

Both Acute and Consecutive Days of Formoterol Stimulation Influence Myogenic, Mitochondrial, and myomiR Gene Expression in Human Skeletal Muscle Cells
 ,
,
Abstract
:1. Introduction
2. Results
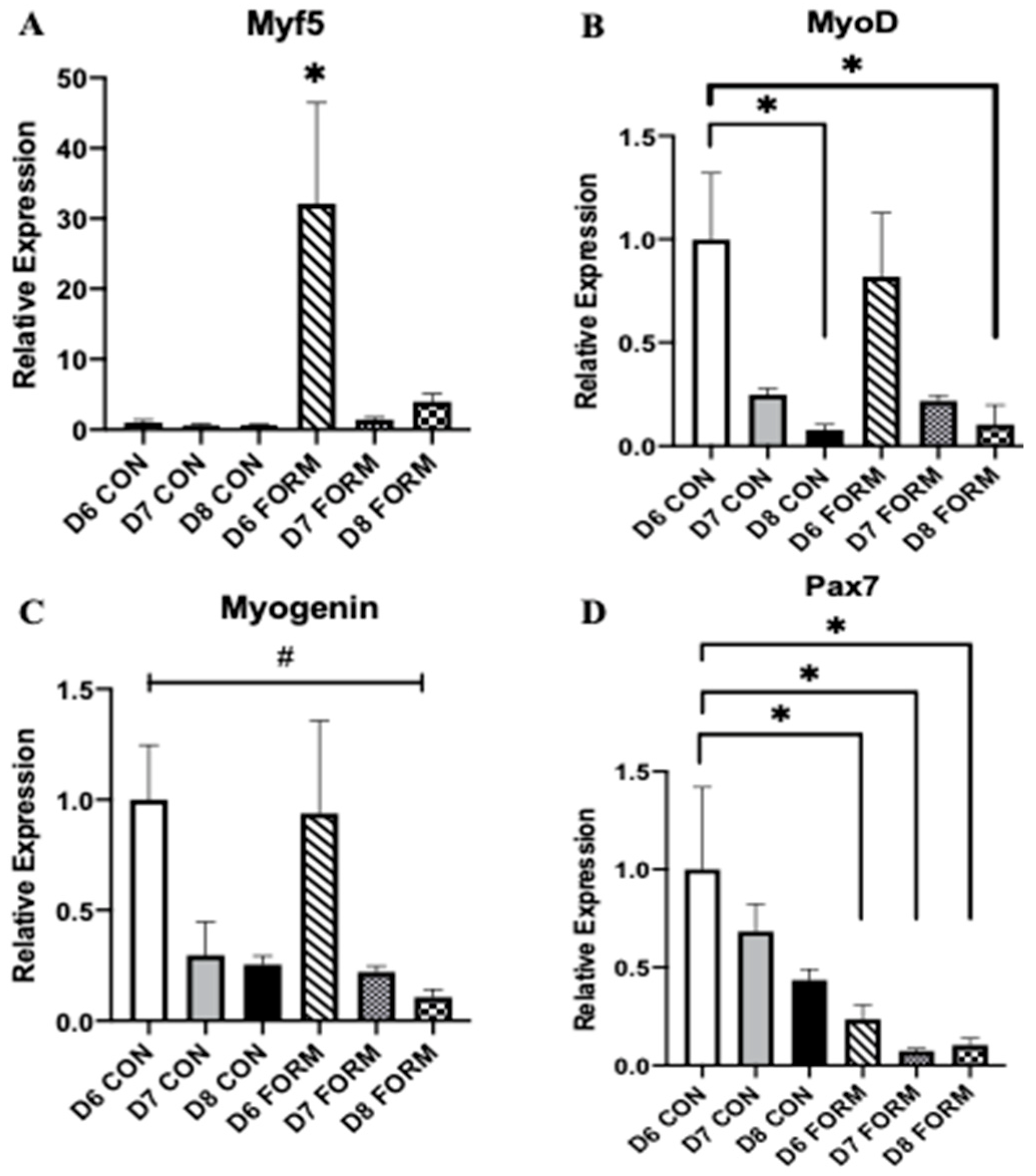
2.1. Formoterol Stimulates Gene Expression of Myogenic Regulatory Factors
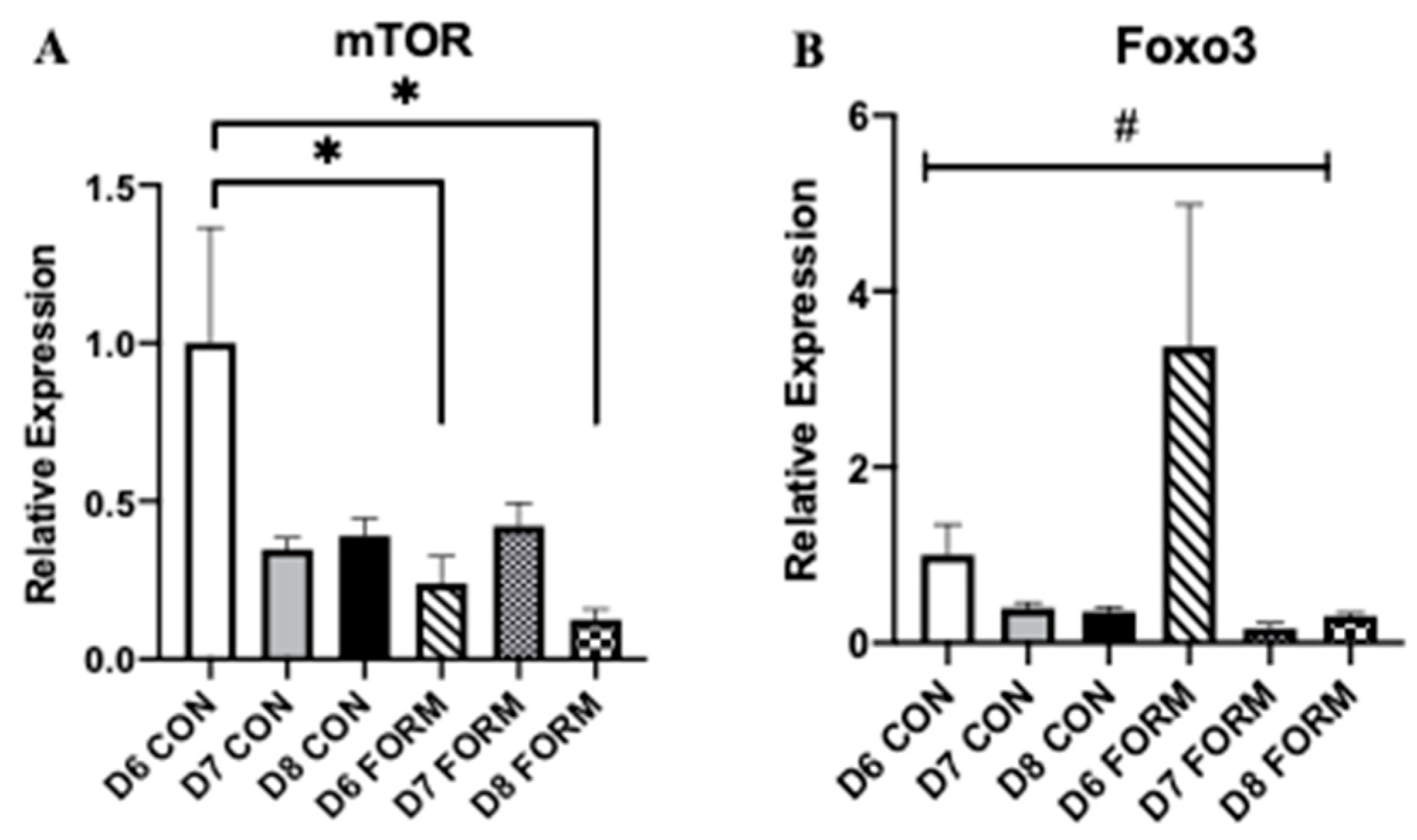
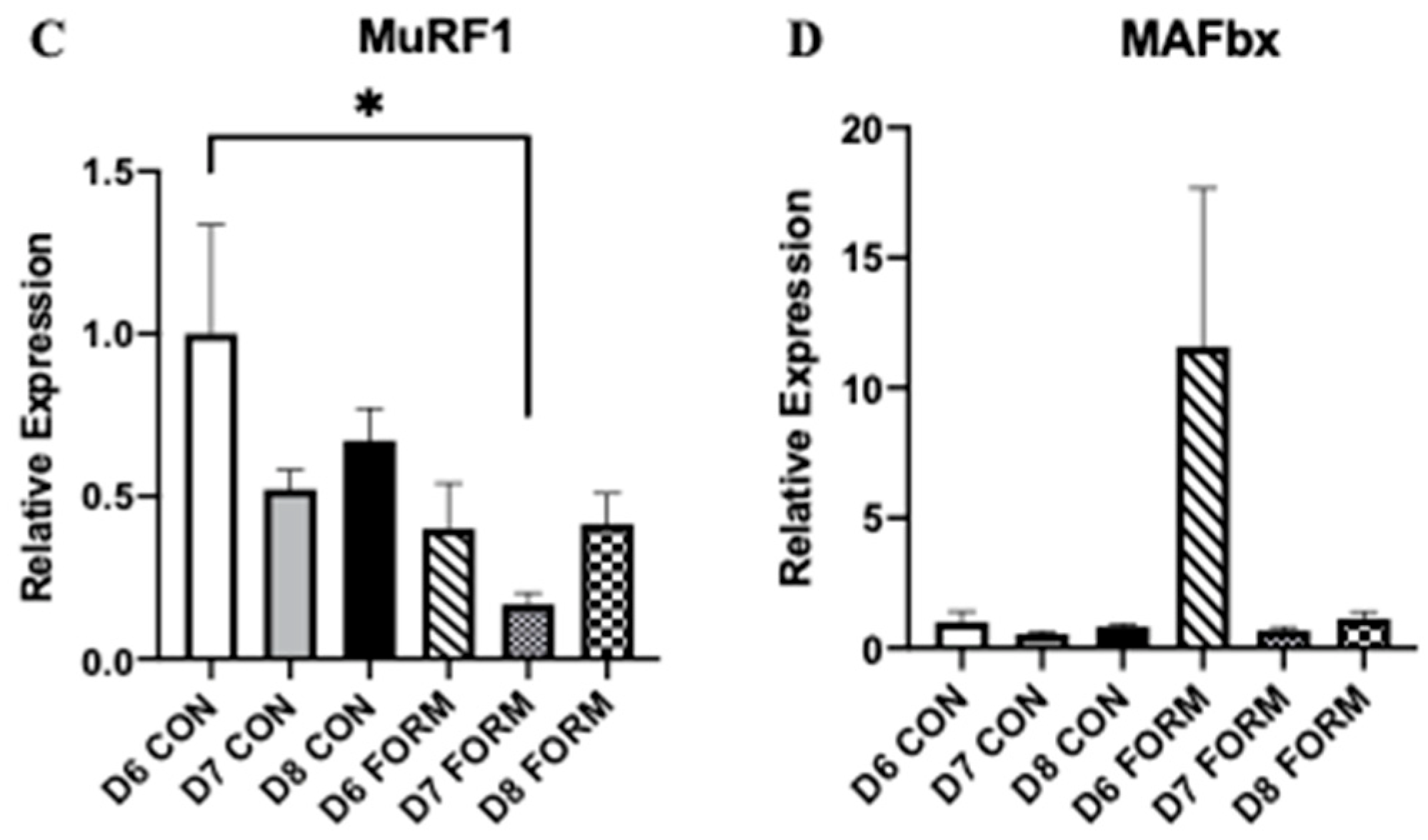
2.2. Formoterol Influences the Expression of Regulators of Skeletal Muscle Mass
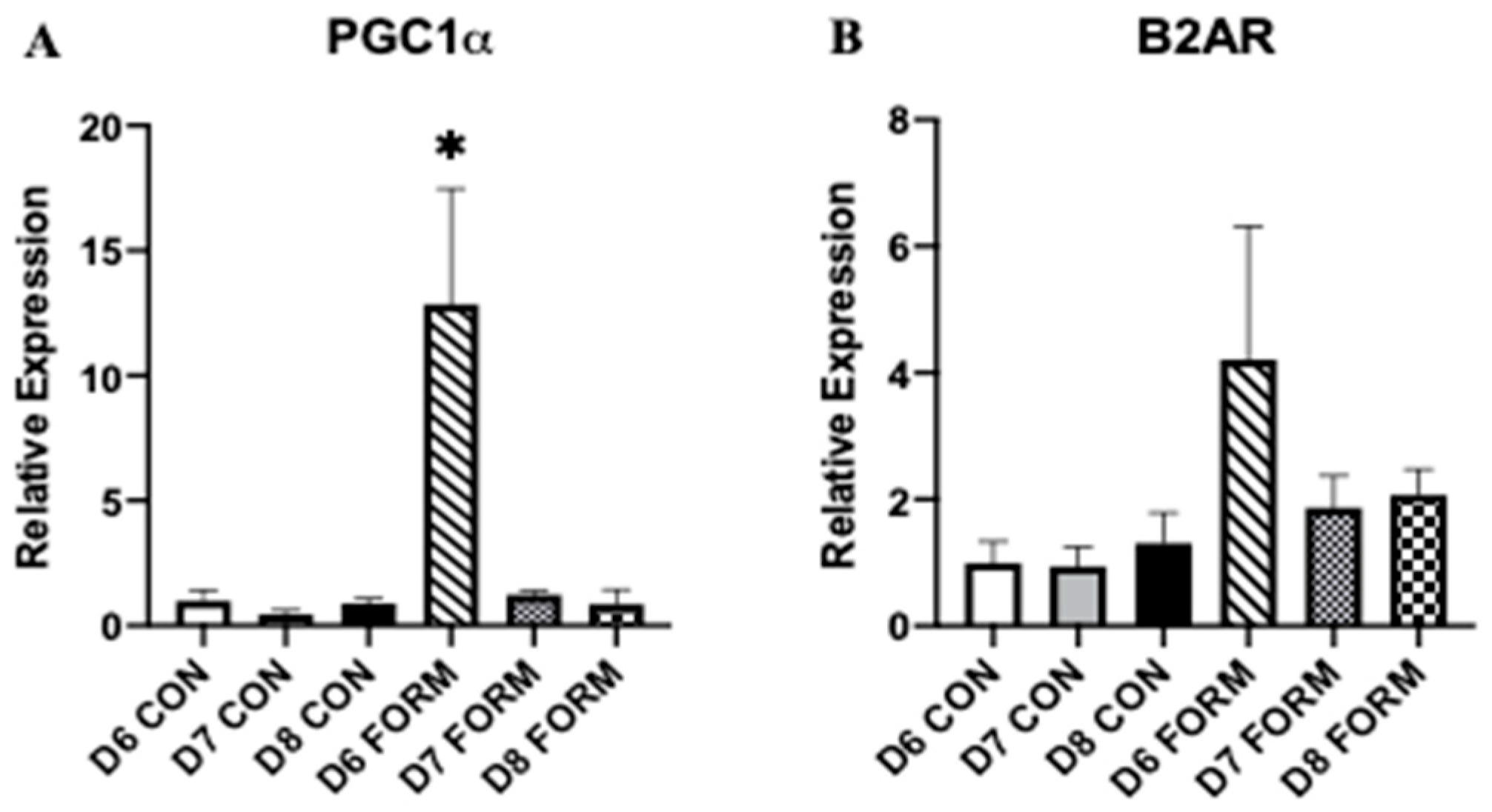
2.3. Formoterol’s Effects on Gene Expression Related to Mitochondrial Biogenesis and Beta-2 Adrenergic Receptors
2.4. Formoterol Stimulates and Influences the Gene Expression of miR-206
3. Discussion
4. Materials and Methods
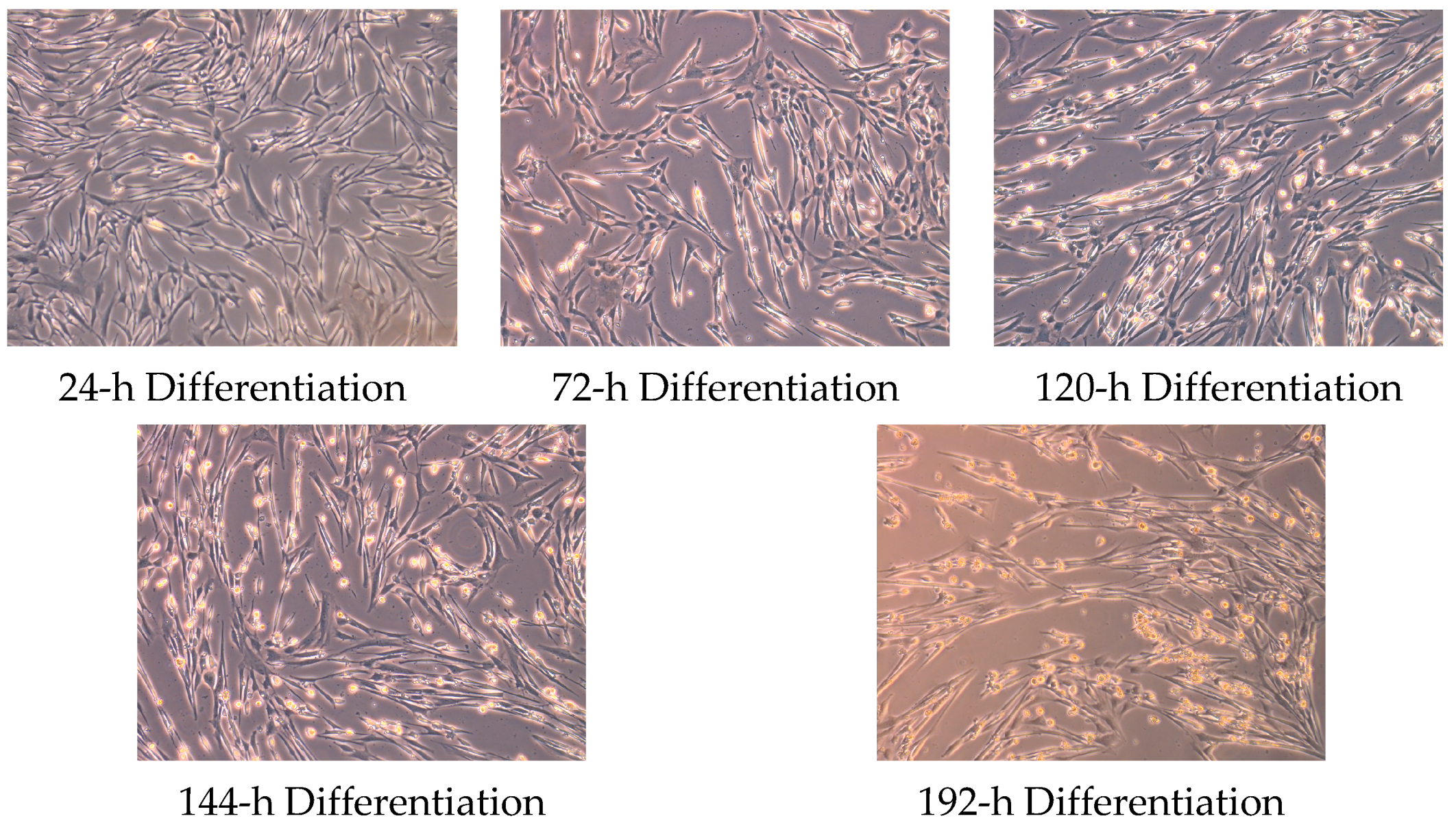
4.1. Cell Culture and Myotube Formation
4.2. Exercise Mimetic
4.3. Experimental Conditions
4.4. RNA Extraction
4.5. Gene Expression of Markers Related to Myogenesis, Muscle Growth, and Mitochondrial Actvitiy
4.6. Gene Expression of miRNA
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoppeler, H. Molecular networks in skeletal muscle plasticity. J. Exp. Biol. 2016, 219, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Kuang, S.; Rudnicki, M.A. The emerging biology of satellite cells and their therapeutic potential. Trends Mol. Med. 2008, 14, 82–91. [Google Scholar] [CrossRef]
- Bazgir, B.; Fathi, R.; Valojerdi, M.R.; Mozdziak, P.; Asgari, A. Satellite cells contribution to exercise mediated muscle hypertrophy and repair. Cell J. 2017, 18, 473. [Google Scholar] [CrossRef]
- Brook, M.S.; Wilkinson, D.J.; Smith, K.; Atherton, P.J. It’s not just about protein turnover: The role of ribosomal biogenesis and satellite cells in the regulation of skeletal muscle hypertrophy. Eur. J. Sport Sci. 2019, 19, 952–963. [Google Scholar] [CrossRef]
- Asfour, H.A.; Allouh, M.Z.; Said, R.S. Myogenic regulatory factors: The orchestrators of myogenesis after 30 years of discovery. Exp. Biol. Med. 2018, 243, 118–128. [Google Scholar] [CrossRef]
- Zammit, P.S. Function of the Myogenic Regulatory Factors Myf5, MyoD, Myogenin and MRF4 in Skeletal Muscle, Satellite Cells and Regenerative Myogenesis. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2017; Volume 72, pp. 19–32. [Google Scholar] [CrossRef]
- Kirby, T.J.; Chaillou, T.; McCarthy, J.J. The role of microRNAs in skeletal muscle health and disease. Front. Biosci. Landmark Ed. 2015, 20, 37. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.H. MicroRNA in myogenesis and muscle atrophy. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 258. [Google Scholar] [CrossRef] [Green Version]
- Giagnorio, E.; Malacarne, C.; Mantegazza, R.; Bonanno, S.; Marcuzzo, S. MyomiRs and their multifaceted regulatory roles in muscle homeostasis and amyotrophic lateral sclerosis. J. Cell Sci. 2021, 134, jcs258349. [Google Scholar] [CrossRef]
- Horak, M.; Novak, J.; Bienertova-Vasku, J. Muscle-specific microRNAs in skeletal muscle development. Dev. Biol. 2016, 410, 1–13. [Google Scholar] [CrossRef]
- Small, E.M.; O’Rourke, J.R.; Moresi, V.; Sutherland, L.B.; McAnally, J.; Gerard, R.D.; Richardson, J.A.; Olson, E.N. Regulation of PI3-kinase/Akt signaling by muscle-enriched microRNA-486. Proc. Natl. Acad. Sci. USA 2010, 107, 4218–4223. [Google Scholar] [CrossRef] [Green Version]
- Popov, D.V.; Lysenko, E.A.; Bachinin, A.V.; Miller, T.F.; Kurochkina, N.S.; Kravchenko, I.V.; Furalyov, V.A.; Vinogradova, O.L. Influence of resistance exercise intensity and metabolic stress on anabolic signaling and expression of myogenic genes in skeletal muscle. Muscle Nerve 2015, 51, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Joanisse, S.; Lim, C.; McKendry, J.; Mcleod, J.C.; Stokes, T.; Phillips, S.M. Recent advances in understanding resistance exercise training-induced skeletal muscle hypertrophy in humans. F1000Research 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Kirby, T.J.; McCarthy, J.J. MicroRNAs in skeletal muscle biology and exercise adaptation. Free. Radic. Biol. Med. 2013, 64, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Silva, G.J.; Bye, A.; El Azzouzi, H.; Wisløff, U. MicroRNAs as important regulators of exercise adaptation. Prog. Cardiovasc. Dis. 2017, 60, 130–151. [Google Scholar] [CrossRef] [PubMed]
- Hawley, J.A.; Joyner, M.J.; Green, D.J. Mimicking exercise: What matters most and where to next? J. Physiol. 2021, 599, 791–802. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Yi, S.J.; Lee, H.; Kim, J.H.; Oh, M.J.; Song, E.J.; Kim, K.; Jhun, B.H. β2-Adrenergic receptor (β2-AR) agonist formoterol suppresses differentiation of L6 myogenic cells by blocking PI3K–AKT pathway. Anim. Cells Syst. 2019, 23, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Sabourin, L.A.; Rudnicki, M.A. The molecular regulation of myogenesis. Clin. Genet. 2000, 57, 16–25. [Google Scholar] [CrossRef]
- Bentzinger, C.F.; Wang, Y.X.; Rudnicki, M.A. Building muscle: Molecular regulation of myogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008342. [Google Scholar] [CrossRef] [Green Version]
- Zanou, N.; Gailly, P. Skeletal muscle hypertrophy and regeneration: Interplay between the myogenic regulatory factors (MRFs) and insulin-like growth factors (IGFs) pathways. Cell. Mol. Life Sci. 2013, 70, 4117–4130. [Google Scholar] [CrossRef]
- Xu, M.; Chen, X.; Chen, D.; Yu, B.; Li, M.; He, J.; Huang, Z. Regulation of skeletal myogenesis by microRNAs. J. Cell. Physiol. 2020, 235, 87–104. [Google Scholar] [CrossRef]
- Pearen, M.A.; Ryall, J.G.; Lynch, G.S.; Muscat, G.E. Expression profiling of skeletal muscle following acute and chronic β 2-adrenergic stimulation: Implications for hypertrophy, metabolism and circadian rhythm. BMC Genom. 2009, 10, 448. [Google Scholar] [CrossRef] [Green Version]
- Bodine, S.C. mTOR signaling and the molecular adaptation to resistance exercise. Med. Sci. Sport. Exerc. 2006, 38, 1950–1957. [Google Scholar] [CrossRef]
- Yoon, M.S. mTOR as a key regulator in maintaining skeletal muscle mass. Front. Physiol. 2017, 8, 788. [Google Scholar] [CrossRef] [Green Version]
- Joassard, O.R.; Amirouche, A.; Gallot, Y.S.; Desgeorges, M.M.; Castells, J.; Durieux, A.C.; Berthon, P.; Freyssenet, D.G. Regulation of Akt-mTOR, ubiquitin-proteasome and autophagy-lysosome pathways in response to formoterol administration in rat skeletal muscle. Int. J. Biochem. Cell Biol. 2013, 45, 2444–2455. [Google Scholar] [CrossRef]
- Salazar-Degracia, A.; Busquets, S.; Argilés, J.M.; Bargalló-Gispert, N.; López-Soriano, F.J.; Barreiro, E. Effects of the beta2 agonist formoterol on atrophy signaling, autophagy, and muscle phenotype in respiratory and limb muscles of rats with cancer-induced cachexia. Biochimie 2018, 149, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Lynch, G.S.; Ryall, J.G. Role of B-adrenergic signaling in skeletal muscle: Implications for muscle wasting and disease. Physiol. Rev. 2008, 88, 729–767. [Google Scholar] [CrossRef] [Green Version]
- Margolis, L.M.; Rivas, D.A. Potential role of microRNA in the anabolic capacity of skeletal muscle with aging. Exerc. Sport Sci. Rev. 2018, 46, 86. [Google Scholar] [CrossRef]
- Duplanty, A.A.; Siggins, R.W.; Allerton, T.; Simon, L.; Molina, P.E. Myoblast mitochondrial respiration is decreased in chronic binge alcohol administered simian immunodeficiency virus-infected antiretroviral-treated rhesus macaques. Physiol. Rep. 2018, 6, e13625. [Google Scholar] [CrossRef] [Green Version]
- Wills, L.P.; Trager, R.E.; Beeson, G.C.; Lindsey, C.C.; Peterson, Y.K.; Beeson, C.C.; Schnellmann, R.G. The β2-adrenoceptor agonist formoterol stimulates mitochondrial biogenesis. J. Pharmacol. Exp. Ther. 2012, 342, 106–118. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Sequence (5′-3′) |
|---|---|---|
| Myf5 | Forward | GCTTCTAGTTCCAGGCTTATC |
| Reverse | GCCTTCTTCTTCCTGTGTATTA | |
| MyoD | Forward Reverse | CACAACGGACGACTTCTATG GTGCTCTTCGGGTTTCAG |
| Myogenin | Forward Reverse | CCCTGAATTGAGAGAGAAGAAG CGGATGGCAGCTTTACAA |
| Pax7 | Forward Reverse | GAAGACGAAGAAGACGGAAAG GGACACTTCCAAAGGGAATC |
| mTOR | Forward Reverse | GGACTACAGGGAGAAGAAGA CATCAGAGTCAAGTGGTCATAG |
| Foxo3 | ForwardReverse | CCACCCTTGGCCTCTAAATAA GGTAACAGGTATCAGGTTCTGG |
| MuRF1 | Forward Reverse | CAGCTGGACAAGTCCACAAA GCGTCTGCTATGTGCTCTAAAT |
| MAFbx | Forward Reverse | GCATGCCCTTGGCAAATAAG ATGTGGGTTGTGTGCTATTGA |
| PGC-1α | Forward Reverse | TCTCTCTCTCTCTCTCTCTCT CATGGGTGTCAGGATTAAGG |
| B2AR | Forward Reverse | CCTGCTGACCAAGAATAAGG GCAGGTCTCATTGGCATAG |
| RPS13 | Forward Reverse | GCATCTTGAGAGGAACAGAAA AGGACTCGCTTGGTCTTAT |
| hsa-miR-206 | Mature Sequence | UGGAAUGUAAGGAAGUGUGUGG |
| hsa-miR-486 | Mature Sequence | UCCUGUACUGAGCUGCCCCGAG |
| hsa-miR-92a | Mature Sequence | UAUUGCACUUGUCCCGGCCUGU |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gordon, R.A.; Zumbro, E.L.; Guerin, G.D.; Sokoloski, M.L.; Ben-Ezra, V.; Brower, C.S.; Rigby, R.B.; Duplanty, A.A. Both Acute and Consecutive Days of Formoterol Stimulation Influence Myogenic, Mitochondrial, and myomiR Gene Expression in Human Skeletal Muscle Cells. Muscles 2023, 2, 86-96. https://doi.org/10.3390/muscles2010008
Gordon RA, Zumbro EL, Guerin GD, Sokoloski ML, Ben-Ezra V, Brower CS, Rigby RB, Duplanty AA. Both Acute and Consecutive Days of Formoterol Stimulation Influence Myogenic, Mitochondrial, and myomiR Gene Expression in Human Skeletal Muscle Cells. Muscles. 2023; 2(1):86-96. https://doi.org/10.3390/muscles2010008
Chicago/Turabian StyleGordon, Ryan A., Emily L. Zumbro, Gena D. Guerin, Matthew L. Sokoloski, Vic Ben-Ezra, Christopher S. Brower, Rhett B. Rigby, and Anthony A. Duplanty. 2023. "Both Acute and Consecutive Days of Formoterol Stimulation Influence Myogenic, Mitochondrial, and myomiR Gene Expression in Human Skeletal Muscle Cells" Muscles 2, no. 1: 86-96. https://doi.org/10.3390/muscles2010008