
Role and Regulation of Clp Proteases: A Target against Gram-Positive Bacteria
by
 , and
, and
Camila Queraltó
1,†, 
Ricardo Álvarez
1,†,
Constanza Ortega
1,
Fernando Díaz-Yáñez
1,2,
Daniel Paredes-Sabja
1,2,3 and
Fernando Gil
1,2,* 1
Microbiota-Host Interactions and Clostridia Research Group, Departamento de Ciencias Biológicas, Facultad de Ciencias de la Vida, Universidad Andres Bello, Santiago 8370251, Chile
2
ANID-Millennium Science Initiative Program-Millennium Nucleus in the Biology of the Intestinal Microbiota, Santiago 8370251, Chile
3
Department of Biology, Texas A&M University, College Station, TX 77843, USA
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Bacteria 2023, 2(1), 21-36; https://doi.org/10.3390/bacteria2010002
Submission received: 17 November 2022
/
Revised: 14 December 2022
/
Accepted: 29 December 2022
/
Published: 6 January 2023
Abstract
:Bacterial proteases participate in the proteolytic elimination of misfolded or aggregated proteins, carried out by members of the AAA+ protein superfamily such as Hsp100/Clp, Lon, and FtsH. It is estimated that the Clp and Lon families perform around 80% of cellular proteolysis in bacteria. These functions are regulated, in part, through the spatial and/or temporal use of adapter proteins, which participate in the recognition and delivery of specific substrate proteins to proteases. The proteolysis plays an important role in maintaining and controlling the quality of the proteins, avoiding the accumulation and aggregation of unfolded or truncated proteins. However, this is not their only function, since they play an important role in the formation of virulent phenotypes and in the response to different types of stress faced when entering the host or that occur in the environment. This review summarizes the structural and functional aspects of the Clp proteases and their role in Gram-positive microorganisms.
1. Introduction
Proteolysis mediates the selective renewal of several cellular proteins, eliminating those that are defective or unwanted, thus allowing quality control of proteins and the different cellular processes [1]. One of the functions of bacterial proteases is the proteolytic elimination of misfolded or aggregated proteins, carried out by members of the AAA+ protein superfamily (ATPase associated with various cellular activities), such as Hsp100/Clp (heat shock protein-100/caseinolytic protease), Lon, and FtsH [2]. It has been estimated that the Clp and Lon families perform around 80% of cellular proteolysis in bacteria [3,4]. In addition, they control the proteolysis of regulatory proteins, such as key transcription factors that control the cell cycle and bacterial development or adaptation. These two opposite functions are regulated, in part, through the spatial and/or temporal use of adapter proteins, which participate in the recognition and delivery of specific substrate proteins to proteases [5,6].
The HSP100/Clp family of ATPases plays crucial roles in the folding, assembly, and degradation of proteins during normal growth and, mainly, under stress-inducing conditions [7,8]. This family is formed by several ATPase chaperones and the peptidase ClpP (caseinolytic protease proteolytic subunit). Chaperones are divided into two classes: Class I, whose members are ClpA (caseinolytic protease subunit (A), ClpB (caseinolytic protease subunit (B), and ClpC (caseinolytic protease subunit (C), which have two ATP-binding domains separated by a spacer region; and Class II, which includes ClpX (caseinolytic protease subunit X) and ClpY (caseinolytic protease subunit Y), that present only one domain of binding to ATP. Most of the chaperones bind to ClpP peptidase to form a proteolytic complex, with the exception of ClpY which only interacts with ClpQ, forming ClpQY peptidase, also known as HsIUV [9]. ClpQ is part of the Clp family, and like ClpP, is an ATP-dependent peptidase. However, it is one of the least-studied and its biological function and regulation are still not very clear. In addition, it has been shown to exhibit differences in the active site between Gram-positive and Gram-negative organisms [10].
The chaperone–ClpP complex is capable of degrading proteins in a specific manner where the chaperones can use ATP to promote protein folding changes and direct protein degradation by ClpP [11,12]. Although ClpP is part of this family, it does not have the same functions, since, unlike the rest, it is an ATP-dependent peptidase that, when associated with one of the chaperones, has serine protease activity [13]. Most bacteria contain the ClpXP protease, which makes ClpXP the most ubiquitous of the Clp proteases. On the other hand, ClpA and ClpC are orthologous; ClpA is usually found in Gram-negative bacteria, while ClpC is found in Gram-positive bacteria and cyanobacteria. ClpYQ exists together with ClpAP in most Gram-negative bacteria, and is also found in certain Gram-positive bacteria. Protein degradation dependent on these proteases has been studied in detail in the Gram-negative bacterium Escherichia coli (E. coli), whereas ClpCP has been characterized in the Gram-positive, spore-forming bacterium, Bacillus subtilis (B. subtilis) [9,14].
At the end of the 1990s, ClpP began to attract attention due to its potential as an antibacterial target, demonstrating that it participates in the bacterial virulence of the Gram-positive pathogens Staphylococcus aureus (S. aureus) and Listeria monocytogenes (L. monocytogenes). Since then, its use as a pharmaceutical target has been described in both Gram-positive and Gram-negative bacteria, being effective mostly in Gram-positive bacteria, where it proved to be effective in eliminating S. aureus, Enterococcus faecalis (E. faecalis), Streptococcus pneumoniae (S. pneumoniae), B. subtilis, and L. monocytogenes [9,15].
In this review, we summarize and discuss the structural and functional aspects of the Clp machinery. We will analyze the role of Clp proteases and chaperones in Gram-positive microorganisms. Moreover, we will describe several compounds that target Clp, focusing on ClpP.
2. Clp Protease Families
To date, ATP-dependent proteases Lon, FtsH, and Clp have been characterized [1,2,16]. These complexes are responsible for maintaining a proper balance between protein synthesis and degradation at the cellular level, helping to maintain homeostasis. The peptidases of the Hsp100/Clp family are part of the quality control system of proteins both in normal growth and under stress conditions, playing an important role [17] (Figure 1 and Table 1).
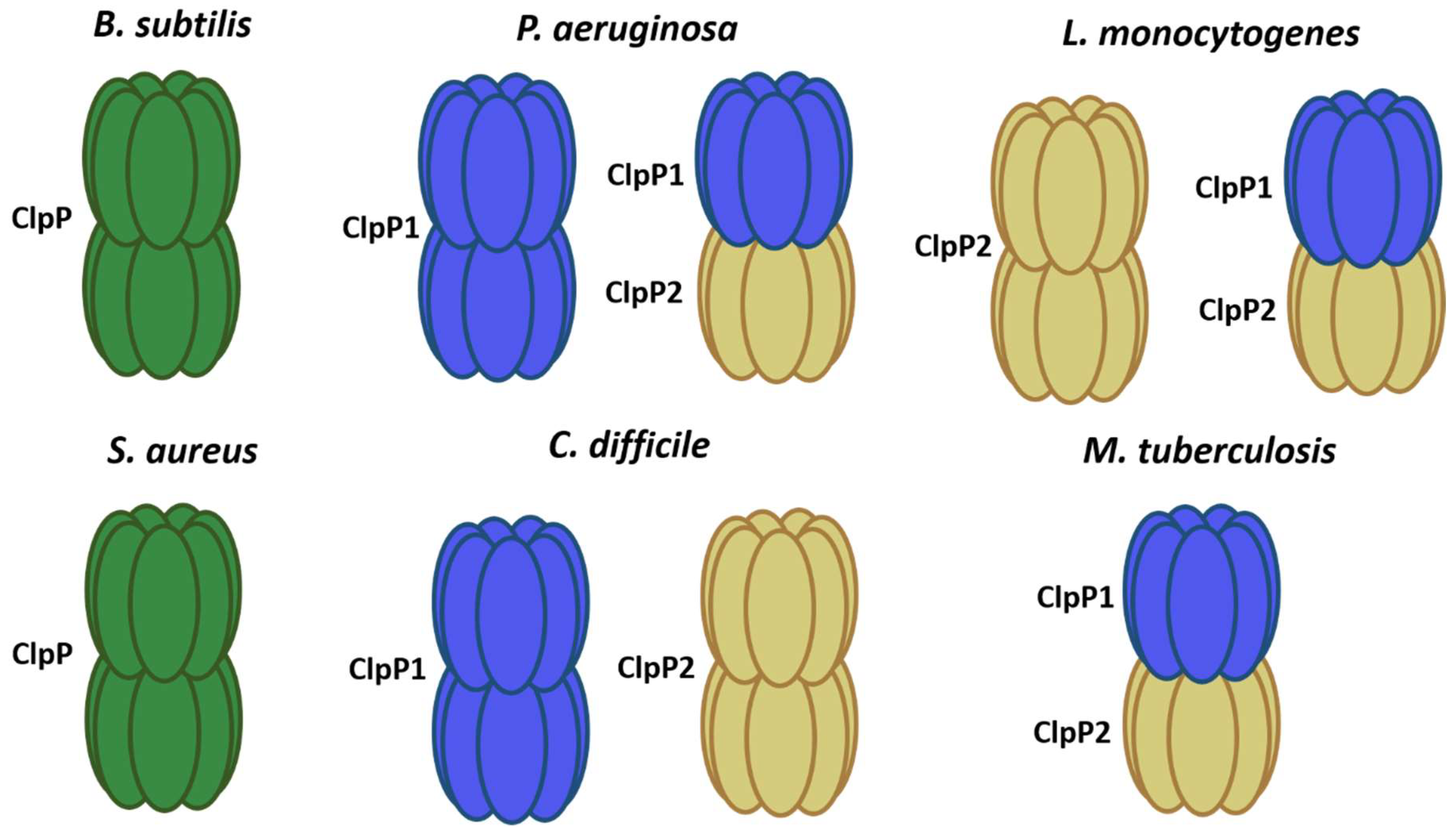
The ClpP peptidase is well-conserved and characterized in different bacterial species. It has a structure formed by two rings of heptamers. The axial pore of each ring acts as an entrance to the proteolytic chamber, where there are 14 active sites formed by the catalytic triad serine-histidine-aspartate. In most organisms, the ClpP is formed by 14 identical monomers [1,15]. However, it can not only be found as a homodimer but also as a heterodimer, as in the case of ClpP1P2 from Mycobacterium tuberculosis (M. tuberculosis), where the active peptidase is composed of two isoforms of ClpP [18]. Pathogenic bacteria such as Pseudomonas aeruginosa (P. aeruginosa) and Clostridioides difficile (C. difficile) also have two isoforms of ClpP; nevertheless, each isoform can produce an active homotetradecamer [19,20] (Figure 1).
This protein acts as a peptidase capable of degrading short peptides that can enter its proteolytic chamber through its narrow axial pores, so for more efficient proteolysis and ability to degrade larger proteins, it forms complexes with any of the chaperones described above [1] (Figure 2A). These chaperones recognize, unfolded through hydrolysis, and introduce in the proteolytic chamber of ClpP a range of substrates for their degradations. Although it has been seen that proteins that are already deployed can enter the ClpP proteolytic chamber without the help of chaperones, this occurs much slower than in the presence of chaperones [21,22].
On the one hand, ClpX and ClpC chaperones are highly conserved ATP-dependent proteins in Gram-positive bacteria, in addition to their classic chaperone functions, and can associate with ClpP to form the proteolytic complexes ClpXP and ClpCP [23], while it is believed that ClpB do not interact with ClpP [5] (Table 1).
The proteolysis carried out by the Clp complexes plays an important role in maintaining and controlling protein quality, avoiding the accumulation and aggregation of unfolded proteins, and eliminating proteins that have already completed their life cycle or are truncated. This is not their only function, since they play an important role in bacterial pathogenesis, participating in the formation of virulent phenotypes and in the response to the different types of stress faced when entering the host or that occur in the environment [1,15]. Considering this, Clp proteases appear as a promising target against bacteria due to their participation in different essential cellular processes and phenotypes related to virulence.
It has been observed that ClpXP peptidase is necessary for the degradation of proteins whose translation is stalled by a labeling system. ClpXP can recognize the SsrA-tag at the C-terminal end of the unfinished protein, degrading it and thus preventing its aggregation [24,25]. Meanwhile, the ClpCP peptidase acts in the control of protein quality by degrading unfolded, misfolded, or aggregated proteins, which accumulate under stress conditions such as heat shock [17,26]. In addition, it has been seen that they are participating in the controlled degradation of transcription factors such as the master regulator of competition ComK [27], the anti-sigma factor SpoIIAB involved in sporulation [28], and the oxidative stress transcription factor Spx [29].
In the case of ClpEP, its expression is tightly controlled and is only induced after a strong heat shock, suggesting that it could act as an additional system in other severe stress conditions [30].

{kind=link}
{kind=link}
{kind=link}
Table 1.
Different proteases and chaperones in Gram-positive bacteria.
| Organism | Protease | Chaperone | Reference |
|---|---|---|---|
| B. subtilis | ClpP–ClpQ | ClpC–ClpE–ClpX | [17] |
| S. aureus | ClpP | ClpC–ClpX | [23] |
| M. tuberculosis | ClpP1P2 | ClpX–ClpC–ClpB | [31] |
| C. difficile | ClpP1–ClpP2 | ClpB–ClpC–ClpX | [20] |
Regulation of Complex
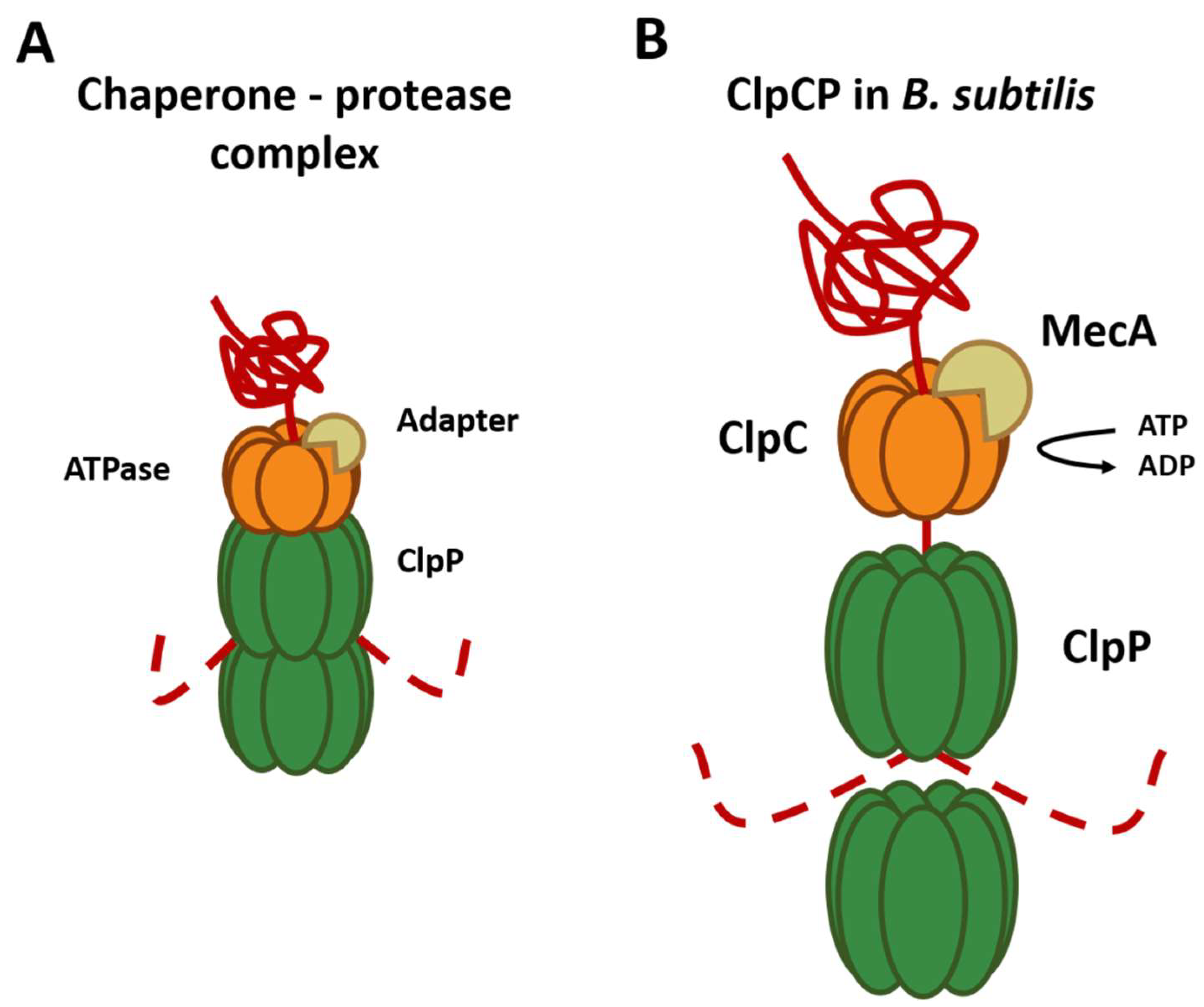
As mentioned above, chaperones can recognize the substrates that will be degraded by ClpP. The best-described substrate class comprises proteins tagged with the ssrA tag, a short peptide sequence C-terminally added to proteins by the tmRNA system to rescue stalled ribosomes. However, for this, the presence of specific substrate adapter proteins, such as YjbH, TrfA, and McsB is required [32]. For example, ClpC requires adapter proteins for all its functions since it is only capable of forming a hexameric ring in the presence of the adapter protein MecA [33] (Figure 2B). These adapter proteins are also regulated through anti-adapter proteins or their phosphorylation of adapters mediated by different signals, for example, the anti-adapter protein ComS and the phosphorylation of the adapter proteins RssB and McsB. This series of regulations, in conjunction with the different Clp ATPases, allows an extensive regulation of the selection of the substrate that will be degraded by the ClpP peptidase.
Other findings showed that the ClpP1P2 complex of M. tuberculosis requires the presence of an activating peptide; this peptide can be N-blocked dipeptide, usually Z-Leu-Leu (Benzyloxycarbonyl-L-Leucyl-L-Leucine), Z-Leu-Leu-H (Benzyloxycarbonyl-L-Leucyl-L-Leucinal), or a similar molecule, that binds near the active sites of the proteolytic particle and stabilizes the active conformation of the ClpP1P2 double-ring [34]. This functional conformation of the complex is also stabilized by the presence of the chaperone ClpX and protein substrate, acting synergistically with the activator peptide [35,36]. The requirement for activators seems to be a unique feature of Actinobacteria, as other species that contain two ClpP proteoforms, such as Listeria monocytogenes [37,38] and Chlamydia trachomatis [39], are functional in absence of activators. In the absence of proteolytic degradation via ClpP1P2, the levels of misfolded proteins can reach toxic levels, leading to cell death. Interestingly, E. coli ClpX rings can interact with M. tuberculosis ClpP1P2 complex and even promote substrate degradation more than ten-fold faster compared to the M. tuberculosis chaperones [36].
Another microorganism that possesses two isoforms of ClpP (ClpP1 and ClpP2) is C. difficile, but their functions are not yet known in detail. ClpP2 was only reported in hypervirulent strains, while ClpP1 has been reported in strains of ribotype 630 and hypervirulent strains, and it has been suggested that the ClpP isoforms act in an independent manner and possess different functions [40]. ClpP1 and ClpP2 have 74% and 63% identity, respectively, with ClpP from B. subtilis, an evolutionarily related organism, and sequence alignment showed that key regions including the catalytic triad are conserved. The evaluation of the proteolytic activity of each isoform was studied; however, only ClpP1 has proteolytic activity. On the other hand, it was revealed that both isoforms can form a complex with ClpX and degrade substrates labeled with the SsrA-tag, suggesting that the binding of the chaperone produces a change at the level of the ClpP2 peptidase that produces its activation [20].
3. Role of the Clp Complexes
3.1. Response to Heat Shock and Oxidative Stress
As previously mentioned, Clp complexes participate in different cellular processes both under homeostasis conditions and, mainly, under stress conditions. In B. subtilis, it has been shown that the participation of adapter proteins together with the chaperones can detect multiple signals to control the activity and stability of different proteins in stressful conditions. In the case of the adapter protein YjbH and the chaperone ClpX, it is known to regulate the activity of Spx [29,41]. Under normal conditions, this complex of ClpXP-YjbH degrades Spx, which controls a regulon involved in the response to oxidative stress and has also been shown to be essential for thermotolerance [42,43]. YjbH is responsible for exposing regions of recognition of ClpXP by Spx, but when the cell faces these types of stress the YjbH protein tends to form aggregates, while the ClpC chaperone that contains regions sensitive to oxidative stress becomes inactive; both processes lead to the accumulation of Spx, as it is not being degraded, allowing the tolerance of the microorganism against oxidative stress and the increase in temperature [44,45].
There are other mechanisms of response to heat and oxidative stress, likewise the regulation of CtsR by ClpCP. CtsR is the repressor of protein quality control genes both in B. subtilis and in all Gram-positive microorganisms, being a key part in the adaptation of the cell to environmental changes [46]. It regulates the expression of its operon which contains ctsR, mcsA, mcsB, and clpC. In addition, it is also found to regulate clpP and clpE [14]. These proteins are inhibited by CtsR and induced under stress conditions, so they can play a crucial role in adaptation. In normal growth, CtsR binds and represses its operator; however, an increase in the temperature induces a conformational change, affecting CtsR binding to DNA, inducing the expression of genes under its control. This inactivation of CtsR molecules leads to protein degradation by ClpCP [47,48]. In addition, ClpE, unlike ClpC, is known to be involved in the maintenance of the repressive activity of CtsR, though it is still not clear how both degradation and reactivation mechanisms work [49]. These proteases were recently shown to be involved in nutrient-dependent cell growth. Nutrients can affect both bacterial growth and morphology [50]. Moreover, it has been disclosed that Clp proteases are responsible for the degradation of UgtP in low carbon conditions, becoming this molecule in an antagonist responsible for the size coordination of cells in response to the availability of nutrients [50].
As in B. subtilis, the ClpCP complex from S. aureus degrades non-native proteins and is carried out in a process that also requires the adapter protein, MecA, which is necessary not only for substrate recognition but also for the oligomerization of ClpC into a hexamer and binding to ClpP [26,33]. A homolog of MecA is also found in S. aureus, where it is the teicoplanin resistance factor designated A (TrfA) [51]. TrfA is a target of Clp protease [52]. and a mutant lacking the corresponding gene is temperature-sensitive, indicating that TrfA can help ClpCP-mediated proteolysis of non-native proteins. McsB is another adapter-like protein also found in B. subtilis that was trapped as a Clp target [46,52,53]. Inactivation of McsB in S. aureus affects growth at high temperatures, as well as in the presence of heavy metals, osmotic pressure, oxidative stress, and low pH [53]. In this case, the sensitivity to heat may be due to the lack of expression of ClpC, since McsB in B. subtilis is necessary for the degradation of the negative heat-shock regulator CtsR, that controls the clpC expression [49].
Surprisingly, the inactivation of chaperone ClpX in S. aureus improved survival at high temperatures, indicating that ClpX does not function as a classic heat shock chaperone in this bacterium [54]. Consistent with this notion, clpX transcription is not inducible by heat stress in S. aureus [54]. However, inactivation of ClpX in S. aureus severely impaired growth at 30 °C or less [54]. The molecular mechanism underlying this phenotype is unknown and is not shared by a mutant lacking clpP, suggesting that it is the ClpP-independent chaperone activity of ClpX that is important for S. aureus growth at 30 °C. [54]. Inactivation of clpX allows S. aureus to grow at higher temperatures than wild-type cells. This shows that Clp proteins contribute to virulence through stress-independent (through ClpXP) and stress-dependent (ClpCP and ClpB) pathways [54,55].
Additionally, in deletion mutants of the clpP, clpC, and clpB, an inhibition or reduction of the S. aureus growth at 45 °C has been observed [54,55,56]. However, ClpB is not functional partners with ClpP, which suggests that the ClpCP complex would be degrading non-native proteins in S. aureus, which was confirmed by trapping protein substrates at high and environmental temperatures, respectively [52]. The likely contribution of ClpB to stress survival is as chaperone to prevent protein development or promote disaggregation, being required for heat-induced thermal tolerance, where pre-exposure to intermediate to high temperatures improves survival at high temperatures [55,57].
In addition, it has been shown that ClpB forms part of a protein bichaperone network together with the DnaK protein. [58,59,60]. DnaK is predicted to be essential for growth due to its involvement in folding nascent proteins and reactivating aggregated proteins along with ClpB [10,61,62,63,64,65,66]. Non-replicating bacterial cells have been shown to contain greater amounts of protein aggregates than those in an actively replicating state [62,66,67,68,69] Factors promoting this increase in protein aggregation include a suboptimal function of protein quality control machinery, lack of nutrients to support protein synthesis, and an increase in cellular oxidants sufficient to cause irreversible protein modifications [70]. M. tuberculosis lacking clpB exhibits defects in recovery after achieving a stationary growth phase, which is one non-replicative state.
3.2. Role of Clp in Natural Competence, Motility, and Biofilm
An important cell development process in which Clp proteases participate is competence, a process in which the bacteria actively absorb extracellular DNA during the stationary growth phase. This process takes the cell to a state known as the K state, in which they exhibit different properties, such as growth inhibition, characteristic of persister cells, which also give them an adaptive advantage over different stressors such as antibiotics [71]. The state of competition, or K state, is regulated by the transcription factor ComK, which induces the transcription of competition genes that lead to the formation of DNA receptors and the transport to the cell [72]. In B. subtilis, during the exponential growth phase, ComK is being degraded by ClpCP, while in the stationary phase ComK remains free and accumulates [73]. This regulation is given by the adapter protein MecA, which is capable of binding ComK, inhibiting it and leading to degradation by ClpCP, but through quorum signaling molecules ComS binds more strongly to MecA, leaving ComK free and leading to the state of competition [27]. On the other hand, it has been seen that the degradation of ComK by ClpCP regulates motility, negatively regulating the expression of the anti-sigma factor FlgM [74]. FlgM inhibits the activity of sigma factor D (SigD), which positively regulates flagellar genes transcription [75]. ClpCP was also shown to regulate motility through DegU, which is a member of the two-component system DegS/DegU, positively or negatively regulating the expression of several related genes with different functions, including flagella formation, biofilm formation, and competition for DNA absorption [76]. Furthermore, it has been reported that the regulation of Spx occurs by ClpXP by an unknown mechanism. In the absence of this peptidase, an increase occurs in the levels of Spx and DegU phosphorylated, which would negatively regulate the expression of the B. subtilis fla/che operon, leading to a decrease in SigD, decreased the expression of late flagellar genes, and consequently a decreased swimming motility [8,77].
In contrast to this, several bacteria can move from a motile to a sessile state, where they form a larger quantity of biofilms. In B. subtilis, the transition to a sessile state depends on the protein SlrR. In a mobile state, SlrR is present in low quantity in bacteria, allowing the expression of genes related to motility. On the contrary, if the levels of SlrR increase, the motility is diminished, giving a phenotype of sessile cell chains and biofilm formation. The change in the concentrations of SlrR depends on its degradation, although the mechanism is still unknown, but it has been shown that ClpCP is participating in the stability of SlrR [78].
3.3. Role of Clp in Sporulation
In unfavorable conditions, some microorganisms are capable of forming spores, which are metabolically inactive and resistant to most environmental changes and types of stress. In B. subtilis, it has been seen that both ClpXP and ClpCP have roles in the formation of these spores through the regulation of different factors [79]. Activation of SigF is strictly necessary for the initiation of sporulation; therefore, the degradation of SpoIIAB by ClpC pate allows to maintain a free SigF and will lead to the formation of spores [28]. The mechanism by which ClpCP degrades SpoIIAB is unknown, but the chaperone is known to be able to recognize an amino acid sequence at the N-terminus of SpoIIAB [80]. On the other hand, ClpXP together with the adapter CmpA participates in the quality control of the spore envelope. If there are defects in the maturation of the spore envelope, CmpA mediates the degradation by ClpXP of the coat protein SpoIVA, which causes instability and lysis of the spore. However, in conditions where the spore does not present defects in the envelope, ClpXP degrades to CmpA. This mechanism is part of those that ensure that only correctly assembled spores are produced, affecting sporulation [79,81].
In the spore-forming pathogen Bacillus anthracis (B. anthracis), sporulation tests have also been performed using mutants in the genes coding for Clp proteases, and it was found that clpC and mcsB are necessary for the formation and germination of spores [82]. Furthermore, it was shown that they also participate in growth under heat stress conditions, as in B. subtilis, and that they are required for the formation of the septum and the maintenance of cell shape [82,83]. The participation of ClpC in sporulation was also demonstrated in the enteropathogenic Bacillus thuringiensis (B. thuringiensis), which, like B. anthracis, has two isoforms of ClpP, ClpP1 and ClpP2, which have been related to the regulation of different types of stress [84,85]. As mentioned above, in B. subtilis, SpoIIAB is regulated by ClpCP, so a similar regulation was suggested in B. anthracis that would explain the decrease in sporulation, since in clpC mutants there is a loss of envelope of the spore [80,82]. Another spore-forming pathogen is C. difficile, where it was shown that the 630 strain in sporulation process expressed only the ClpP1 peptidase and ClpX and ClpC chaperone. However, recent studies in the hypervirulent strain R20291 described that clpP1 and clpP2 participate directly in sporulation, considering that an absence of each ClpP isoform leads to altered sporulation phenotypes and the clpP1/clpP2 double mutant exhibiting asporogenic behavior [86] (Table 1).
ClpP1 is in the genome in an operon along with ClpX, while ClpP2 is in a chromosomally distant region. C. difficile also possesses the ClpB and ClpC chaperones [20] (Table 1). Genomic and proteomic studies have been carried out that have evidenced the presence of these proteases under different conditions. In C. difficile 630, in the sporulation process and subjected to heat shock, it showed the presence only of the ClpP1 peptidase and ClpX and ClpC chaperones. In contrast, a microarray analysis detected both isoforms, but ClpP1 is the only one that increases under pH stress and antibiotic challenge [87,88,89]. On the other hand, ClpP2 has been involved in hypervirulent and resistant phenotypes, since its transcription has been reported in the NAP125 strain and the CD38-2 prophage.
3.4. Role of Clp in Survival and Virulence
As mentioned above, these proteolytic complexes are involved in processes such as sporulation and motility, which are related to the virulence of different pathogens. In this sense, studies have been carried out to determine the role of these proteases in the virulence and survival of pathogens, demonstrating that they are necessary for an effective infection [55,90] In S. aureus, ClpP is the most abundant peptidase under standard laboratory growth conditions [16,54]. Furthermore, the proteolytic activity of ClpP is critical for the virulence of S. aureus [54]. In clpX or clpP mutant strains, a virulence deficiency was observed in a skin abscess model where the infection did not extend from the inoculation site, and it was demonstrated that this deficiency was due to the decrease in the secretion of hemolysin and other factors encoded by the accessory gene regulator (agr locus). In addition, it was found that ClpP regulates the levels of the Isd protein responsible for extracting iron, which is necessary for pathogenicity, while ClpC showed significant defects in long-term survival and intracellular replication of this pathogen [55,90]. Virulence was reduced in the clpP mutant strain, most likely due to the repression of agr-regulated virulence genes [54]. The expression of the global regulatory agr gene was decreased in this mutant strain, leading to reduced production of α-toxin and induction of urease activity. Furthermore, the global regulator MgrA, which is involved in antibiotic resistance and virulence, was found to be significantly regulated by clpP clearance. Therefore, ClpP can control the transcription of numerous virulence factors, such as the urease operon and hla, by modulating the level of MgrA [56,90]. Furthermore, there is evidence that Rot (toxin repressor) in complex with RNAIII is a substrate for the serine protease SspA under the control of Clp-dependent degradation [16].
The ClpX chaperone is highly conserved in S. aureus and has a dual role in the cell, associating with the peptidase ClpP and, independently of ClpP, facilitating protein folding and interactions [6]. In S. aureus, inactivation of clpX severely reduced virulence in localized and systemic models of infections, suggesting that ClpX is indispensable for the pathogenesis of S. aureus [54,91,92]. Consistent with this observation, ClpX controls, by an unknown mechanism, the transcription of several major virulence genes and the translation of protein A and the global virulence regulator Rot [16,54,93]. It has also been shown that inactivation of clpC in S. aureus increases long-term intracellular survival in non-professional phagocytes, suggesting that ClpC modulates intracellular persistence. This is likely to be mediated, at least in part, due to an interaction of ClpC with AgrA and ClpCP-mediated degradation of MazE. The Type II MazEF Toxin–antitoxin system is one of the best characterized in S. aureus [94]. This system is also found in other clinically important bacteria such as E. coli or P. aeuruginosa, among others [95,96,97]. Several studies were carried out to characterize the MazEF locus by studying its activation and transcriptional function [98,99,100,101,102,103]. MazEF is composed of MazF toxin, and its activity is modulated by the MazE antitoxin. Under normal growth conditions, the high level of MazE ensures the formation of the toxin–antitoxin complex and, consequently, the inactivity of MazF [104]. MazE is cleaved by the ClpCP degradation module, where ClpC is an active chaperone and ClpP is a peptidase. The degradation of MazE is assisted by the adapter protein TrfA, which provides specificity for ClpCP and facilitates the recognition of MazE [32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105].
In this regard, using a bacterial adenylate cyclase two-hybrid (BACTH) screening system in M. tuberculosis demonstrated that protein interactome ClpC1 chaperone of the ClpCP chaperone-protease complex found 196 unique interactors. Among the interactors with an assigned function, according to the Mycobrowser database classification [106], toxin–antitoxin (TA) systems are present as a major group with 24 unique TA system hits, including members of the VapBC, MazEF, and ParDE type II families [107]. Furthermore, in vitro degradation assays show that antitoxins of the VapBC and RelBE system are degraded by ClpCP [106].
On the other hand, in B. anthracis, the proteolytic activity of ClpX has been related to the degradation of antimicrobial peptides, such as cathelicidin, an antimicrobial part of the first line of the innate defense of the host. clpX mutant strains are rapidly eliminated in vitro by the antimicrobial peptides cathelicidin, α-defensin, and lysozyme [81]. It has also been suggested that ClpX participates in maintaining resistance to antimicrobial peptides LL-37 and nisine, and to the antibiotic penicillin and doptamicine [84]. Additionally, Listeria monocytogenes, a Gram-positive intracellular pathogen, possesses two ClpP isoforms, ClpP2 and ClpP1P2 (Figure 1). It was established that ClpP was essential in the growth and survival of the bacteria under stress conditions, being susceptible to the bactericidal activity of the host without the presence of these peptidases. In this sense, it was determined that ClpP2 was necessary for the expression of the virulence factor Listeriosin O, which forms pores to allow the exit of phagocytic vacuoles after the invasion of the host [108].
More recently, it has been reported that ClpB is secreted in the extracellular environment and interacts with host macrophages. When ClpB is exposed on the surface, it can mediate inflammatory immune response, which may help in maintaining the integrity of tuberculous granulomas that contain the pathogen [109]. In addition, these strains deficient in clpB are more sensitive than wild-type to isoniazid, a frontline TB drug that induces oxidative damage in mycobacteria and is attenuated in a mouse infection model [66]. ClpB has also been shown to be important in M. tuberculosis, where strains deficient in this chaperone show a defect in recovering from stationary phase or antibiotic stress in standard culture conditions. ClpB has a key role in stress tolerance and is also involved in virulence. In addition to ClpB, M. tuberculosis also encodes for other members of the Hsp100/Clp family, the proteins ClpC, ClpX, and two ClpP paralogues [31] (Figure 1 and Table 1), which are involved in both general and regulatory proteolysis [110,111]. Interestingly, it was demonstrated that clpC1, clpP1, and clpP2 are essential for the growth and virulence of M. tuberculosis [65,110]. The two clpP genes, clpP1, and clpP2 are co-expressed on a single operon that is activated by ClgR, and encode for ClpP1 and ClpP2, respectively [112,113]. Initial structure–function studies concluded that ClpP1 and ClpP2 were separate enzymes that, in isolation, formed mixtures of homo-heptameric and homo-tetradecameric complexes that lacked activity [31].
In summary, Clp is involved in virulence, exhibiting that it is necessary for the degradation of proteins involved in adaptation to the stationary phase, response to temperature stress, biofilm formation, and motility [54]. During infection, S. aureus is faced with environmental changes such as temperature, oxidative stress, antimicrobial peptides, among others. These stresses can lead to the development of misfolded proteins. The removal of these misfolded and non-native proteins is necessary for cell growth and functionality [114]. Selective degradation by the ClpP (Figure 1) and ClpQ proteases remove damaged proteins and regulate the abundance of functional proteins such as stress-induced transcriptional regulators [115,116].
Finally, ClpQ is present in Gram-positive bacteria, but contrasting to its counterpart in E. coli, it does not possess a catalytic triad of the threonine type, but biochemical studies suggest that it is a serine peptidase as well as ClpP,; unlike ClpP that forms heptamers, ClpQ forms hexamers, which produce a different interaction between the peptidase and the chaperones. Recent studies have shown a defect in motility by swarming and swimming in a mutant of the clpY and clpQ gene in B. subtilis. They also observed that the mutant showed a significant reduction in the production of γ-PGA, which has been related to the formation of biofilms and the colonization of surfaces. It was also suggested that ClpYQ has a dispensable role in the response to heat shock in both B. subtilis and B. cereus [10].
4. Clp Family as a Target against Bacteria: Focus on ClpP
Various studies have shown that both uncontrolled activation and inhibition of ClpP peptidase affect different aspects of bacterial pathogenesis (Figure 3). In this regard, ClpP has generated great interest, emerging as a potentially viable option as a new drug target [7,15].
4.1. Inhibition of ClpP
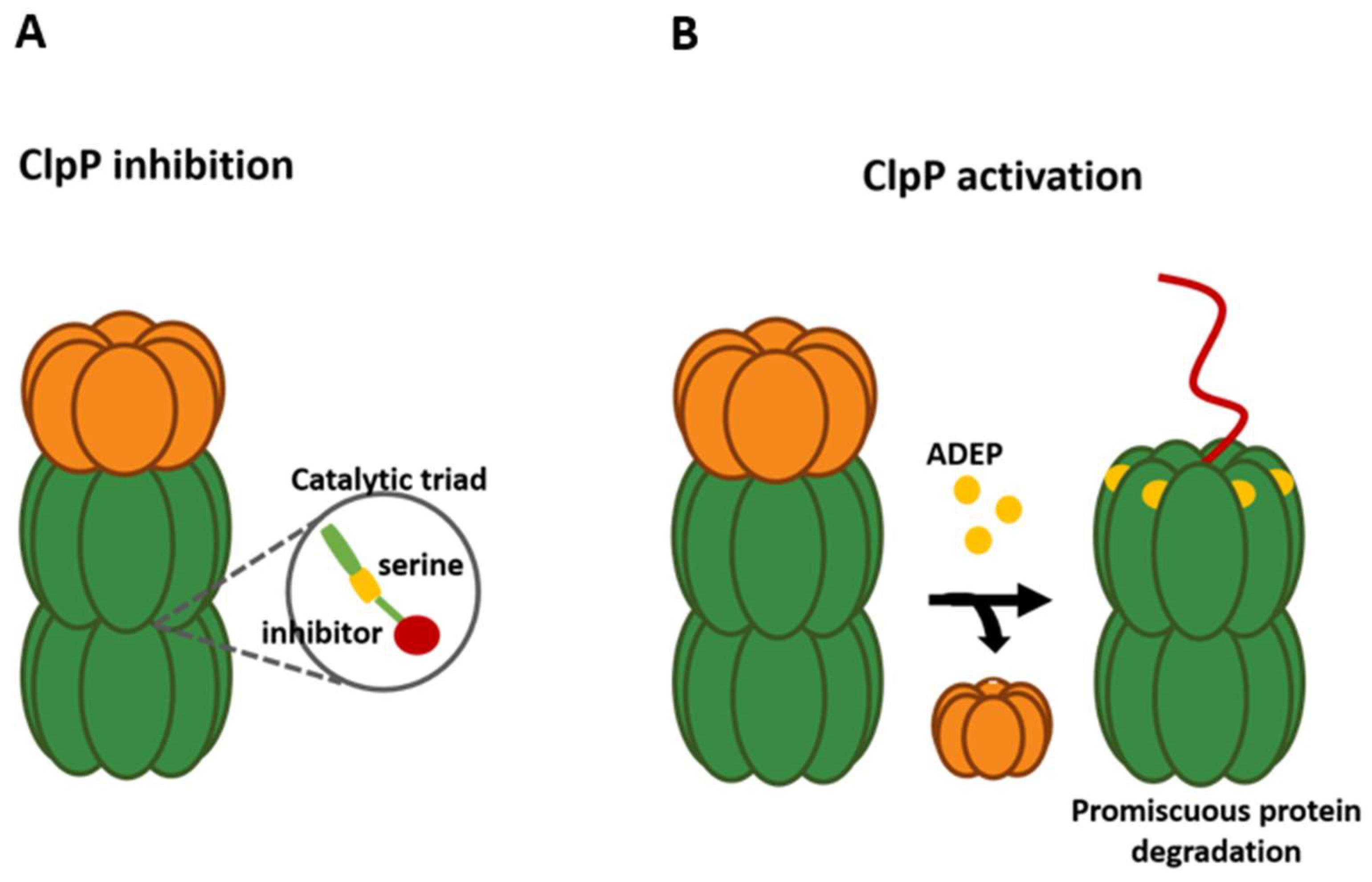
Many pathways to alter the function of the ClpP complex have been explored; for example, the specific inhibition of ClpP has been achieved with the use of β-lactone, phenyl esters, and β-sultams, all compounds which bind to the active site in an irreversible way (Figure 3A). β-lactones, for their part, are effective against methicillin-resistant S. aureus, where compound D3 generates a decrease in virulence factors, such as hemolysins and proteases, and affects the ability to generate necrosis and inflammation [116]. To optimize these compounds, U1 was synthesized, improving the efficiency between 3 to 5 times and being able to inhibit not only ClpP of S. aureus but also of L. monocytogenes, where a decrease in the synthesis of listeriolysin O and phospholipase was observed [117,118,119]. Other analogs were effective against ClpP1/P2 of M. tuberculosis [120]. Despite the effectiveness of β-lactones, their clinical use is limited due to their plasma instability and their low selectivity. In this sense, phenyl esters arose, which have greater efficiency and plasma stability than β-lactones. However, as a result of this, their activity against virulent phenotypes was significantly lower and their inhibitory activity was not maintained [121]. On the other hand, the use of β-sultams demonstrated a new way of inhibiting peptidases, since they can modify the serine of the catalytic site of ClpP, preventing nucleophilic attack [122]. Another compound that can inhibit ClpP is the boron-derived compound Bortezomib, a human 26S proteasome drug that is used for cancer and has shown an effective inhibition of ClpP1/P2 of M. tuberculosis by covalently binding to the Ser98 active site through its boron atom. However, its high cost and human proteasome inhibition preclude its use as a treatment [123].
On the other hand, the first non-covalent binding inhibitor to ClpP was developed, compound AV145, which does not bind to the active site, but to the management region of each monomer, which induces an inactive conformation of the tetradecameric complex [124]. However, its action is not effective since it does not inhibit the binding of ClpX with ClpP and, therefore, its effects are not considerable [124]. In addition, its high cost and proteasome inhibition prevented its use as a treatment [123]. Although the way to inhibit ClpP has been extensively explored, all the aforementioned compounds are effective against Gram-positive bacteria, while the inhibition of ClpP in Gram-negative bacteria has not been explored. Chloromethyl ketone is one of the compounds studied in E. coli, and it has been reported that it is capable of inhibiting ClpP. Recently, a new class of inhibitor was reported: the compound α-amino diarylphosphonate, which can bind irreversibly to ClpP [15,124,125].
4.2. ClpP Modulation
Currently, the most successful strategy is to target the interaction between ClpP and its ATPases through allosteric activation of ClpP, leading to uncontrolled and unregulated proteolysis (Figure 3B). A group of compounds of the acyldepsipeptide family (ADEP) was shown to be capable of activating the peptidase function in a deregulated manner, because this molecule competes with ATPases for the same site of binding to ClpP, managing to displace it, which produces a loss of regulatory function provided by chaperones. In addition, ADEPs mimicking the binding of chaperones can open the axial pore of the protease, activating it and allowing it to indiscriminately degrade different unstable and unfolded proteins or nascent ribosome chains [1,126]. With improved stability, ADEP has been shown to have potent activity against Gram-positive bacteria such as M. tuberculosis, S. aureus, and S. pneumoniae. Specifically, ADEP4 in combination with rifampicin demonstrated the ability to eradicate biofilms and eliminate persister cells of S. aureus by activating the uncontrolled proteolysis of more than 400 proteins in a mouse model, and was able to eradicate M. tuberculosis by only preventing binding between ClpX and essential ClpP1P2 [7,34,127,128]. Most studies focus on Gram-positive bacteria due to the limitations of ADEP compared to Gram-negative, mainly due to their susceptibility and limited penetration of the outer membrane. Nevertheless, studies showed that ADEP26 was potent against two Gram-negative pathogens, Neisseria meningitidis and Neisseria gonorrhoeae, while it was more potent than other ADEPs against Gram-positive bacteria. However, more studies are required on ADEP because of its toxicity and implementation for its clinical use [129]. Other activators have been discovered, such as ACP (activators of self-compartmentalized proteases) and sclerotiamide, a natural product, which have been presented to have similar activity to ADEPs in E. coli [130,131].
5. Conclusions
Fast adaptation to environmental changes ensures bacterial survival, and proteolysis represents a key cellular process in adaptation. The Clp protease system is a multi-component mechanism responsible for protein homeostasis, protein quality control, and targeted proteolysis of transcriptional regulators in bacteria. Important specific contributions of the Clp protease system to bacterial survival range from the degradation of defective, misfolded, or aggregated proteins and the general rescue of stalled ribosomes to precise regulatory control of the heat-shock response, natural competence, sporulation, motility, virulence, and life-cycle management. The identification of new approaches and/or drug targets that could reduce bacterial infection are expected to find high utility in prophylaxis, transmission, and attenuation of symptoms.
Author Contributions
Conceptualization, C.Q. and F.G.; validation, R.Á., D.P.-S. and F.G.; formal analysis, F.G.; investigation, C.Q., R.Á. and C.O.; resources, F.G.; data curation, F.D.-Y. and F.G.; writing—original draft preparation, C.Q., R.Á., C.O., F.D.-Y. and F.G.; writing—review and editing, C.Q., F.D.-Y. and F.G.; visualization, F.G.; supervision, F.G.; project administration, F.G.; funding acquisition, D.P.-S. and F.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by a grant from ANID (FONDECYT Grant 1171397 to F.G. and funded by ANID—Millennium Science Initiative Program—NCN17_093” to D.P-S & F.G.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bhandari, V.; Wong, K.S.; Zhou, J.L.; Mabanglo, M.F.; Batey, R.A.; Houry, W.A. The Role of ClpP Protease in Bacterial Pathogenesis and Human Diseases. ACS Chem. Biol. 2018, 13, 1413–1425. [Google Scholar] [CrossRef] [PubMed]
- Striebel, F.; Kress, W.; Weber-Ban, E. Controlled destruction: AAA ATPases in protein degradation from bacteria to eukaryotes. Curr. Opin. Struct. Biol. 2009, 19, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Maurizi, M.R.; Thompson, M.W.; Singh, S.K.; Kim, S. Endopeptidase Clp: ATP-dependent Clp protease from Escherichia coli. Methods Enzymol. Proteolytic Enzym. Serine Cysteine Pept. 1994, 244, 314–331. [Google Scholar] [CrossRef]
- Goldberg, A.L.; Moerschell, R.P.; Hachung, C.; Maurizi, M.R. ATP-dependent protease La (Lon) from Escherichia coli. Methods Enzymol. Proteolytic Enzym. Serine Cysteine Pept. 1994, 244, 350–375. [Google Scholar] [CrossRef]
- Frees, D.; Savijoki, K.; Varmanen, P.; Ingmer, H. Clp ATPases and ClpP proteolytic complexes regulate vital biological processes in low GC, Gram-positive bacteria. Mol. Microbiol. 2007, 63, 1285–1295. [Google Scholar] [CrossRef] [PubMed]
- Kirstein, J.; Molière, N.; Dougan, D.A.; Turgay, K. Adapting the machine: Adaptor proteins for Hsp100/Clp and AAA proteases. Nat. Rev. Microbiol. 2009, 7, 589–599. [Google Scholar] [CrossRef]
- Brötz-Oesterhelt, H.; Beyer, D.; Kroll, H.P.; Endermann, R.; Ladel, C.; Schroeder, W.; Hinzen, B.; Raddatz, S.; Paulsen, H.; Henninger, K.; et al. Dysregulation of bacterial proteolytic machinery by a new class of antibiotics. Nat. Med. 2005, 11, 1082–1087. [Google Scholar] [CrossRef]
- Molière, N.; Hoßmann, J.; Schäfer, H.; Turgay, K. Role of Hsp100/Clp Protease Complexes in Controlling the Regulation of Motility in Bacillus subtilis. Front. Microbiol. 2016, 7, 315. [Google Scholar] [CrossRef] [Green Version]
- Kress, W.; Mutschler, H.; Weber-Ban, E. Both ATPase Domains of ClpA Are Critical for Processing of Stable Protein Structures. J. Biol. Chem. 2009, 284, 31441–31452. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Yan, F.; He, Y.; Qin, Y.; Chen, Y.; Chai, Y.; Guo, J.H. The ClpY-ClpQ protease regulates multicellular development in Bacillus subtilis. Microbiology 2018, 164, 848–862. [Google Scholar] [CrossRef]
- Schirmer, E.C.; Glover, J.R.; Singer, M.A.; Lindquist, S. HSP100/Clp proteins: A common mechanism explains diverse functions. Trends Biochem. Sci. 1996, 21, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Lemos, J.A.; Burne, R.A. Regulation and Physiological Significance of ClpC and ClpP in Streptococcus mutans. J. Bacteriol. 2002, 184, 6357–6366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottesman, S.; Maurizi, M.R.; Wickner, S. Regulatory Subunits of Energy-Dependent Proteases. Cell 1997, 91, 435–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruger, E. Clp-mediated proteolysis in Gram-positive bacteria is autoregulated by the stability of a repressor. EMBO J. 2001, 20, 852–863. [Google Scholar] [CrossRef]
- Moreno-Cinos, C.; Goossens, K.; Salado, I.G.; Veken, P.V.; Winter, H.D.; Augustyns, K. ClpP Protease, a Promising Antimicrobial Target. Int. J. Mol. Sci. 2019, 20, 2232. [Google Scholar] [CrossRef] [Green Version]
- Frees, D.; Thomsen, L.E.; Ingmer, H. Staphylococcus aureus ClpYQ plays a minor role in stress survival. Arch. Microbiol. 2005, 183, 286–291. [Google Scholar] [CrossRef]
- Kruger, E.; Witt, E.; Ohlmeier, S.; Hanschke, R.; Hecker, M. The Clp Proteases of Bacillus subtilis Are Directly Involved in Degradation of Misfolded Proteins. J. Bacteriol. 2000, 182, 3259–3265. [Google Scholar] [CrossRef] [Green Version]
- Akopian, T.; Kandror, O.; Raju, R.M.; Unnikrishnan, M.; Rubin, E.J.; Goldberg, A.L. The active ClpP protease from M. tuberculosis is a complex composed of a heptameric ClpP1 and a ClpP2 ring. EMBO J. 2012, 31, 1529–1541. [Google Scholar] [CrossRef] [Green Version]
- Hall, B.M.; Breidenstein, E.B.; Fuente-Núñez, C.D.; Reffuveille, F.; Mawla, G.D.; Hancock, R.E.; Baker, T.A. Two Isoforms of Clp Peptidase in Pseudomonas aeruginosa Control Distinct Aspects of Cellular Physiology. J. Bacteriol. 2017, 199, e00568-16. [Google Scholar] [CrossRef] [Green Version]
- Lavey, N.P.; Shadid, T.; Ballard, J.D.; Duerfeldt, A.S. Clostridium difficile ClpP Homologues are Capable of Uncoupled Activity and Exhibit Different Levels of Susceptibility to Acyldepsipeptide Modulation. ACS Infect. Dis. 2019, 5, 79–89. [Google Scholar] [CrossRef]
- Olivares, A.O.; Baker, T.A.; Sauer, R.T. Mechanical Protein Unfolding and Degradation. Annu. Rev. Physiol. 2018, 80, 413–429. [Google Scholar] [CrossRef] [PubMed]
- Gerth, U.; Kock, H.; Kusters, I.; Michalik, S.; Switzer, R.L.; Hecker, M. Clp-Dependent Proteolysis Down-Regulates Central Metabolic Pathways in Glucose-Starved Bacillus subtilis. J. Bacteriol. 2007, 190, 321–331. [Google Scholar] [CrossRef] [Green Version]
- Frees, D.; Gerth, U.; Ingmer, H. Clp chaperones and proteases are central in stress survival, virulence and antibiotic resistance of Staphylococcus aureus. Int. J. Med. Microbiol. 2014, 304, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Ujiie, H.; Matsutani, T.; Tomatsu, H.; Fujihara, A.; Ushida, C.; Miwa, Y.; Fujita, Y.; Himeno, H.; Muto, A. Trans-Translation is Involved in the CcpA-Dependent Tagging and Degradation of TreP in Bacillus subtilis. J. Biochem. 2008, 145, 59–66. [Google Scholar] [CrossRef]
- Sauer, R.T.; Baker, T.A. AAA Proteases: ATP-Fueled Machines of Protein Destruction. Annu. Rev. Biochem. 2011, 80, 587–612. [Google Scholar] [CrossRef] [PubMed]
- Schlothauer, T.; Mogk, A.; Dougan, D.A.; Bukau, B.; Turgay, K. MecA, an adaptor protein necessary for ClpC chaperone activity. Proc. Natl. Acad. Sci. USA 2003, 100, 2306–2311. [Google Scholar] [CrossRef] [Green Version]
- Turgay, K.; Hahn, J.; Burghoorn, J.; Dubnau, D. Competence in Bacillus subtilis is controlled by regulated proteolysis of a transcription factor. EMBO J. 1998, 17, 6730–6738. [Google Scholar] [PubMed] [Green Version]
- Pan, Q.; Garsin, D.A.; Losick, R. Self-Reinforcing Activation of a Cell-Specific Transcription Factor by Proteolysis of an Anti-σ Factor in B. subtilis. Mol. Cell 2001, 8, 873–883. [Google Scholar] [CrossRef]
- Nakano, S.; Zheng, G.; Nakano, M.M.; Zuber, P. Multiple Pathways of Spx (YjbD) Proteolysis in Bacillus subtilis. J. Bacteriol. 2002, 184, 3664–3670. [Google Scholar] [CrossRef] [Green Version]
- Miethke, M.; Hecker, M.; Gerth, U. Involvement of Bacillus subtilis ClpE in CtsR Degradation and Protein Quality Control. J. Bacteriol. 2006, 188, 4610–4619. [Google Scholar] [CrossRef]
- Benaroudj, N.; Raynal, B.; Miot, M.; Ortiz-Lombardia, M. Assembly and proteolytic processing of mycobacterial ClpP1 and ClpP2. BMC Biochem. 2011, 12, 61. [Google Scholar] [CrossRef] [PubMed]
- Donegan, N.P.; Marvin, J.S.; Cheung, A.L. Role of adaptor TrfA and ClpPC in controlling levels of SsrA-tagged proteins and antitoxins in Staphylococcus aureus. J. Bacteriol. 2014, 196, 4140–4151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirstein, J.; Schlothauer, T.; Dougan, D.A.; Lilie, H.; Tischendorf, G.; Mogk, A.; Bukau, B.; Turgay, K. Adaptor protein controlled oligomerization activates the AAA protein ClpC. EMBO J. 2006, 25, 1481–1491. [Google Scholar] [CrossRef] [Green Version]
- Famulla, K.; Sass, P.; Malik, I.; Akopian, T.; Kandror, O.; Alber, M.; Hinzen, B.; Ruebsamen-Schaeff, H.; Kalscheuer, R.; Goldberg, A.L.; et al. Acyldepsipeptide antibiotics kill mycobacteria by preventing the physiological functions of the ClpP1P2 protease. Mol. Microbiol. 2016, 101, 194–209. [Google Scholar] [CrossRef] [Green Version]
- Leodolter, J.; Warweg, J.; Weber-Ban, E. The Mycobacterium tuberculosis ClpP1P2 Protease Interacts Asymmetrically with Its ATPase Partners ClpX and ClpC1. PLoS ONE 2015, 10, e0125345. [Google Scholar] [CrossRef]
- Schmitz, K.R.; Sauer, R.T. Substrate delivery by the AAA ClpX and ClpC1 unfoldases activates the mycobacterial ClpP1P2 peptidase. Mol. Microbiol. 2014, 93, 617–628. [Google Scholar] [CrossRef] [Green Version]
- Zeiler, E.; List, A.; Alte, F.; Gersch, M.; Wachtel, R.; Poreba, M.; Drag, M.; Groll, M.; Sieber, S.A. Structural and functional insights into caseinolytic proteases reveal an unprecedented regulation principle of their catalytic triad. Proc. Natl. Acad. Sci. USA 2013, 110, 11302–11307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahmen, M.; Vielberg, M.; Groll, M.; Sieber, S.A. Structure and Mechanism of the Caseinolytic Protease ClpP1/2 Heterocomplex from Listeria monocytogenes. Angew. Chem. Int. Ed. 2015, 54, 3598–3602. [Google Scholar] [CrossRef]
- Pan, S.; Malik, I.T.; Thomy, D.; Henrichfreise, B.; Sass, P. The functional ClpXP protease of Chlamydia trachomatis requires distinct clpP genes from separate genetic loci. Sci. Rep. 2019, 9, 14129. [Google Scholar] [CrossRef] [Green Version]
- Sekulovic, O.; Fortier, L. Global Transcriptional Response of Clostridium difficile Carrying the ϕCD38-2 Prophage. Appl. Environ. Microbiol. 2014, 81, 1364–1374. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.M.; Hahn, E.; Zuber, P. Adaptor bypass mutations of Bacillus subtilis spxsuggest a mechanism for YjbH-enhanced proteolysis of the regulator Spx by ClpXP. Mol. Microbiol. 2014, 93, 426–438. [Google Scholar] [CrossRef] [Green Version]
- Leelakriangsak, M.; Kobayashi, K.; Zuber, P. Dual Negative Control of spx Transcription Initiation from the P3 Promoter by Repressors PerR and YodB in Bacillus subtilis. J. Bacteriol. 2006, 189, 1736–1744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Runde, S.; Molière, N.; Heinz, A.; Maisonneuve, E.; Janczikowski, A.; Elsholz, A.K.; Gerth, U.; Hecker, M.; Turgay, K. The role of thiol oxidative stress response in heat-induced protein aggregate formation during thermotolerance in Bacillus subtilis. Mol. Microbiol. 2014, 91, 1036–1052. [Google Scholar] [CrossRef] [PubMed]
- Garg, S.K.; Kommineni, S.; Henslee, L.; Zhang, Y.; Zuber, P. The YjbH Protein of Bacillus subtilis Enhances ClpXP-Catalyzed Proteolysis of Spx. J. Bacteriol. 2008, 191, 1268–1277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engman, J.; Wachenfeldt, C.V. Regulated protein aggregation: A mechanism to control the activity of the ClpXP adaptor protein YjbH. Mol. Microbiol. 2014, 95, 51–63. [Google Scholar] [CrossRef]
- Elsholz, A.K.; Michalik, S.; Zühlke, D.; Hecker, M.; Gerth, U. CtsR, the Gram-positive master regulator of protein quality control, feels the heat. EMBO J. 2010, 29, 3621–3629. [Google Scholar] [CrossRef] [Green Version]
- Kirstein, J.; Dougan, D.A.; Gerth, U.; Hecker, M.; Turgay, K. The tyrosine kinase McsB is a regulated adaptor protein for ClpCP. EMBO J. 2007, 26, 2061–2070. [Google Scholar] [CrossRef] [Green Version]
- Fuhrmann, J.; Schmidt, A.; Spiess, S.; Lehner, A.; Turgay, K.; Mechtler, K.; Charpentier, E.; Clausen, T. McsB Is a Protein Arginine Kinase That Phosphorylates and Inhibits the Heat-Shock Regulator CtsR. Science 2009, 324, 1323–1327. [Google Scholar] [CrossRef] [Green Version]
- Elsholz AK, W.; Hempel, K.; Michalik, S.; Gronau, K.; Becher, D.; Hecker, M.; Gerth, U. Activity control of the ClpC adaptor McsB in Bacillus subtilis. J. Bacteriol. 2011, 193, 3887–3893. [Google Scholar] [CrossRef] [Green Version]
- Hill, N.S.; Zuke, J.D.; Buske, P.J.; Chien, A.; Levin, P.A. A nutrient-dependent division antagonist is regulated post-translationally by the Clp proteases in Bacillus subtilis. BMC Microbiol. 2018, 18, 29. [Google Scholar] [CrossRef]
- Renzoni, A.; Kelley, W.L.; Barras, C.; Monod, A.; Huggler, E.; François, P.; Schrenzel, J.; Studer, R.; Vaudaux, P.; Lew, D.P. Identification by genomic and genetic analysis of two new genes playing a key role in intermediate glycopeptide resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 2009, 53, 903–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.; Michalik, S.; Varming, A.N.; Andersen, J.H.; Albrecht, D.; Jelsbak, L.; Krieger, S.; Ohlsen, K.; Hecker, M.; Gerth, U.; et al. Trapping and proteomic identification of cellular substrates of the ClpP protease in Staphylococcus aureus. J. Proteome Res. 2013, 12, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Wozniak, D.J.; Tiwari, K.B.; Soufan, R.; Jayaswal, R.K. The mcsB gene of the clpC operon is required for stress tolerance and virulence in Staphylococcus aureus. Microbiology 2012, 158 Pt 10, 2568. [Google Scholar] [CrossRef] [Green Version]
- Frees, D.; Qazi, S.N.; Hill, P.J.; Ingmer, H. Alternative roles of ClpX and ClpP in Staphylococcus aureus stress tolerance and virulence. Mol. Microbiol. 2003, 48, 1565–1578. [Google Scholar] [CrossRef] [PubMed]
- Frees, D.; Chastanet, A.; Qazi, S.; Sørensen, K.; Hill, P.; Msadek, T.; Ingmer, H. Clp ATPases are required for stress tolerance, intracellular replication and biofilm formation in Staphylococcus aureus. Mol. Microbiol. 2004, 54, 1445–1462. [Google Scholar] [CrossRef]
- Frees, D.; Andersen, J.H.; Hemmingsen, L.; Koskenniemi, K.; Bæk, K.T.; Muhammed, M.K.; Gudeta, D.D.; Nyman, T.A.; Sukura, A.; Varmanen, P.; et al. New insights into Staphylococcus aureus stress tolerance and virulence regulation from an analysis of the role of the ClpP protease in the strains Newman, COL, and SA564. J. Proteome Res. 2012, 11, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Glover, J.R.; Lindquist, S. Hsp104, Hsp70, and Hsp40: A novel chaperone system that rescues previously aggregated proteins. Cell 1998, 94, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Calloni, G.; Chen, T.; Schermann, S.M.; Chang, H.C.; Genevaux, P.; Agostini, F.; Tartaglia, G.G.; Hayer-Hartl, M.; Hartl, F.U. DnaK Functions as a Central Hub in the E. coli Chaperone Network. Cell Rep. 2012, 1, 251–264. [Google Scholar] [CrossRef] [Green Version]
- Mogk, A. Identification of thermolabile Escherichia coli proteins: Prevention and reversion of aggregation by DnaK and ClpB. EMBO J. 1999, 18, 6934–6949. [Google Scholar] [CrossRef]
- Ziȩtkiewicz, S.; Krzewska, J.; Liberek, K. Successive and Synergistic Action of the Hsp70 and Hsp100 Chaperones in Protein Disaggregation. J. Biol. Chem. 2004, 279, 44376–44383. [Google Scholar] [CrossRef]
- DeJesus, M.A.; Gerrick, E.R.; Xu, W.; Park, S.W.; Long, J.E.; Boutte, C.C.; Rubin, E.J.; Schnappinger, D.; Ehrt, S.; Fortune, S.M.; et al. Comprehensive Essentiality Analysis of the Mycobacterium tuberculosis Genome via Saturating Transposon Mutagenesis. mBio 2017, 8, e02133-16. [Google Scholar] [CrossRef] [Green Version]
- Fay, A.; Glickman, M.S. An Essential Nonredundant Role for Mycobacterial DnaK in Native Protein Folding. PLoS Genet. 2014, 10, e1004516. [Google Scholar] [CrossRef] [PubMed]
- Lupoli, T.J.; Fay, A.; Adura, C.; Glickman, M.S.; Nathan, C.F. Reconstitution of a Mycobacterium tuberculosis proteostasis network highlights essential cofactor interactions with chaperone DnaK. Proc. Natl. Acad. Sci. USA 2016, 113, E7947–E7956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupoli, T.J.; Vaubourgeix, J.; Burns-Huang, K.; Gold, B. Targeting the Proteostasis Network for Mycobacterial Drug Discovery. ACS Infect. Dis. 2018, 4, 478–498. [Google Scholar] [CrossRef] [Green Version]
- Sassetti, C.M.; Boyd, D.H.; Rubin, E.J. Genes required for mycobacterial growth defined by high density mutagenesis. Mol. Microbiol. 2003, 48, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Vaubourgeix, J.; Lin, G.; Dhar, N.; Chenouard, N.; Jiang, X.; Botella, H.; Lupoli, T.; Mariani, O.; Yang, G.; Ouerfelli, O.; et al. Stressed Mycobacteria Use the Chaperone ClpB to Sequester Irreversibly Oxidized Proteins Asymmetrically within and between Cells. Cell Host Microbe 2015, 17, 178–190. [Google Scholar] [CrossRef] [Green Version]
- Kwiatkowska, J.; Matuszewska, E.; Kuczyńska-Wiśnik, D.; Laskowska, E. Aggregation of Escherichia coli proteins during stationary phase depends on glucose and oxygen availability. Res. Microbiol. 2008, 159, 651–657. [Google Scholar] [CrossRef]
- Maisonneuve, E.; Ezraty, B.; Dukan, S. Protein Aggregates: An Aging Factor Involved in Cell Death. J. Bacteriol. 2008, 190, 6070–6075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llorens, J.M.; Tormo, A.; Martínez-García, E. Stationary phase in gram-negative bacteria. FEMS Microbiol. Rev. 2010, 34, 476–495. [Google Scholar] [CrossRef] [Green Version]
- Josefson, R.; Andersson, R.; Nyström, T. How and why do toxic conformers of aberrant proteins accumulate during ageing? Essays Biochem. 2017, 61, 317–324. [Google Scholar] [CrossRef]
- Yüksel, M.; Power, J.J.; Ribbe, J.; Volkmann, T.; Maier, B. Fitness Trade-Offs in Competence Differentiation of Bacillus subtilis. Front. Microbiol. 2016, 7, 888. [Google Scholar] [CrossRef] [Green Version]
- Hahn, J.; Maier, B.; Haijema, B.J.; Sheetz, M.; Dubnau, D. Transformation Proteins and DNA Uptake Localize to the Cell Poles in Bacillus subtilis. Cell 2005, 122, 59–71. [Google Scholar] [CrossRef] [Green Version]
- Persuh, M.; Turgay, K.; Mandic-Mulec, I.; Dubnau, D. The N- and C-terminal domains of MecA recognize different partners in the competence molecular switch. Mol. Microbiol. 1999, 33, 886–894. [Google Scholar] [CrossRef]
- Liu, J.; Zuber, P. A Molecular Switch Controlling Competence and Motility: Competence Regulatory Factors ComS, MecA, and ComK Control ςD-Dependent Gene Expression in Bacillus subtilis. J. Bacteriol. 1998, 180, 4243–4251. [Google Scholar] [CrossRef] [Green Version]
- Márquez, L.M.; Helmann, J.D.; Ferrari, E.; Parker, H.M.; Ordal, G.W.; Chamberlin, M.J. Studies of sigma D-dependent functions in Bacillus subtilis. J. Bacteriol. 1990, 172, 3435–3443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K. Gradual activation of the response regulator DegU controls serial expression of genes for flagellum formation and biofilm formation in Bacillus subtilis. Mol. Microbiol. 2007, 66, 395–409. [Google Scholar] [CrossRef] [PubMed]
- Tsukahara, K.; Ogura, M. Promoter selectivity of the Bacillus subtilis response regulator DegU, a positive regulator of the fla/che operon and sacB. BMC Microbiol. 2008, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, Y.; Norman, T.; Kolter, R.; Losick, R. An epigenetic switch governing daughter cell separation in Bacillus subtilis. Genes Dev. 2010, 24, 754–765. [Google Scholar] [CrossRef] [Green Version]
- Tan, I.; Weiss, C.; Popham, D.; Ramamurthi, K. A Quality-Control Mechanism Removes Unfit Cells from a Population of Sporulating Bacteria. Dev. Cell 2015, 34, 682–693. [Google Scholar] [CrossRef] [Green Version]
- Pan, Q.; Losick, R. Unique Degradation Signal for ClpCP in Bacillus subtilis. J. Bacteriol. 2003, 185, 5275–5278. [Google Scholar] [CrossRef]
- McGillivray, S.M.; Ebrahimi, C.M.; Fisher, N.; Sabet, M.; Zhang, D.X.; Chen, Y.; Haste, N.M.; Aroian, R.V.; Gallo, R.L.; Guiney, D.G.; et al. ClpX Contributes to Innate Defense Peptide Resistance and Virulence Phenotypes of Bacillus anthracis. J. Innate Immun. 2009, 1, 494–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, L.K.; Dhasmana, N.; Sajid, A.; Kumar, P.; Bhaduri, A.; Bharadwaj, M.; Gandotra, S.; Kalia, V.C.; Das, T.K.; Goel, A.K.; et al. ClpC operon regulates cell architecture and sporulation in Bacillus anthracis. Environ. Microbiol. 2014, 17, 855–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Msadek, T.; Kunst, F.; Rapoport, G. MecB of Bacillus subtilis, a member of the ClpC ATPase family, is a pleiotropic regulator controlling competence gene expression and growth at high temperature. Proc. Natl. Acad. Sci. USA 1994, 91, 5788–5792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, L.; Evans, C.R.; Do, V.D.; Losefsky, Q.P.; Ngo, D.Q.; Mcgillivray, S.M. Loss of the ClpXP Protease Leads to Decreased Resistance to Cell-Envelope Targeting Antimicrobials in Bacillus anthracis Sterne. Front. Microbiol. 2021, 12, 719548. [Google Scholar] [CrossRef]
- Fedhila, S.; Msadek, T.; Nel P y Lereclus, D. Distintos genes clpP controlan respuestas adaptativas específicas en Bacillus thuringiensis. Rev. Bacteriol. 2002, 184, 5554–5562. [Google Scholar] [CrossRef] [Green Version]
- Bishop, C.E.; Shadid, T.M.; Lavey, N.P.; Kempher, M.L.; Ballard, J.D.; Duerfeldt, A.S. Identification of ClpP Dual Isoform Disruption as an Antisporulation Strategy for Clostridioides difficile. J. Bacteriol. 2022, 204, e00411-21. [Google Scholar] [CrossRef]
- Emerson, J.E.; Stabler, R.A.; Wren, B.W.; Fairweather, N.F. Microarray analysis of the transcriptional responses of Clostridium difficile to environmental and antibiotic stress. J. Med. Microbiol. 2008, 57, 757–764. [Google Scholar] [CrossRef]
- Jain, S.; Graham, R.L.; Mcmullan, G.; Ternan, N.G. Proteomic analysis of the insoluble subproteome of Clostridium difficile strain 630. FEMS Microbiol. Lett. 2010, 312, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawley, T.D.; Croucher, N.J.; Yu, L.; Clare, S.; Sebaihia, M.; Goulding, D.; Pickard, D.J.; Parkhill, J.; Choudhary, J.; Dougan, G. Proteomic and Genomic Characterization of Highly Infectious Clostridium difficile 630 Spores. J. Bacteriol. 2009, 191, 5377–5386. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, I.; Becker, P.; Grundmeier, M.; Bischoff, M.; Somerville, G.A.; Peters, G.; Sinha, B.; Harraghy, N.; Proctor, R.A.; Herrmann, M. Staphylococcus aureus ClpC is required for stress resistance, aconitase activity, growth recovery, and death. J. Bacteriol. 2005, 187, 4488–4496. [Google Scholar] [CrossRef]
- Sun, F.; Zhou, L.; Zhao, B.C.; Deng, X.; Cho, H.; Yi, C.; Jian, X.; Song, C.X.; Luan, C.H.; Bae, T.; et al. Targeting MgrA-mediated virulence regulation in Staphylococcus aureus. Chem. Biol. 2011, 18, 1032–1041. [Google Scholar] [CrossRef] [Green Version]
- Farrand, A.J.; Reniere, M.L.; Ingmer, H.; Frees, D.; Skaar, E.P. Regulation of host hemoglobin binding by the Staphylococcus aureus Clp proteolytic system. J. Bacteriol. 2013, 195, 5041–5050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jelsbak, L.; Ingmer, H.; Valihrach, L.; Cohn, M.T.; Christiansen, M.H.; Kallipolitis, B.H.; Frees, D. The chaperone ClpX stimulates expression of Staphylococcus aureus protein A by Rot dependent and independent pathways. PLoS ONE 2010, 5, e12752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, C.F.; Bertram, R. Toxin-antitoxin systems of Staphylococcus aureus. Toxins 2016, 8, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittenhuber, G. Occurrence of MazEF-like antitoxin/toxin systems in bacteria. J. Mol. Microbiol. Biotechnol. 1999, 1, 295–302. [Google Scholar] [PubMed]
- Nguyen, D.; Joshi-Datar, A.; Lepine, F.; Bauerle, E.; Olakanmi, O.; Beer, K.; McKay, G.; Siehnel, R.; Schafhauser, J.; Wang, Y.; et al. Active starvation responses mediate antibiotic tolerance in biofilms and nutrient-limited bacteria. Science 2011, 334, 982–986. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.; Carr, A.N.; Whitworth, L.; Johnson, B.; Wilson, K.S. MazEF toxin-antitoxin proteins alter Escherichia coli cell morphology and infrastructure during persister formation and regrowth. Microbiology 2017, 163, 308–321. [Google Scholar] [CrossRef]
- Donegan, N.P.; Cheung, A.L. Regulation of the mazEF toxin-antitoxin module in Staphylococcus aureus and its impact on sigB expression. J. Bacteriol. 2009, 191, 2795–2805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Z.; Tamber, S.; Memmi, G.; Donegan, N.P.; Cheung, A.L. Overexpression of MazFsa in Staphylococcus aureus induces bacteriostasis by selectively targeting mRNAs for cleavage. J. Bacteriol. 2009, 191, 2051–2059. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Inoue, K.; Yoshizumi, S.; Kobayashi, H.; Zhang, Y.; Ouyang, M.; Kato, F.; Sugai, M.; Inouye, M. Staphylococcus aureus MazF specifically cleaves a pentad sequence, UACAU, which is unusually abundant in the mRNA for pathogenic adhesive factor SraP. J. Bacteriol. 2009, 191, 3248–3255. [Google Scholar] [CrossRef]
- Zorzini, V.; Haesaerts, S.; Donegan, N.P.; Fu, Z.; Cheung, A.L.; van Nuland, N.A.; Loris, R. Crystallization of the Staphylococcus aureus MazF mRNA interferase. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2011, 67, 386–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorzini, V.; Buts, L.; Sleutel, M.; Garcia-Pino, A.; Talavera, A.; Haesaerts, S.; Greve, H.D.; Cheung, A.; van Nuland, N.A.; Loris, R. Structural and biophysical characterization of Staphylococcus aureus Sa MazF shows conservation of functional dynamics. Nucleic Acids Res. 2014, 42, 6709–6725. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Yokota, A.; Ota, Y.; Tsuruga, M.; Aoi, R.; Tsuneda, S.; Noda, N. Nitrosomonas europaea MazF specifically recognises the UGG motif and promotes selective RNA degradation. Front. Microbiol. 2018, 9, 2386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Z.; Donegan, N.P.; Memmi, G.; Cheung, A.L. Characterization of MazF Sa, an endoribonuclease from Staphylococcus aureus. J. Bacteriol. 2007, 189, 8871–8879. [Google Scholar] [CrossRef] [Green Version]
- Donegan, N.P.; Thompson, E.T.; Fu, Z.; Cheung, A.L. Proteolytic regulation of toxin-antitoxin systems by ClpPC in Staphylococcus aureus. J. Bacteriol. 2010, 192, 1416–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziemski, M.; Leodolter, J.; Taylor, G.; Kerschenmeyer, A.; Weber-Ban, E. Genome-wide interaction screen for Mycobacterium tuberculosis ClpCP protease reveals toxin–antitoxin systems as a major substrate class. FEBS J. 2020, 288, 99–114. [Google Scholar] [CrossRef]
- Sala, A.; Bordes, P.; Genevaux, P. Multiple Toxin-Antitoxin Systems in Mycobacterium tuberculosis. Toxins 2014, 6, 1002–1020. [Google Scholar] [CrossRef] [Green Version]
- Gaillot, O.; Pellegrini, E.; Bregenholt, S.; Nair, S.; Berche, P. The ClpP serine protease is essential for the intracellular parasitism and virulence of Listeria monocytogenes. Mol. Microbiol. 2002, 35, 1286–1294. [Google Scholar] [CrossRef]
- Tripathi, P.; Singh, L.K.; Kumari, S.; Hakiem, O.R.; Batra, J.K. ClpB is an essential stress regulator of Mycobacterium tuberculosis and endows survival advantage to dormant bacilli. Int. J. Med. Microbiol. 2020, 310, 151402. [Google Scholar] [CrossRef]
- Raju, R.M.; Unnikrishnan, M.; Rubin, D.H.; Krishnamoorthy, V.; Kandror, O.; Akopian, T.N.; Goldberg, A.L.; Rubin, E.J. Mycobacterium tuberculosis ClpP1 and ClpP2 Function Together in Protein Degradation and Are Required for Viability in vitro and During Infection. PLoS Pathog. 2012, 8, e1002511. [Google Scholar] [CrossRef]
- Raju, R.M.; Jedrychowski, M.P.; Wei, J.R.; Pinkham, J.T.; Park, A.S.; O’Brien, K.; Rehren, G.; Schnappinger, D.; Gygi, S.P.; Rubin, E.J. Post-Translational Regulation via Clp Protease Is Critical for Survival of Mycobacterium tuberculosis. PLoS Pathog. 2014, 10, e1003994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhuwaider, A.A.; Dougan, D.A. AAA Machines of Protein Destruction in Mycobacteria. Front. Mol. Biosci. 2017, 4, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Personne, Y.; Brown, A.C.; Schuessler, D.L.; Parish, T. Mycobacterium tuberculosis ClpP Proteases Are Co-transcribed but Exhibit Different Substrate Specificities. PLoS ONE 2013, 8, e60228. [Google Scholar] [CrossRef]
- Truscott, K.N.; Bezawork-Geleta, A.; Dougan, D.A. Unfolded protein responses in bacteria and mitochondria: A central role for the ClpXP machine. IUBMB Life 2011, 63, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.I.; Levchenko, I.; Fraczkowska, K.; Woodruff, R.V.; Sauer, R.T.; Baker, T.A. Molecular determinants of complex formation between Clp/Hsp100 ATPases and the ClpP peptidase. Nat. Struct. Biol. 2001, 8, 230–233. [Google Scholar] [CrossRef]
- Singh, S.K.; Rozycki, J.; Ortega, J.; Ishikawa, T.; Lo, J.; Steven, A.C.; Maurizi, M.R. Functional domains of the ClpA and ClpX molecular chaperones identified by limited proteolysis and deletion analysis. J. Biol. Chem. 2001, 276, 29420–29429. [Google Scholar] [CrossRef] [Green Version]
- Böttcher, T.; Sieber, S. β-Lactones as Privileged Structures for the Active-Site Labeling of Versatile Bacterial Enzyme Classes. Angew. Chem. Int. Ed. 2008, 47, 4600–4603. [Google Scholar] [CrossRef]
- Böttcher, T.; Sieber, S. β-Lactones Decrease the Intracellular Virulence of Listeria monocytogenesin Macrophages. ChemMedChem 2009, 4, 1260–1263. [Google Scholar] [CrossRef]
- Weinandy, F.; Lorenz-Baath, K.; Korotkov, V.S.; Böttcher, T.; Sethi, S.; Chakraborty, T.; Sieber, S.A. A β-Lactone-Based Antivirulence Drug Ameliorates Staphylococcus aureus Skin Infections in Mice. ChemMedChem 2014, 9, 710–713. [Google Scholar] [CrossRef]
- Compton, C.L.; Schmitz, K.R.; Sauer, R.T.; Sello, J.K. Antibacterial Activity of and Resistance to Small Molecule Inhibitors of the ClpP Peptidase. ACS Chem. Biol. 2013, 8, 2669–2677. [Google Scholar] [CrossRef]
- HHackl, M.W.; Lakemeyer, M.; Dahmen, M.; Glaser, M.; Pahl, A.; Lorenz-Baath, K.; Menzel, T.; Sievers, S.; Böttcher, T.; Antes, I.; et al. Phenyl Esters Are Potent Inhibitors of Caseinolytic Protease P and Reveal a Stereogenic Switch for Deoligomerization. J. Am. Chem. Soc. 2015, 137, 8475–8483. [Google Scholar] [CrossRef] [PubMed]
- Gersch, M.; Kolb, R.; Alte, F.; Groll, M.; Sieber, S.A. Disruption of Oligomerization and Dehydroalanine Formation as Mechanisms for ClpP Protease Inhibition. J. Am. Chem. Soc. 2013, 136, 1360–1366. [Google Scholar] [CrossRef] [PubMed]
- Moreira, W.; Ngan, G.J.; Low, J.L.; Poulsen, A.; Chia, B.C.; Ang, M.J.; Yap, A.; Fulwood, J.; Lakshmanan, U.; Lim, J.; et al. Target Mechanism-Based Whole-Cell Screening Identifies Bortezomib as an Inhibitor of Caseinolytic Protease in Mycobacteria. mBio 2015, 6, e00253-15. [Google Scholar] [CrossRef] [Green Version]
- Pahl, A.; Lakemeyer, M.; Vielberg, M.T.; Hackl, M.W.; Vomacka, J.; Korotkov, V.S.; Stein, M.L.; Fetzer, C.; Lorenz-Baath, K.; Richter, K.; et al. Reversible Inhibitors Arrest ClpP in a Defined Conformational State that Can Be Revoked by ClpX Association. Angew. Chem. Int. Ed. 2015, 54, 15892–15896. [Google Scholar] [CrossRef] [PubMed]
- Szyk, A.; Maurizi, M.R. Crystal structure at 1.9 Å of E. coli ClpP with a peptide covalently bound at the active site. J. Struct. Biol. 2006, 156, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Malik, I.T.; Brötz-Oesterhelt, H. Conformational control of the bacterial Clp protease by natural product antibiotics. Nat. Prod. Rep. 2017, 34, 815–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sowole, M.A.; Alexopoulos, J.A.; Cheng, Y.; Ortega, J.; Konermann, L. Activation of ClpP Protease by ADEP Antibiotics: Insights from Hydrogen Exchange Mass Spectrometry. J. Mol. Biol. 2013, 425, 4508–4519. [Google Scholar] [CrossRef]
- Conlon, B.P.; Nakayasu, E.S.; Fleck, L.E.; LaFleur, M.D.; Isabella, V.M.; Coleman, K.; Leonard, S.N.; Smith, R.D.; Adkins, J.N.; Lewis, K. Activated ClpP kills persisters and eradicates a chronic biofilm infection. Nature 2013, 503, 365–370. [Google Scholar] [CrossRef] [Green Version]
- Goodreid, J.D.; Janetzko, J.; Santa Maria Jr, J.P.; Wong, K.S.; Leung, E.; Eger, B.T.; Bryson, S.; Pai, E.F.; Gray-Owen, S.D.; Walker, S.; et al. Development and Characterization of Potent Cyclic Acyldepsipeptide Analogues with Increased Antimicrobial Activity. J. Med. Chem. 2016, 59, 624–646. [Google Scholar] [CrossRef]
- Leung, E.; Datti, A.; Cossette, M.; Goodreid, J.; McCaw, S.E.; Mah, M.; Nakhamchik, A.; Ogata, K.; El Bakkouri, M.; Cheng, Y.Q.; et al. Activators of Cylindrical Proteases as Antimicrobials: Identification and Development of Small Molecule Activators of ClpP Protease. Chem. Biol. 2011, 18, 1167–1178. [Google Scholar] [CrossRef]
- Lavey, N.P.; Coker, J.A.; Ruben, E.A.; Duerfeldt, A.S. Sclerotiamide: The First Non-Peptide-Based Natural Product Activator of Bacterial Caseinolytic Protease P. J. Nat. Prod. 2016, 79, 1193–1197. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
ClpP models in different bacteria. In green, it shows single ClpP peptidases forming 2 rings (B. subtilis and S. aureus). ClpP1 subunit in blue and ClpP2 in yellow.
Figure 1.
ClpP models in different bacteria. In green, it shows single ClpP peptidases forming 2 rings (B. subtilis and S. aureus). ClpP1 subunit in blue and ClpP2 in yellow.

Figure 2.
Composition and mechanism of the chaperone–peptidase complex. Substrates are recognized and unfolded by the hexameric chaperone (orange) using an adapter (brown). The unfolded substrate (red) is transferred into the proteolytic chamber of the ClpP (green), where proteolysis is carried out. (A) General composition of the chaperone protease complex. (B) Composition of the ClpCP chaperone–protease complex in B. subtilis.
Figure 2.
Composition and mechanism of the chaperone–peptidase complex. Substrates are recognized and unfolded by the hexameric chaperone (orange) using an adapter (brown). The unfolded substrate (red) is transferred into the proteolytic chamber of the ClpP (green), where proteolysis is carried out. (A) General composition of the chaperone protease complex. (B) Composition of the ClpCP chaperone–protease complex in B. subtilis.

Figure 3.
Mechanisms of alteration of the ATPase–ClpP complex. (A) Inhibition chaperone–peptidase complex by compounds that block the active site of ClpP by binding to serine of the catalytic triad (S98). (B) Activation of ClpP through an ADEP molecule binding produces a promiscuous protein degradation.
Figure 3.
Mechanisms of alteration of the ATPase–ClpP complex. (A) Inhibition chaperone–peptidase complex by compounds that block the active site of ClpP by binding to serine of the catalytic triad (S98). (B) Activation of ClpP through an ADEP molecule binding produces a promiscuous protein degradation.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Queraltó, C.; Álvarez, R.; Ortega, C.; Díaz-Yáñez, F.; Paredes-Sabja, D.; Gil, F. Role and Regulation of Clp Proteases: A Target against Gram-Positive Bacteria. Bacteria 2023, 2, 21-36. https://doi.org/10.3390/bacteria2010002
AMA Style
Queraltó C, Álvarez R, Ortega C, Díaz-Yáñez F, Paredes-Sabja D, Gil F. Role and Regulation of Clp Proteases: A Target against Gram-Positive Bacteria. Bacteria. 2023; 2(1):21-36. https://doi.org/10.3390/bacteria2010002
Chicago/Turabian StyleQueraltó, Camila, Ricardo Álvarez, Constanza Ortega, Fernando Díaz-Yáñez, Daniel Paredes-Sabja, and Fernando Gil. 2023. "Role and Regulation of Clp Proteases: A Target against Gram-Positive Bacteria" Bacteria 2, no. 1: 21-36. https://doi.org/10.3390/bacteria2010002