
Habituation and Behavioural Response of Confinement-Induced Anxiety Conditions in a Zebrafish Model

Abstract
:1. Introduction
2. Methodology
2.1. Animal Husbandry
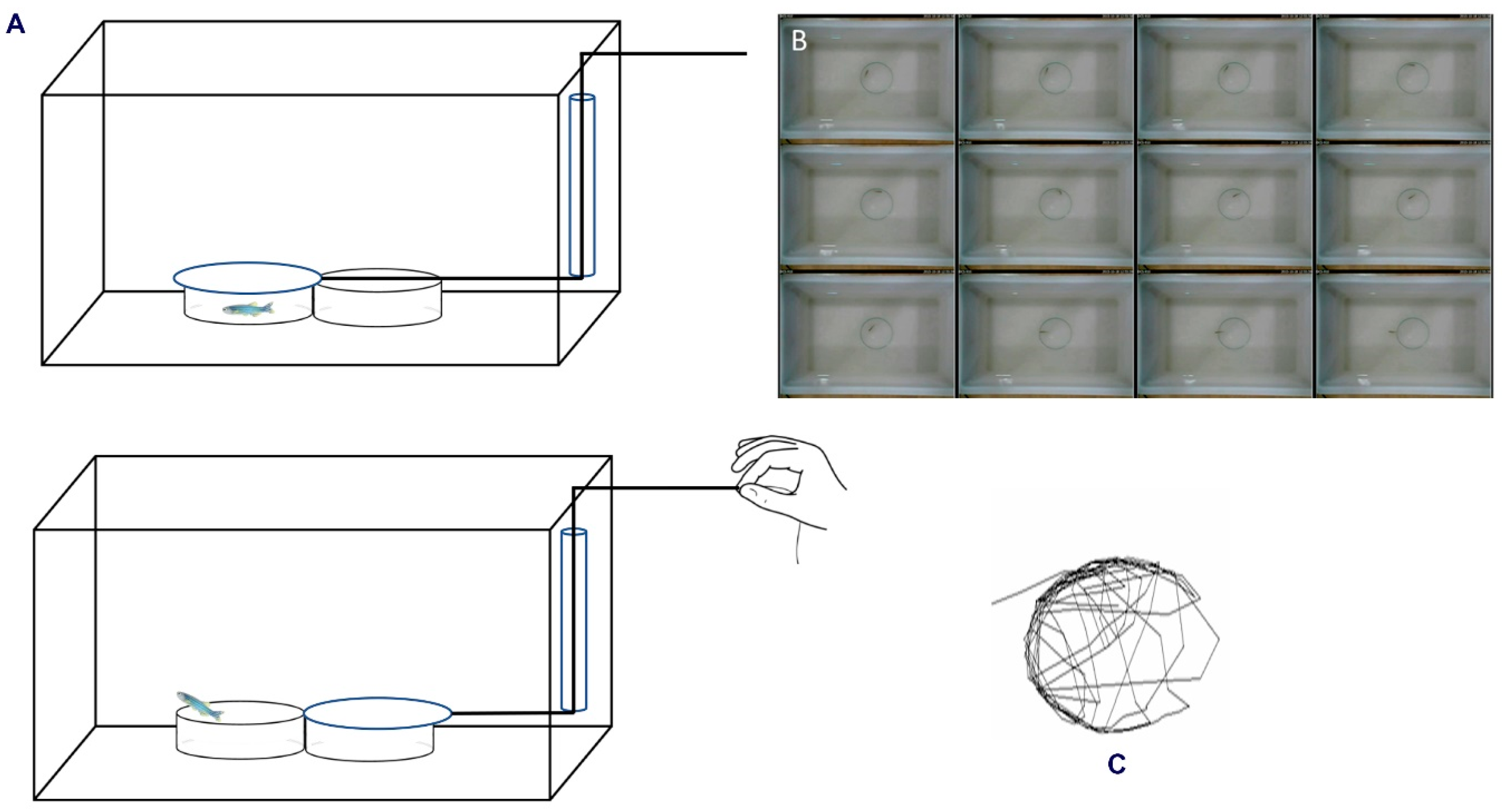
2.2. Experimental Setup
2.3. Behaviour Analysis
2.4. Statistical Analysis
3. Results
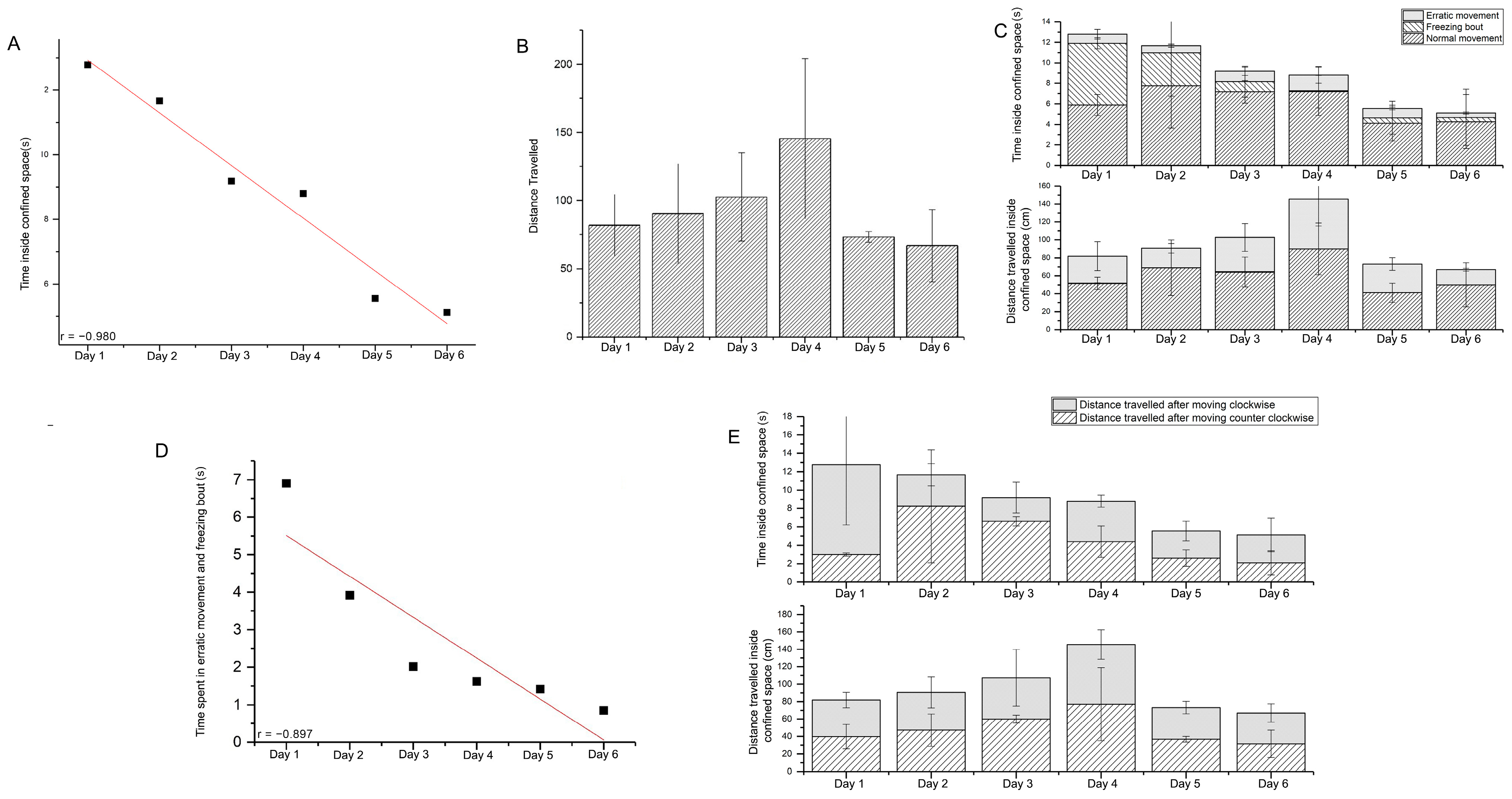
3.1. Response Time for Escaping from Confinement
3.2. Effect of Confined Space on the Movement of Zebrafish
3.3. Effect of Confined Space on Anxiety
3.4. Effect of Confined Space on Rotational Movement
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Guo, S. Linking Genes to Brain, Behavior and Neurological Diseases: What Can We Learn from Zebrafish? Genes. Brain. Behav. 2004, 3, 63–74. [Google Scholar] [CrossRef]
- Roberts, A.C.; Bill, B.R.; Glanzman, D.L. Learning and Memory in Zebrafish Larvae. Front. Neural Circuits 2013, 7, 126. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, Y.; Talpos, A.; Gerlai, R. Towards the Characterization of Short-Term Memory of Zebrafish: Effect of Fixed versus Random Reward Location. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2015, 56, 189–195. [Google Scholar] [CrossRef]
- Vuaden, F.C.; Savio, L.E.B.; Piato, A.L.; Pereira, T.C.; Vianna, M.R.; Bogo, M.R.; Bonan, C.D.; Wyse, A.T.S. Long-Term Methionine Exposure Induces Memory Impairment on Inhibitory Avoidance Task and Alters Acetylcholinesterase Activity and Expression in Zebrafish (Danio Rerio). Neurochem. Res. 2012, 37, 1545–1553. [Google Scholar] [CrossRef]
- Girdhar, K.; Gruebele, M.; Chemla, Y.R. The Behavioral Space of Zebrafish Locomotion and Its Neural Network Analog. PLoS ONE 2015, 10, e0128668. [Google Scholar] [CrossRef] [Green Version]
- Stewart, A.; Kadri, F.; Dileo, J.; Chung, K.M.; Cachat, J.; Goodspeed, J.; Suciu, C.; Roy, S.; Gaikwad, S.; Wong, K.; et al. The Developing Utility of Zebrafish in Modeling Neurobehavioral Disorders. Int. J. Comp. Psychol. 2010, 23, 104–120. [Google Scholar] [CrossRef]
- Dabrowski, K.; Miller, M. Contested Paradigm in Raising Zebrafish (Danio Rerio). Zebrafish 2018, 15, 295–309. [Google Scholar] [CrossRef] [Green Version]
- Pather, S.; Gerlai, R. Shuttle Box Learning in Zebrafish (Danio Rerio). Behav. Brain Res. 2009, 196, 323–327. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Scott-Scheiern, T.; Kempker, L.; Simons, K. Active Avoidance Conditioning in Zebrafish (Danio Rerio). Neurobiol. Learn. Mem. 2007, 87, 72–77. [Google Scholar] [CrossRef]
- Aoki, R.; Tsuboi, T.; Okamoto, H. Y-Maze Avoidance: An Automated and Rapid Associative Learning Paradigm in Zebrafish. Neurosci. Res. 2015, 91, 69–72. [Google Scholar] [CrossRef]
- Mathur, P.; Lau, B.; Guo, S. Conditioned Place Preference Behavior in Zebrafish. Nat. Protoc. 2011, 6, 338–345. [Google Scholar] [CrossRef]
- Blaser, R.E.; Chadwick, L.; McGinnis, G.C. Behavioral Measures of Anxiety in Zebrafish (Danio Rerio). Behav. Brain Res. 2010, 208, 56–62. [Google Scholar] [CrossRef]
- Morales, E.E.; Wingert, R.A. Zebrafish as a Model of Kidney Disease. In Results and Problems in Cell Differentiation; Springer: New York, NY, USA, 2017; Volume 60, pp. 55–75. ISBN 9789529253333. [Google Scholar]
- Stewart, A.; Wong, K.; Cachat, J.; Gaikwad, S.; Kyzar, E.; Wu, N.; Hart, P.; Piet, V.; Utterback, E.; Elegante, M.; et al. Zebrafish Models to Study Drug Abuse-Related Phenotypes. Rev. Neurosci. 2011, 22, 95–105. [Google Scholar] [CrossRef]
- Becker, T.S.; Rinkwitz, S. Zebrafish as a Genomics Model for Human Neurological and Polygenic Disorders. Dev. Neurobiol. 2012, 72, 415–428. [Google Scholar] [CrossRef]
- Gaikwad, S.; Stewart, A.; Hart, P.; Wong, K.; Piet, V.; Cachat, J.; Kalueff, A.V. Acute Stress Disrupts Performance of Zebrafish in the Cued and Spatial Memory Tests: The Utility of Fish Models to Study Stress-Memory Interplay. Behav. Process. 2011, 87, 224–230. [Google Scholar] [CrossRef]
- Ninkovic, J.; Bally-Cuif, L. The Zebrafish as a Model System for Assessing the Reinforcing Properties of Drugs of Abuse. Methods 2006, 39, 262–274. [Google Scholar] [CrossRef]
- Rey, S.; Huntingford, F.A.; Boltaña, S.; Vargas, R.; Knowles, T.G.; Mackenzie, S. Fish Can Show Emotional Fever: Stress-Induced Hyperthermia in Zebrafish. Proceedings. Biol. Sci. 2015, 282, 658–667. [Google Scholar] [CrossRef] [Green Version]
- Kalueff, A.V.; Gebhardt, M.; Stewart, A.M.; Cachat, J.M.; Brimmer, M.; Chawla, J.S.; Craddock, C.; Kyzar, E.J.; Roth, A.; Landsman, S.; et al. Towards a Comprehensive Catalog of Zebrafish Behavior 1.0 and Beyond. Zebrafish 2013, 10, 70–86. [Google Scholar]
- Jesuthasan, S. Fear, Anxiety, and Control in the Zebrafish. Dev. Neurobiol. 2012, 72, 395–403. [Google Scholar] [CrossRef]
- Avdesh, A.; Martin-Iverson, M.T.; Mondal, A.; Chen, M.; Askraba, S.; Morgan, N.; Lardelli, M.; Groth, D.M.; Verdile, G.; Martins, R.N. Evaluation of Color Preference in Zebrafish for Learning and Memory. J. Alzheimer’s Dis. 2012, 28, 459–469. [Google Scholar] [CrossRef]
- Cerutti, D.; Levin, E. Behavioral Neuroscience of Zebrafish, Methods of Behavior Analysis in Neuroscience, 2nd ed.Taylor & Francis Group LLC: Oxfordshire, UK, 2008; pp. 293–310. ISBN 9781420052343. [Google Scholar]
- Lindeyer, C.M.; Reader, S.M. Social Learning of Escape Routes in Zebrafish and the Stability of Behavioural Traditions. Anim. Behav. 2010, 79, 827–834. [Google Scholar] [CrossRef]
- Zelikowsky, M.; Hui, M.; Karigo, T.; Choe, A.; Yang, B.; Blanco, M.R.; Beadle, K.; Gradinaru, V.; Deverman, B.E.; Anderson, D.J. The Neuropeptide Tac2 Controls a Distributed Brain State Induced by Chronic Social Isolation Stress. Cell 2018, 173, 1265–1279.e19. [Google Scholar] [CrossRef] [Green Version]
- Cacioppo, J.T.; Cacioppo, S.; Capitanio, J.P.; Cole, S.W. The Neuroendocrinology of Social Isolation. Annu. Rev. Psychol. 2015, 66, 733–767. [Google Scholar] [CrossRef] [Green Version]
- Cacioppo, J.T.; Hawkley, L.C. Social Isolation and Health, with an Emphasis on Underlying Mechanisms. Perspect. Biol. Med. 2003, 46, S39–S52. [Google Scholar] [CrossRef]
- Borbély, A.A.; Baumann, F.; Brandeis, D.; Strauch, I.; Lehmann, D. Sleep Deprivation: Effect on Sleep Stages and EEG Power Density in Man. Electroencephalogr. Clin. Neurophysiol. 1981, 51, 483–493. [Google Scholar] [CrossRef]
- Stewart, A.M.; Gaikwad, S.; Kyzar, E.; Kalueff, A.V. Understanding Spatio-Temporal Strategies of Adult Zebrafish Exploration in the Open Field Test. Brain Res. 2012, 1451, 44–52. [Google Scholar] [CrossRef]
- Lopez-Luna, J.; Al-Jubouri, Q.; Al-Nuaimy, W.; Sneddon, L.U. Impact of Stress, Fear and Anxiety on the Nociceptive Responses of Larval Zebrafish. PLoS ONE 2017, 12, e0181010. [Google Scholar] [CrossRef] [Green Version]
- Amo, R.; Aizawa, H.; Takahoko, M.; Kobayashi, M.; Takahashi, R.; Aoki, T.; Okamoto, H. Identification of the Zebrafish Ventral Habenula as a Homolog of the Mammalian Lateral Habenula. J. Neurosci. 2010, 30, 1566–1574. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.; Mathuru, A.S.; Teh, C.; Kibat, C.; Korzh, V.; Penney, T.B.; Jesuthasan, S. The Habenula Prevents Helpless Behavior in Larval Zebrafish. Curr. Biol. 2010, 20, 2211–2216. [Google Scholar] [CrossRef] [Green Version]
- Agetsuma, M.; Aizawa, H.; Aoki, T.; Nakayama, R.; Takahoko, M.; Goto, M.; Sassa, T.; Amo, R.; Shiraki, T.; Kawakami, K.; et al. The Habenula Is Crucial for Experience-Dependent Modification of Fear Responses in Zebrafish. Nat. Neurosci. 2010, 13, 1354–1356. [Google Scholar] [CrossRef]
- Aggarwal, N.T.; Wilson, R.S.; Beck, T.L.; Bienias, J.L.; Bennett, D.A. Mild Cognitive Impairment in Different Functional Domains and Incident Alzheimer’s Disease. J. Neurol. Neurosurg. Psychiatry 2005, 76, 1479–1484. [Google Scholar] [CrossRef]
- Avdesh, A.; Chen, M.; Martin-Iverson, M.T.; Mondal, A.; Ong, D.; Rainey-Smith, S.; Taddei, K.; Lardelli, M.; Groth, D.M.; Verdile, G.; et al. Regular Care and Maintenance of a Zebrafish (Danio Rerio) Laboratory: An Introduction. J. Vis. Exp. 2012, 2012, e4196. [Google Scholar] [CrossRef] [Green Version]
- Egan, R.J.; Bergner, C.L.; Hart, P.C.; Cachat, J.M.; Canavello, P.R.; Elegante, M.F.; Elkhayat, S.I.; Bartels, B.K.; Tien, A.K.; Tien, D.H.; et al. Understanding Behavioral and Physiological Phenotypes of Stress and Anxiety in Zebrafish. Behav. Brain Res. 2009, 205, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Sison, M.; Gerlai, R. Behavioral Performance Altering Effects of MK-801 in Zebrafish (Danio Rerio). Behav. Brain Res. 2011, 220, 331–337. [Google Scholar] [CrossRef] [Green Version]
- Brown, D. Combining Computational Physics with Video Analysis in Tracker. 2007. Available online: https://www.compadre.org/osp/items/detail.cfm?ID=9686 (accessed on 6 October 2022).
- Wee, L.K.; Tat, L.L. Video Analysis and Modeling Tool for Physics Education: A workshop for Redesigning Pedagogy. arXiv: Physics Education. 2012. Available online: https://www.semanticscholar.org/paper/Video-Analysis-and-Modeling-Tool-for-Physics-A-for-Wee-Lee/b86be2b53e910bd44f41d7c08a48ff1ecfd5b64c (accessed on 6 October 2022).
- Sivaji, K.; Kannan, R.R. Polysorbate 80 Coated Gold Nanoparticle as a Drug Carrier for Brain Targeting in Zebrafish Model. J. Clust. Sci. 2019, 30, 897–906. [Google Scholar] [CrossRef]
- Burne, T.; Scott, E.; van Swinderen, B.; Hilliard, M.; Reinhard, J.; Claudianos, C.; Eyles, D.; McGrath, J. Big Ideas for Small Brains: What Can Psychiatry Learn from Worms, Flies, Bees and Fish? Mol. Psychiatry 2011, 16, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Gerlai, R. High-Throughput Behavioral Screens: The First Step towards Finding Genes Involved in Vertebrate Brain Function Using Zebrafish. Molecules 2010, 15, 2609–2622. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Stewart, A.M.; Gerlai, R. Zebrafish as an Emerging Model for Studying Complex Brain Disorders. Trends Pharmacol. Sci. 2014, 35, 63–75. [Google Scholar]
- Piato, L.; Maria, E.; Rosemberg, D.B.; Rico, E.P.; Mussulini, B.H.M.; Bonan, C.D.; Dias, R.D.; Blaser, R.E.; Souza, D.O.; De, D.L. Differences in Spatio-Temporal Behavior of Zebrafish in the Open Tank Paradigm after a Short-Period Confinement into Dark and Bright Environments. PLoS ONE 2011, 6, e19397. [Google Scholar] [CrossRef] [Green Version]
- Baraban, S.C.; Taylor, M.R.; Castro, P.A.; Baier, H. Pentylenetetrazole Induced Changes in Zebrafish Behavior, Neural Activity and c-Fos Expression. Neuroscience 2005, 131, 759–768. [Google Scholar] [CrossRef]
- Bass, S.L.S.; Gerlai, R. Zebrafish (Danio Rerio) Responds Differentially to Stimulus Fish: The Effects of Sympatric and Allopatric Predators and Harmless Fish. Behav. Brain Res. 2008, 186, 107–117. [Google Scholar] [CrossRef]
- Barcellos, L.J.G.; Ritter, F.; Kreutz, L.C.; Quevedo, R.M.; da Silva, L.B.; Bedin, A.C.; Finco, J.; Cericato, L. Whole-Body Cortisol Increases after Direct and Visual Contact with a Predator in Zebrafish, Danio Rerio. Aquaculture 2007, 272, 774–778. [Google Scholar] [CrossRef]
- Levin, E.D.; Bencan, Z.; Cerutti, D.T. Anxiolytic Effects of Nicotine in Zebrafish. Physiol. Behav. 2007, 90, 54–58. [Google Scholar] [CrossRef]
- Champagne, D.L.; Hoefnagels, C.C.M.; de Kloet, R.E.; Richardson, M.K. Translating Rodent Behavioral Repertoire to Zebrafish (Danio Rerio): Relevance for Stress Research. Behav. Brain Res. 2010, 214, 332–342. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Behaviour Category | Behaviour Definition | Description |
|---|---|---|
| Anxiety | Freezing Bout | A complete cessation of movement (except for gills and eyes) by the fish, resulting from high stress/anxiety or as part of the submissive behavior. |
| Erratic movement/Burst swimming | Complex behavior characterized by sharp changes in direction or velocity and repeated rapid darting. | |
| Zig-zagging | The direction of movement changes in a seemingly alternating (zig-zag-like) manner. | |
| Darting | A single fast acceleration in one direction. | |
| Escape | Alarm reaction | An adaptive escape reaction which serves as an anti-predatory response exhibited in the context of fear-inducing stimulation; typically characterized by increased speed of movement and rapid directional changes, a response set that is often referred to as erratic movement (see also zig-zagging). The alarm reaction may also include freezing. |
| Turn | A simple change in swimming direction. | |
| Startled | A quick startle response in which the fish body first curves to form a C-shape, and then the fish propels itself away at an angle from its previous position using a fast swim. | |
| Escape | The ability of a zebrafish to escape from the confined space. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wilson Alphonse, C.R.; Rajaretinam, R.K. Habituation and Behavioural Response of Confinement-Induced Anxiety Conditions in a Zebrafish Model. Appl. Biosci. 2022, 1, 315-323. https://doi.org/10.3390/applbiosci1030020
Wilson Alphonse CR, Rajaretinam RK. Habituation and Behavioural Response of Confinement-Induced Anxiety Conditions in a Zebrafish Model. Applied Biosciences. 2022; 1(3):315-323. https://doi.org/10.3390/applbiosci1030020
Chicago/Turabian StyleWilson Alphonse, Carlton Ranjith, and Rajesh Kannan Rajaretinam. 2022. "Habituation and Behavioural Response of Confinement-Induced Anxiety Conditions in a Zebrafish Model" Applied Biosciences 1, no. 3: 315-323. https://doi.org/10.3390/applbiosci1030020