Northup, D.E. Comparison of bacterial communities from lava cave microbial mats to overlying surface soils from Lava Beds National Monument, USA. PLoS ONE 2017, 12, e0169339. [Google Scholar] [CrossRef] [PubMed]
- Weng, M.M.; Zaikova, E.; Millan, M.; Williams, A.J.; McAdam, A.C.; Knudson, C.A.; Fuqua, S.R.; Wagner, N.Y.; Craft, K.; Nawotniak, S.K.; et al. Life underground: Investigating microbial communities and their biomarkers in Mars-analog lava tubes at Craters of the Moon National Monument and Preserve. J. Geophys. Res. Planets 2022, 127, e2022JE007268. [Google Scholar] [CrossRef]
- Nicolosi, G.; Gonzalez-Pimentel, J.L.; Piano, E.; Isaia, M.; Miller, A.Z. First insights into the bacterial diversity of Mount Etna volcanic caves. Microb. Ecol. 2023, 85. in press. [Google Scholar] [CrossRef] [PubMed]
- Barton, H.A.; Taylor, N.M.; Kreate, M.P.; Springer, A.C.; Oehrle, S.A.; Bertog, J.L. The impact of host rock geochemistry on bacterial community structure in oligotrophic cave environments. Int. J. Speleol. 2007, 36, 93–104. [Google Scholar] [CrossRef]
- Ghezzi, D.; Sauro, F.; Columbu, A.; Carbone, C.; Hong, P.-Y.; Vergara, F.; De Waele, J.; Cappelletti, M. Transition from unclassified Ktedonobacterales to Actinobacteria during amorphous silica precipitation in a quartzite cave environment. Sci. Rep. 2021, 11, 3921. [Google Scholar] [CrossRef]
- Weber, C.F. Reduced vertical stratification of soil bacterial community structure and composition is associated with Bromus tectorum invasion of sagebrush steppe. J. Arid Environ. 2015, 115, 90–99. [Google Scholar] [CrossRef]
- Osman, J.R.; Fernandes, G.; Regeard, C.; Jaubert, C.; DuBow, M.S. Examination of the bacterial biodiversity of coastal eroded surface soils from the Padza de Dapani (Mayotte Island). Geomicrobiol. J. 2018, 35, 355–365. [Google Scholar] [CrossRef]
- Lambrechts, S.; Willems, A.; Tahon, G. Uncovering the uncultivated majority in Antarctic soils: Toward a synergistic approach. Front. Microbiol. 2019, 10, 242. [Google Scholar] [CrossRef]
- Li, J.; Wu, Z.; Yuan, J. Impact of agro-farming activities on microbial diversity of acidic red soils in a Camellia Oleifera Forest. Rev. Bras. Cien. Solo 2019, 43, e0190044. [Google Scholar] [CrossRef]
- Zhenqing, Z.; Binglin, Z.; Wei, Z.; Guangxiu, L.; Tuo, C.; Yang, L.; Jingwei, C.; Mao, T. Distribution characteristics and anti-radiation activity of culturable bacteria in black gobi ecosystem of the Hexi Corridor. J. Desert Res. 2020, 40, 52–62. [Google Scholar]
- Bossolani, J.; Crusciol, C.A.C.; Leite, M.F.A.; Merloti, L.F.; Moretti, L.G.; Pascoaloto, I.M.; Kuramae, E.E. Modulation of the soil microbiome by long-term Ca-based soil amendments boosts soil organic carbon and physicochemical quality in a tropical no-till crop rotation system. Soil Biol. Biochem. 2021, 156, 108188. [Google Scholar] [CrossRef]
- Chen, B.; Jiao, S.; Luo, S.; Ma, B.; Qi, W.; Cao, C.; Zhao, Z.; Du, G.; Ma, X. High soil pH enhances the network interactions among bacterial and archaeal microbiota in alpine grasslands of the Tibetan Plateau. Environ. Microbiol. 2021, 23, 464–477. [Google Scholar] [CrossRef]
- Liu, X.; Liu, Y.; Zhang, L.; Yin, R.; Wu, G.-L. Bacterial contributions of bio-crusts and litter crusts to nutrient cycling in the Mu Us Sandy Land. Catena 2021, 199, 105090. [Google Scholar] [CrossRef]
- Liu, Z.; Yang, Y.; Ji, S.; Dong, D.; Li, Y.; Wang, M.; Han, L.; Chen, X. Effects of elevation and distance from highway on the abundance and community structure of bacteria in soil along Qinghai-Tibet highway. Int. J. Environ. Res. Public Health 2021, 18, 13137. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Lipus, D.; Arens, F.L.; Baqué, M.; Bornemann, T.L.V.; de Vera, J.-P.; Flury, M.; Frösler, J.; Heinz, J.; Hwang, Y.; et al. Microbial hotspots in lithic microhabitats inferred from DNA fractionation and metagenomics in the Atacama Desert. Microorganisms 2021, 9, 1038. [Google Scholar] [CrossRef]
- Xie, J.; Wu, Z.; Zhang, X.; Peng, T.; Yang, C.; Zhang, J.; Liang, J. Diversity and structural characteristics of soil microbial communities in different habitats of wild Lilium regale Wilson in Wenchuan area. Bioengineered 2021, 12, 10457–10469. [Google Scholar] [CrossRef] [PubMed]
- Benaud, N.; Chelliah, D.S.; Wong, S.Y.; Ferrari, B.C. Soil substrate culturing approaches recover diverse members of Actinomycetota from desert soils of Herring Island, East Antarctica. Extremophiles 2022, 26, 24. [Google Scholar] [CrossRef] [PubMed]
- Guerra, V.A.; Beule, L.; Mackowiak, C.L.; Dubeux, J.C.B., Jr.; Blount, A.R.S.; Wang, X.-B.; Rowland, D.L.; Liao, H.-L. Soil bacterial community response to rhizoma peanut incorporation into Florida pastures. J. Environ. Qual. 2022, 51, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Ke, M.; Xu, N.; Zhang, Z.; Qiu, D.; Kang, J.; Lu, T.; Wang, T.; Peijnenburg, W.J.G.M.; Sun, L.; Hu, B.; et al. Development of a machine-learning model to identify the impacts of pesticides characteristics on soil microbial communities from high-throughput sequencing data. Environ. Microbiol. 2022, 24, 5561–5573. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, H. Alien species invasion of deep-sea bacteria into terrestrial soil. J. Clean. Product. 2022, 371, 133662. [Google Scholar] [CrossRef]
- Sun, H.; Peng, Q.; Guo, J.; Zhang, H.; Bai, J.; Mao, H. Effects of short-term soil exposure of different doses of ZnO nanoparticles on the soil environment and the growth and nitrogen fixation of alfalfa. Environ. Pollut. 2022, 309, 119817. [Google Scholar] [CrossRef]
- Topalovic, O.; Santos, S.S.; Heuer, H.; Nesme, J.; Kanfra, X.; Hallmann, J.; Sørensen, S.J.; Vestergård, M. Deciphering bacteria associated with a pre-parasitic stage of the root-knot nematode Meloidogyne hapla in nemato-suppressive and nemato-conducive soils. Appl. Soil Ecol. 2022, 172, 104344. [Google Scholar] [CrossRef]
- Wang, L.; Peng, C.; Gong, B.; Yang, Z.; Song, J.; Li, L.; Xu, L.; Yue, T.; Wang, X.; Yang, M.; et al. Actinobacteria community and their antibacterial and cytotoxic activity on the Weizhou and Xieyang volcanic islands in the Beibu Gulf of China. Front. Microbiol. 2022, 13, 911408. [Google Scholar] [CrossRef]
- Feng, Z.; Sun, H.; Qin, Y.; Zhou, Y.; Zhu, H.; Yao, Q. A synthetic community of siderophore-producing bacteria increases soil selenium bioavailability and plant uptake through regulation of the soil microbiome. Sci. Total Environ. 2023, 871, 162076. [Google Scholar] [CrossRef]
- Conte, A. Phylogenetic Diversity and Metabolic Potential of Prokaryotic Communities in Permafrost and Brine Pockets of Perennially Frozen Antarctic Lakes (Northern Victoria Land). Ph.D. Thesis, Università Degli Studi di Messina, Messina, Italy, 2017. [Google Scholar]
- Perez-Mon, C.; Stierli, B.; Plötze, M.; Frey, B. Fast and persistent responses of alpine permafrost microbial communities to in situ warming. Sci. Total Environ. 2022, 807, 150720. [Google Scholar] [CrossRef]
- Narendrula, R. Biochemical and Molecular Characterization of Microbial Communities from a Metal Contaminated and Reclaimed Region. Ph.D. Thesis, Laurentian University, Sudbury, ON, Canada, 2017. [Google Scholar]
- Lin, J.; He, F.; Su, B.; Sun, M.; Owens, G.; Chen, Z. The stabilizing mechanism of cadmium in contaminated soil using green synthesized iron oxide nanoparticles under long-term incubation. J. Hazard. Mater. 2019, 379, 120832. [Google Scholar] [CrossRef]
- Dong, S.; Liu, S.; Cui, S.; Zhou, X.; Gao, Q. Responses of soil properties and bacterial community to the application of sulfur fertilizers in black and sandy soils. Pol. J. Environ. Stud. 2022, 31, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Pu, C.; Liu, H.; Ding, G.; Sun, Y.; Yu, X.; Chen, J.; Ren, J.; Gong, X. Impact of direct application of biogas slurry and residue in fields: In situ analysis of antibiotic resistance genes from pig manure to fields. J. Hazard. Mater. 2018, 344, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Zhang, Y.; Yin, Y.; Zhu, X.; Zhu, W.; Zhou, Y. Comparison of soil bacterial community and functional characteristics following afforestation in the semi-arid areas. PeerJ 2019, 7, e7141. [Google Scholar] [CrossRef]
- Deng, J.; Zhou, Y.; Zhu, W.; Yin, Y. Effects of afforestation with Pinus sylvestris var. mongolica plantations combined with enclosure management on soil microbial community. PeerJ 2020, 8, e8857. [Google Scholar] [PubMed]
- Liu, K.; Ding, X.; Wang, J. Soil metabolome correlates with bacterial diversity and co-occurrence patterns in root-associated soils on the Tibetan Plateau. Sci. Total Environ. 2020, 735, 139572. [Google Scholar] [CrossRef]
- Jiang, H.; Chen, Y.; Hu, Y.; Wang, Z.; Lu, X. Soil bacterial communities and diversity in alpine grasslands on the Tibetan Plateau based on 16S rRNA gene sequencing. Front. Ecol. Evol. 2021, 9, 630722. [Google Scholar] [CrossRef]
- Solon, A.J.; Mastrangelo, C.; Vimercati, L.; Sommers, P.; Darcy, J.L.; Gendron, E.M.S.; Porazinska, D.L.; Schmidt, S.K. Gullies and moraines are islands of biodiversity in an arid, mountain landscape, Asgard Range, Antarctica. Front. Microbiol. 2021, 12, 654135. [Google Scholar] [CrossRef]
- Rodríguez-Berbel, N.; Ortega, R.; Lucas-Borja, M.E.; Solé-Benet, A.; Miralles, I. Long-term effects of two organic amendments on bacterial communities of calcareous mediterranean soils degraded by mining. J. Environ. Manag. 2020, 271, 110920. [Google Scholar] [CrossRef]
- Chuvochina, M.S.; Alekhina, I.A.; Normand, P.; Petit, J.-R.; Bulat, S.A. Three events of Saharan dust deposition on the Mont Blanc glacier associated with different snow-colonizing bacterial phylotypes. Microbiology 2011, 80, 125–131. [Google Scholar] [CrossRef]
- Hui, N.; Sun, N.; Du, H.; Umair, M.; Kang, H.; Liu, X.; Romantschuk, M.; Liu, C. Karst rocky desertification does not erode ectomycorrhizal fungal species richness but alters microbial community structure. Plant Soil 2019, 445, 383–396. [Google Scholar] [CrossRef]
- Reverdy, A.; Hathaway, D.; Jha, J.; Michaels, G.; Sullivan, J.; Diaz Mac-Adoo, D.; Riquelme, C.; Chai, Y.; Godoy, V.G. Insights into the diversity and survival strategies of soil bacterial isolates from the Atacama Desert. bioRxiv 2020. [Google Scholar] [CrossRef]
- Biderre-Petit, C.; Hochart, C.; Gardon, H.; Dugat-Bony, E.; Terrat, S.; Jouan-Dufournel, I.; Paris, R. Analysis of bacterial and archaeal communities associated with Fogo volcanic soils of different ages. FEMS Microbiol. Ecol. 2020, 96, fiaa104. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Du, W.; Xu, Y.; Wang, Y.L. Soil bacterial diversity and function in semi-arid forest parks in Baotou City. Biodivers. Sci. 2022, 30, 22245. [Google Scholar] [CrossRef]
- Li, Y.; Gao, W.; Wang, C.; Gao, M. Distinct distribution patterns and functional potentials of rare and abundant microorganisms between plastisphere and soils. Sci. Total Environ. 2023, 873, 162413. [Google Scholar] [CrossRef]
- Zhang, S.; Pei, L.; Zhao, Y.; Shan, J.; Zheng, X.; Xu, G.; Sun, Y.; Wang, F. Effects of microplastics and nitrogen deposition on soil multifunctionality, particularly C and N cycling. J. Hazard. Mater. 2023, 451, 131152. [Google Scholar] [CrossRef]
- Lin, Y.; Yang, L.; Chen, Z.; Gao, Y.; Kong, J.; He, Q.; Su, Y.; Li, J.; Qiu, Q. Seasonal variations of soil bacterial and fungal communities in a subtropical Eucalyptus plantation and their responses to throughfall reduction. Front. Microbiol. 2023, 14, 1113616. [Google Scholar] [CrossRef]
- De Tender, C.; Haegeman, A.; Vandecasteele, B.; Clement, L.; Cremelie, P.; Dawyndt, P.; Maes, M.; Debode, J. Dynamics in the strawberry rhizosphere microbiome in response to biochar and Botrytis cinerea leaf infection. Front. Microbiol. 2016, 7, 2062. [Google Scholar] [CrossRef]
- Echeverría Molinar, A. Efecto de Factores Abióticos y Bióticos Sobre la Estructura de la Comunidad Microbiana del Suelo en un Ambiente Oligotrófico. Master’s Thesis, Instituto Potosino de Investigación Científica y Tecnológica, San Luis Potosí, México, 2017. [Google Scholar]
- Visioli, G.; Sanangelantoni, A.M.; Vamerali, T.; Dal Cortivo, C.; Blandino, M. 16S rDNA profiling to reveal the influence of seed-applied biostimulants on the rhizosphere of young maize plants. Molecules 2018, 23, 1461. [Google Scholar] [CrossRef]
- Gao, X.; Wu, Z.; Liu, R.; Wu, J.; Zeng, Q.; Qi, Y. Rhizosphere bacterial community characteristics over different years of sugarcane ratooning in consecutive monoculture. BioMed Res. Int. 2019, 2019, 4943150. [Google Scholar] [CrossRef]
- Chen, Y.; Tian, W.; Shao, Y.; Li, Y.-J.; Lin, L.-A.; Zhang, Y.-J.; Han, H.; Chen, Z.-J. Miscanthus cultivation shapes rhizosphere microbial community structure and function as assessed by Illumina MiSeq sequencing combined with PICRUSt and FUNGUIld analyses. Arch. Microbiol. 2020, 202, 1157–1171. [Google Scholar] [CrossRef] [PubMed]
- López-Lozano, N.E.; Echeverría Molinar, A.; Ortiz Durán, E.A.; Hernández Rosales, M.; Souza, V. Bacterial diversity and interaction networks of Agave lechuguilla rhizosphere differ significantly from bulk soil in the oligotrophic basin of Cuatro Cienegas. Front. Plant Sci. 2020, 11, 1028. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, L.C.P. Comunidades Microbianas Rizosféricas de Plantas em Coexistência Sob Diferentes Condições Edáficas. Ph.D. Thesis, Universidade Federal de Viçosa, Viçosa, Brasil, 2020. [Google Scholar]
- Bettermann, A.; Zethof, J.H.T.; Babin, D.; Cammeraat, L.H.; Solé-Benet, A.; Lázaro, R.; Luna, L.; Nesme, J.; Sorensen, S.J.; Kalbitz, K.; et al. Importance of microbial communities at the root-soil interface for extracellular polymeric substances and soil aggregation in semiarid grasslands. Soil Biol. Biochem. 2021, 159, 108301. [Google Scholar] [CrossRef]
- Deng, Q.; Zhang, T.; Xie, D.; Yang, Y. Rhizosphere microbial communities are significantly affected by optimized phosphorus management in a slope farming system. Front. Microbiol. 2021, 12, 739844. [Google Scholar] [CrossRef]
- Engelbrecht, G.; Claassens, S.; Mienie, C.M.S.; Fourie, H. Screening of rhizosphere bacteria and nematode populations associated with soybean roots in the Mpumalanga Highveld of South Africa. Microorganisms 2021, 9, 1813. [Google Scholar] [CrossRef]
- Li, C.; Chen, G.; Zhang, J.; Zhu, P.; Bai, X.; Hou, Y.; Zhang, X. The comprehensive changes in soil properties are continuous cropping obstacles associated with American ginseng (Panax quinquefolius) cultivation. Sci. Rep. 2021, 11, 5068. [Google Scholar] [CrossRef]
- Ye, F.; Wang, X.; Wang, Y.; Wu, S.; Wu, J.; Hong, Y. Different pioneer plant species have similar rhizosphere microbial communities. Plant Soil 2021, 464, 165–181. [Google Scholar] [CrossRef]
- Zuo, J.; Zu, M.; Liu, L.; Song, X.; Yuan, Y. Composition and diversity of bacterial communities in the rhizosphere of the Chinese medicinal herb Dendrobium. BMC Plant Biol. 2021, 21, 127. [Google Scholar] [CrossRef]
- Gu, Y.-Y.; Zhang, H.-Y.; Liang, X.-Y.; Fu, R.; Li, M.; Chen, C.-J. Impact of biochar and bioorganic fertilizer on rhizosphere bacteria in saline–alkali soil. Microorganisms 2022, 10, 2310. [Google Scholar] [CrossRef]
- He, C.; Wang, R.; Ding, W.; Li, Y. Effects of cultivation soils and ages on microbiome in the rhizosphere soil of Panax ginseng. Appl. Soil Ecol. 2022, 174, 104397. [Google Scholar] [CrossRef]
- Jara-Servin, A.; Silva, A.; Barajas, H.; Cruz-Ortega, R.; Tinoco-Ojanguren, C.; Alcaraz, L.D. The Allelopathic Buffelgrass and Its Rhizosphere Microbiome. Available online: https://papers.ssrn.com/sol3/papers.cfm?abstract_id=4263505 (accessed on 28 February 2023).
- Kushwaha, P.; Neilson, J.W.; Maier, R.M.; Babst-Kostecka, A. Soil microbial community and abiotic soil properties influence Zn and Cd hyperaccumulation differently in Arabidopsis halleri. Sci. Total Environ. 2022, 803, 150006. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Zhang, Y.; Liang, X.; Duan, R.; Yang, L.; Du, Y.; Wu, L.; Huang, J.; Xiang, G.; Bai, J.; et al. Assessment of rhizosphere bacterial diversity and composition in a metal hyperaccumulator (Boehmeria nivea) and a nonaccumulator (Artemisia annua) in an antimony mine. J. Appl. Microbiol. 2022, 132, 3432–3443. [Google Scholar] [CrossRef] [PubMed]
- Lizano-Bastardín, A.L.; Villadas, P.J.; Pulido-Suárez, L.; Fernández-López, M.; León-Barrios, M. The Rhizosphere Microbiome Associated with the Legume Spartocytisus supranubius in the High Mountain Ecosystem of Teide N.P. First Spanish-Portuguese Congress on Beneficial Plant-Microbe Interactions, Poster S1-P-03. Available online: https://events.iniav.pt/bemiplant/images/book-abstracts_15-10-2022.pdf (accessed on 9 February 2023).
- Ren, H.; Islam, M.S.; Wang, H.; Guo, H.; Wang, Z.; Qi, X.; Zhang, S.; Guo, J.; Wang, Q.; Li, B. Effect of humic acid on soil physical and chemical properties, microbial community structure, and metabolites of decline diseased bayberry. Int. J. Mol. Sci. 2022, 23, 14707. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, T.; Isobe, K.; Imada, S.; Eltayeb, M.; Akaji, Y.; Nakayama, M.; Allen, M.F.; Aronson, E. Root endophytic bacterial and fungal communities in a natural hot desert are differentially regulated in dry and wet seasons by stochastic processes and functional traits. Available online: https://ssrn.com/abstract=4363629 (accessed on 25 February 2023).
- Zhou, Y.; Pang, Z.; Jia, H.; Yuan, Z.; Ming, R. Responses of roots and rhizosphere of female papaya to the exogenous application of GA3. BMC Plant Biol. 2023, 23, 35. [Google Scholar] [CrossRef] [PubMed]
- Obieze, C.C.; George, P.B.L.; Boyle, B.; Khasa, D.P. Black pepper rhizomicrobiome: Spectrum of plant health indicators, critical environmental factors and community compartmentation in Vietnam. Appl. Soil Ecol. 2023, 187, 104857. [Google Scholar] [CrossRef]
- Deng, J.; Yin, Y.; Zhu, W.; Zhou, Y. Response of soil environment factors and microbial communities to phytoremediation with Robinia pseudoacacia in an open-cut magnesite mine. Land Degrad. Dev. 2020, 31, 2340–2355. [Google Scholar] [CrossRef]
- Ezeokoli, O.T.; Bezuidenhout, C.C.; Maboeta, M.S.; Khasa, D.P.; Adeleke, R.A. Structural and functional differentiation of bacterial communities in post-coal mining reclamation soils of South Africa: Bioindicators of soil ecosystem restoration. Sci. Rep. 2020, 10, 1759. [Google Scholar] [CrossRef]
- Thompson, E.; Erickson, M.; Malik, N.; Mettler, R.; Reman, B.; Ren, Y.; Bergmann, D. Culture-independent characterization of “cave silver” biofilms from the 1470 m level of the Sanford Underground Research Facility, Lead, SD. Proc. South Dak. Acad. Sci. 2020, 99, 29–55. [Google Scholar]
- Arif, S.; Nacke, H.; Schliekmann, E.; Reimer, A.; Arp, G.; Hoppert, M. Composition and niche-specific characteristics of microbial consortia colonizing Marsberg copper mine in the Rhenish Massif. Biogeosciences 2021, 19, 4883–4902. [Google Scholar] [CrossRef]
- Shi, A.; Hu, Y.; Zhang, X.; Zhou, D.; Xu, J.; Rensing, C.; Zhang, L.; Xing, S.; Ni, W.; Yang, W. Biochar loaded with bacteria enhanced Cd/Zn phytoextraction by facilitating plant growth and shaping rhizospheric microbial community. Environ. Pollut. 2023, 327, 121559. [Google Scholar] [CrossRef]
- Diaz-Herraiz, M.; Jurado, V.; Cuezva, S.; Laiz, L.; Pallecchi, P.; Tiano, P.; Sanchez-Moral, S.; Saiz-Jimenez, C. Deterioration of an Etruscan tomb by bacteria from the order Rhizobiales. Sci. Rep. 2014, 4, 3610. [Google Scholar] [CrossRef] [PubMed]
- Zacharenka, F. Study of the Bacterial Diversity on Ancient Mural Paintings from Kalliroi’s Fountain and Pana’s Sanctuary. Master’s Thesis, University of Thessaly, Volos, Greece, 2014. [Google Scholar]
- Duan, Y.; Wu, F.; Wang, W.; He, D.; Gu, J.-D.; Feng, H.; Chen, T.; Liu, G.; An, L. The microbial community characteristics of ancient painted sculptures in Maijishan Grottoes, China. PLoS ONE 2017, 12, e0179718. [Google Scholar]
- Li, Q.; Zhang, B.; Wang, L.; Ge, Q. Distribution and diversity of bacteria and fungi colonizing ancient Buddhist statues analyzed by high-throughput sequencing. Int. Biodeter. Biodegr. 2017, 117, 245–254. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, B.; Yang, X.; Ge, Q. Deterioration-associated microbiome of stone monuments: Structure, variation, and assembly. Appl. Environ. Microbiol. 2018, 84, e02680-17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ge, Q.; Zhu, Z.; Deng, Y.; Gu, J.-D. Microbiological community of the Royal Palace in Angkor Thom and Beng Mealea of Cambodia by Illumina sequencing based on 16S rRNA gene. Int. Biodeter. Biodegr. 2018, 134, 127–135. [Google Scholar] [CrossRef]
- Dyda, M.; Pyzik, A.; Wilkojc, E.; Kwiatkowska-Kopka, B.; Sklodowska, A. Bacterial and fungal diversity inside the medieval building constructed with sandstone plates and lime mortar as an example of the microbial colonization of a nutrient-limited extreme environment (Wawel Royal Castle, Krakow, Poland). Microorganisms 2019, 7, 416. [Google Scholar] [CrossRef]
- Jroundi, F.; Elert, K.; Ruiz-Agudo, E.; Gonzalez-Muñoz, M.T.; Rodriguez-Navarro, C. Bacterial diversity evolution in Maya plaster and stone following a bio-conservation treatment. Front. Microbiol. 2020, 11, 599144. [Google Scholar] [CrossRef]
- Louati, M.; Ennis, N.J.; Ghodhbane-Gtari, F.; Hezbri, K.; Sevigny, J.L.; Fahnestock, M.F.; Cherif-Silini, H.; Bryce, J.G.; Tisa, L.S.; Gtari, M. Elucidating the ecological networks in stone-dwelling microbiomes. Environ. Microbiol. 2020, 22, 1467–1480. [Google Scholar] [CrossRef]
- Schröer, L.; De Kock, T.; Cnudde, V.; Boon, N. Differential colonization of microbial communities inhabiting Lede stone in the urban and rural environment. Sci. Total Environ. 2020, 733, 139339. [Google Scholar] [CrossRef]
- Yang, S.; Wu, L.; Wu, B.; Zhang, Y.; Wang, H.; Tan, X. Diversity and structure of soil microbiota of the Jinsha earthen relic. PLoS ONE 2020, 15, e0236165. [Google Scholar] [CrossRef]
- Coelho, C.; Mesquita, N.; Costa, I.; Soares, F.; Trovão, J.; Freitas, H.; Portugal, A.; Tiago, I. Bacterial and archaeal structural diversity in several biodeterioration patterns on the limestone walls of the Old Cathedral of Coimbra. Microorganisms 2021, 9, 709. [Google Scholar] [CrossRef] [PubMed]
- Ipekci, E. Evaluation of Stone Deterioration Problems of Anavarza Archaeological Site for the Purpose of Conservation. Ph.D. Thesis, İzmir Institute of Technology, Urla, Turkey, 2021. [Google Scholar]
- Sansupa, C.; Suphaphimol, N.; Nonthijun, P.; Ronsuek, T.; Yimklan, S.; Semakul, N.; Khrueraya, T.; Suwannarach, N.; Purahong, W.; Disayathanoowat, T. The microbiome of a 13th century Lan Na mural painting: Diversity, taxonomic distribution and their biodeterioration potentials. Microorganisms 2023, 11. in press. [Google Scholar]
- Chen, P.; Zhang, L.; Guo, X.; Dai, X.; Liu, L.; Xi, L.; Wang, J.; Song, L.; Wang, Y.; Zhu, Y.; et al. Diversity, biogeography, and biodegradation potential of actinobacteria in the deep-sea sediments along the Southwest Indian Ridge. Front. Microbiol. 2016, 7, 1340. [Google Scholar] [CrossRef] [PubMed]
- Pietrzak, K.; Otlewska, A.; Dybka, K.; Danielewicz, D.; Pangallo, D.; Demnerová, K.; Ďurovič, M.; Kraková, L.; Scholtz, V.; Bučková, M.; et al. A modern approach to biodeterioration assessment and disinfection of historical book. In A Modern Approach to Biodeterioration Assessment and the Disinfection of Historical Book Collections; Gutarowska, B., Ed.; Institute of Fermentation Technology and Microbiology, Łódź University of Technology: Łódź, Poland, 2016; pp. 81–125. [Google Scholar]
- Gozari, M.; Bahador, N.; Jassbi, A.R.; Mortazavi, M.S.; Hamzehei, S.; Eftekhar, E. Isolation, distribution and evaluation of cytotoxic and antioxidant activity of cultivable actinobacteria from the Oman Sea sediments. Acta Oceanol. Sin. 2019, 38, 84–90. [Google Scholar] [CrossRef]
- Van Assche, A. Characterization of the Bacterial Community Composition in Drinking Water Production and Distribution Systems, Emphasizing Acinetobacter species. Ph.D. Thesis, University of Leuven, Leuven, Belgium, 2019. [Google Scholar]
- Argudo Fernández, M. Microbial Communities Responses in Fluvial Biofilms under Metal Stressed Scenarios. Ph.D. Thesis, Universitat de Girona, Girona, Spain, 2020. [Google Scholar]
- Cao, W.; Xiong, Y.; Zhao, D.; Tan, H.; Qu, J. Bryophytes and the symbiotic microorganisms, the pioneers of vegetation restoration in karst rocky desertification areas in southwestern China. Appl. Microbiol. Biotechnol. 2020, 104, 873–891. [Google Scholar] [CrossRef] [PubMed]
- Caffrey, P.; Hogan, M.; Song, Y. New glycosylated polyene macrolides: Refining the ore from genome mining. Antibiotics 2022, 11, 334. [Google Scholar] [CrossRef]
- Yang, Y.; Qiu, J.; Wang, X. Exploring the dynamic of bacterial communities in Manila clam (Ruditapes philippinarum) during refrigerated storage. Front. Microbiol. 2022, 13, 882629. [Google Scholar] [CrossRef]
- Boquet, E.; Boronat, A.; Ramos-Cormenzana, A. Production of calcite (calcium carbonate) crystals by soil bacteria is a general phenomenon. Nature 1973, 246, 527–528. [Google Scholar] [CrossRef]
- Barton, H.A.; Northup, D.E. Geomicrobiology in cave environments: Past, current and future perspectives. J. Cave Karst Stud. 2007, 69, 163–178. [Google Scholar]
- Banks, E.D.; Taylor, N.M.; Gulley, J.; Lubbers, B.R.; Giarrizzo, J.G.; Bullen, H.A.; Hoehler, T.M.; Barton, H.A. Bacterial calcium carbonate precipitation in cave environments: A function of calcium homeostasis. Geomicrobiol. J. 2010, 27, 444–454. [Google Scholar] [CrossRef]
- Cañaveras, J.C.; Hoyos, M.; Sanchez-Moral, S.; Sanz-Rubio, E.; Bedoya, J.; Soler, V.; Groth, I.; Schumann, P.; Laiz, L.; Gonzalez, I.; et al. Microbial communities associated to hydromagnesite and needle fiber aragonite deposits in a karstic cave (Altamira, Northern Spain). Geomicrobiol. J. 1999, 16, 9–25. [Google Scholar]
- Sanchez-Moral, S.; Cañaveras, J.C.; Laiz, L.; Saiz-Jimenez, C.; Bedoya, J.; Luque, L. Biomediated precipitation of calcium carbonate metastable phases in hypogean environments: A short review. Geomicrobiol. J. 2003, 20, 491–500. [Google Scholar] [CrossRef]
- Cuezva, S.; Fernandez-Cortes, A.; Porca, E.; Pasic, L.; Jurado, V.; Hernandez-Marine, M.; Serrano-Ortiz, P.; Cañaveras, J.C.; Sanchez-Moral, S.; Saiz-Jimenez, C. The biogeochemical role of Actinobacteria in Altamira Cave, Spain. FEMS Microbiol. Ecol. 2012, 81, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Maciejewska, M.; Adam, D.; Naômé, A.; Martinet, L.; Tenconi, E.; Calusinska, M.; Delfosse, P.; Hanikenne, M.; Baurain, D.; Compère, P.; et al. Assessment of the potential role of Streptomyces in cave moonmilk formation. Front. Microbiol. 2017, 8, 1181. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, T.D.; Reeksting, B.J.; Gebhard, S. Bacteria-induced mineral precipitation: A mechanistic review. Microbiology 2021, 167, 001049. [Google Scholar] [CrossRef]
- Tomé, D. Yeast extracts: Nutritional and flavoring food ingredients. ACS Food Sci. Technol. 2021, 1, 487–494. [Google Scholar] [CrossRef]
- Joubert, J.J.; van Rensburg, E.J.; Pitout, M.J. A plate method for demonstrating the breakdown of heparin and chrondoitin sulphate by bacteria. J. Microbiol. Methods 1984, 2, 197–202. [Google Scholar] [CrossRef]
- Sanchez-Moral, S.; Luque, L.; Cañaveras, J.C.; Laiz, L.; Jurado, V.; Saiz-Jimenez, C. Bioinduced barium precipitation in St. Callixtus and Domitilla catacombs. Ann. Microbiol. 2004, 54, 1–12. [Google Scholar]
- Sanchez-Moral, S.; Bedoya, J.; Luque, L.; Cañaveras, J.C.; Jurado, V.; Laiz, L.; Saiz-Jimenez, C. Biomineralization of different crystalline phases by bacteria isolated from catacombs. In Molecular Biology and Cultural Heritage; Saiz-Jimenez, C., Ed.; Balkema: Lisse, The Netherlands, 2003; pp. 179–185. [Google Scholar]
- Rivadeneyra, M.A.; Ramos-Cormenzana, A.; García-Cervigón, A. Bacterial formation of struvite. Geomicrobiol. J. 1983, 3, 151–163. [Google Scholar] [CrossRef]
- Rivadeneyra, M.A.; Pérez-García, I.; Ramos-Cormenzana, A. Influence of ammonium ion on bacterial struvite production. Geomicrobiol. J. 1992, 10, 125–137. [Google Scholar] [CrossRef]
- Manzoor, M.A.P.; Singh, B.; Agrawal, A.K.; Arun, A.B.; Mujeeburahiman, M.; Rekha, P.-D. Morphological and micro-tomographic study on evolution of struvite in synthetic urine infected with bacteria and investigation of its pathological biomineralization. PLoS ONE 2018, 13, e0202306. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Román, M.; Rivadeneyra, M.A.; Vasconcelos, C.; McKenzie, J.A. Biomineralization of carbonate and phosphate by moderately halophilic bacteria. FEMS Microbiol. Ecol. 2007, 61, 273–284. [Google Scholar] [CrossRef]
- Cañaveras, J.C.; Cuezva, S.; Sanchez-Moral, S.; Lario, J.; Laiz, L.; Gonzalez, J.M.; Saiz-Jimenez, C. On the origin of fiber calcite crystals in moonmilk deposits. Naturwissenschaften 2006, 93, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Onac, B.P.; Ghergari, L. Moonmilk mineralogy in some Romanian and Norwegian Caves. Cave Sci. 1993, 20, 106–120. [Google Scholar]
- Cañaveras, J.C.; Sanchez–Moral, S.; Soler, V.; Saiz-Jimenez, C. Microorganisms and microbially induced fabrics in cave walls. Geomicrobiol. J. 2001, 18, 223–240. [Google Scholar]
- Baskar, S.; Baskar, R.; Routh, J. Biogenic evidences of moonmilk deposition in the Mawmluh Cave, Meghalaya, India. Geomicrobiol. J. 2011, 28, 252–265. [Google Scholar]
- Jones, B.; Peng, X. Abiogenic growth of needle-fiber calcite in spring towers at Shiqiang, Yunnan province, China. J. Sediment. Res. 2014, 84, 1021–1040. [Google Scholar] [CrossRef]
- Cuezva, S.; Sanchez-Moral, S.; Saiz-Jimenez, C.; Cañaveras, J.C. Microbial communities and associated mineral fabrics in Altamira Cave, Spain. Int. J. Speleol. 2009, 38, 83–92. [Google Scholar]
- Sanchez-Moral, S.; Portillo, M.C.; Janices, I.; Cuezva, S.; Fernández-Cortés, A.; Cañaveras, J.C.; Gonzalez, J.M. The role of microorganisms in the formation of calcitic moonmilk deposits and speleothems in Altamira Cave. Geomorphology 2012, 139–140, 285–292. [Google Scholar] [CrossRef]
- Maciejewska, M.; Calusinska, M.; Cornet, L.; Adam, D.; Pessi, I.S.; Malchair, S.; Delfosse, P.; Baurain, D.; Barton, H.; Carnol, M.; et al. High-throughput sequencing analysis of the actinobacterial spatial diversity in moonmilk deposits. Antibiotics 2018, 7, 27. [Google Scholar] [CrossRef]
- Groth, I.; Schumann, P.; Laiz, L.; Sanchez–Moral, S.; Cañaveras, J.C.; Saiz-Jimenez, C. Geomicrobiological study of the Grotta dei Cervi, Porto Badisco, Italy. Geomicrobiol. J. 2001, 18, 241–258. [Google Scholar] [CrossRef]
- Park, S.; Cho, Y.-J.; Jung, D.; Jo, K.; Lee, E.-J.; Lee, J.-S. Microbial diversity in moonmilk of Baeg-nyong Cave, Korean CZO. Front. Microbiol. 2020, 11, 613. [Google Scholar] [CrossRef] [PubMed]
- Cirigliano, A.; Tomassetti, M.C.; Di Pietro, M.; Mura, F.; Maneschi, M.L.; Gentili, M.D.; Cardazzo, B.; Arrighi, C.; Mazzoni, C.; Negri, R.; et al. Calcite moonmilk of microbial origin in the Etruscan Tomba degli Scudi in Tarquinia, Italy. Sci. Rep. 2018, 8, 15839. [Google Scholar] [CrossRef] [PubMed]
- Maciejewska, M.; Pessi, I.S.; Arguelles-Arias, A.; Noirfalise, P.; Luis, G.; Ongena, M.; Barton, H.; Carnol, M.; Rigali, S. Streptomyces lunaelactis sp. nov., a novel ferroverdin A-producing Streptomyces species isolated from a moonmilk speleothem. Anton. Leeuw. 2015, 107, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Svec, P.; Kosina, M.; Zeman, M.; Holochová, P.; Králová, S.; Nemcová, E.; Micenková, L.; Urvashi, M.; Gupta, V.; Sood, U.; et al. Pseudomonas karstica sp. nov. and Pseudomonas spelaei sp. nov., isolated from calcite moonmilk deposits from caves. Int. J. Syst. Evol. Microbiol. 2020, 70, 5131–5140. [Google Scholar] [CrossRef]
- Adam, D.; Maciejewska, M.; Naômé, A.; Martinet, L.; Coppieters, W.; Karim, L.; Baurain, D.; Rigali, S. Isolation, Characterization, and antibacterial activity of hard-to-culture Actinobacteria from cave moonmilk deposits. Antibiotics 2018, 7, 28. [Google Scholar] [CrossRef]
- Jaroszewicz, W.; Bielanska, P.; Lubomska, D.; Kosznik-Kwasnicka, K.; Golec, P.; Grabowski, Ł.; Wieczerzak, E.; Drózdz, W.; Gaffke, L.; Pierzynowska, K.; et al. Antibacterial, antifungal and anticancer activities of compounds produced by newly isolated Streptomyces strains from the Szczelina Chochołowska Cave (Tatra Mountains, Poland). Antibiotics 2021, 10, 1212. [Google Scholar] [CrossRef]
- Jaroszewicz, W.; Bielanska, P.; Lubomska, D.; Kosznik-Kwasnicka, K.; Golec, P.; Grabowski, Ł.; Wieczerzak, E.; Drózdz, W.; Gaffke, L.; Pierzynowska, K.; et al. Antimicrobial activities of compounds produced by newly isolated Streptomyces strains from Mountain Caves. Med. Sci. Forum 2022, 12, 7. [Google Scholar]
- Maciejewska, M.; Adam, D.; Martinet, L.; Naômé, A.; Całusińska, M.; Delfosse, P.; Carnol, M.; Barton, H.A.; Hayette, M.-P.; Smargiasso, N.; et al. A Phenotypic and genotypic analysis of the antimicrobial potential of cultivable Streptomyces isolated from cave moonmilk deposits. Front. Microbiol. 2016, 7, 1455. [Google Scholar] [CrossRef]
- Martinet, L.; Naômé, A.; Rezende, L.C.D.; Tellatin, D.; Pignon, B.; Docquier, J.-D.; Sannio, F.; Baiwir, D.; Mazzucchelli, G.; Frédérich, M.; et al. Lunaemycins, new cyclic hexapeptide antibiotics from the cave moonmilk-dweller Streptomyces lunaelactis MM109T. Int. J. Mol. Sci. 2023, 24, 1114. [Google Scholar] [CrossRef]
- Martinet, L.; Naômé, A.; Baiwir, D.; De Pauw, E.; Mazzucchelli, G.; Rigali, S. On the risks of phylogeny-based strain prioritization for drug discovery: Streptomyces lunaelactis as a case study. Biomolecules 2020, 10, 1027. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Cave | Relative Abundance | Genus | Type of Sample | References |
|---|---|---|---|---|
| Pindal | 16.0–27.1 | Crossiella | Moonmilk | [26,28,31] |
| 1.4–1.7 | Crossiella | Sediment under moonmilk | ||
| 11.3–11.7 | Crossiella | Top-layer sediments | ||
| 6.0–9.0 | Crossiella | Sediments | ||
| 5.3–7.9 | Crossiella | Yellow biofilm | ||
| 2.0–8.0 7.0–8.0 | Crossiella Crossiella | Grey biofilms Pink biofilms | ||
| Fuente de la Canaria | 12.6–12.8 | Crossiella | Mucous formations | [29] |
| 12.3 | Crossiella | Moonmilk | ||
| 6.7 | Crossiella | Brown and yellow deposits | ||
| Bucara II | 38.9 24.9 | Crossiella Crossiella | Pink deposit Moonmilk | [29] |
| Nerja | 0.1–1.5 | Crossiella | Phototrophic biofilms | [30] |
| Castañar | 15.0 | Crossiella | Grey biofilm | [31] |
| Altamira | >20.0 27.0 38.0 | Crossiella Crossiella Crossiella | Grey biofilms White biofilms Yellow biofilms | [8] |
| Covadura | 26.4–54.1 21.8–51.9 4.5–19.7 | Crossiella Crossiella Crossiella | White biofilm Yellow biofilm Sediments | Unpublished data |
| Yeso | 1.3–13.3 | Crossiella | Sediments | Unpublished data |
| Thyssen Museum basement | 16.6 64.2 2.8–7.4 | Crossiella Crossiella Crossiella | White biofilm Grey biofilm Sediment | [32] |
| Karstic Caves | Relative Abundance% | Genus | Type of Sample (Method) | References |
|---|---|---|---|---|
| Heshang | n.d. | Crossiella | Weathered rocks (NGS) | [35] |
| Laugerie-Haute | 4.0 | Crossiella | Salt efflorescences (clones) | [36] |
| Sorcerers | 30.0 | Crossiella | Salt efflorescences (NGS) | [37] |
| Pillier | n.d. | Crossiella | Wall rock (NGS) | [38] |
| Yixing Shanjuan | 3.9 | Crossiella | Speleothem (NGS) | [39] |
| Shuanghe | 9.5 | Crossiella | Rock (NGS) | [40] |
| Manao-Pee | 4.1 | Crossiella | Soil (NGS) | [41] |
| KN14 | 27.1–52.3 | Crossiella | Rock/Clay (NGS) | [42] |
| RN5 | 1.0–17.9 | Crossiella | Rock/Clay/Mud (NGS) | [42] |
| Maijishan Grottoes | n.d. | Crossiella | Walls paintings (NGS) | [43] |
| Heshang | n.d. | Crossiella | Weathered rocks (NGS) | [44] |
| Kashmir and Tiser | 11.9–36.6 | Crossiella | Soil (NGS) | [45] |
| Zhijin | 4.1 | Crossiella | Wall rock (NGS) | [46] |
| Rouffignac | ~70.0 | Crossiella | Wall rock (NGS) | [47] |
| Stiffe | 9.9 | Crossiella | Biofilms (NGS) | [48] |
| Heshang | n.d. | Crossiella | Weathered rocks (NGS) | [49] |
| Cave Church | 0.1–4.9 | Crossiella | Fresco (NGS) | [50] |
| Volcanic Caves | ||||
| Azorean caves | 18.6 | Crossiella | Biofilms (clones) | [51] |
| Hawaiian caves | n.d. | Crossiella | Biofilms (NGS) | [52] |
| Californian caves | n.d. | Crossiella | Biofilms (NGS) | [53] |
| Idahoan caves | n.d. | Crossiella | Biofilms (NGS) | [54] |
| Sicilian caves | 62.5–77.6 | Crossiella | Biofilms (NGS) | [55] |
| Other Cave Types | ||||
| Carlsbad Cavern | n.d. | Crossiella | Rocks (clones) | [56] |
| Imawarì Yeuta | n.d. | Crossiella | Patina/Speleothems (NGS) | [57] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin-Pozas, T.; Gonzalez-Pimentel, J.L.; Jurado, V.; Laiz, L.; Cañaveras, J.C.; Fernandez-Cortes, A.; Cuezva, S.; Sanchez-Moral, S.; Saiz-Jimenez, C. Crossiella, a Rare Actinomycetota Genus, Abundant in the Environment. Appl. Biosci. 2023, 2, 194-210. https://doi.org/10.3390/applbiosci2020014
Martin-Pozas T, Gonzalez-Pimentel JL, Jurado V, Laiz L, Cañaveras JC, Fernandez-Cortes A, Cuezva S, Sanchez-Moral S, Saiz-Jimenez C. Crossiella, a Rare Actinomycetota Genus, Abundant in the Environment. Applied Biosciences. 2023; 2(2):194-210. https://doi.org/10.3390/applbiosci2020014
Chicago/Turabian StyleMartin-Pozas, Tamara, Jose Luis Gonzalez-Pimentel, Valme Jurado, Leonila Laiz, Juan Carlos Cañaveras, Angel Fernandez-Cortes, Soledad Cuezva, Sergio Sanchez-Moral, and Cesareo Saiz-Jimenez. 2023. "Crossiella, a Rare Actinomycetota Genus, Abundant in the Environment" Applied Biosciences 2, no. 2: 194-210. https://doi.org/10.3390/applbiosci2020014