
The Isolation, Screening, and Characterization of Polyhydroxyalkanoate-Producing Bacteria from Hypersaline Lakes in Kenya


Abstract
:1. Introduction
2. Results
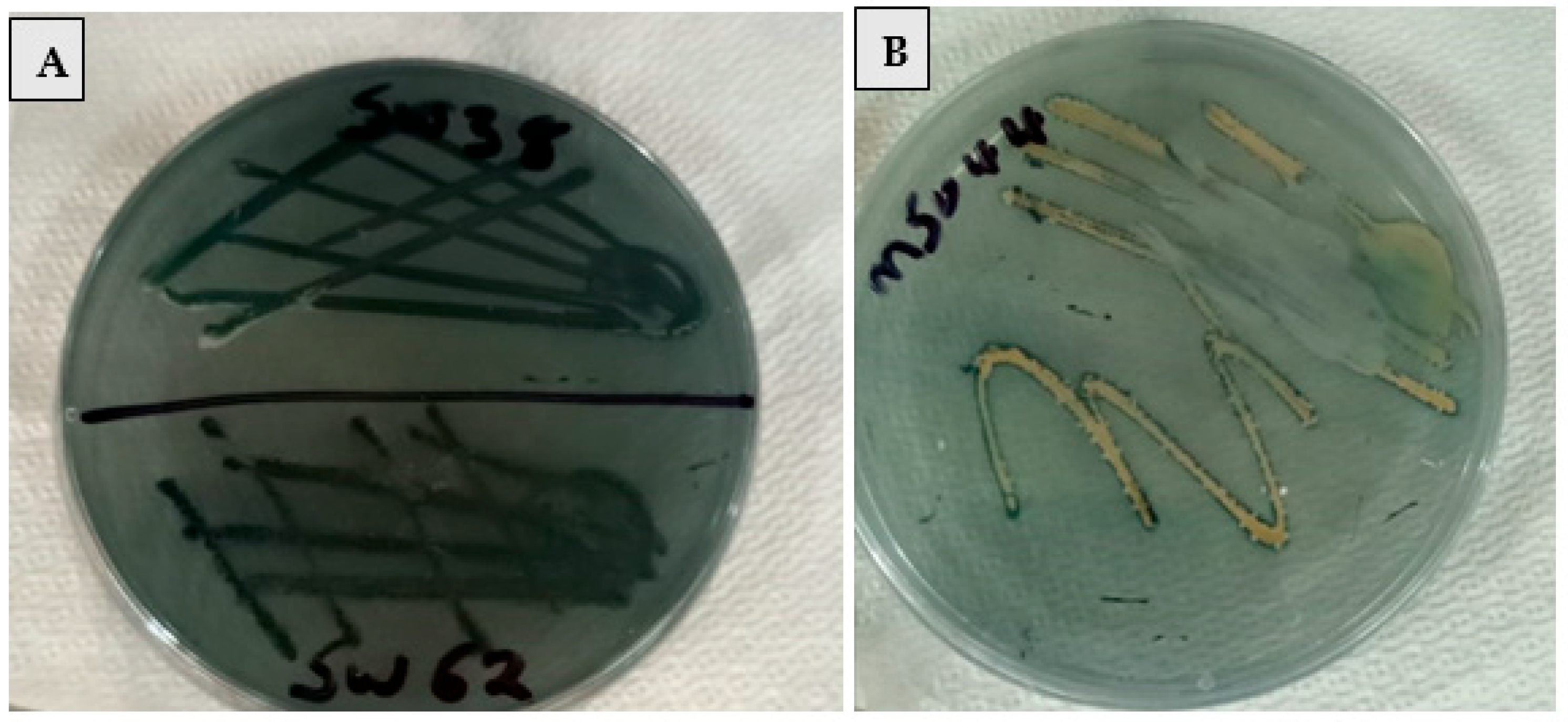
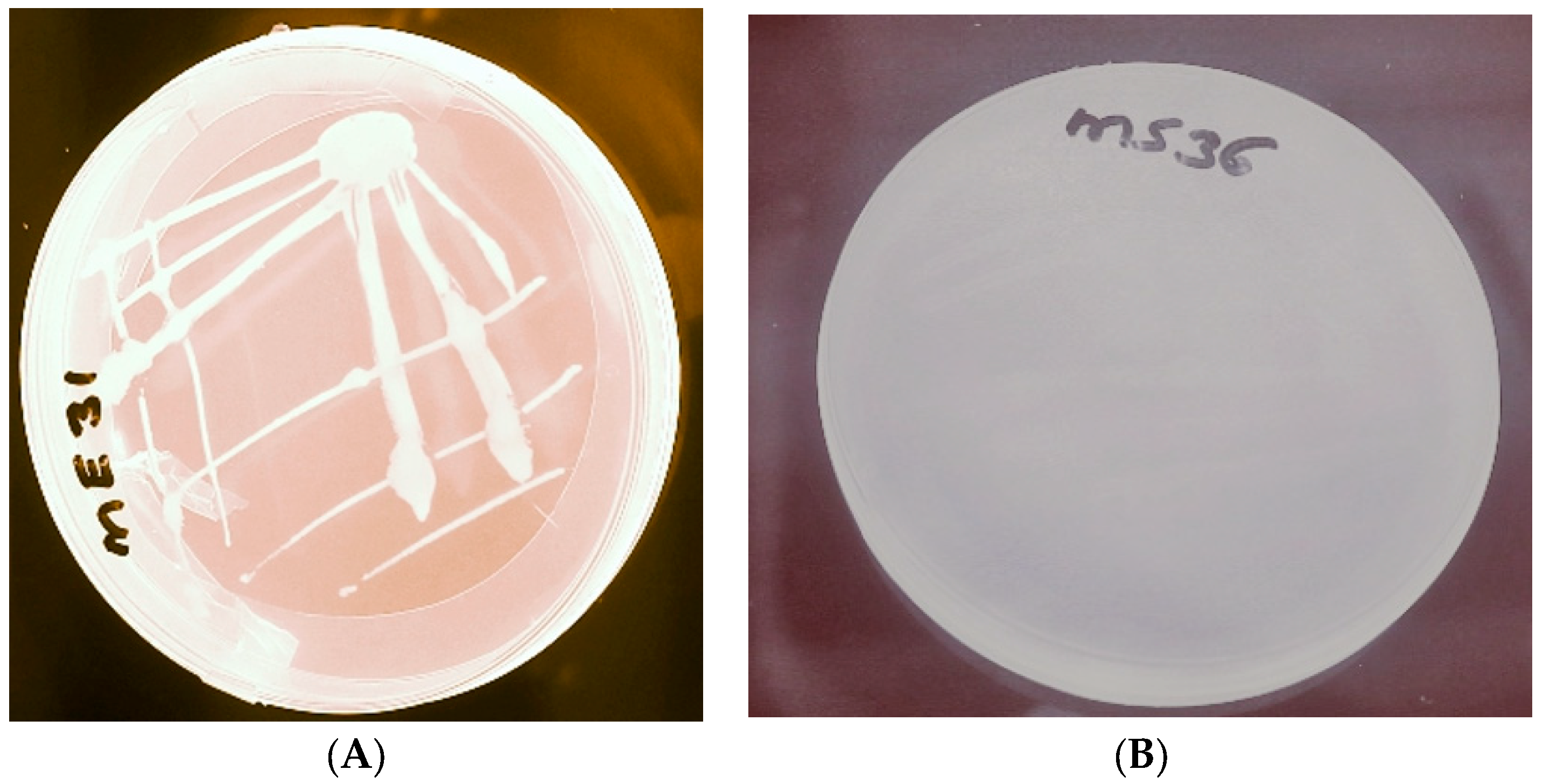
2.1. Isolation and Screening of PHA-Producing Bacteria
2.2. Characterization of PHA-Producing Bacteria
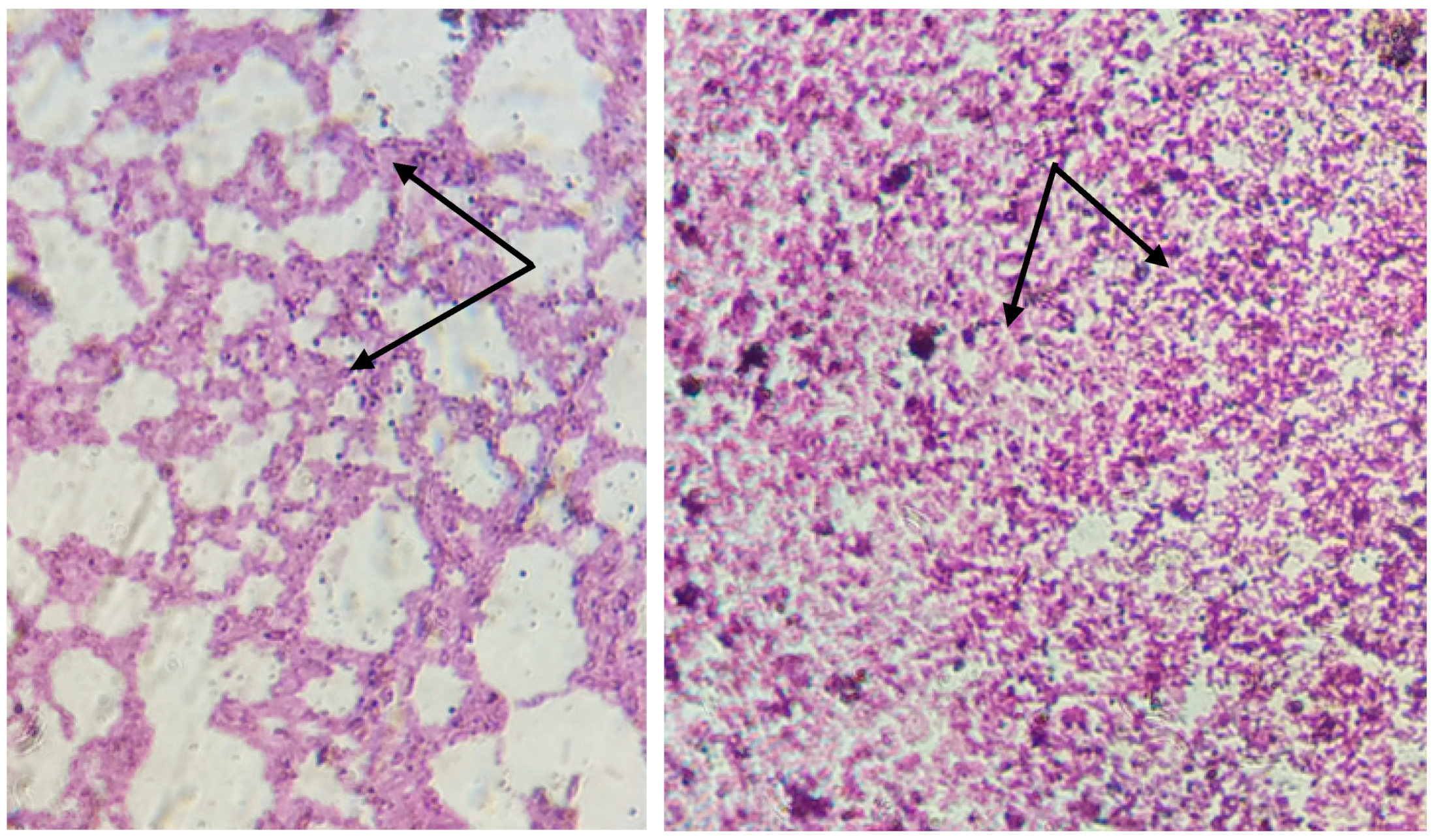
2.2.1. Morphological and Biochemical Characterization
2.2.2. Molecular Characterization
2.3. Production of Polyhydroxyalkanoates by Bacteria Isolated from Hypersaline Lakes in Kenya
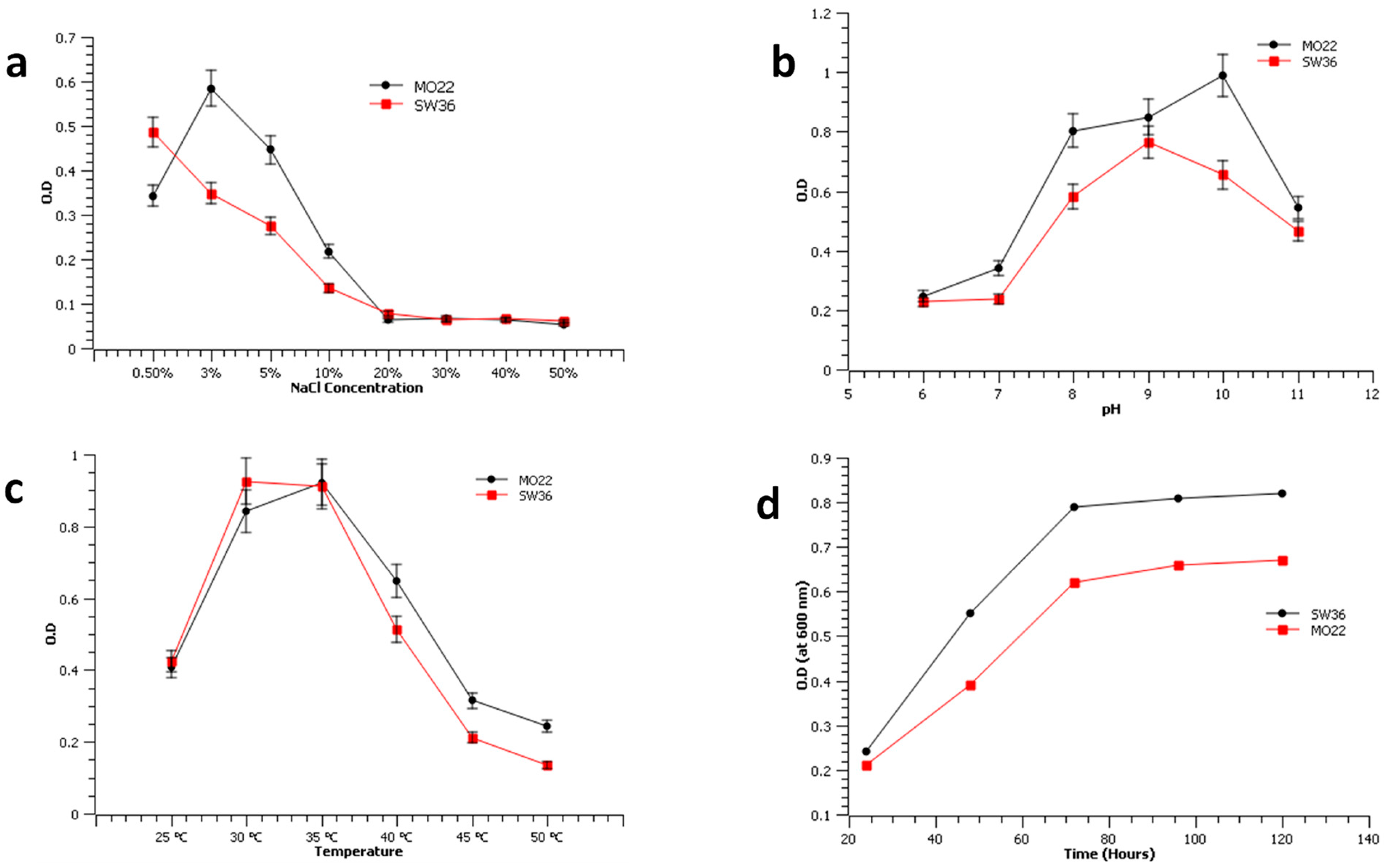
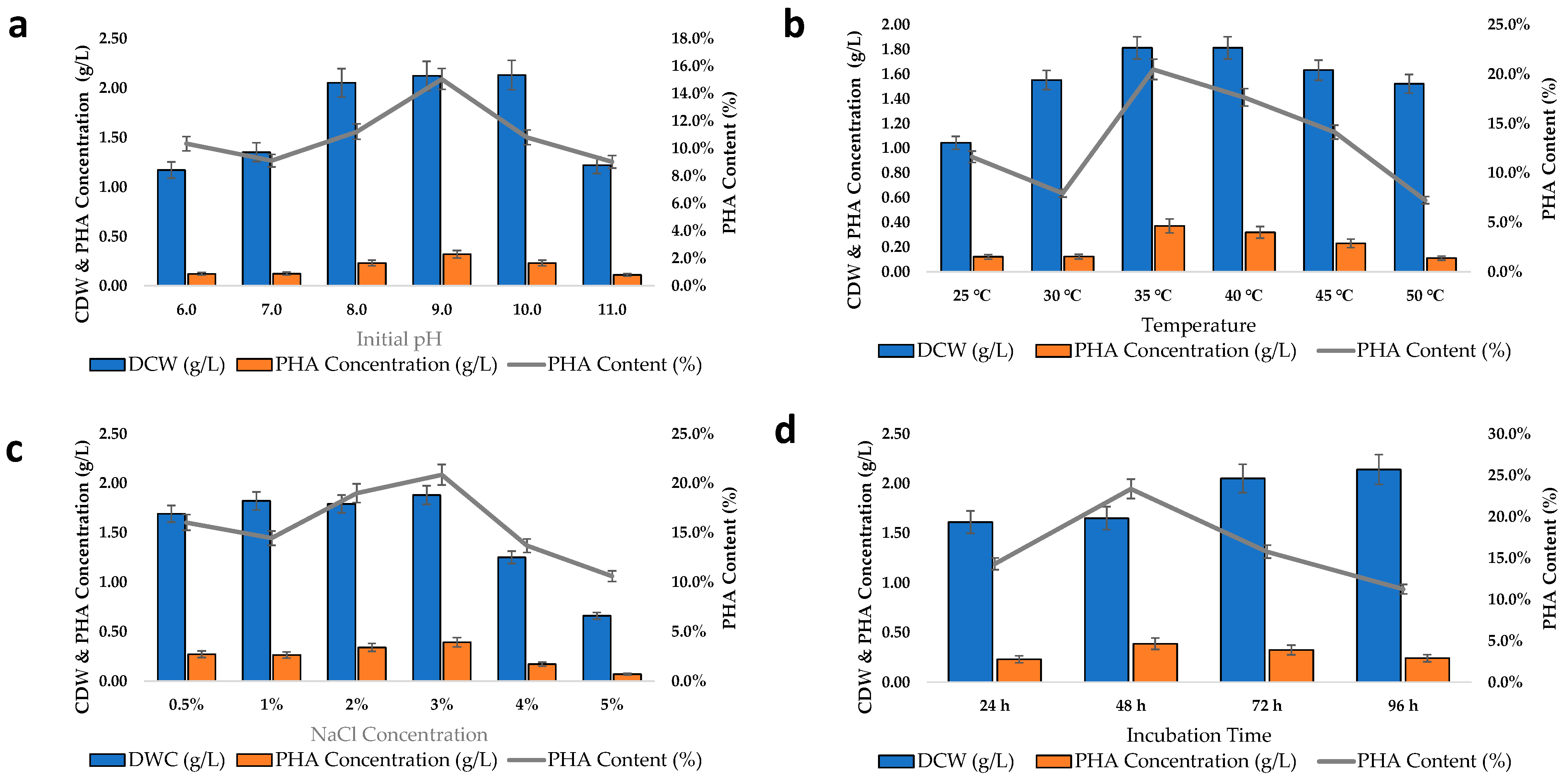
2.4. Bacteria Cell Growth Dynamics
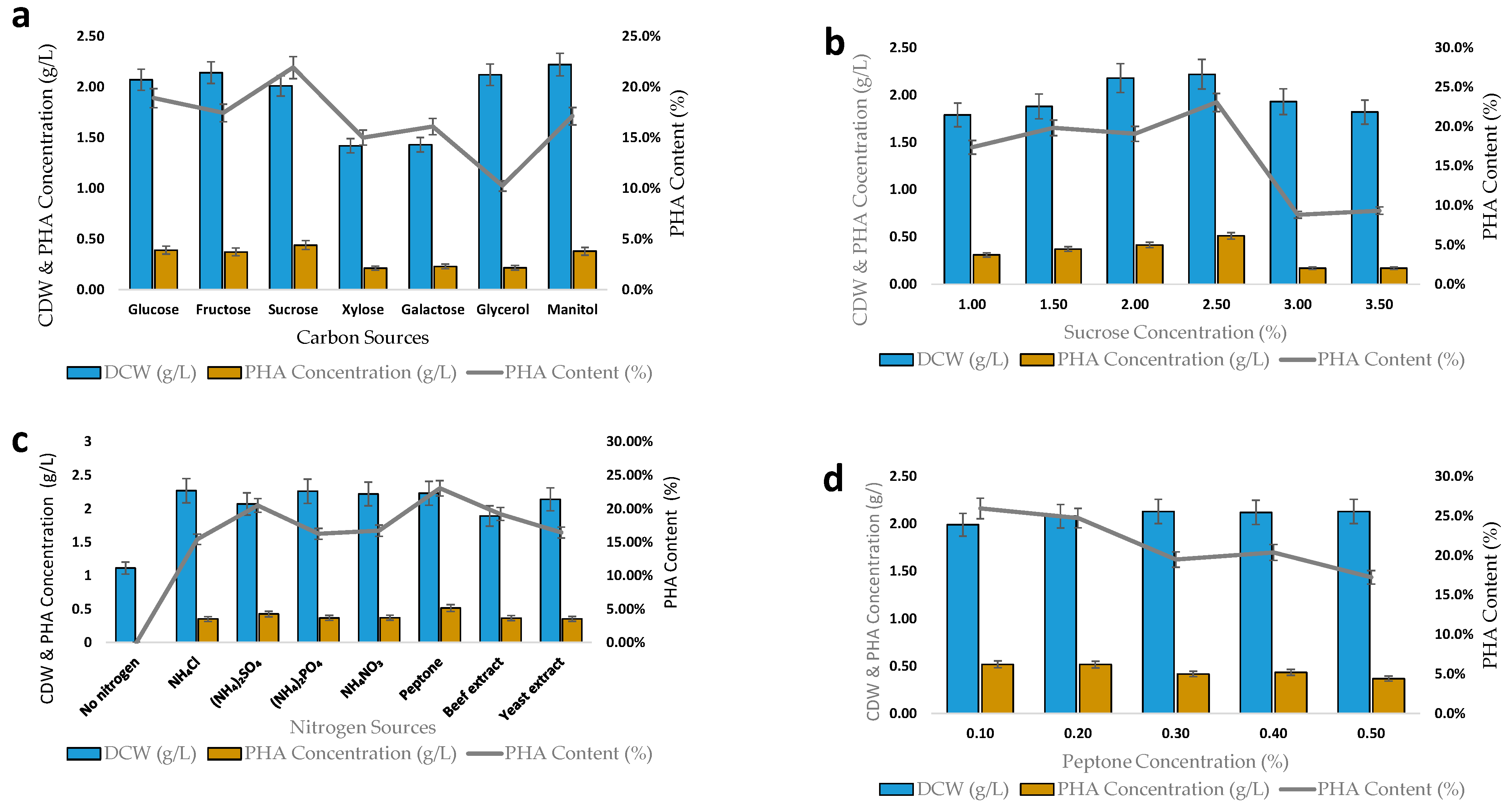
2.5. Production of Polyhydroxyalkanoates on Different Carbon and Nitrogen Sources
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Isolation of PHA-Producing Bacteria
4.2. Screening of PHA-Producing Bacteria
4.3. Quantification of Cell Growth
4.4. PHA Production and Recovery in Submerged Fermentation
4.5. Characterization of PHA-Producing Bacteria and Phylogenetic Analysis
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- OECD. Plastic Pollution Is Growing Relentlessly as Waste Management and Recycling Fall Short; OECD: Paris, France, 2022. [Google Scholar]
- Cabernard, L.; Pfister, S.; Oberschelp, C.; Hellweg, S. Growing Environmental Footprint of Plastics Driven by Coal Combustion. Nat. Sustain. 2022, 5, 139–148. [Google Scholar] [CrossRef]
- Moshood, T.D.; Nawanir, G.; Mahmud, F.; Mohamad, F.; Ahmad, M.H.; AbdulGhani, A. Sustainability of Biodegradable Plastics: New Problem or Solution to Solve the Global Plastic Pollution? Curr. Res. Green Sustain. Chem. 2022, 5, 100273. [Google Scholar] [CrossRef]
- Briassoulis, D.; Pikasi, A.; Hiskakis, M. Recirculation Potential of Post-Consumer /Industrial Bio-Based Plastics through Mechanical Recycling—Techno-Economic Sustainability Criteria and Indicators. Polym. Degrad. Stab. 2021, 183, 109217. [Google Scholar] [CrossRef]
- Podkościelna, B.; Gargol, M.; Goliszek, M.; Klepka, T.; Sevastyanova, O. Degradation and Flammability of Bioplastics Based on PLA and Lignin. Polym. Test. 2022, 111, 107622. [Google Scholar] [CrossRef]
- Singh Saharan, B.; Grewal, A.; Kumar, P. Biotechnological Production of Polyhydroxyalkanoates: A Review on Trends and Latest Developments. Chin. J. Biol. 2014, 2014, 802984. [Google Scholar] [CrossRef]
- Amir, M.; Bano, N.; Baker, A.; Zia, Q.; Banawas, S.; Zaheer, M.R.; Shariq, M.; Nawaz, M.S.; Khan, M.F.; Azad, Z.R.A.A.; et al. Isolation and Optimization of Extracellular PHB Depolymerase Producer Aeromonas Caviae Kuk1-(34) for Sustainable Solid Waste Management of Biodegradable Polymers. PLoS ONE 2022, 17, e0264207. [Google Scholar] [CrossRef]
- Zhou, W.; Colpa, D.I.; Geurkink, B.; Euverink, G.J.W.; Krooneman, J. The Impact of Carbon to Nitrogen Ratios and PH on the Microbial Prevalence and Polyhydroxybutyrate Production Levels Using a Mixed Microbial Starter Culture. Sci. Total Environ. 2022, 811, 152341. [Google Scholar] [CrossRef]
- Schlegel, H.G.; Gottschalk, G.; Von Bartha, R. Formation and Utilization of Poly-β-Hydroxybutyric Acid by Knallgas Bacteria (Hydrogenomonas). Nature 1961, 191, 463–465. [Google Scholar] [CrossRef]
- Anbukarasu, P.; Sauvageau, D.; Elias, A. Tuning the Properties of Polyhydroxybutyrate Films Using Acetic Acid via Solvent Casting. Sci. Rep. 2015, 5, 17884. [Google Scholar] [CrossRef]
- Madison, L.L.; Huisman, G.W. Metabolic Engineering of Poly(3-Hydroxyalkanoates): From DNA to Plastic. Microbiol. Mol. Biol. Rev. 1999, 63, 21–53. [Google Scholar] [CrossRef]
- Mohapatra, S.; Mohanta, P.R.; Sarkar, B.; Daware, A.; Kumar, C.; Samantaray, D.P. Production of Polyhydroxyalkanoates (PHAs) by Bacillus Strain Isolated from Waste Water and Its Biochemical Characterization. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2017, 87, 459–466. [Google Scholar] [CrossRef]
- Kaur, L.; Khajuria, R.; Parihar, L.; Dimpal Singh, G. Polyhydroxyalkanoates: Biosynthesis to Commercial Production—A Review. J. Microbiol. Biotechnol. Food Sci. 2017, 6, 1098–1106. [Google Scholar] [CrossRef]
- Kumar, M.; Rathour, R.; Singh, R.; Sun, Y.; Pandey, A.; Gnansounou, E.; Andrew Lin, K.Y.; Tsang, D.C.W.; Thakur, I.S. Bacterial Polyhydroxyalkanoates: Opportunities, Challenges, and Prospects. J. Clean. Prod. 2020, 263, 121500. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, P.; Ray, S.; Kalia, V.C. Challenges and Opportunities for Customizing Polyhydroxyalkanoates. Indian J. Microbiol. 2015, 55, 235–249. [Google Scholar] [CrossRef] [PubMed]
- Ganesh Saratale, R.; Cho, S.K.; Dattatraya Saratale, G.; Kadam, A.A.; Ghodake, G.S.; Kumar, M.; Naresh Bharagava, R.; Kumar, G.; Su Kim, D.; Mulla, S.I.; et al. A Comprehensive Overview and Recent Advances on Polyhydroxyalkanoates (PHA) Production Using Various Organic Waste Streams. Bioresour. Technol. 2021, 325, 124685. [Google Scholar] [CrossRef]
- Kourmentza, C.; Plácido, J.; Venetsaneas, N.; Burniol-Figols, A.; Varrone, C.; Gavala, H.N.; Reis, M.A.M. Recent Advances and Challenges towards Sustainable Polyhydroxyalkanoate (PHA) Production. Bioengineering 2017, 4, 55. [Google Scholar] [CrossRef]
- Mitra, R.; Xu, T.; Xiang, H.; Han, J. Current Developments on Polyhydroxyalkanoates Synthesis by Using Halophiles as a Promising Cell Factory. Microb. Cell Fact. 2020, 19, 86. [Google Scholar] [CrossRef]
- Chen, G.-Q.; Jiang, X.-R. Next Generation Industrial Biotechnology Based on Extremophilic Bacteria. Curr. Opin. Biotechnol. 2018, 50, 94–100. [Google Scholar] [CrossRef]
- Zhang, X.; Lin, Y.; Chen, G.Q. Halophiles as Chassis for Bioproduction. Adv. Biosyst. 2018, 2, 1800088. [Google Scholar] [CrossRef]
- Ben Abdallah, M.; Karray, F.; Sayadi, S. Production of Polyhydroxyalkanoates by Two Halophilic Archaeal Isolates from Chott El Jerid Using Inexpensive Carbon Sources. Biomolecules 2020, 10, 109. [Google Scholar] [CrossRef]
- Chanprateep, S. Current Trends in Biodegradable Polyhydroxyalkanoates. J. Biosci. Bioeng. 2010, 110, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Gomez, J.G.C.; Rodrigues, M.F.A.; Alli, R.C.P.; Torres, B.B.; Bueno Netto, C.L.; Oliveira, M.S.; Da Silva, L.F. Evaluation of Soil Gram-Negative Bacteria Yielding Polyhydroxyalkanoic Acids from Carbohydrates and Propionic Acid. Appl. Microbiol. Biotechnol. 1996, 45, 785–791. [Google Scholar] [CrossRef]
- Pernicova, I.; Novackova, I.; Sedlacek, P.; Kourilova, X.; Kalina, M.; Kovalcik, A.; Koller, M.; Nebesarova, J.; Krzyzanek, V.; Hrubanova, K.; et al. Introducing the Newly Isolated Bacterium Aneurinibacillus Sp. H1 as an Auspicious Thermophilic Producer of Various Polyhydroxyalkanoates (PHA) Copolymers-1. Isolation and Characterization of the Bacterium. Polymers 2020, 12, 1235. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, K.; Ohta, A.; Hyakutake, M.; Ichinomiya, Y.; Tsuge, T. Isolation of Polyhydroxyalkanoate-Producing Bacteria from a Polluted Soil and Characterization of the Isolated Strain Bacillus Cereus YB-4. Polym. Degrad. Stab. 2010, 95, 1335–1339. [Google Scholar] [CrossRef]
- Możejko-Ciesielska, J.; Kiewisz, R. Bacterial Polyhydroxyalkanoates: Still Fabulous? Microbiol. Res. 2016, 192, 271–282. [Google Scholar] [CrossRef]
- Sehgal, R.; Gupta, R. Polyhydroxyalkanoate and Its Efficient Production: An Eco-Friendly Approach towards Development. 3 Biotech 2020, 10, 549. [Google Scholar] [CrossRef]
- Das, R.; Pal, A.; Paul, A.K. Assessment and Characterization of Poly(3-Hydroxybutyrate) Accumulated by Endophytic Bacterium Exiguobacterium acetylicum BNL 103 from Oleaginous Plant Brassica napus L. Environ. Sustain. 2022, 5, 103–117. [Google Scholar] [CrossRef]
- Kumar, P.; Kim, B.S. Valorization of Polyhydroxyalkanoates Production Process by Co-Synthesis of Value-Added Products. Bioresour. Technol. 2018, 269, 544–556. [Google Scholar] [CrossRef]
- Pernicova, I.; Kucera, D.; Nebesarova, J.; Kalina, M.; Novackova, I.; Koller, M.; Obruca, S. Production of Polyhydroxyalkanoates on Waste Frying Oil Employing Selected Halomonas Strains. Bioresour. Technol. 2019, 292, 122028. [Google Scholar] [CrossRef]
- Tao, W.; Lv, L.; Chen, G.Q. Engineering Halomonas Species TD01 for Enhanced Polyhydroxyalkanoates Synthesis via CRISPRi. Microb. Cell Fact. 2017, 16, 48. [Google Scholar] [CrossRef]
- El-malek, F.A.; Farag, A.; Omar, S.; Khairy, H. Polyhydroxyalkanoates (PHA) from Halomonas Pacifica ASL10 and Halomonas Salifodiane ASL11 Isolated from Mariout Salt Lakes. Int. J. Biol. Macromol. 2020, 161, 1318–1328. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.B.; Li, H.C.; Li, D.Q.; Nawaz, H.; You, T.T.; Xu, F. Efficiently Unsterile Polyhydroxyalkanoate Production from Lignocellulose by Using Alkali-Halophilic Halomonas Alkalicola M2. Bioresour. Technol. 2022, 351, 126919. [Google Scholar] [CrossRef]
- Thomas, T.; Sudesh, K.; Bazire, A.; Elain, A.; Tan, H.T.; Lim, H.; Bruzaud, S. PHA Production and Pha Synthases of the Halophilic Bacterium Halomonas Sp. SF2003. Bioengineering 2020, 7, 29. [Google Scholar] [CrossRef] [PubMed]
- Hammami, K.; Souissi, Y.; Souii, A.; Ouertani, A.; El-Hidri, D.; Jabberi, M.; Chouchane, H.; Mosbah, A.; Masmoudi, A.S.; Cherif, A.; et al. Extremophilic Bacterium Halomonas desertis G11 as a Cell Factory for Poly-3-Hydroxybutyrate-Co-3-Hydroxyvalerate Copolymer’s Production. Front. Bioeng. Biotechnol. 2022, 10, 878843. [Google Scholar] [CrossRef]
- Abdelmalek, F.; Steinbüchel, A.; Rofeal, M. The Hyperproduction of Polyhydroxybutyrate Using Bacillus Mycoides ICRI89 through Enzymatic Hydrolysis of Affordable Cardboard. Polymers 2022, 14, 2810. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.; Kumar, A. Production and Characterization of Polyhydroxyalkanoates from Industrial Waste Using Soil Bacterial Isolates. Braz. J. Microbiol. 2021, 52, 715–726. [Google Scholar] [CrossRef]
- Elain, A.; Le Grand, A.; Corre, Y.M.; Le Fellic, M.; Hachet, N.; Le Tilly, V.; Loulergue, P.; Audic, J.L.; Bruzaud, S. Valorisation of Local Agro-Industrial Processing Waters as Growth Media for Polyhydroxyalkanoates (PHA) Production. Ind. Crops Prod. 2016, 80, 1–5. [Google Scholar] [CrossRef]
- Cui, Y.W.; Zhang, H.Y.; Lu, P.F.; Peng, Y.Z. Effects of Carbon Sources on the Enrichment of Halophilic Polyhydroxyalkanoate-Storing Mixed Microbial Culture in an Aerobic Dynamic Feeding Process. Sci. Rep. 2016, 6, 30766. [Google Scholar] [CrossRef]
- Carvalho, J.M.; Marreiros, B.C.; Reis, M.A.M. Polyhydroxyalkanoates Production by Mixed Microbial Culture under High Salinity. Sustainability 2022, 14, 1346. [Google Scholar] [CrossRef]
- Corrado, I.; Petrillo, C.; Isticato, R.; Casillo, A.; Corsaro, M.M.; Sannia, G.; Pezzella, C. The Power of Two: An Artificial Microbial Consortium for the Conversion of Inulin into Polyhydroxyalkanoates. Int. J. Biol. Macromol. 2021, 189, 494–502. [Google Scholar] [CrossRef]
- Tan, B.; Zheng, Y.; Yan, H.; Liu, Y.; Li, Z.J. Metabolic Engineering of Halomonas Bluephagenesis to Metabolize Xylose for Poly-3-Hydroxybutyrate Production. Biochem. Eng. J. 2022, 187, 108623. [Google Scholar] [CrossRef]
- Mohanrasu, K.; Rao, R.G.R.; Dinesh, G.H.; Zhang, K.; Prakash, G.S.; Song, D.P.; Muniyasamy, S.; Pugazhendhi, A.; Jeyakanthan, J.; Arun, A. Optimization of Media Components and Culture Conditions for Polyhydroxyalkanoates Production by Bacillus Megaterium. Fuel 2020, 271, 117522. [Google Scholar] [CrossRef]
- Anjali, M.; Sukumar, C.; Kanakalakshmi, A.; Shanthi, K. Enhancement of Growth and Production of Polyhydroxyalkanoates by Bacillus Subtilis from Agro-Industrial Waste as Carbon Substrates. Compos. Interfaces 2014, 21, 111–119. [Google Scholar] [CrossRef]
- Pan, W.; Perrotta, J.A.; Stipanovic, A.J.; Nomura, C.T.; Nakas, J.P. Production of Polyhydroxyalkanoates by Burkholderia Cepacia ATCC 17759 Using a Detoxified Sugar Maple Hemicellulosic Hydrolysate. J. Ind. Microbiol. Biotechnol. 2012, 39, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Zainab-L, I.; Uyama, H.; Li, C.; Shen, Y.; Sudesh, K. Production of Polyhydroxyalkanoates from Underutilized Plant Oils by Cupriavidus necator. Clean 2018, 46, 1700542. [Google Scholar] [CrossRef]
- Al-Battashi, H.; Annamalai, N.; Al-Kindi, S.; Nair, A.S.; Al-Bahry, S.; Verma, J.P.; Sivakumar, N. Production of Bioplastic (Poly-3-Hydroxybutyrate) Using Waste Paper as a Feedstock: Optimization of Enzymatic Hydrolysis and Fermentation Employing Burkholderia Sacchari. J. Clean. Prod. 2019, 214, 236–247. [Google Scholar] [CrossRef]
- Dubey, S.; Mishra, S. Efficient Production of Polyhydroxyalkanoate Through Halophilic Bacteria Utilizing Algal Biodiesel Waste Residue. Front. Bioeng. Biotechnol. 2021, 9, 624859. [Google Scholar] [CrossRef] [PubMed]
- Aljuraifani, A.A.; Berekaa, M.M.; Ghazwani, A.A. Bacterial Biopolymer (Polyhydroxyalkanoate) Production from Low-Cost Sustainable Sources. Microbiologyopen 2019, 8, e00755. [Google Scholar] [CrossRef] [PubMed]
- Nishida, M.; Tanaka, T.; Hayakawa, Y.; Nishida, M. Solid-State Nuclear Magnetic Resonance (NMR) and Nuclear Magnetic Relaxation Time Analyses of Molecular Mobility and Compatibility of Plasticized Polyhydroxyalkanoates (PHA) Copolymers. Polymers 2018, 10, 506. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, H.G.; Lafferty, R.; Krauss, I. The Isolation of Mutants Not Accumulating Poly-β-Hydroxybutyric Acid. Arch. Mikrobiol. 1970, 71, 283–294. [Google Scholar] [CrossRef]
- Wei, Y.H.; Chen, W.C.; Huang, C.K.; Wu, H.S.; Sun, Y.M.; Lo, C.W.; Janarthanan, O.M. Screening and Evaluation of Polyhydroxybutyrate-Producing Strains from Indigenous Isolate Cupriavidus taiwanensis Strains. Int. J. Mol. Sci. 2011, 12, 252–265. [Google Scholar] [CrossRef] [PubMed]
- Spiekermann, P.; Rehm, B.H.A.; Kalscheuer, R.; Baumeister, D.; Steinbüchel, A. A Sensitive, Viable-Colony Staining Method Using Nile Red for Direct Screening of Bacteria That Accumulate Polyhydroxyalkanoic Acids and Other Lipid Storage Compounds. Arch. Microbiol. 1999, 171, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Hahn, S.K.; Chang, Y.K.; Kim, B.S.; Chang, H.N. Optimization of Microbial Poly(3-hydroxybutyrate) Recover Using Dispersions of Sodium Hypochlorite Solution and Chloroform. Biotechnol. Bioeng. 1994, 44, 256–261. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Isolates/Observations | |||
|---|---|---|---|---|
| SW32 | SW36 | MO22 | ME81 | |
| Colony shape | Circular | Circular | Circular | Circular |
| Color | Yellowish | Off white | Yellow | Cream |
| Cell shape | Rod | Rod | Rod | Rod |
| Gram reaction | + | − | + | + |
| Catalase | + | + | + | + |
| Oxidase | + | + | + | + |
| H2S production | − | − | − | − |
| Voges Proskauer | + | − | − | + |
| Urease activity | − | − | − | + |
| Indole production | − | − | − | − |
| Citrate utilization | + | − | + | + |
| Glucose | + | + | + | + |
| Sucrose | + | + | + | + |
| Xylose | + | − | + | + |
| Lactose | + | + | + | + |
| Starch | − | − | − | − |
| Maltose | + | − | + | + |
| Galactose | + | + | + | + |
| Fructose | + | + | + | + |
| Mannose | + | + | − | − |
| Cellobiose | + | − | + | + |
| Glycerol | + | + | + | + |
| Mannitol | − | − | + | − |
| Isolate Code | Organism | Query Cover | Percent Identity | Accession No. |
|---|---|---|---|---|
| SO13 | Bacillus sp. 01105 | 99% | 98.52% | EU520307.1 |
| SO31 | Bacillus subtilis strain OTPB28 | 100% | 88.60% | KT265083.1 |
| SO75 | Bacillus sp. strain FA2-253 | 98% | 98.16% | KY476210.1 |
| SE42 | Paracoccus sp. TMN-21-1 | 100% | 96.76% | JX950033.1 |
| SE83 | Exiguobacterium sp. QZS4_8 | 99% | 97.51% | KX364032.1 |
| SE84 | Arthrobacter sp. strain C15 | 100% | 97.96% | MK182873.1 |
| SE89 | Paracoccus sp. TMN-21-1 | 99% | 97.62% | JX950033.1 |
| SE93 | Bacillus sp. ISO_06_Kulunda | 99% | 96.31% | EU676884.1 |
| SW32 | Bacillus pumilus strain 37 | 99% | 94.96% | MK327816.1 |
| SW36 | Halomonas alkalicola EXT | 100% | 97.49% | MK478810 |
| SW38 | Bacillus sp. strain MEB205 | 100% | 97.28% | MN809475.1 |
| SW62 | Bacillus subtilis strain QD9 | 98% | 98.52% | EF488088.1 |
| MO12 | Bacillus sp. KVD-DM52 | 97% | 88.90% | KJ872838.1 |
| MO15 | Bacillus sp. KVD-DM52 | 100% | 96.38% | KJ872838.1 |
| MO22 | Bacillus sp. strain JSM-1684023 | 96% | 96.80% | MG893133.1 |
| MO25 | Rhodobaca sp. strain ZN9W | 99% | 96.89% | MH463983.1 |
| ME31 | Bacillus safensis strain Ter61 | 95% | 97.41% | MW672512.1 |
| ME32 | Bacillus sp. strain CHA410 | 96% | 87.04% | MT355498.1 |
| ME33 | Bacillus pumilus strain 37 | 99% | 98.00% | MK327816.1 |
| ME42 | Rhodobaca sp. strain HJB301 | 100% | 97.47% | MT892652.1 |
| ME51 | Bacillus sp. E-127 | 95% | 95.44% | FJ764772.1 |
| ME54 | Bacillus sp. strain JSM 1684086 | 98% | 97.47% | MG893132.1 |
| ME81 | Bacillus sp. E-127 | 100% | 96.43% | FJ764772.1 |
| ME82 | Bacillus sp. ISO_06_Kulunda | 97% | 96.35% | EU676884.1 |
| ME88 | Bacillus sp. ISO_09_Wadi-Natrun | 95% | 96.85% | EU676885.1 |
| MS42 | Bacillus sp. ISO_06_Kulunda | 99% | 98.63% | EU676884.1 |
| MS44 | Exiguobacterium aurantiacum GBRS02 | 99% | 99.00% | MT373550.1 |
| MS52 | Bacillus sp. ISO_06_Kulunda | 99% | 98.63% | EU676884.1 |
| MW27 | Bacillus sp. A-09 | 76% | 76.47% | AY347311.1 |
| MW52 | Bacillus sp. BAB-1831 | 91% | 99.00% | KF771913.1 |
| MW54 | Bacillus sp. (in: firmicutes) | 99% | 97.49% | KX816446.1 |
| Isolate | Organism | DWC (g/L) | PHA Concentration (g/L) | PHA Content | PHA Yield (g/g) |
|---|---|---|---|---|---|
| ME31 | Bacillus safensis strain Ter61 | 2.881 | 0.151 | 5.24% | 0.008 |
| ME32 | Bacillus sp. strain CHA410 | 2.117 | 0.090 | 4.25% | 0.005 |
| ME33 | Bacillus pumilus strain 37 | 1.890 | 0.215 | 11.38% | 0.011 |
| ME42 | Rhodobaca sp. strain HJB301 | 1.984 | 0.150 | 7.56% | 0.008 |
| ME51 | Bacillus sp. E-127 | 1.678 | 0.110 | 6.56% | 0.006 |
| ME54 | Bacillus sp. strain JSM 1684086 | 0.935 | 0.085 | 9.09% | 0.004 |
| ME81 | Bacillus sp. E-127 | 1.718 | 0.150 | 8.73% | 0.008 |
| ME82 | Bacillus sp. ISO_06_Kulunda | 1.610 | 0.196 | 12.17% | 0.010 |
| ME88 | Bacillus sp. ISO_09_Wadi-Natrun | 0.877 | 0.080 | 9.12% | 0.004 |
| MO12 | Bacillus sp. KVD-DM52 | 1.025 | 0.090 | 8.78% | 0.005 |
| MO15 | Bacillus sp. KVD-DM52 | 1.043 | 0.078 | 7.48% | 0.004 |
| MO22 | Bacillus sp. strain JSM-1684023 | 1.515 | 0.290 | 19.14% | 0.015 |
| MO25 | Rhodobaca sp. strain ZN9W | 2.619 | 0.108 | 4.12% | 0.005 |
| MS42 | Bacillus sp. ISO_06_Kulunda | 1.292 | 0.170 | 13.16% | 0.009 |
| MS44 | Exiguobacterium aurantiacum GBRS02 | 1.888 | 0.144 | 7.63% | 0.007 |
| MS52 | Bacillus sp. ISO_06_Kulunda | 1.305 | 0.095 | 7.28% | 0.005 |
| MW27 | Bacillus sp. A-09 | 3.506 | 0.275 | 7.84% | 0.015 |
| MW52 | Bacillus sp. BAB-1831 | 0.877 | 0.055 | 6.27% | 0.003 |
| MW54 | Bacillus sp. (in: firmicutes) | 1.458 | 0.064 | 4.38% | 0.003 |
| SE42 | Paracoccus sp. TMN-21-1 | 1.130 | 0.079 | 6.99% | 0.004 |
| SE83 | Exiguobacterium sp. QZS4_8 | 1.411 | 0.062 | 4.39% | 0.003 |
| SE84 | Arthrobacter sp. strain C15 | 0.689 | 0.052 | 7.55% | 0.003 |
| SE89 | Paracoccus sp. TMN-21-1 | 1.082 | 0.085 | 7.86% | 0.004 |
| SE93 | Bacillus sp. ISO_06_Kulunda | 1.360 | 0.151 | 11.10% | 0.008 |
| SO13 | Bacillus sp. 01105 | 1.194 | 0.078 | 6.53% | 0.004 |
| SO31 | Bacillus subtilis strain OTPB28 | 1.401 | 0.052 | 3.71% | 0.003 |
| SO75 | Bacillus sp. strain FA2-253 | 1.232 | 0.082 | 6.66% | 0.004 |
| SW32 | Bacillus pumilus strain 37 | 1.772 | 0.235 | 13.26% | 0.012 |
| SW36 | Halomonas alkalicola EXT | 2.412 | 0.397 | 16.46% | 0.020 |
| SW38 | Bacillus sp. strain MEB205 | 1.942 | 0.189 | 9.73% | 0.009 |
| SW62 | Bacillus subtilis strain QD9 | 3.520 | 0.188 | 5.34% | 0.009 |
| Carbon Source | DCW (g/L) | PHA Concentration (g/L) | PHA Content (%) | PHA Yield (g/g) |
|---|---|---|---|---|
| Glucose | 2.07 | 0.39 | 18.89% | 0.02 |
| Fructose | 2.14 | 0.37 | 17.43% | 0.02 |
| Sucrose | 2.01 | 0.44 | 21.89% | 0.02 |
| Xylose | 1.42 | 0.21 | 15.00% | 0.01 |
| Galactose | 1.43 | 0.23 | 16.08% | 0.01 |
| Glycerol | 2.12 | 0.22 | 10.24% | 0.01 |
| Mannitol | 2.22 | 0.38 | 17.12% | 0.02 |
| * Sucrose [23] | - | - | - | * 0.50 |
| Strain | Substrate | PHA Accumulation | Reference |
|---|---|---|---|
| Halomonas bluephagenesis | Xylose | 5.37 g/L | [42] |
| Bacillus mycoides | Cardboard | 2.63 g/L | [36] |
| * Bacillus megaterium | Glucose | 5.61 g/L | [43] |
| Bacillus subtilis | Sugarcane molasses | 2.5 g/L | [44] |
| * Burkholderia cepacia ATCC 17759 | Sugar maple | 8.72 g/L | [45] |
| Cupriavidus necator | Plant oils | 6.0 g/L | [46] |
| Bacillus subtilis | Glucose | 3.09 g/L | [12] |
| Burkholderia sacchari | Wastepaper | 1.6 g/L | [47] |
| ** Bacillus sp. strain JSM-1684023 | Sucrose | 0.516 g/L | ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muigano, M.N.; Anami, S.E.; Onguso, J.M.; Omare, G.M. The Isolation, Screening, and Characterization of Polyhydroxyalkanoate-Producing Bacteria from Hypersaline Lakes in Kenya. Bacteria 2023, 2, 81-97. https://doi.org/10.3390/bacteria2020007
Muigano MN, Anami SE, Onguso JM, Omare GM. The Isolation, Screening, and Characterization of Polyhydroxyalkanoate-Producing Bacteria from Hypersaline Lakes in Kenya. Bacteria. 2023; 2(2):81-97. https://doi.org/10.3390/bacteria2020007
Chicago/Turabian StyleMuigano, Martin N., Sylvester E. Anami, Justus M. Onguso, and Godfrey M. Omare. 2023. "The Isolation, Screening, and Characterization of Polyhydroxyalkanoate-Producing Bacteria from Hypersaline Lakes in Kenya" Bacteria 2, no. 2: 81-97. https://doi.org/10.3390/bacteria2020007