
Probiotic Regulation to Modulate Aging Gut and Brain Health: A Concise Review
by
 , , and
, , and
Mrinal Samtiya
1 ,
,
Anil Kumar Puniya
2,
Monica Puniya
3,
Nagendra P. Shah
4,
Tejpal Dhewa
1,* and
Ravichandra Vemuri
5,* 1
Department of Nutrition Biology, Central University of Haryana, Mahendergarh 123031, Haryana, India
2
Dairy Microbiology Division, ICAR-National Dairy Research Institute, Karnal 132001, Haryana, India
3
Science and Standards Division, Food Safety and Standards Authority of India (FSSAI), FDA Bhawan, Kotla Road, New Delhi 110002, India
4
Food and Nutritional Science, School of Biological Sciences, The University of Hong Kong, Pokfulam Road, Pokfulam, Hong Kong
5
Department of Pathology, Wake Forest School of Medicine, Winston-Salem, NC 27101, USA
*
Authors to whom correspondence should be addressed.
Bacteria 2022, 1(4), 250-265; https://doi.org/10.3390/bacteria1040019
Submission received: 27 May 2022
/
Revised: 6 October 2022
/
Accepted: 17 October 2022
/
Published: 2 November 2022
Abstract
:The human gastrointestinal (GI) tract contains a diverse mixture of commensal and pathogenic microbes, forming the gut microbiome. These gut microbes and their potential to improve human health are a topic of great interest to the scientific community. Many intestinal and age-related complications are linked to dysbiosis of the gut microbiome, often associated with a weakened immune system. A decrease in beneficial microbes, generally, along with decreased microbial diversity in the gut, can, in many cases, result in disease, particularly in older individuals. Probiotics, which are ingestible beneficial microorganisms, have the potential to positively modulate the indigenous gut microbiota. There are two predominant and conventional classes of lactic acid bacterial probiotics, lactobacilli and bifidobacteria, which have been confirmed for their health benefits and role in preventing certain gut-related disorders. The proper use of probiotics and/or supplements, along with a consistently healthy lifestyle, is a promising holistic approach to maintaining or improving gut health and minimizing other age-linked disorders. There are many properties that bacterial probiotics possess, which may allow for these beneficial effects in the gut. For instance, probiotics have adhesion capacities (capability to stay in GI tract) that are effective in excluding pathogens, while other probiotics have the potential to stimulate or modulate the intestinal immune system by regulating genes that reside within and outside of the gut environment. This review discussed the possible underlying mechanics of probiotics, evidence of probiotic-based mitigation of age-related disease, and the role of probiotics in modulating gut health and, in turn, maintaining brain health.
1. Introduction
Probiotics are viable microorganisms, defined as “live microorganisms that, when administered in adequate amounts, confer a health benefit on the host” [1]. Traditionally, probiotics are thought to be Gram-positive bacteria, although more recently, certain yeasts have shown some therapeutic promise. Bacillus, Bifidobacterium, Lactobacillus, Streptococcus, Enterococcus and Escherichia (strain Nissle 1917) are some genera that are frequently used as probiotics [2,3,4]. Probiotics have enhanced the curiosity of researchers due to their immense potential when used in combination with the human nutritional diet. It has been demonstrated that the consumption of probiotics promotes health by regulating the age-linked oxidative stress, which is accompanied by inflammation [5], gradual immune-system weakening (i.e., immunosenescence) [6,7], and gut microbiota disturbance (i.e., dysbiosis) [5,6,7]. Although age-linked disorders can be caused by a variety of conditions, previous studies have shown that low- or chronic-grade inflammation is a major driver of successive aging and several age-linked diseases [8,9,10]. Diminishing cognitive function, learning capacities, and memory, as well as stress-linked behaviors, are all perceived as normal in regard to the progression of aging [11]. The gradual disruption of anatomical and physiological activities leads to aging that microbes perceive due to their complex biological developments. Thus, in multicellular organisms, the gradual weakening in utilities of multiple cells and tissues is observed [12]. As per the World Health Organization (WHO), the aging population is growing worldwide, and it is predicted that the number of people above 60 years of age will almost double (12% to 22%) from 2015 to 2050, making a more thorough understanding of aging and potentially related therapeutics all the more relevant [13]. The gradual deterioration in homeostatic biological activities in the elderly increases the risk and hastens the onset of health issues, such as metabolic decline, cancer, immunosenescence, neurodegeneration, and heart-related illnesses. It is essential to find an effective solution that can regulate these concerns to ultimately attain a healthy life during aging [14].
Probiotics can influence other bacteria’s actions by producing metabolites and compounds containing antimicrobial properties and lowering the pH in such a way that hinders the growth of other microbes [15]. Several probiotics interact more intimately with the immune system by regulating cytokine expression via activating toll-like receptors [16]. The gut microbiome is composed of roughly 100 trillion diverse bacteria that are necessary for regulating host health. For host immunity, these commensal bacteria protect from pathogens and contribute significantly to the metabolism of drugs and nutrients [17].
Given their beneficial properties and tight links to the immune system, probiotics can be offered as a viable therapeutic approach against neurodegenerative illnesses by modulating the gut microbiota in such a way as to ameliorate the intestinal epithelial barrier and also regulate the progression of inflammatory functions, as well as microglial initiation [18]. The complete mechanistic function of probiotic activity that allows for their beneficial attributes has not yet been fully elucidated [19]. In certain studies, probiotics have demonstrated promising effects on host health, including maintenance of previously disrupted intestinal microbiota, maturation of the gastrointestinal tract, reduction of infectious and harmful bacteria, and enhancement of immune functions [20].
Studies have reported that probiotics with specific health-associated properties can be useful in modulating age-linked gut microbiota imbalance. The most reported beneficial attributes of probiotics are the reduction of diarrheal disorders, protection against pathogens, amelioration of intestinal epithelial barrier, improvement in gastrointestinal motility and gut disorders, and immunomodulatory functions [12]. In addition to these reported effects, probiotics also support the growth of beneficial bacteria and have been shown to regulate tight junctions and mRNA expression [21,22,23]. The primary aim of this concise review was to summarize the recent evidence of probiotics to alleviate/improve aging gut- and brain-health-related disorders. In addition, it also briefly contains the possible mechanisms and pathways of probiotics that might be involved in regulating age-linked illnesses and gut health. This concise review provides the basic information related to the up-to-date probiotics’ functioning. It could benefit emerging researchers who have just started their research careers in this specific area of probiotics.
2. Mechanism of Actions of Probiotics
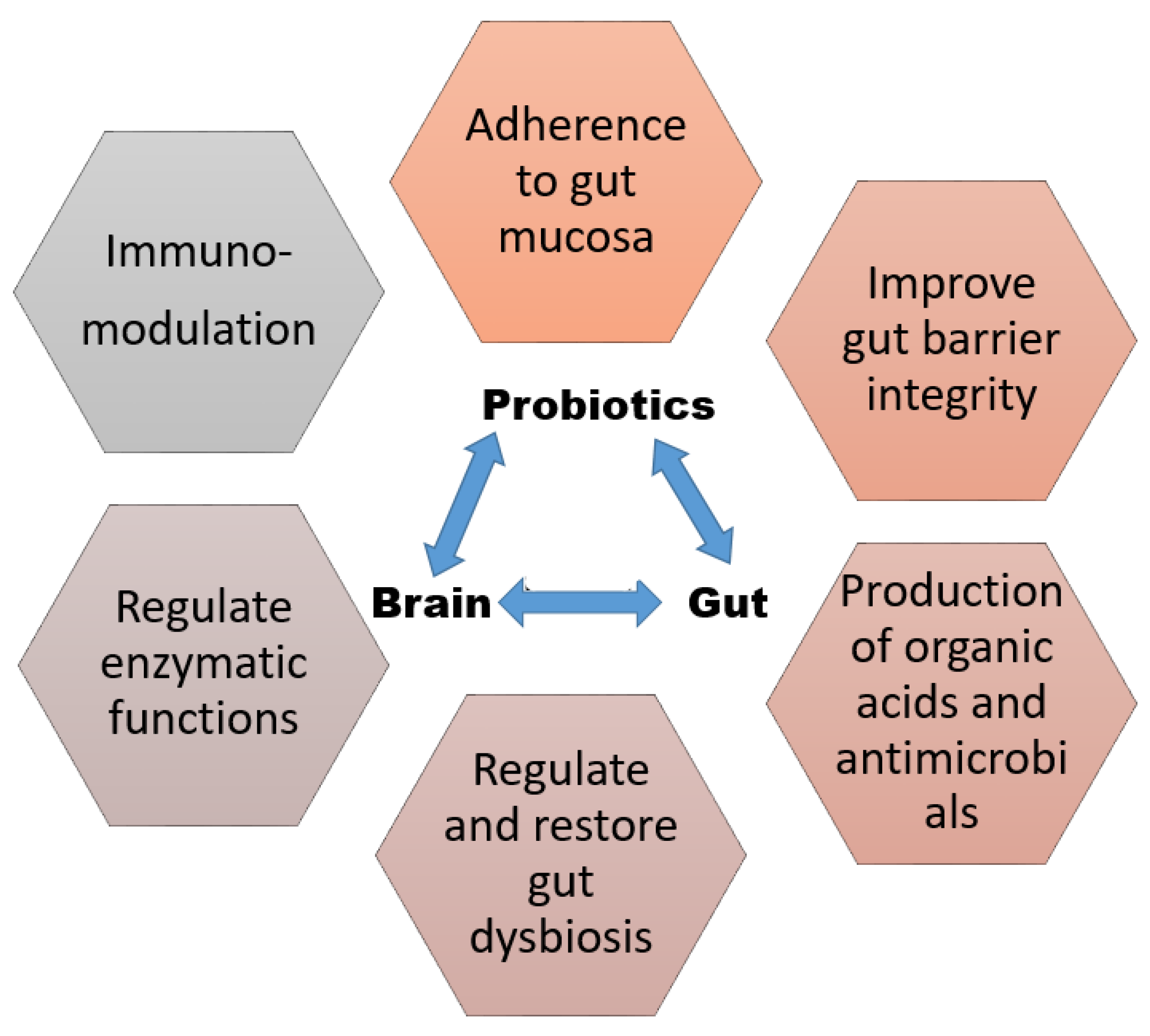
Probiotics meant for health promotion should be of known/human origin, meet safety standards, and be vector-free (as these may transmit resistance to antimicrobials and other toxicity/infectious factors). Furthermore, a potential probiotic should also be capable of surviving and possibly even growing under intestinal environmental factors, such as the acidic pH, strong enzymatic actions, incoming biliary salts, etc., [19,24]. Probiotics function via different modes of action depending upon genetic influences and their own set of metabolic activities, including immunomodulation [25], stimulation of genes related to oxidation of fatty acids, enhancement of the production of satiety-inducing peptides, improvement of serum lipid profiles [26], enzyme production, production of antimicrobials and organic acids [27], competitive exclusion via adherence to the mucosal epithelium, and improvement of the gut barrier permeability [28]. Figure 1 presents a brief overview of these probiotic-related mechanisms of action.
Most probiotics that have been previously tested in the literature are lactic acid producers (Lactobacillus and Bifidobacterium) that have already been identified as having health-associated attributes (treating/preventing/improving) concerning human disorders [29]. Somewhat unusually, lactic acid bacteria (LAB) produce exopolysaccharides (EPS) that have been reported to be involved in several activities, including immunomodulation, and have antioxidative properties, antimicrobial attributes, and significant applications in food technology for enhancing the characteristics of yogurt and cheese. Hence, most existing genetic modifications for improved galactose utilization by LAB are closely related to the production of exopolysaccharides (EPSs) for use in the food industry [30]. The concept of probiotics has been mainly focused on the modulation of the host microbiome through direct interaction with the commensal microbiota of the host. Probiotics belonging to the Lactobacillus and Bifidobacterium genera commonly produce acetic and lactic acids as their principal carbohydrate metabolism end products. When made in situ, these organic acids could decrease the luminal pH and subsequently inhibit pathogenic invasion [27]. The cell walls of probiotics contain varying constituents, such as teichoic acids, capsular polysaccharides, lipoproteins, and peptidoglycan. These cell-wall components illustrate the microbe’s associated molecular patterns, which are important to and accessed by particular host intestinal receptors, i.e., pattern-recognition receptors that regulate the immune functions to obstruct pathogens [3,31]. Nothaft et al. [32] observed that co-supplementation of Lactobacillus reuteri and Anaerosporobacter mobilis with an N-glycan-based vaccine in broiler chickens and specific-pathogen-free (SPF) leghorn chickens led to the betterment of IgY antibodies in serum (i.e., immune function) and a beneficial composition of intestinal microbiota, along with the reduction of intestinal colonization by Campylobacter jejuni. The gut microbiota interacts with mucin via diverse levels; they stimulate the proliferation of mucosal cells along with the synthesis and degradation of mucin. Furthermore, probiotics also promote the growth of goblet cells and mucin production [3]. Engevik et al. [33] reported the potential of Bifidobacterium dentium for intestinal improvement in a mouse model. Bifidobacterium dentium adheres to the intestinal mucus and releases γ-aminobutyric acid, which increases mucin production by upregulating the MUC2 functions of goblet cells. A large range of metabolites are produced by probiotics, including tryptamine, short-chain fatty acids (SCFAs), vitamins, bacteriocins, γ-aminobutyric acid (GABA), and indole. There are three SCFAs (i.e., acetate, butyrate, and propionate) that are commonly produced by the intestinal microbiome, and all are thought to be part of a healthy metabolome. Propionate, in particular, potentially limits the development of Salmonella typhimurium via disrupting the intracellular pH balance that S. typhimurium relies on [34,35]. To confer potential beneficial effects to the host, probiotics must have the ability to persist through early digestion, as well as have intestinal-adherence potential. The adherence capability of probiotics is determined by the hydrophobicity and autoaggregation ability, qualities which are inherent to each strain [36]. Within the complex and heavily populated gut microbiome, bacterial species compete with one another for the same receptor in the gastrointestinal tract and maintain the earned niches through competitive exclusion. Some of the probiotic types, such as lactobacilli and bifidobacteria, maintain their particular niches through the inhibition of the growth of particular pathogens via the production of antimicrobial peptides (i.e., bacteriocins) [19]. Huang et al. [37] assessed the attributes of Lactobacillus plantarum ZDY 2013 in relation to the modulation of gut microbiota. The results from quantitative PCR testing have shown that L. plantarum ZDY 2013 improved the abundance of Bifidobacterium and Lactobacillus populations in either the colon or cecum and, consequently, decreased populations of enteropathogenic bacteria.
Probiotic L. rhamnosus pretreatment increases the Akt phosphorylation apart from enhancing the protein expression of zonula occludens-1 and occludin. Furthermore, some probiotics demonstrate the capability to maintain the intestinal epithelial barrier and inspire epithelial cell initiation in response to bacterial infections of the intestinal tract [38]. A study involving human subjects by Mujagic et al. [39] assessed the effects of three L. plantarum strains on the small-intestinal epithelial barrier function and transcription of the gut mucosal gene. Gene transcription pathways related to cell-cell adhesion were modulated by L. plantarum TIFN101. The consumption of probiotics has been correlated with the reduction in opportunistic pathogens such as Clostridium difficile, C. cluster XI, C. perfringens, Campylobacter, and Enterococcus faecium, all of which show population increases during aging [40]. In 1907, Elie Metchnikoff reported the significant observation that individuals who regularly consumed Lactobacillus bulgaricus-containing yogurt lived longer than their counterparts. Though there has been longstanding and continued interest in the potential ameliorative effects of probiotics, there are a number of unanswered questions about the specific underlying mechanisms of probiotics that may contribute to a healthy lifespan [41,42]. Elie Metchnikoff’s report was the first evidence suggesting that probiotics improve life span longevity. Previous studies stated that the population of beneficial bacteria (i.e., lactobacilli) is reducing as the age of people increases, and the administration of lactobacilli provides health outcomes in the aged people by improving gut health and immune-modulatory actions and by regulating the composition of the gut mycobiome [43,44,45]. Probiotics may positively impact healthy aging through several mechanistic ways, such as by reducing oxidative stress, improving cognition and inflammatory responses, reducing fat accumulation, reducing pathogenic protections, and alleviating metabolic illnesses [46]. For example, a recent study reported the positive impact of probiotics on the healthy aging of Caenorhabditis elegans [46]. Results have shown that Lactobacillus plantarum JBC5 (LPJBC5) enhances longevity and improves age-related biomarkers to maintain healthy aging in Caenorhabditis elegans. LPJBC5 decreases the reactive oxygen species and improves the serotonin-signaling gene expressions (maintains cognitive function) and mitochondria function (decreases apoptosis). Moreover, LPJBC5 increased the tight junction proteins that maintain gut barrier integrity and decreased fat accumulation by downregulating the gene-encoding enzymes and substrates of fat metabolism. Overall study results concluded that LPJBC5 improved healthy aging, cognitive functions, and gut barrier health [46]. In elderly persons, the decline of the immune system is linked to increased production of proinflammatory cytokines (i.e., inflammaging) [47,48]. Probiotics can protect the host’s immune system via stimulation, modulation, and regulation of specific immune functions. Intestinal mucosal immunity is the key factor to look at when it comes to gut-linked immunomodulation [49,50]. For instance, L. fermentum MTCC 5898, administered orally to mice for 28 days, substantially decreased the IFN-γ (i.e., proinflammatory cytokine) levels and significantly increased the TGF-β (i.e., regulatory cytokine) levels in intestinal fluid [23].
3. Gut Health Modulation by Probiotics
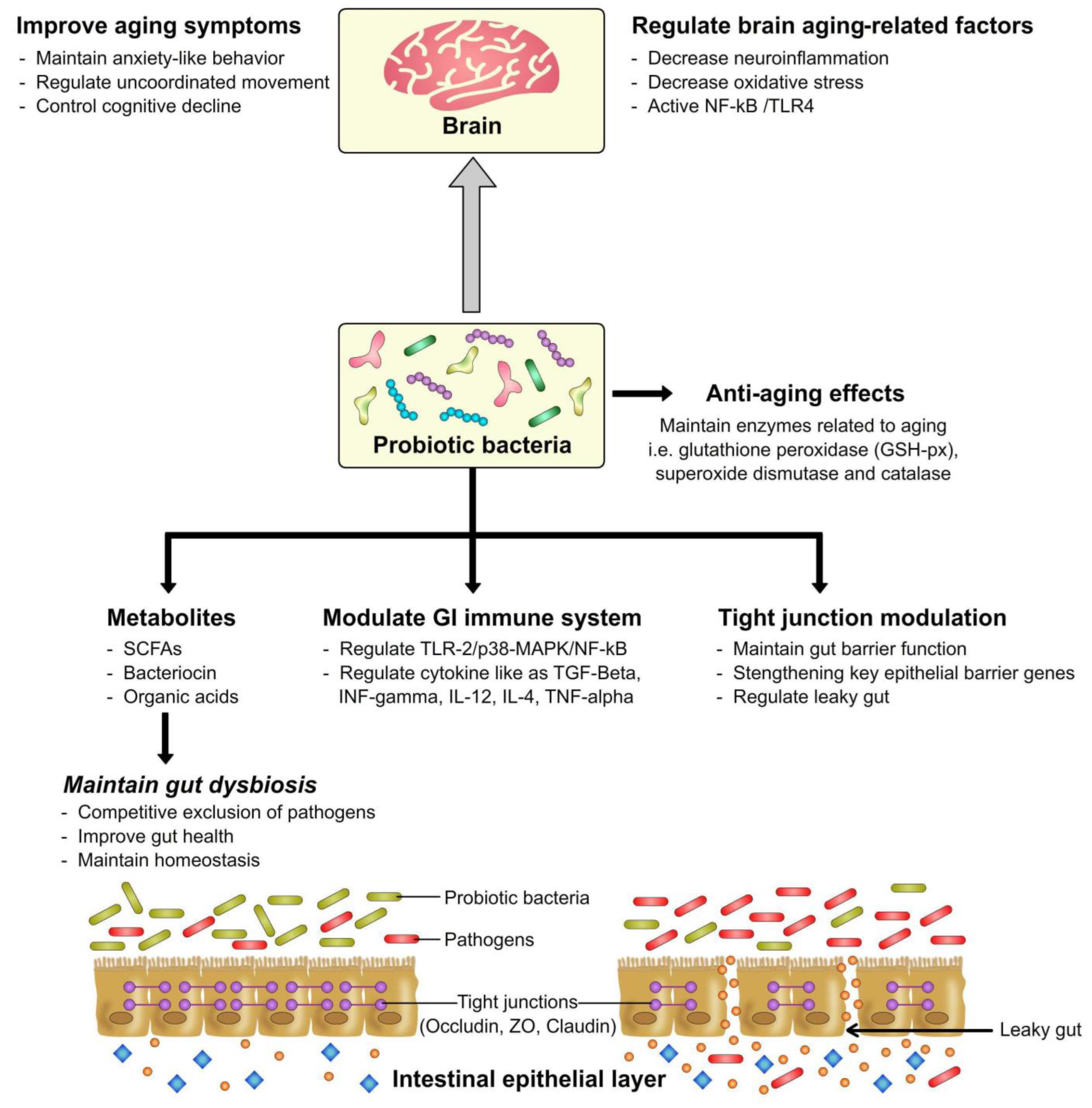
Hippocrates already realized that “all diseases begin in the gut”. These days, researchers working on the human microbiota have concluded that in order to temporarily improve the microbes in the gut, fermented foods need to be consumed as part of a healthy diet [36]. Nearly a hundred trillion microbes reside in the human gut; that includes nearly 1000 known bacterial species. In addition to an array of microorganisms, the gut also contains about 70% of the body’s immune system via gut-associated lymphoid tissue. Therefore, a major portion of the immune system resides in the intestinal tract and aids in maintaining the balance of gut microbiota [10]. Probiotics have been reported to have prophylactic properties to improve the diverse gut-linked concerns or diseases via the modulation of general gut microbiota functions, such as immune-system processes and metabolic pathways [27]. Figure 2 shows these promising attributes of probiotics in relation to the regulation of aging and gut health modulation.
The immune system also controls intestinal inflammation and allergies using SCFAs that are produced via the microbial breakdown of dietary constituents such as fiber [51]. SCFAs are the key metabolic products of microbes formed during the fermentation of indigestible and partially digestible polysaccharides in the gut. These SCFAs can regulate the immune functions and homeostasis in the gut via antimicrobial, anti-inflammatory, and antitumorigenic activities. Probiotics target SCFA production by modulating the gut microbiota composition in the gastrointestinal tract, which may alter the metabolic profile and, thus, the metabolome [17,52]. A recent study by Kong et al. [53] confirmed that probiotic supplementation improves the gut microbiota diversity in high-fat-diet (HFD) and high-sucrose-diet (HCD) mice models. Results have found that probiotic supplementation increased the abundance of microbiota negatively associated with obesity, including Akkermansia, Lactococcus, and Bifidobacterium. In addition, Escherichia, Shigella, Blautia, Acinetobacter, and Oscillibacter significantly increased in the HCD group; Ruminococcus, Olsenella, and Allobaculum were significantly changed in the HFD group. Another study confirmed that probiotic Lactobacillus rhamnosus GG (LGG) improved the gut microbiota diversity using Chao1, ACE, Simpson, and Shannon indices (Alpha diversity indices). Chao1 and ACE indices are mainly used to evaluate gut microbiota richness, whereas Simpson and Shannon indices could measure gut microbiota diversity. Results have found that LGG promotes alpha diversity in the gut microbiota. Furthermore, results concluded that low doses of the LGG group increase the abundances of Bacteroidetes and Firmicutes, while leading to a decrease in the Verrucomicrobia, Proteobacteria, and Deferribacteres abundances [54]. The gut epithelial barrier integrity, integral to overall host health, is controlled by the tight junction proteins that form complexes among the intestinal epithelial cells. Deviations in the tight junction proteins disturb gut mucosal barrier integrity, consequently permitting the undesirable movement of infectious microbes within and outside of the gut via the intestinal epithelium. Probiotics directly improve gut barrier functions via the inhibition of pathogens by increasing the tight junctions’ protein expression or reorganizing the tight junction proteins [3,55]. Studies on in vitro cell lines and in vivo animal models have established the potential effects of probiotics for improving the tight junction proteins to modulate the gastrointestinal tract’s health. Wang et al. [29] reported that lipoteichoic acid, a cell-wall component of human-originated L. paracasei D3-5, has the potential to reduce age-related leaky gut and inflammation, as well as to enhance the Muc2 expression by modifying the TLR-2/p38-MAPK/NF-kB-signaling pathway in mice models. Samtiya et al. [23] reported that L. fermentum MTCC-5898 could improve gut integrity. When L. fermentum was administered orally to mice for 28 days, it improved the mRNA gene expression of Claudin-1 and MUC-2, demonstrating the development of gut barrier functions. A clinical trial of 20 healthy Italian volunteers provided evidence that B. longum BB536 and L. rhamnosus HN001 modulated the gut microbiota composition when preprandially and postprandially administrated for 1 month [56]. The supplementation of probiotic lactobacilli corrected preexisting gut dysbiosis, regulated amino acid (i.e., 5-oxoproline, urocanic acid, cystamine, and citrulline) metabolism, and improved antioxidative possibilities in the aged rats (i.e., d-galactose senescence-induced), highlighting potential anti-aging effects of probiotics [57]. Marteau et al. [58] indicated that a fermented milk product comprising Bifidobacterium animalis subsp. lactis CNCM I-2494 and lactic acid bacteria have improved digestive symptoms in women after four weeks of ingestion. Human-originated cocktail of probiotics (i.e., 5 strains of each Lactobacillus and Enterococcus) has shown therapeutic potential in mice. This probiotic cocktail prevented high-fat-diet-induced leaky gut, gut dysbiosis, and inflammation, together with other metabolic irregularities and weakened the physical functions of the aged mice after ten weeks of feeding [59]. Similarly, a probiotic mixture isolated from human origin could control intestinal health and enhance the production of the native SCFAs. A single oral dose, as well as an additional five consecutive dosages of this probiotic mixture (10 strains), in mice controls the gut microbiome and increases the production of SCFAs. Therefore, the human-originated probiotic lactobacilli and enterococci may improve gut dysbiosis and could be utilized as a possible treatment for dysbiosis-related diseases [60].
Bifidobacterium has been reported as one of the most abundant strains in breast milk and has immense neonatal health benefits [61,62]. Various Bifidobacterium bifidis strains have also been reported for their support in helping the host overcome irritable bowel syndrome and allergic reactions [63]. A recent study has confirmed the potential of Lactobacillus helveticus-containing citrulline-enriched fermented milk (LHFM_Cit) to improve the intestinal epithelial barrier function and inflammatory response caused by pathogenic E. coli on IPEC-J2 cells. Results have found that treatment of IPEC-J2 cells with 5% (v/v) of fermented milk with live LH511 and 4 mM citrulline (5%LHFM_Cit-4mM) improved the transepithelial electrical resistance (TEER), regulated the tight junction proteins (zonula occluden-1, occludin, and claudin-1), and amended the toll-like receptors (TLRs), as well as negative controllers of the TLR signaling pathway (A20 and IRAK-M) [64]. Yu et al. [65] assessed the capability of probiotics to modulate gut health in pre-weaned mouse pups. Maternal Lactobacillus acidophilus and Bifidobacterium infantis exposure facilitated the intestinal epithelial cell differentiation; prohibited the loss of mucin; conserved the intestinal integrity and barrier functions; and reduced the levels of serum interleukin-1β, tumor necrosis factor α, and interleukin-6 in the pre-weaned offspring. The potential of probiotics to modulate gut morphology and intestinal absorption strength has been reported. L. fermentum IKP 23, L. fermentum IKP 111, and L. salivarius IKP 333 can regulate histomorphology of the small intestine and improve the D-xylose absorption ability in broiler chicken challenged with Salmonella enteritidis [55]. Table 1 shows the factors related to the use of probiotics as a gut health modulator.
4. Probiotics as Anti-Aging Agents in Relation to the Gut-Brain Axis
Aging is a multifaceted and progressively distorted process that comes with the key risk of severe chronic conditions and disorders in elderly persons. Efforts have been made to accurately understand the human aging process, and diverse theories have been established which consider both the genetic and epigenetic aspects that may play a role [66]. In relation to the gut, as mentioned in the above sections, aging is associated with gut dysbiosis [10,67], or the loss of the number and diversity of the gut microbiota, leading to multiple age-related disorders. Recent attention has been given to the interaction between the gut microbiome and the brain in the context of neurodegenerative conditions such as Alzheimer’s disease, wherein neuroinflammation and cognitive decline play an important role. This concept is termed as the “gut–brain axis” and is defined as the cross talk between the central nervous system, immune system, and endocrine system, with cognitive centers of the brain and peripheral intestinal cellular function [68,69,70]. Bidirectional communication can occur through multiple channels, including tryptophan metabolism, and acquires particular importance in the context of age-related diseases [71,72]. Thus, use of novel cellular and microbiome-altering strategies is needed to investigate therapeutics designed to preserve cognitive function in the expanding older adult population.
Overall, reactive oxygen species accretion has been reported as a vital moderator of the diverse phases of age-linked cellular deterioration due to their capacity to interrupt inflammatory and cellular oxidative balance. In addition, reactive oxygen species speed up the gradual destruction of the telomere, which is thought to have a role in the process of aging [7]. As a potential aging modulator, probiotics have the potential to regulate age-linked injuries and pathologies. This is particularly confirmed in studies regarding gastrointestinal health and immunosenescence [56,73,74,75]. Molecular mechanisms involved in the aging progression are not well understood, though oxidative stress and nervous inflammation are both considered typical features of the aging brain [76,77]. Stimulation of the toll-like receptor 4/nuclear-factor-κB plays a role in the production of tumor necrosis factor-α, interleukin-6, and interleukin-1β, finally leading to cognitive dysfunction and neuroinflammation [78]. A reducing sugar generally found in the body (i.e., D-galactose) causes different age-related disorders, including cognitive decline, enhanced malondialdehyde activity, and the reduction of antioxidant enzyme functions such as superoxide dismutase (an antioxidant enzyme) and glutathione peroxidase [79,80]. To this end, it is clear that the gut and its microbes have a role to play in brain health. Psychobiotic bacteria are defined as “live organisms that, when ingested in adequate amounts, produce a brain health benefits” [81]. Although, all interventions targeting microbiota to regulate the microbiota-brain-related functioning are currently categorized as “psychobiotics”, most were limited to gut health [82].
Wang et al. [29] reported that a human-originated, heat-killed Lactobacillus paracasei D3-5 improved their physical and cognitive function when fed to old mice. Bifidobacterium longum and B. animalis show the potential to ameliorate aging, anxiety-like behavior, cognitive deterioration, and unconscious movement when orally supplemented to D-galactose-treated mice. B. longum and B. animalis, both alone and in combination, improved the age-linked cognitive functions by NF-κB/TLR4-prompted neuroinflammation inhibition. These strains also ameliorated oxidative stress by upregulating Sirt 1 protein expression [11]. Sharma et al. [83] documented that probiotic interventions can benefit aged people via health improvements with the reduction of inflammatory illnesses in aging mice models. Supplementation of Lactobacillus fermentum MBC2 to the model species, C. elegans, improved general longevity and weakened the presence of age-linked indicators such as lipofuscin accumulation, nematode body bending, and pumping rate [84].
A study conducted by Yu et al. [85] assessed the anti-aging potential of three strains of Lactobacillus (i.e., L. mucosae LMU1001, and L. plantarum LPL0902 and LPL0302) that were isolated from the gastrointestinal tract of Gaotian villagers in China (92 years old). Results concluded that L. mucosae LMU1001-containing yogurt had the most effective attributes out of the studied strains to improve the antioxidant activity, thus potentially diminishing certain aging effects. Additionally, a significant increase was confirmed in glutathione peroxidase activities in serum and liver superoxide dismutase in the d-galactose-induced aging mice. Lin et al. [86] established the potential of L. plantarum AR113 and AR501 to promote anti-aging activity. The oral supplementation of probiotics amended the antioxidant status of D-galactose-induced oxidative stress mice and regulated abnormal activities to the homeostatic baseline levels of catalase, glutathione peroxidase, and superoxide dismutase. Oral supplementation also increased the nuclear factor erythroid-2 related factor 2 gene expression. Similarly, another study reported that oral supplementation of Lactobacillus brevis OW38 improved age-related colitis and memory losses in aging mice models via p16 expression, intestinal microbiota lipopolysaccharide production, and NF- ĸB activation [5]. Likewise, Lactobacillus paracasei PS23 administration slowed down the age-linked cognitive decay in senescence-accelerated mouse prone 8 models that were defined by an early commencement of age-linked modifications [87]. Hor et al. [88] have also reported the potential of Lactobacillus strains in improving the aging of the musculoskeletal system in D-galactose-induced aging rats. L. fermentum DR9 increased the expression of AMPK-α2, while reducing the expression of IL-6 and tartrate-resistant acid phosphatase in the tibia. Consequently, L. fermentum DR9 may be used as a promising anti-aging agent. These findings suggest that probiotic therapies of human-originated cocktails could be beneficial for inhibiting age-linked metabolic dysfunctions, physical deterioration, and inflammation, while promoting healthy aging [59]. Table 2 presents studies related to probiotics as an anti-aging agent in relation to aging-brain health.
5. Prospects and Developments
The growth of the aging population has subsequently led to a rising number of different age-linked conditions, such as neurodegenerative issues and dementia. Accordingly, due to the lack of effective therapy to restrain and/or regulate the mechanisms related to age-linked cognitive degeneration, it is essential to shift our research efforts towards the prevention or development of lifestyle programs that aid in maintaining health for long life spans [91]. The gut flora is colonized by a wide variety of microbes that can be either pathogenic or have a beneficial relationship with the host [92]. Probiotics and their corresponding prebiotics are beneficial in preventing and treating many diseases, including aging and gastrointestinal problems [66,93]. Increasing awareness of these benefits has led to the increased commercialization of pro- and prebiotic-based supplements and foods. Many foods, such as yogurt, cheese, kefir, and kombucha, naturally contain probiotics, and an increasing number of commercial probiotics are available in the form of supplements, drinks, and fermented milk products. In order to be effective, probiotics need to be consumed on a regular basis [94]. Presently, the international market of functional foods, as well as probiotics, is growing at a rapid pace. On account of the large response of consumers, the market has moved towards the increased production of functional dairy or nondairy products containing probiotics. These probiotic-containing foods or supplements are vastly produced in three different forms: for direct or indirect human ingestion (freeze-dried form), for industrial or home usages (fermented or nonfermented formula), or in powder/capsule/tablets form [95]. The general public has more recently been made aware of the functional correlation between diet and health, and now they need only those products that provide those suitable nutrients and defined constituents that claim to improve life quality, longevity, and/or health [36]. The experts’ panel of the European Society for Primary Care Gastroenterology stated that specifically tested probiotics should be considered for use in the regulation of different irritable bowel syndrome symptoms [96,97].
The microbiota-targeted treatments can be used as a possible therapeutic for specific age-linked disorders and can potentially play a role in regulating the health concerns that accompany the aging process [67]. The probiotics-based dietary therapies, nutraceuticals, and fiber constituent’s additions to the gut microbiota are increasingly being studied, especially regarding their potential ability to improve the gut imbalance for improved human health. A study also showed the protective effects and fundamental mechanisms of particular probiotics that may be used to develop potential remedies for a better aging life [14,60]. Several technologies, including next-generation sequencing, have been developed, making it possible to identify and evaluate intestinally culturable and nonculturable bacteria. There have been different studies on the relationship between illnesses and alterations in the intestinal microbiome. These reports indicated that the intestinal microbiota plays a significant role between human health and disorders [98]. The probiotics could modulate the intestinal microbiome and improve gut dysbiosis.
Consequently, probiotics can be one of the key interventions for regulating the healthy gut microbiome and improving an imbalanced gut and inflammation. However, it remains primarily unknown via which mechanism the probiotics moderate the gut and inflammation, particularly in aging people [59]. The probiotics used as or in dietary complements and diets include bacteria or yeast and are available in the market as tablets, freeze-dried powder, capsules, and fermented foods such as yogurt. Probiotic products contain a single strain or a more diverse cocktail of different strains [99]. Cerium dioxide nanoparticles, which are well-known for UV shielding, neurotropic, and anti-aging characteristics, have the potential for various biomedical applications. The practices of probiotics with nanoparticle-based management and food complements seem to be promising, with many potential avenues for future research [100].
6. Conclusions
The human gut microbiome is a multifaceted community of microbes that synergistically cooperate with the host by generating SCFAs, influencing intestinal inflammation and regulating immune function. Aging is a universal process that has globally confronted researchers, all seeking to understand its mechanisms and, subsequently, how to slow down the process through corresponding pathways. Aging is a stochastic, damaging, and progressive event that significantly contributes to decreased quality of life and increased strain on the medical system. Given that the elderly population is set to increase, it is all the more important to understand this general process better. Aging itself is not an ailment, but the related declines are, and efforts have been made to recognize its source, causes, and genetic role. The role of the microbiome also cannot be ignored, as this consortia of microorganisms in the gut have been closely linked to both the onset and the prevention of many pathologies. By increasing the composition of beneficial microbes and reducing the progression of pathogens and native pathobionts, probiotics could modulate the microbial composition in such a way as to maintain homeostasis in the intestinal tract. Probiotics have health-promoting attributes and can be part of an effective strategy to enhance longevity and health by regulating different metabolic pathways and disorders. As of now, there are only a few studies that highlight probiotics as being promising therapeutic agents for cognitive and brain-related disorders. Nonetheless, these few studies begin to support the effectiveness of probiotics as a therapeutic approach for improving aging and gut health modulation. Probiotics could be used as one of the multiple effective interventions for the regulation of anti-aging and gut-related ailments in humans. However, further in vitro and in vivo studies are needed before postulating or conducting any clinical or human trials to validate these claims.
Author Contributions
Conceptualization: M.S., A.K.P. and T.D.; writing—original draft preparation: M.S.; writing—review and editing, M.S., A.K.P., M.P., N.P.S., T.D. and R.V. All authors have read and agreed to the published version of the manuscript.
Funding
The author (M.S.) is thankful to DST-Haryana (HSCSIT/R&D/2021/461), India, for the award of Junior Research Fellowship (Financial support).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
References
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fijan, S. Microorganisms with claimed probiotic properties: An overview of recent literature. Int. J. Environ. Res. Public Health 2014, 11, 4745–4767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.; Moore, R.J.; Stanley, D.; Chousalkar, K.K. The gut microbiota of laying hens and its manipulation with prebiotics and probiotics to enhance gut health and food safety. Appl. Environ. Microbiol. 2020, 86, e00600-20. [Google Scholar] [CrossRef] [PubMed]
- Abdou, A.M.; Hedia, R.H.; Omara, S.T.; Mahmoud, M.A.; Kandil, M.M.; Bakry, M.A. Interspecies comparison of probiotics isolated from different animals. Vet. World 2018, 11, 227. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.J.; Kim, K.A.; Hwang, Y.J.; Han, M.J.; Kim, D.H. Anti-inflammaging effects of Lactobacillus brevis OW38 in aged mice. Benef. Microbes 2016, 7, 707–718. [Google Scholar] [CrossRef]
- Kawase, M.; He, F.; Miyazawa, K.; Kubota, A.; Yoda, K.; Hiramatsu, M. Orally administered heat-killed Lactobacillus gasseri TMC0356 can upregulate cell-mediated immunity in senescence-accelerated mice. FEMS Microbiol. Lett. 2012, 326, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Padwad, Y. Probiotic bacteria as modulators of cellular senescence: Emerging concepts and opportunities. Gut Microbes 2020, 11, 335–349. [Google Scholar] [CrossRef]
- Arai, Y.; Martin-Ruiz, C.M.; Takayama, M.; Abe, Y.; Takebayashi, T.; Koyasu, S.; Suematsu, M.; Hirose, N.; von Zglinicki, T. Inflammation, but not telomere length, predicts successful ageing at extreme old age: A longitudinal study of semi-supercentenarians. EBioMedicine 2015, 2, 1549–1558. [Google Scholar] [CrossRef] [Green Version]
- Lumeng, C.N.; Liu, J.; Geletka, L.; Delaney, C.; Delproposto, J.; Desai, A.; Oatmen, K.; Martinez-Santibanez, G.; Julius, A.; Garg, S.; et al. Aging is associated with an increase in T cells and inflammatory macrophages in visceral adipose tissue. J. Immunol. 2011, 187, 6208–6216. [Google Scholar] [CrossRef] [Green Version]
- Buford, T.W. (Dis) Trust your gut: The gut microbiome in age-related inflammation, health, and disease. Microbiome 2017, 5, 80. [Google Scholar] [CrossRef]
- Xia, C.; Cao, X.; Cui, L.; Liu, H.; Wang, S.; Chen, T. Anti-aging effect of the combination of Bifidobacterium longum and B. animalis in a d-galactose-treated mice. J. Funct. Foods 2020, 69, 103938. [Google Scholar] [CrossRef]
- Landete, J.M.; Gaya, P.; Rodríguez, E.; Langa, S.; Peirotén, Á.; Medina, M.; Arqués, J.L. Probiotic bacteria for healthier aging: Immunomodulation and metabolism of phytoestrogens. BioMed Res. Int. 2017, 2017, 5939818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Aging and Health. 2018. Available online: https://www.who.int/news-room/fact-sheets/detail/ageing-and-health (accessed on 12 September 2020).
- Nagpal, R.; Mainali, R.; Ahmadi, S.; Wang, S.; Singh, R.; Kavanagh, K.; Kitzman, D.W.; Kushugulova, A.; Marotta, F.; Yadav, H. Gut microbiome and aging: Physiological and mechanistic insights. Nutr. Healthy Aging 2018, 4, 267–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunningham, M.; Azcarate-Peril, M.A.; Barnard, A.; Benoit, V.; Grimaldi, R.; Guyonnet, D.; Holscher, H.D.; Hunter, K.; Manurung, S.; Obis, D.; et al. Shaping the future of probiotics and prebiotics. Trends Microbiol. 2021, 29, 667–685. [Google Scholar] [CrossRef] [PubMed]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-inflammatory and immunomodulatory effects of probiotics in gut inflammation: A door to the body. Front. Immunol. 2021, 12, 178. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Matar, C.; Ismail, N. Adolescence and aging: Impact of adolescence inflammatory stress and microbiota alterations on brain development, aging, and neurodegeneration. J. Gerontol. Ser. A 2020, 75, 1251–1257. [Google Scholar] [CrossRef] [Green Version]
- Kelly, J.R.; Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G.; Hyland, N.P. Breaking down the barriers: The gut microbiome, intestinal permeability and stress-related psychiatric disorders. Front. Cell. Neurosci. 2015, 9, 392. [Google Scholar] [CrossRef] [Green Version]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of action of probiotics. Adv. Nutr. 2019, 10 (Suppl. 1), S49–S66. [Google Scholar] [CrossRef] [Green Version]
- Szajewska, H. What are the indications for using probiotics in children? Arch. Dis. Child 2016, 101, 398–403. [Google Scholar] [CrossRef] [Green Version]
- Stewart, L.; Crumley, B.; Walton, K. Effects of probiotic VSL# 3 on cytokine and tight junction protein expression in intestinal epithelial cells. FASEB J. 2015, 29, 1010–1014. [Google Scholar] [CrossRef]
- Yousefi, B.; Eslami, M.; Ghasemian, A.; Kokhaei, P.; Salek Farrokhi, A.; Darabi, N. Probiotics importance and their immunomodulatory properties. J. Cell Physiol. 2019, 234, 8008–8018. [Google Scholar] [CrossRef] [PubMed]
- Samtiya, M.; Bhat, M.I.; Gupta, T.; Kapila, S.; Kapila, R. Safety assessment of potential probiotic Lactobacillus fermentum MTCC-5898 in murine model after repetitive dose for 28 days (Sub-Acute Exposure). Probiot. Antimicrob. Proteins 2020, 12, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Díaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Immune-mediated mechanisms of action of probiotics and synbiotics in treating pediatric intestinal diseases. Nutrients 2018, 10, 42. [Google Scholar] [CrossRef] [Green Version]
- Wan, L.Y.M.; Chen, Z.J.; Shah, N.P.; El-Nezami, H. Modulation of intestinal epithelial defense responses by probiotic bacteria. Crit. Rev. Food. Sci. Nutr. 2016, 56, 2628–2641. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, D.K.; Puniya, M.; Shandilya, U.K.; Dhewa, T.; Kumar, N.; Kumar, S.; Puniya, A.K.; Shukla, P. Gut microbiota modulation and its relationship with obesity using prebiotic fibers and probiotics: A review. Front. Microbiol. 2017, 8, 563. [Google Scholar] [CrossRef]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef]
- Szabóová, R. Optimal Criteria for the Selection of Probiotics, Based on their Mode of Action. Folia Vet. 2019, 63, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Ahmadi, S.; Nagpal, R.; Jain, S.; Mishra, S.P.; Kavanagh, K.; Zhu, X.; Wang, Z.; McClain, D.A.; Kritchevsky, S.B.; et al. Lipoteichoic acid from the cell wall of a heat killed Lactobacillus paracasei D3-5 ameliorates aging-related leaky gut, inflammation and improves physical and cognitive functions: From C. elegans to mice. Geroscience 2020, 42, 333–352. [Google Scholar] [CrossRef]
- Wu, Q.; Cheung, C.K.; Shah, N.P. Towards galactose accumulation in dairy foods fermented by conventional starter cultures: Challenges and strategies. Trends Food Sci. Technol. 2015, 41, 24–36. [Google Scholar] [CrossRef]
- Bron, P.A.; Van Baarlen, P.; Kleerebezem, M. Emerging molecular insights into the interaction between probiotics and the host intestinal mucosa. Nat. Rev. Microbiol. 2012, 10, 66–78. [Google Scholar] [CrossRef]
- Nothaft, H.; Perez-Muñoz, M.E.; Gouveia, G.J.; Duar, R.M.; Wanford, J.J.; Lango-Scholey, L.; Panagos, C.G.; Srithayakumar, V.; Plastow, G.S.; Coros, C.; et al. Coadministration of the Campylobacter jejuni N-glycan-based vaccine with probiotics improves vaccine performance in broiler chickens. Appl. Environ. Microbiol. 2017, 83, e01523-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engevik, M.A.; Luk, B.; Chang-Graham, A.L.; Hall, A.; Herrmann, B.; Ruan, W.; Endres, B.T.; Shi, Z.; Garey, K.W.; Hyser, J.M.; et al. Bifidobacterium dentium fortifies the intestinal mucus layer via autophagy and calcium signaling pathways. mBio 2019, 10, e01087-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Shah, N.P. High γ-aminobutyric acid production from lactic acid bacteria: Emphasis on Lactobacillus brevis as a functional dairy starter. Crit. Rev. Food Sci. Nutr. 2017, 57, 3661–3672. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, A.; Lam, L.; Rajendram, M.; Tamburini, F.; Honeycutt, J.; Pham, T.; Van Treuren, W.; Pruss, K.; Stabler, S.R.; Lugo, K.; et al. A gut commensal-produced metabolite mediates colonization resistance to Salmonella infection. Cell Host Microbe 2018, 24, 296–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valero-Cases, E.; Cerdá-Bernad, D.; Pastor, J.J.; Frutos, M.J. Non-dairy fermented beverages as potential carriers to ensure probiotics, prebiotics, and bioactive compounds arrival to the gut and their health benefits. Nutrients 2020, 12, 1666. [Google Scholar] [CrossRef]
- Huang, R.; Tao, X.; Wan, C.; Li, S.; Xu, H.; Xu, F.; Shah, N.P.; Wei, H. In vitro probiotic characteristics of Lactobacillus plantarum ZDY 2013 and its modulatory effect on gut microbiota of mice. J. Dairy Sci. 2015, 98, 5850–5861. [Google Scholar] [CrossRef]
- Zhang, W.; Zhu, Y.H.; Yang, J.C.; Yang, G.Y.; Zhou, D.; Wang, J.F. A selected Lactobacillus rhamnosus strain promotes EGFR-independent Akt activation in an enterotoxigenic Escherichia coli K88-infected IPEC-J2 cell model. PLoS ONE 2015, 10, e0125717. [Google Scholar] [CrossRef]
- Mujagic, Z.; De Vos, P.; Boekschoten, M.V.; Govers, C.; Pieters, H.J.H.; De Wit, N.J.; Bron, P.A.; Masclee, A.A.; Troost, F.J. The effects of Lactobacillus plantarum on small intestinal barrier function and mucosal gene transcription; a randomized double-blind placebo controlled trial. Sci. Rep. 2017, 7, 40128. [Google Scholar] [CrossRef]
- Cӑtoi, A.F.; Corina, A.; Katsiki, N.; Vodnar, D.C.; Andreicuț, A.D.; Stoian, A.P.; Rizzo, M.; Pérez-Martínez, P. Gut microbiota and aging-A focus on centenarians. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165765. [Google Scholar] [CrossRef]
- Metchnikoff, E. Optimistic Studies; Putman’s Sons: New York, NY, USA, 1908; pp. 161–183. [Google Scholar]
- Ayala, F.R.; Bauman, C.; Cogliati, S.; Leñini, C.; Bartolini, M.; Grau, R. Microbial flora, probiotics, Bacillus subtilis and the search for a long and healthy human longevity. Microb. Cell 2017, 4, 133. [Google Scholar] [CrossRef]
- Patel, P.J.; Singh, S.K.; Panaich, S.; Cardozo, L. The aging gut and the role of prebiotics, probiotics, and synbiotics: A review. J. Clin. Gerontol. Geriatr. 2014, 5, 3–6. [Google Scholar] [CrossRef] [Green Version]
- Lew, L.; Hor, Y.; Jaafar, M.; Lau, A.; Ong, J.; Chuah, L.; Yap, K.; Azzam, G.; Azlan, A.; Liong, M. Lactobacilli modulated AMPK activity and prevented telomere shortening in ageing rats. Benef. Microbes 2019, 10, 883–892. [Google Scholar] [CrossRef] [PubMed]
- Nam, B.; Kim, S.A.; Nam, W.; Jeung, W.H.; Park, S.-D.; Lee, J.-L.; Sim, J.-H.; Jang, S.S. Lactobacillus plantarum HY7714 restores TNF-α induced defects on tight junctions. Prev. Nutr. Food Sci. 2019, 24, 64. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Joishy, T.; Das, S.; Kalita, M.C.; Mukherjee, A.K.; Khan, M.R. A potential probiotic Lactobacillus plantarum JBC5 improves longevity and healthy aging by modulating antioxidative, innate immunity and serotonin-signaling pathways in Caenorhabditis elegans. Antioxidants 2022, 11, 268. [Google Scholar] [CrossRef]
- Sivamaruthi, B.S.; Fern, L.A.; Hj, D.S.N.R.P.; Chaiyasut, C. The influence of probiotics on bile acids in diseases and aging. Biomed. Pharmacother. 2020, 128, 110310. [Google Scholar] [CrossRef]
- Calder, P.C.; Bosco, N.; Bourdet-Sicard, R.; Capuron, L.; Delzenne, N.; Doré, J.; Franceschi, C.; Lehtinen, M.J.; Recker, T.; Salvioli, S.; et al. Health relevance of the modification of low grade inflammation in ageing (inflammageing) and the role of nutrition. Ageing Res. Rev. 2017, 40, 95–119. [Google Scholar] [CrossRef]
- Fong, F.L.Y.; Shah, N.P.; Kirjavainen, P.; El-Nezami, H. Mechanism of action of probiotic bacteria on intestinal and systemic immunities and antigen-presenting cells. Int. Rev. Immunol. 2016, 35, 179–188. [Google Scholar] [CrossRef]
- Yang, H.; Sun, Y.; Cai, R.; Chen, Y.; Gu, B. The impact of dietary fiber and probiotics in infectious diseases. Microb. Pathog. 2020, 140, 103931. [Google Scholar] [CrossRef]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef] [Green Version]
- Westfall, S.; Lomis, N.; Kahouli, I.; Dia, S.Y.; Singh, S.P.; Prakash, S. Microbiome, probiotics and neurodegenerative diseases: Deciphering the gut brain axis. Cell Mol. Life Sci. 2017, 74, 3769–3787. [Google Scholar] [CrossRef]
- Kong, C.; Gao, R.; Yan, X.; Huang, L.; Qin, H. Probiotics improve gut microbiota dysbiosis in obese mice fed a high-fat or high-sucrose diet. Nutrition 2019, 60, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.W.; Cheng, M.Y.; Yang, X.; Lu, Y.Y.; Yin, H.D.; Zeng, Y.; Wang, R.Y.; Jiang, Y.L.; Yang, W.T.; Wang, J.Z.; et al. Probiotic Lactobacillus rhamnosus GG Promotes Mouse Gut Microbiota Diversity and T Cell Differentiation. Front. Microbiol. 2020, 11, 3216. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Nawaz, M.; Anjum, A.A.; Ahmad, M.U.D.; Mehmood, A.; Rabbani, M.; Mustafa, A.; Ali, M.A. Effect of Indigenous Probiotics on Gut Morphology and Intestinal Absorption Capacity in Broiler Chicken Challenged with Salmonella enteritidis. Pak. J. Zool. 2020, 52, 1825. [Google Scholar] [CrossRef]
- Toscano, M.; De Grandi, R.; Stronati, L.; De Vecchi, E.; Drago, L. Effect of Lactobacillus rhamnosus HN001 and Bifidobacterium longum BB536 on the healthy gut microbiota composition at phyla and species level: A preliminary study. World J. Gastroenterol. 2017, 23, 2696. [Google Scholar] [CrossRef] [PubMed]
- Hor, Y.Y.; Lew, L.C.; Jaafar, M.H.; Lau, A.S.Y.; Ong, J.S.; Kato, T.; Nakanishi, Y.; Azzam, G.; Azlan, A.; Ohno, H.; et al. Lactobacillus sp. improved microbiota and metabolite profiles of aging rats. Pharmacol. Res. 2019, 146, 104312. [Google Scholar] [CrossRef] [PubMed]
- Marteau, P.; Le Nevé, B.; Quinquis, L.; Pichon, C.; Whorwell, P.J.; Guyonnet, D. Consumption of a fermented milk product containing Bifidobacterium lactis CNCM I-2494 in women complaining of minor digestive symptoms: Rapid response which is independent of dietary fibre intake or physical activity. Nutrients 2019, 11, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadi, S.; Wang, S.; Nagpal, R.; Wang, B.; Jain, S.; Razazan, A.; Mishra, S.P.; Zhu, X.; Wang, Z.; Kavanagh, K.; et al. A human-origin probiotic cocktail ameliorates aging-related leaky gut and inflammation via modulating the microbiota/taurine/tight junction axis. JCI Insight 2020, 5, e132055. [Google Scholar] [CrossRef]
- Nagpal, R.; Wang, S.; Ahmadi, S.; Hayes, J.; Gagliano, J.; Subashchandrabose, S.; Kitzman, D.W.; Becton, T.; Read, R.; Yadav, H. Human-origin probiotic cocktail increases short-chain fatty acid production via modulation of mice and human gut microbiome. Sci. Rep. 2018, 8, 12649. [Google Scholar] [CrossRef] [Green Version]
- Arboleya, S.; Ruas-Madiedo, P.; Margolles, A.; Solís, G.; Salminen, S.; Clara, G.; Gueimonde, M. Characterization and in vitro properties of potentially probiotic Bifidobacterium strains isolated from breast-milk. Int. J. Food Microbiol. 2011, 149, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Miles, M.P. Probiotics and Gut Health in Athletes. Curr. Nutr. Rep. 2020, 9, 129–136. [Google Scholar] [CrossRef]
- Ku, S.; Park, M.S.; Ji, G.E.; You, H.J. Review on Bifidobacterium bifidum BGN4: Functionality and nutraceutical applications as a probiotic microorganism. Int. J. Mol. Sci. 2016, 17, 1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, S.W.; El-Nezami, H.; Shah, N.P. The protective effects of enriched citrulline fermented milk with Lactobacillus helveticus on the intestinal epithelium integrity against Escherichia coli infection. Sci. Rep. 2020, 10, 499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Lu, J.; Oliphant, K.; Gupta, N.; Claud, K.; Lu, L. Maternal administration of probiotics promotes gut development in mouse offsprings. PLoS ONE 2020, 15, e0237182. [Google Scholar] [CrossRef]
- Sharma, R.; Padwad, Y. Perspectives of the potential implications of polyphenols in influencing the interrelationship between oxi-inflammatory stress, cellular senescence and immunosenescence during aging. Trends Food Sci. Technol. 2020, 98, 41–52. [Google Scholar] [CrossRef]
- Vaiserman, A.M.; Koliada, A.K.; Marotta, F. Gut microbiota: A player in aging and a target for anti-aging intervention. Ageing Res. Rev. 2017, 35, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.A.; McVey Neufeld, K.A. Gut-brain axis: How the microbiome influences anxiety and depression. Trends Neurosci. 2013, 36, 305–312. [Google Scholar] [CrossRef]
- El Aidy, S.; Dinan, T.G.; Cryan, J.F. Gut Microbiota: The Conductor in the Orchestra of Immune-Neuroendocrine Communication. Clin. Ther. 2015, 37, 954–967. [Google Scholar] [CrossRef]
- Sun, Y.; Baptista, L.C.; Roberts, L.M.; Jumbo-Lucioni, P.; McMahon, L.L.; Buford, T.W.; Carter, C.S. The gut microbiome as a therapeutic target for cognitive impairment. J. Gerontol. A 2020, 75, 1242–1250. [Google Scholar] [CrossRef]
- Forsythe, P.; Bienenstock, J.; Kunze, W.A. Vagal pathways for microbiome-brain-gut axis communication. In Microbial Endocrinology: The Microbiota-Gut-Brain Axis in Health and Disease; Lyte, M., Cryan, J., Eds.; Springer: New York, NY, USA, 2014; pp. 115–133. [Google Scholar] [CrossRef]
- Calvani, R.; Picca, A.; Lo Monaco, M.R.; Landi, F.; Bernabei, R.; Marzetti, E. Of microbes and minds: A narrative review on the second brain aging. Front. Med. 2018, 5, 53. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Kapila, R.; Kapasiya, M.; Saliganti, V.; Dass, G.; Kapila, S. Dietary supplementation of milk fermented with probiotic Lactobacillus fermentum enhances systemic immune response and antioxidant capacity in aging mice. Nutr. Res. 2014, 34, 968–981. [Google Scholar] [CrossRef]
- Malaguarnera, G.; Leggio, F.; Vacante, M.; Motta, M.; Giordano, M.; Biondi, A.; Basile, F.; Mastrojeni, S.; Mistretta, A.; Malaguarnera, M.; et al. Probiotics in the gastrointestinal diseases of the elderly. J. Nutr. Health Aging 2012, 16, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.; Kalam, A.; Sarker, M.; Li, T.; Yin, J. Probiotic species in the modulation of gut microbiota: An overview. BioMed Res. Int. 2018, 2018, 9478630. [Google Scholar] [CrossRef] [Green Version]
- Cabello-Verrugio, C.; Simon, F.; Trollet, C.; Santibañez, J.F. Oxidative stress in disease and aging: Mechanisms and therapies 2016. Oxid. Med. Cell Longev. 2017, 2017, 4310469. [Google Scholar] [CrossRef] [PubMed]
- Kure, C.; Timmer, J.; Stough, C. The immunomodulatory effects of plant extracts and plant secondary metabolites on chronic neuroinflammation and cognitive aging: A mechanistic and empirical review. Front Pharmacol. 2017, 8, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Huang, H.; Dong, S.; Sha, H.; Wei, W.; Liu, C. High mobility group box-1 mediates hippocampal inflammation and contributes to cognitive deficits in high-fat high-fructose diet-induced obese rats. Brain Behav. Immun. 2019, 82, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Sang, Y.; Zhang, F.; Wang, H.; Yao, J.; Chen, R.; Zhou, Z.; Yang, K.; Xie, Y.; Wan, T.; Ding, H. Apigenin exhibits protective effects in a mouse model of d-galactose-induced aging via activating the Nrf2 pathway. Food Funct. 2017, 8, 2331–2340. [Google Scholar] [CrossRef]
- Yang, W.; Shi, L.; Chen, L.; Zhang, B.; Ma, K.; Liu, Y.; Qian, Y. Protective effects of perindopril on d-galactose and aluminum trichloride induced neurotoxicity via the apoptosis of mitochondria-mediated intrinsic pathway in the hippocampus of mice. Brain Res. Bull. 2014, 109, 46–53. [Google Scholar] [CrossRef]
- Dinan, T.G.; Stanton, C.; Cryan, J.F. Psychobiotics: A novel class of psychotropic. Biol. Psychiatry 2013, 74, 720–726. [Google Scholar] [CrossRef]
- Mörkl, S.; Butler, M.I.; Holl, A.; Cryan, J.F.; Dinan, T.G. Probiotics and the microbiota-gut-brain axis: Focus on psychiatry. Curr. Nutr. Rep. 2020, 9, 171–182. [Google Scholar] [CrossRef]
- Sharma, R.; Kapila, R.; Dass, G.; Kapila, S. Improvement in Th1/Th2 immune homeostasis, antioxidative status and resistance to pathogenic E. coli on consumption of probiotic Lactobacillus rhamnosus fermented milk in aging mice. Age 2014, 36, 9686. [Google Scholar] [CrossRef]
- Schifano, E.; Zinno, P.; Guantario, B.; Roselli, M.; Marcoccia, S.; Devirgiliis, C.; Uccelletti, D. The foodborne strain Lactobacillus fermentum MBC2 triggers pept-1-dependent pro-longevity effects in Caenorhabditis elegans. Microorganisms 2019, 7, 45. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Li, S.; Yang, D.; Qiu, L.; Wu, Y.; Wang, D.; Shah, N.P.; Xu, F.; Wei, H. A novel strain of Lactobacillus mucosae isolated from a Gaotian villager improves in vitro and in vivo antioxidant as well as biological properties in D-galactose-induced aging mice. J. Dairy Sci. 2016, 99, 903–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Xia, Y.; Wang, G.; Xiong, Z.; Zhang, H.; Lai, F.; Ai, L. Lactobacillus plantarum AR501 Alleviates the Oxidative Stress of D-Galactose-Induced Aging Mice Liver by Upregulation of Nrf2-Mediated Antioxidant Enzyme Expression. J. Food Sci. 2018, 83, 1990–1998. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.Y.; Chen, L.H.; Wang, M.F.; Hsu, C.C.; Chan, C.H.; Li, J.X.; Huang, H.Y. Lactobacillus paracasei PS23 delays progression of age-related cognitive decline in senescence accelerated mouse prone 8 (SAMP8) mice. Nutrients 2018, 10, 894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hor, Y.Y.; Ooi, C.H.; Lew, L.C.; Jaafar, M.H.; Lau, A.Y.; Lee, B.K.; Azlan, A.; Choi, S.B.; Azzam, G.; Liong, M.T. The molecular mechanisms of probiotic strains in improving ageing bone and muscle of d-galactose-induced ageing rats. J. Appl. Microbiol. 2021, 130, 1307–1322. [Google Scholar] [CrossRef]
- Kim, C.S.; Cha, L.; Sim, M.; Jung, S.; Chun, W.Y.; Baik, H.W.; Shin, D.M. Probiotic supplementation improves cognitive function and mood with changes in gut microbiota in community-dwelling older adults: A randomized, double-blind, placebo-controlled, multicenter trial. J. Gerontol. Ser. A 2021, 76, 32–40. [Google Scholar] [CrossRef]
- Akbari, E.; Asemi, Z.; Daneshvar Kakhaki, R.; Bahmani, F.; Kouchaki, E.; Tamtaji, O.R.; Hamidi, G.A.; Salami, M. Effect of probiotic supplementation on cognitive function and metabolic status in Alzheimer’s disease: A randomized, double-blind and controlled trial. Front Aging Neurosci. 2016, 10, 256. [Google Scholar] [CrossRef] [Green Version]
- Flanagan, E.; Lamport, D.; Brennan, L.; Burnet, P.; Calabrese, V.; Cunnane, S.C.; De Wilde, M.C.; Dye, L.; Farrimond, J.A.; Lombardo, N.E.; et al. Nutrition and the ageing brain: Moving towards clinical applications. Ageing Res. Rev. 2020, 62, 101079. [Google Scholar] [CrossRef]
- Khalesi, S.; Vandelanotte, C.; Thwaite, T.; Russell, A.M.; Dawson, D.; Williams, S.L. Awareness and attitudes of gut health, probiotics and prebiotics in Australian adults. J. Diet. Suppl. 2021, 18, 418–432. [Google Scholar] [CrossRef]
- Guarino, A.; Guandalini, S.; Vecchio, A.L. Probiotics for prevention and treatment of diarrhea. J. Clin. Gastroenterol. 2015, 49, S37–S45. [Google Scholar] [CrossRef]
- Khalesi, S.; Bellissimo, N.; Vandelanotte, C.; Williams, S.; Stanley, D.; Irwin, C. A review of probiotic supplementation in healthy adults: Helpful or hype? Eur. J. Clin. Nutr. 2019, 73, 24–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misra, S.; Mohanty, D.; Mohapatra, S. Applications of probiotics as a functional ingredient in food and gut health. J. Food Nutr. Res. 2019, 7, 213–223. [Google Scholar] [CrossRef]
- Sanders, M.E.; Merenstein, D.; Merrifield, C.A.; Hutkins, R. Probiotics for human use. Nutr. Bull. 2018, 43, 212–225. [Google Scholar] [CrossRef] [Green Version]
- Pitchumoni, C.; Mishra, S.P.; Yadav, H. Gut Microbiota and Aging: A Broad Perspective. In Geriatric Gastroenterology; Spinger: Berlin/Heidelberg, Germany, 2020; pp. 1–21. [Google Scholar] [CrossRef]
- Kaku, N.; Matsumoto, N.; Sasaki, D.; Tsuda, K.; Kosai, K.; Uno, N.; Morinaga, Y.; Tagami, A.; Adachi, S.; Hasegawa, H.; et al. Effect of probiotics on gut microbiome in patients with administration of surgical antibiotic prophylaxis: A randomized controlled study. J. Infect. Chemother. 2020, 26, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Van Schoor, J. Probiotics and gut health. SA Pharm. Assist. 2020, 20, 32–33. [Google Scholar] [CrossRef]
- Bubnov, R.V.; Spivak, M.Y.; Lazarenko, L.M.; Bomba, A.; Boyko, N.V. Probiotics and immunity: Provisional role for personalized diets and disease prevention. EPMA J. 2015, 6, 14. [Google Scholar] [CrossRef]
Figure 1.
Probiotic mechanisms of action brief overview.

Figure 2.
Probiotics relation to regulate aging and gut health modulation.


{kind=link}
{kind=link}
Table 1.
Probiotics as a gut health modulator.
| Probiotic(s) | Animal Model | Effects on Host | Results | References |
|---|---|---|---|---|
| Lactobacillus fermentum | Weanling Mice | Gut health modulation | Increased mRNA expression of claudin-1 and MUC-2 in intestinal epithelial cells. | [23] |
| Lactobacillus paracasei | Old mice | Gut health modulation | Improved mucin production, decreased leaky gut and inflammation. | [29] |
| Bifidobacterium longum and Lactobacillus rhamnosus | Healthy volunteers | Gut health modulation | Decreased Firmicutes abundance, the overall reduction of potentially harmful bacteria, and an increase in beneficial bacteria. | [56] |
| 5 Lactobacillus and 5 Enterococcus strains (Coaktail-Human originated) | Older mice | Gut health modulation | Increased the beneficial commensals in older HFD mice and decreased the expression of IL-6, TNF-α, and IL-1β, while increasing the anti-inflammatory markers, such as IL-10 and TGF-β expression in the colon tissues. Improved the mRNA expression of tight junction proteins such as Zo1 and Ocln in the intestinal tissues. | [59] |
| Lactobacillus fermentum, Lactobacillus fermentum, and Lactobacillus salivarius | Broiler chicken | Gut health modulation | Improved the villus height and villus-height-to-crypt-depth ratio improved the gut morphometric parameters and absorption capacity. | [55] |
| Lactobacilli strains | D-Galactose senescence-induced aging rats | Gut health modulation | Reduced Bacteroides, increased the ratio of Firmicutes/Bacteroidetes. | [57] |
| Lactobacillus acidophilus and Bifidobacterium infantis | Mouse pre-weaned pups | Gut health modulation | Supported intestinal epithelial-cell differentiation, reduced loss of mucin, protected the intestinal integrity and barrier function, and reduced serum levels of IL-1β, TNF-α, and IL-6. | [65] |
Table 2.
Probiotics as anti-aging agent (Gut–Brain axis).
| Probiotic(s) | Animal Model | Effects on Host | Results | References |
|---|---|---|---|---|
| Lactobacillus brevis | Aged mice | Anti-aging | Suppressed the expression of senescence markers p16, p53, and SAMHD1 and restored expression of brain-derived neurotrophic factor and doublecortin in aged mice. | [5] |
| Bifidobacterium longum and Bifidobacterium animalis | D-Galactose-treated mice | Anti-aging | Improved the anxiety-like behavior, uncoordinated movement, cognitive decline, and hippocampus senescence; ameliorate age-related cognitive degeneration by inhibiting NF-κB/TLR4-induced-neuroinflammation and oxidative stress. | [11] |
| Lactobacillus paracasei | Old mice | Anti-aging | Improved physical and cognitive functions, modulating the TLR-2/p38-MAPK/NF-kB pathway, which may reduce age-related leaky gut and inflammation. | [29] |
| Lactobacillus fermentum | C. elegans | Anti-aging | Improved pumping rate, lipofuscin accumulation, and body bending. | [84] |
| Lactobacillus plantarum | D-Galactose-treated mice | Anti-aging | Reduced abnormal activities of superoxide dismutase, glutathione peroxidase, and catalase. Reduced expressions of several antioxidant genes, such as glutathione reductase, glutathione S-transferase, glutamate–cysteine ligase catalytic subunit, glutamate–cysteine ligase modifier subunit, and NAD(P)H quinone oxidoreductase 1. | [86] |
| Lactobacillus paracasei | Senescence-accelerated mouse prone 8 (SAMP8) mice | Anti-aging | Reduced senescence and low-serious anxiety-like behaviors and memory impairment and enhanced the antioxidative enzymes superoxide dismutase (SOD) and glutathione peroxidase (GPx). | [87] |
| Lactobacillus fermentum | D-Galactose induced aging rats | Anti-aging | Improved expression of SOD in bone and muscle, increased AMPK-α2 expression, and reduced the expression of IL-6 and TRAP in tibia. | [88] |
| B. bifidum BGN4 and B. longum BORI | Older adults | Brain health (gut–brain axis) | Alleviated stress and improved mental flexibility in older adults, along with modulating gut microbiota. | [89] |
| L. acidophilus, L. casei, B. bifidum, and L. fermentum | Alzheimer’s disease AD (60–95 years age) | Brain health | Findings of the study suggested that the probiotic-treated group had shown considerable improvement in Mini-mental state examination score. Overall study concluded that probiotic supplementation for 12 weeks could improve the metabolic status and cognitive functioning in the AD patients. | [90] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Samtiya, M.; Puniya, A.K.; Puniya, M.; Shah, N.P.; Dhewa, T.; Vemuri, R. Probiotic Regulation to Modulate Aging Gut and Brain Health: A Concise Review. Bacteria 2022, 1, 250-265. https://doi.org/10.3390/bacteria1040019
AMA Style
Samtiya M, Puniya AK, Puniya M, Shah NP, Dhewa T, Vemuri R. Probiotic Regulation to Modulate Aging Gut and Brain Health: A Concise Review. Bacteria. 2022; 1(4):250-265. https://doi.org/10.3390/bacteria1040019
Chicago/Turabian StyleSamtiya, Mrinal, Anil Kumar Puniya, Monica Puniya, Nagendra P. Shah, Tejpal Dhewa, and Ravichandra Vemuri. 2022. "Probiotic Regulation to Modulate Aging Gut and Brain Health: A Concise Review" Bacteria 1, no. 4: 250-265. https://doi.org/10.3390/bacteria1040019