
Canada Lynx (Lynx canadensis) as Potential Reservoirs and Sentinels of Toxoplasma gondii in Northern Canada
 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
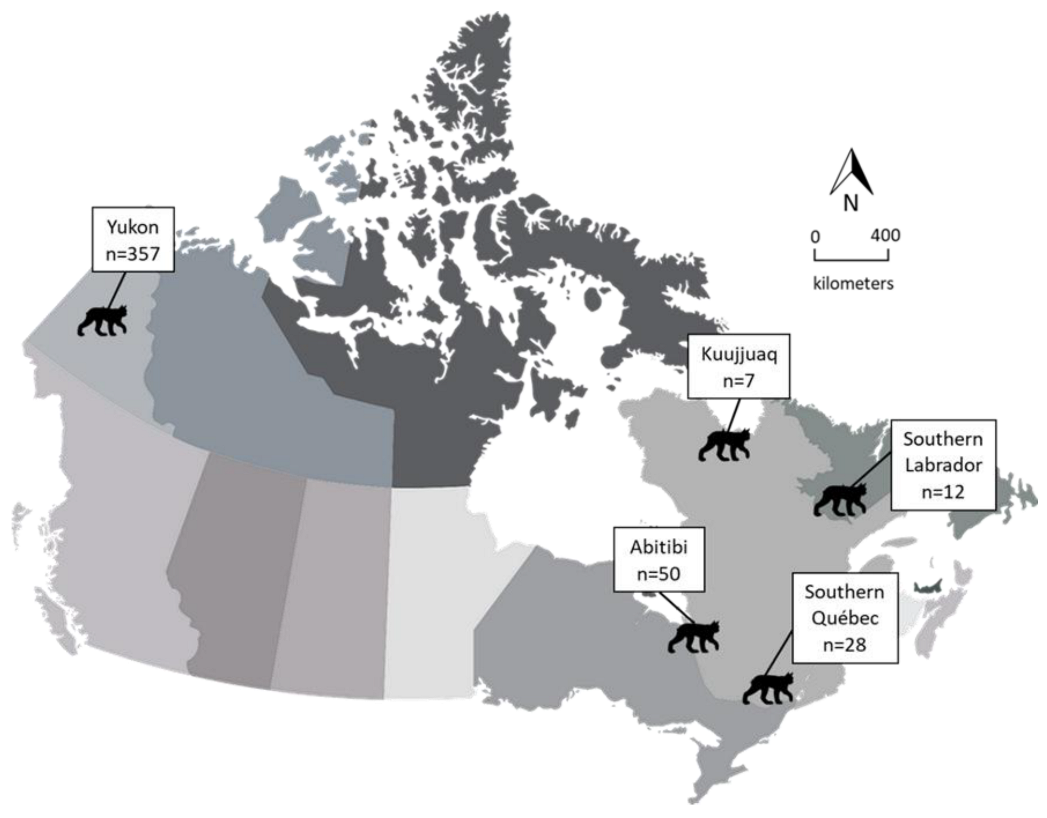
2.1. Sample Locations and Lynx Sampling
2.2. Serological Analysis
2.3. Molecular Analysis
2.3.1. Detection of DNA of T. gondii in Tissue Samples
2.3.2. Detection of DNA of T. gondii in Fecal Samples
2.4. Fecal Flotation
2.5. Statistical Analysis
3. Results
3.1. Descriptive Analysis
3.2. Cut-off Values and Best-Performing Serological Assay
3.3. Association between serological and molecular tests
3.4. Prevalence and Potential Risk Factors Associated with T. gondii
3.5. Quantification of Infection Intensity and Risk Factors
3.6. CoproPCR and DNA Sequencing
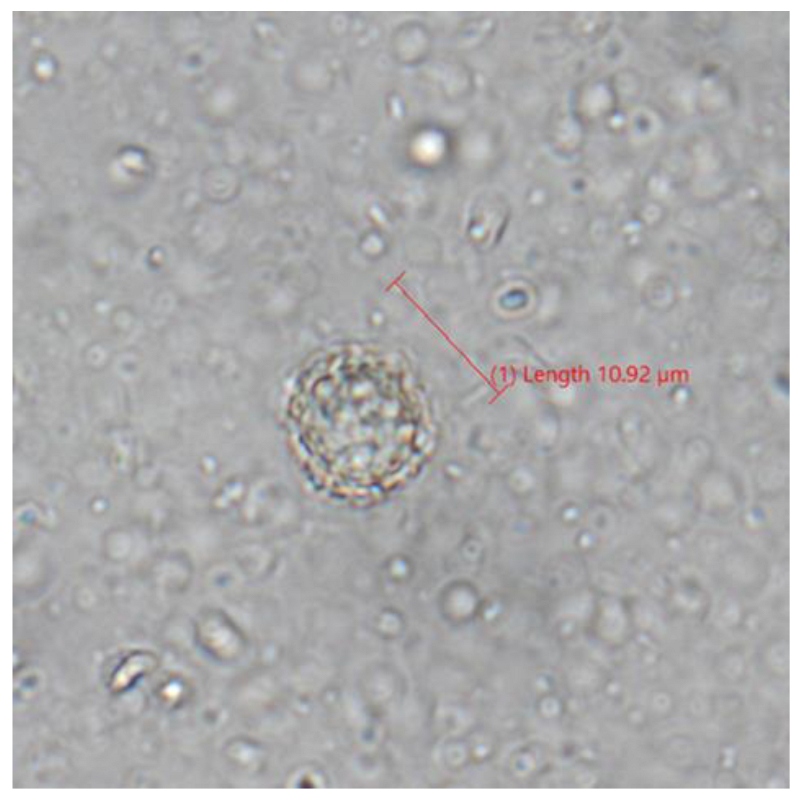
3.7. Fecal Flotation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tenter, A.M.; Heckeroth, A.R.; Weiss, L.M. Toxoplasma gondii: From animals to humans. Int. J. Parasitol. 2000, 30, 1217–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubey, J.P. Toxoplasmosis of Animals and Humans, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2010; p. 313. [Google Scholar]
- Miller, N.L.; Frenkel, J.K.; Dubey, J.P. Oral infections with Toxoplasma cysts and oocysts in felines, other mammals, and in birds. J. Parasitol. 1972, 58, 928–937. [Google Scholar] [CrossRef] [PubMed]
- Bowie, W.R.; King, A.S.; Werker, D.H.; Isaac-Renton, J.L.; Bell, A.; Eng, S.B.; Marion, S.A. Outbreak of toxoplasmosis associated with municipal drinking water. The BC Toxoplasma Investigation Team. Lancet 1997, 350, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Aramini, J.J.; Stephen, C.; Dubey, J.P.; Engelstoft, C.; Schwantje, H.; Ribble, C.S. Potential contamination of drinking water with Toxoplasma gondii oocysts. Epidemiol. Infect. 1999, 122, 305–315. [Google Scholar] [CrossRef] [PubMed]
- García-Bocanegra, I.; Dubey, J.P.; Martínez, F.; Vargas, A.; Cabezón, O.; Zorrilla, I.; Arenas, A.; Almería, S. Factors affecting seroprevalence of Toxoplasma gondii in the endangered Iberian lynx (Lynx pardinus). Vet. Parasitol. 2010, 167, 36–42. [Google Scholar] [CrossRef]
- Hatam-Nahavandi, K.; Calero-Bernal, R.; Rahimi, M.T.; Pagheh, A.S.; Zarean, M.; Dezhkam, A.; Ahmadpour, E. Toxoplasma gondii infection in domestic and wild felids as public health concerns: A systematic review and meta-analysis. Sci. Rep. 2021, 11, 9509. [Google Scholar] [CrossRef]
- Mowat, G.; Poole, K.G.; O’Donoghue, M. Ecology of lynx in northern Canada and Alaska. In Ecology and Conservation of Lynx in the United States; Ruggerio, L.F., Aubry, K.B., Buskirk, S.W., Koehler, G.M., Krebs, C.J., McKelvey, K.S., Squires, J.R., Eds.; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2000; pp. 265–306. [Google Scholar]
- Peers, M.J.L.; Konkolics, S.M.; Lamb, C.T.; Majchrzak, Y.N.; Menzies, A.K.; Studd, E.K.; Boonstra, R.; Kenney, A.J.; Krebs, C.J.; Martinig, A.R.; et al. Prey availability and ambient temperature influence carrion persistence in the boreal forest. J. Anim. Ecol. 2020, 89, 2156–2167. [Google Scholar] [CrossRef]
- Jung, T. Raven (Corvus corax) as a novel food item for lynx (Lynx canadensis). Can. Field Nat. 2021, 136, 17–19. [Google Scholar] [CrossRef]
- Zarnke, R.L.; Dubey, J.P.; Ver Hoef, J.M.; McNay, M.E.; Kwok, O.C. Serologic survey for Toxoplasma gondii in lynx from interior Alaska. J. Wildl. Dis. 2001, 37, 36–38. [Google Scholar] [CrossRef] [Green Version]
- Simon, A.; Bigras Poulin, M.; Rousseau, A.N.; Dubey, J.P.; Ogden, N.H. Spatiotemporal dynamics of Toxoplasma gondii infection in Canadian lynx (Lynx canadensis) in western Quebec, Canada. J. Wildl. Dis. 2013, 49, 39–48. [Google Scholar] [CrossRef]
- Al-Adhami, B.H.; Simard, M.; Hernández-Ortiz, A.; Boireau, C.; Gajadhar, A.A. Development and evaluation of a modified agglutination test for diagnosis of Toxoplasma infection using tachyzoites cultivated in cell culture. Food Waterborne Parasitol. 2016, 2, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Labelle, P.; Dubey, J.P.; Mikaelian, I.; Blanchette, N.; Lafond, R.; St-Onge, S.; Martineau, D. Seroprevalence of antibodies to Toxoplasma gondii in lynx (Lynx canadensis) and bobcats (Lynx rufus) from Quebec, Canada. J. Parasitol. 2001, 87, 1194–1196. [Google Scholar] [CrossRef] [PubMed]
- Lalonde, L.F.; Gajadhar, A.A. Detection and differentiation of coccidian oocysts by real-time PCR and melting curve analysis. J. Parasitol. 2011, 97, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.S.; Kukka, P.M.; Peers, M.J.L.; Schmiegelow, F.K.A.; Boonstra, R.; Boutin, S.; Majchrzak, Y.N. Error in trapper-reported sex of lynx (Lynx canadensis) and wolverine (Gulo gulo): Implications for analyses of harvest records. Eur. J. Wildl. Res. 2020, 66, 52. [Google Scholar] [CrossRef]
- Slough, B.G. Estimating lynx population age ratio with pelt data. Wildl. Soc. Bull. 1996, 24, 495–499. [Google Scholar]
- Bouchard, É.; Sharma, R.; Hernández-Ortiz, A.; Buhler, K.; Al-Adhami, B.; Su, C.; Fenton, H.; Gouin, G.G.; Roth, J.D.; Rodrigues, C.W.; et al. Are foxes (Vulpes spp.) good sentinel species for Toxoplasma gondii in northern Canada? Parasites Vectors 2022, 15, 115. [Google Scholar] [CrossRef]
- Sharma, R.; Parker, S.; Al-Adhami, B.; Bachand, N.; Jenkins, E. Comparison of tissues (heart vs. brain) and serological tests (MAT, ELISA and IFAT) for detection of Toxoplasma gondii in naturally infected wolverines (Gulo gulo) from the Yukon, Canada. Food Waterborne Parasitol. 2019, 15, e00046. [Google Scholar] [CrossRef]
- Opsteegh, M.; Langelaar, M.; Sprong, H.; den Hartog, L.; De Craeye, S.; Bokken, G.; Ajzenberg, D.; Kijlstra, A.; van der Giessen, J. Direct detection and genotyping of Toxoplasma gondii in meat samples using magnetic capture and PCR. Int. J. Food Microbiol. 2010, 139, 193–201. [Google Scholar] [CrossRef]
- Bachand, N.; Ravel, A.; Leighton, P.; Stephen, C.; Ndao, M.; Avard, E.; Jenkins, E. Serological and molecular detection of Toxoplasma gondii in terrestrial and marine wildlife harvested for food in Nunavik, Canada. Parasites Vectors 2019, 12, 155. [Google Scholar] [CrossRef] [Green Version]
- Su, C.; Shwab, E.K.; Zhou, P.; Zhu, X.Q.; Dubey, J.P. Moving towards an integrated approach to molecular detection and identification of Toxoplasma gondii. Parasitology 2010, 137, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Dryden, M.W.; Payne, P.A.; Ridley, R.; Smith, V. Comparison of common fecal flotation techniques for the recovery of parasite eggs and oocysts. Vet. Ther. 2005, 6, 15–28. [Google Scholar]
- Sergeant, E.S.G. Epitools Epidemiological Calculators. Ausvet. 2018. Available online: http://epitools.ausvet.com.au (accessed on 15 August 2022).
- Dohoo, I.R.; Martin, W.; Stryhn, H. Veterinary Epidemiologic Research, 2nd ed.; Atlantic Veterinary College Inc., University of Prince Edward Island: Charlottetown, PE, Canada, 2010; p. 865. [Google Scholar]
- Chester, S. The Arctic Guide: Wildlife of the Far North; Princeton University Press: Princeton, NJ, USA, 2016; pp. 71–72. [Google Scholar]
- Simon, A.; Poulin, M.B.; Rousseau, A.N.; Ogden, N.H. Fate and transport of Toxoplasma gondii oocysts in seasonally snow covered watersheds: A conceptual framework from a melting snowpack to the Canadian arctic coasts. Int. J. Environ. Res. Public Health 2013, 10, 994–1005. [Google Scholar] [CrossRef] [PubMed]
- McDonald, J.C.; Gyorkos, T.W.; Alberton, B.; MacLean, J.D.; Richer, G.; Juranek, D. An outbreak of toxoplasmosis in pregnant women in northern Quebec. J. Infect. Dis. 1990, 161, 769–774. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, M.; Renard, A.; Larivière, S. Lynx canadensis (Carnivora: Felidae). Mamm. Species 2019, 51, 136–154. [Google Scholar] [CrossRef]
- Simon, A.; Rousseau, A.N.; Savary, S.; Bigras-Poulin, M.; Ogden, N.H. Hydrological modelling of Toxoplasma gondii oocysts transport to investigate contaminated snowmelt runoff as a potential source of infection for marine mammals in the Canadian Arctic. J. Environ. Manag. 2013, 127, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Murata, F.H.A.; Cerqueira-Cézar, C.K.; Kwok, O.C.H. Recent epidemiologic and clinical Toxoplasma gondii infections in wild canids and other carnivores: 2009–2020. Vet. Parasitol. 2021, 290, 109337. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, A.T.; Fooks, A.R.; Hayman, D.T.; Horton, D.L.; Muller, T.; Plowright, R.; Peel, A.J.; Bowen, R.; Wood, J.L.; Mills, J.; et al. Deciphering serology to understand the ecology of infectious diseases in wildlife. EcoHealth 2013, 10, 298–313. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Parker, S.; Elkin, B.; Mulders, R.; Branigan, M.; Pongracz, J.; Godson, D.L.; Larter, N.C.; Jenkins, E. Risk factors and prevalence of antibodies for Toxoplasma gondii in diaphragmatic fluid in wolverines (Gulo gulo) from the Northwest Territories, Canada. Food Waterborne Parasitol. 2019, 15, e00056. [Google Scholar] [CrossRef]
- Ferreira, S.C.M.; Torelli, F.; Klein, S.; Fyumagwa, R.; Karesh, W.B.; Hofer, H.; Seeber, F.; East, M.L. Evidence of high exposure to Toxoplasma gondii in free-ranging and captive African carnivores. Int. J. Parasitol. Parasites Wildl. 2019, 8, 111–117. [Google Scholar] [CrossRef]
- Bouchard, É.; Elmore, S.A.; Alisauskas, R.T.; Samelius, G.; Gajadhar, A.A.; Schmidt, K.; Ross, S.; Jenkins, E.J. Transmission dynamics of Toxoplasma gondii in Arctic foxes (Vulpes lagopus): A long-term mark-recapture serologic study at Karrak Lake, Nunavut, Canada. J. Wildl. Dis. 2019, 55, 619–626. [Google Scholar] [CrossRef] [Green Version]
- Miernyk, K.M.; Bruden, D.; Parkinson, A.J.; Hurlburt, D.; Klejka, J.; Berner, J.; Stoddard, R.A.; Handali, S.; Wilkins, P.P.; Kersh, G.J.; et al. Human seroprevalence to 11 zoonotic pathogens in the U. S. Arctic, Alaska. Vector Borne Zoonotic Dis. 2019, 19, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Goyette, S.; Cao, Z.R.; Libman, M.; Ndao, M.; Ward, B.J. Seroprevalence of parasitic zoonoses and their relationship with social factors among the Canadian Inuit in Arctic regions. Diagn. Microbiol. Infect. Dis. 2014, 78, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Elton, C.; Nicholson, M. The ten-year cycle in numbers of the lynx in Canada. J. Anim. Ecol. 1942, 11, 215–244. [Google Scholar] [CrossRef]
- Krebs, C.J.; Boonstra, R.; Boutin, S.; Sinclair, A.R.E. What drives the 10-year cycle of snowshoe hares?: The ten-year cycle of snowshoe hares—One of the most striking features of the boreal forest—Is a product of the interaction between predation and food supplies, as large-scale experiments in the yukon have demonstrated. Bioscience 2001, 51, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Frenkel, J.K.; Ruiz, A.; Chinchilla, M. Soil survival of toxoplasma oocysts in Kansas and Costa Rica. Am. J. Trop. Med. Hyg. 1975, 24, 439–443. [Google Scholar] [CrossRef]
- Peers, M.J.L.; Majchrzak, Y.N.; Konkolics, S.M.; Boonstra, R.; Boutin, S. Scavenging by snowshoe hares (Lepus americanus) in Yukon, Canada. Northwest Nat. 2018, 99, 232–235. [Google Scholar] [CrossRef]
- Koethe, M.; Straubinger, R.K.; Pott, S.; Bangoura, B.; Geuthner, A.C.; Daugschies, A.; Ludewig, M. Quantitative detection of Toxoplasma gondii in tissues of experimentally infected turkeys and in retail turkey products by magnetic-capture PCR. Food Microbiol. 2015, 52, 11–17. [Google Scholar] [CrossRef]
- Gisbert Algaba, I.; Verhaegen, B.; Jennes, M.; Rahman, M.; Coucke, W.; Cox, E.; Dorny, P.; Dierick, K.; De Craeye, S. Pork as a source of transmission of Toxoplasma gondii to humans: A parasite burden study in pig tissues after infection with different strains of Toxoplasma gondii as a function of time and different parasite stages. Int. J. Parasitol. 2018, 48, 555–560. [Google Scholar] [CrossRef]
- Kukka, P.M.; Jung, T.S.; Robitaille, J.-F.; Schmiegelow, F.K.A. Temporal variation in the population characteristics of harvested wolverine (Gulo gulo) in northwestern Canada. Wildl. Res. 2017, 44, 497–503. [Google Scholar] [CrossRef]
- Dubey, J.P.; Velmurugan, G.V.; Rajendran, C.; Yabsley, M.J.; Thomas, N.J.; Beckmen, K.B.; Sinnett, D.; Ruid, D.; Hart, J.; Fair, P.A.; et al. Genetic characterisation of Toxoplasma gondii in wildlife from North America revealed widespread and high prevalence of the fourth clonal type. Int. J. Parasitol. 2011, 41, 1139–1147. [Google Scholar] [CrossRef]
- Bouchard, É.; Jokelainen, P.; Sharma, R.; Fenton, H.; Jenkins, E.J. Toxoplasmosis in northern regions. In Arctic One Health: Challenges for Northern Animals and People; Tryland, M., Ed.; Springer International Publishing: Cham, Switzerland, 2022; pp. 297–314. [Google Scholar]
- Bush, E.; Lemmen, D.S. Canada’s Changing Climate Report; Government of Canada: Ottawa, ON, Canada, 2019; p. 444.



{kind=link}
{kind=link}
| Seroprevalence 1 | Tissue-Prevalence 2 | Prevalence 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Variable | % (95% CI); P/T | OR (95% CI) | p Value | % (95% CI); P/T | OR (95% CI) | p Value | % (95% CI); P/T | OR (95% CI) | p Value |
| Sex Female 4 | 19 (13–26); 27/145 | 16 (11–23); 23/145 | 20 (14–27); 29/145 | ||||||
| Male | 29 (23–35); 62/215 | 1.8 (1.1–3) | 0.029 | 23 (18–29); 49/215 | 1.6 (0.9–2.7) | 0.11 | 30 (24–36); 64/215 | 1.7 (1.0–2.8) | 0.039 |
| Age-class Juvenile 4 | 8 (4–18); 5/60 | 7 (3–16); 4/60 | 8.3 (4–18); 5/60 | ||||||
| Adult | 59 (42–74); 20/34 | 15.7 (5.0–49.2) | <0.001 | 44 (29–61); 15/34 | 11.1 (3.3–37.4) | <0.001 | 59 (42–74); 20/34 | 15.7 (5.0–49.2) | <0.001 |
| BCI Low 4 | 13 (8–21); 12/92 | 11 (6–19); 10/92 | 13 (8–21); 12/92 | ||||||
| Med | 19 (14–26); 36/186 | 1.6 (0.8–3.2) | 14 (10–20); 27/186 | 1.4 (0.6–3.0) | 20 (15–26); 37/186 | 1.7 (0.8–3.4) | |||
| High | 32 (17–52); 8/25 | 3.1 (1.1–8.8) | 0.094 | 20 (9–39); 5/25 | 2.1 (0.6–6.7) | 0.468 | 32 (17–52); 8/25 | 3.1 (1.1–8.8) | 0.091 |
| Region Eastern | 41 (30–52); 28/69 | 32 (22–44); 22/69 | 41 (30–52); 28/69 | ||||||
| Labrador, NL | 33 (14–61); 4/12 | 33 (14–61); 4/12 | 33 (14–61); 4/12 | ||||||
| Abitibi, QC | 36 (24–50); 18/50 | 24 (14–37); 12/50 | 36 (24–50); 18/50 | ||||||
| Kuujjuaq, QC | 86 (49–97); 6/7 | 86 (49–97); 6/7 | 86 (49–97); 6/7 | ||||||
| Region Western 4 Yukon | 20 (16–25); 69/340 | 2.7 (1.6–4.6) | <0.001 | 17 (13–21); 57/340 | 2.3 (1.3–4.2) | 0.004 | 22 (17–26); 73/340 | 2.4 (1.4–4.3) | 0.001 |
| Total | 24 (20–28); 97/409 | 19 (16–23); 79/409 | 25 (21–29); 101/409 | ||||||
| Variables (n = 360) | Odds Ratio | 95% CI | p-Value | |
|---|---|---|---|---|
| Sex | Female 1 | 0.021 | ||
| Male | 1.83 | 1.096–3.055 | ||
| Region | Western 1 | 0.002 | ||
| Eastern | 2.49 | 1.409–4.404 |
| Model | β (SE) | 95% Wald CI | Wald Chi-Squared | df | p-Value |
|---|---|---|---|---|---|
| Sex, female | 0.345 (0.241) | −0.128 0.818 | 2.039 | 1 | 0.153 |
| Region, eastern | − 0.710 (0.251) | −1.202 −0.218 | 7.991 | 1 | 0.005 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouchard, É.; Sharma, R.; Hernández-Ortiz, A.; Jung, T.S.; Harms, N.J.; Willier, C.N.; Boonstra, R.; Majchrzak, Y.N.; Peers, M.J.L.; Gouin, G.-G.; et al. Canada Lynx (Lynx canadensis) as Potential Reservoirs and Sentinels of Toxoplasma gondii in Northern Canada. Zoonotic Dis. 2023, 3, 6-17. https://doi.org/10.3390/zoonoticdis3010002
Bouchard É, Sharma R, Hernández-Ortiz A, Jung TS, Harms NJ, Willier CN, Boonstra R, Majchrzak YN, Peers MJL, Gouin G-G, et al. Canada Lynx (Lynx canadensis) as Potential Reservoirs and Sentinels of Toxoplasma gondii in Northern Canada. Zoonotic Diseases. 2023; 3(1):6-17. https://doi.org/10.3390/zoonoticdis3010002
Chicago/Turabian StyleBouchard, Émilie, Rajnish Sharma, Adrián Hernández-Ortiz, Thomas S. Jung, N. Jane Harms, Caitlin N. Willier, Rudy Boonstra, Yasmine N. Majchrzak, Michael J. L. Peers, Géraldine-G. Gouin, and et al. 2023. "Canada Lynx (Lynx canadensis) as Potential Reservoirs and Sentinels of Toxoplasma gondii in Northern Canada" Zoonotic Diseases 3, no. 1: 6-17. https://doi.org/10.3390/zoonoticdis3010002