1. Introduction
During embryonic development, new blood vessels arise from angioblasts within the yolk sac as well as the splanchnopleuric mesoderm of the lateral plate. This process of
de novo blood vessel formation is termed vasculogenesis. From these first vessels, new vessel branches are formed in a process called angiogenesis, which describes the sprouting of endothelial cells from already existing vessels [
1]. The arising primitive vascular networks fuse to form a continuous system and as soon as the embryonic heart starts to beat, vessels are perfused by blood. Upon perfusion, vessel wall maturation and remodeling are induced by the blood pressure and by shear forces [
2].
The typical wall of larger blood vessels consists of three layers. The innermost layer is called
tunica intima. It contains endothelial cells that line the vascular lumen. A layer of smooth muscle cells, called
tunica media, is stored around the intima. The outermost layer of the vascular wall is formed by a loose connective tissue, the so-called
tunica adventitia, which integrates the vessel into the surrounding tissue. Peripheral nerves run here, as well as smaller capillary vessels called
vasa vasorum, which nourish the vessel wall. There is also a versatile stem cell niche that contains vascular and hematopoietic stem cells [
3,
4].
Small capillaries, on the other hand, consist of an endothelial tube that is partially surrounded by perivascular cells. These are embedded in the endothelial basement membrane and are in close contact with the endothelial cells via peg–socket-like structures [
5] at which gap junctional coupling is observed mediated by Connexin43 [
6]. These perivascular cells are called pericytes.
Organoids are stem cell-based 3D-cell culture models which recapitulate different aspects of normal embryonic and fetal tissue development and fulfill at least some of the tissue/organ-specific functions [
7]. Different organoid models have been established within the last years, among these, e.g., brain [
8], intestinal [
9], lung [
10], or blood vessel organoids [
11]. In this study, we use human induced pluripotent stem cells (iPSCs) as the cellular basis of the organoid model. Human iPSCs can be derived from primary somatic cell types such as skin fibroblasts or mononucleated peripheral blood cells via transcription factor-driven epigenetic reprogramming [
12,
13].
In the past, studies on developmental biology were mostly carried out by utilizing animal models. Nowadays, organoids allow such examinations directly on human tissues. Primary human embryonic and fetal tissues are usually hard to access. In contrast, organoids can be easily manufactured in large quantities and closely mimic the real tissues in many aspects. In addition, efficient gene-editing tools such as CRISPR/Cas9 allow researchers to introduce mutations to address the function of genes during development. Moreover, organoids can be used for disease modeling, high throughput drug testing, and toxicity screenings or as building blocks for biofabrication and tissue engineering. Finally, organoids help to reduce the need for animal experimentation.
Here we present a new, reproducible, and cost-effective blood vessel organoid model which recapitulates different aspects of embryonic blood vessel development including vasculogenesis, angiogenesis as well as vessel wall maturation. The resulting vessels display a multi-layered vessel wall and are blood-perfused after transplantation on the chicken chorioallantoic membrane. We assume that our organoid model will help to uncover mechanisms underlying early human blood vessel development and maturation. Moreover, our blood vessel organoids can be used for drug screening or toxicity assessment and might be used as building blocks for biofabrication processes.
2. Material and Methods
2.1. Induced Pluripotent Stem Cell (iPSC) Culture
Two human foreskin-derived male iPSC lines are utilized (STEMCCA NHDF iPSCs and Sendai NHDF iPSCs) as previously described [
14].
iPS cells are cultured on human ESC Matrigel (Corning, Tewksbury, MA, USA) coated 6 well plates in StemMACS iPS Brew XF human (Miltenyi Biotec, Bergisch Gladbach, Germany) medium. Cells are passaged as soon as they reach 80% confluency using StemProAccutase (Gibco/Life Technologies, Westham, MA, USA). Afterward, cells are reseeded in the desired cell density in StemMACS iPS Brew XF human medium supplemented with 10 mM ROCK inhibitor Y-27632 (Miltenyi Biotec, Bergisch Gladbach, Germany).
2.2. Generation of 2D Mesodermal Progenitor Cells (MPCs)
For the induction of MPCs, 80% confluent iPSC cultures are dissociated and detached from the culture dish using StemProAccutase cell detachment solution (Gibco/Life Technologies, Westham, MA, USA). Cells are counted using a Neubauer counting chamber and reseeded at a cell density of 3.5 × 105 cells in 1.5 mL StemMACS iPS Brew medium, supplemented with 10 mM ROCK inhibitor Y-27632, per well. For MPC induction human Matrigel-coated 6-well plates are used. After 24 h cell cultivation, medium was changed daily to mesoderm induction medium (MIM) (Advanced DMEM-F12 (Gibco/Life Technologies, Westham, MA, USA) 100%, L-Glutamine (Gibco/Life Technologies, Westham, MA, USA) 2 mM, Ascorbic acid (Sigma-Aldrich, St. Louis, MO, USA) 0.06 mg/mL (355 µM), CHIR 99,210 (Sigma-Aldrich, St. Louis, MO, USA) 10 µM, BMP4 (Pepro Tech, Cranbury, NJ, USA) 25 ng/mL) for 72 h. MPCs are cultured at 37 °C and 5% CO2 in a humidified incubator.
2.3. Blood Vessel Organoid Generation
For the induction of blood vessel organoids, 80% confluent iPSC cultures are dissociated and detached from the culture dish using StemProAccutase (Gibco/Life Technologies, Westham, MA, USA). Cells are counted using a Neubauer counting chamber. Subsequently, a single cell suspension with the desired cell density (500–4000 cells per 100 µL StemMACS iPS Brew medium (Miltenyi Biotec, Bergisch Gladbach, Germany) supplemented with 10 mM ROCK inhibitor Y-27632) is prepared. A 100 µL iPSC suspension is pipetted per well of an agarose-coated 96-well plate. To prepare agarose-coated wells, 1% agarose (Biozym, Hessisch Oldendorf, Germany,) is boiled in water and 50 µL liquid gel is pipetted into each well of the 96-well plate as previously described [
15]. Cells are cultured for 24 h at 37 °C and 5% CO
2 in a humidified incubator to allow iPSC aggregate formation. A single aggregate forms per well of the 96-well plate. After 24 h, medium is changed to mesoderm induction medium (MIM) (Advanced DMEM-F12 (Gibco/Life Technologies, Westham, MO, USA) 100%, L-Glutamine (Gibco/Life Technologies, Westham, MO, USA) 2 mM, Ascorbic acid (Sigma-Aldrich, St. Louis, MO, USA) 0.06 mg/mL (355 µM), CHIR 99,210 (Sigma-Aldrich, St. Louis, MO, USA) 10 µM, BMP4 (Pepro Tech, Cranbury, NJ, USA) 25 ng/mL) for 72 h. After that, medium is discarded, and organoids are cultivated in vascular growth medium (VGM) (Neurobasal medium/DMEM-F12 (Gibco/Life Technologies, Westham, MO, USA) (50%/50%), B27 without Vitamin A (Gibco Life Technologies, Westham, MO, USA) 1×, N2-Supplement (Gibco Life Technologies, Westham, MO, USA) 1×, L-Glutamine (Gibco Life Technologies, Westham, MO, USA) 2 mM, Ascorbic acid 0.06 mg/mL, VEGF (ProteinTech, Rosemont, IL, USA) 100 ng/mL). After 48 h, medium is changed to organoid maintenance medium (OMM) (Neurobasal medium/DMEM-F12 (50%/50%), B27 without Vitamin A 1×, N2-Supplement 1×, L-Glutamine 2 mM, Ascorbic acid 0.06 mg/mL) and organoids are cultured at 37 °C and 5% CO
2 in a humidified incubator on a cell culture rocking table. Organoids are kept in maintenance medium until they reach the desired age. Medium is changed every other day.
2.4. RT-PCR Analyses
Total RNA samples were extracted using the Direct-Zol RNA MiniPrep Plus Kit (Zymo Research, Freiburg, Germany). cDNA was generated using the GoScriptTM Reverse Transcriptase (Promega, Fitchburg, WI, USA). Semiquantitative RT-PCR analyses were performed using the Red MasterMix (2x) Taq PCR MasterMix (Genaxxon, Ulm, Germany) and a T3 Thermocycler (Biometra, Jena, Germany). The following primer pairs were used: Brachyury (forw: TATGAGCCTCGAATCCACATAGT, rev: CCTCGTTCTGATAAGCAGTCAC), KDR (forw: TTTTTGCCCTTTTGTTCTGTCC, rev: TCATTGTTCCCAGCATTTCA), MEOX (forw: TGACAACTTTGGTATCGTGGA, rev: CCAGTAGAGGCAGGGATGAT), CXCR4 (forw: ACTACACCGAGGAAATGGGCT, rev: CCCACAATGCCAGTTAAGAAGA), PDGFRa (forw: TTTTTGTGACGGTCTTGGAAGT, rev: TGTCTGAGTGTGGTTGTAATAGC), GAPDH (forw: TGACAACTTTGGTATCGTGGA, rev: CCAGTAGAGGCAGGGATGAT).
2.5. Flow Cytometric Analyses
Flow cytometry was performed using a FACSCanto II (BD Biosciences, San Jose, CA, USA) and data were analyzed using the FACSDiva software version 6.1.3 (BD Biosciences, San Jose, CA, USA). To discriminate between dead and living cells, viability stain 450 (BD Biosciences) was used according to the manufacturer’s recommendation. Cells were fixed using 4% PFA in PBS and washed 2× with PBS. Subsequently, samples were transferred in blocking buffer (5% BSA in PBS) and incubated for 1 h. Afterward, cells were incubated in blocking buffer containing 0.1% Triton X-100. Cell suspensions were incubated with specific antibodies targeted against Brachyury (T) (R&D Systems, USA, Minneapolis, MN, AF2085) for 1 h in blocking buffer. Cells were washed 3 times and incubated with Cy5 conjugated secondary antibodies in PBS for 45 min. Unstained cells and cells treated with secondary antibodies alone were used as negative controls.
2.6. Histological Analyses
Organoids are washed gently in PBS and fixed in 4% PFA (in PBS) (Sigma-Aldrich, St. Louis, MO, USA) at 4 °C overnight. Subsequently, fixed organoids are washed in PBS (Sigma-Aldrich, St. Louis, MO, USA) and embedded in agarose gel (1%). Afterward, 5 μm paraffin (Merck) sections are prepared as previously described [
15]. To perform staining, paraffin sections are deparaffinized, rehydrated, and for a general overview of the tissue architecture stained with hematoxylin and eosin. For immunofluorescence analyses, sections are unmasked using Sodium Citrate buffer (10 mM, pH6). Primary antibodies targeted against CD31 (Agilent, M0823, mouse, 1:200), CD31 (Abcam, Cambridge, UK, ab28364, rabbit, 1:200) CD34 (Agilent, Santa Clara, CA, USA, M7165, mouse 1:200), Cx43 (Abcam, Cambridge, UK, ab11370, rabbit, 1:600), SMA (Abcam, Cambridge, UK, ab7817, mouse, 1:200), Gata6 (R&D Systems, St. Louis, MO, USA, AF1700, goat, 1:200), RGS5 (Proteintech, Rosemont, IL, USA, rabbit, 1:200) were diluted in blocking solution (4% BSA, 0.2% Triton-X-100 in PBS) and incubated for 16 h at 4 °C. Secondary Cy2-, Cy3- or Cy5-conjugated antibodies (Dianova, Hamburg, Germany) were used to visualize the primary antibodies. Secondary antibodies were diluted in PBS and incubated for 1 h at room temperature. Nuclei were stained with DAPI solution (1 µg/mL in PBS). Images were taken using an Axiovert 40 CFL microscope (Zeiss, Jena, Germany) or a Nikon Eclipse Ti confocal laser scanning microscope (Nikon, Tokyo, Japan).
2.7. Tissue Clearing
For visualization of blood vessel formation, a whole-mount tissue clearing was performed as previously described [
15].
Following primary antibodies were used: CD31 (Agilent, Santa Clara, CA, USA, mouse, M0823, 1:200), Gata6 (R&D Systems, St. Louis, MO, USA, AF1700, goat, 1:200). Cy2 and Cy3 conjugated secondary antibodies (Dianova, Hamburg, Germany,) were used to visualize the primary antibodies. Imaging is performed using the Nikon Eclipse Ti confocal laser scanning microscope (Nikon, Tokyo, Japan) with long working distance air objectives (4×, 20×) for taking z-stack images. Nikon NIS Elements Confocal software version 4.13.05 (Nikon, Tokyo, Japan) is used for 3D reconstruction of imaged blood vessel organoids.
2.8. Transmission Electron Microscopy (TEM)
Samples were transferred to fixation reagent (2.5% glutaraldehyde, 4% PFA in 0.1 M cacodylate buffer (50 mM cacodylate, 50 mM KCl, 2.5 mM MgCl2, pH7.2)) and incubated for 2 h on ice. Afterward, samples were washed with 0.1 M cacodylate buffer, followed by a second fixation step using 1% osmium tetroxide in 0.1 M cacodylate buffer for 1 h. Subsequently, specimens were washed 2× for 10 min in 0.1 M cacodylate buffer and 1× in a. bidest. Fixed organoids were dehydrated in an ascending EtOH series using 30%, 50%, 70% EtOH, 10 min each, and afterward incubated in 2% uranyl acetate in 70% EtOH for 60 min. Next, samples were dehydrated using an EtOH array of 70%, 80%, 90%, 96%, and 2× 100% for 10 min each and subsequently incubated in propylene oxide (PO) for 2× 30 min. Finally, specimens were incubated in PO/Epon812 (1:1) overnight. The next day, samples were transferred into Epon for 2 h and embedded by Epon polymerization at 60 °C for 48 h.
Specimen were cut using an ultramicrotome, collected on nickel grids, and stained with 2.5% uranyl acetate and 0.2% lead citrate. Pictures were taken using an LEO AB 912 transmission electron microscope (Zeiss, Jena, Germany).
2.9. Grafting of Blood Vessel Organoids on Chicken Chorioallantois Membrane (CAM)
Two 20-days-old blood vessel organoids were transferred in 10 µL Matrigel and placed on a round nylon mesh with a diameter of approximately 1 cm (150 µm grid-size, 50% open surface, 62 µm string diameter, 35 g/m2, PAS2, Hartenstein, Germany, Würzburg). After Matrigel solidification, the nylon mesh was placed upside down on the CAM at embryonic day (ED) 7. After 5–10 days, the mesh with the organoids was carefully explanted and washed in PBS. Afterward, tissue was fixed for 1 h using 4% PFA. Subsequently, 5 µm paraffin sections were prepared. Immunofluorescence analyses were performed as described above. All procedures comply with the EU regulations covering animal experimentation (European Communities Council DIRECTIVE 2010/63/EU) and were conducted in accordance with the animal care guidelines of the University of Würzburg.
3. Results
Human induced pluripotent stem cells (hiPSCs) were differentiated towards a lateral plate mesoderm fate by activation of the Wnt signaling pathway using the specific glycogen synthase-kinase 3 (GSK3) inhibitor Chir99021 [
16] and BMP4 supplementation for 3 days. When the differentiation protocol was applied in 2D cell culture, a striking change in cell morphology was already observed at day 2 of differentiation (
Figure 1A,B). The induced mesodermal progenitor cells (MPCs) showed mRNA expression of the mesendodermal marker genes Brachyury
(T) [
17] and
CXCR4 [
18] as well as the lateral plate mesoderm marker gene
KDR (VEGF receptor 2) [
19]. In contrast, an expression of the paraxial mesoderm markers
PDGFRa [
19] and
MEOX [
20] was hardly detectable indicating differentiation mainly towards the lateral plate mesoderm fate (
Figure 1C). Flow cytometric analysis revealed that 85% of the cells expressed Brachyury at day 2 of differentiation (
Figure 1D).
Next, we applied the same differentiation protocol to iPSC aggregates in 3D suspension culture. For that purpose, an iPSC single-cell suspension was prepared and a defined number of 500 cells was seeded per well of an agarose-coated 96-well plate. The agarose coating prevents cells from attaching to the culture dish and forms a conical mold in which the cells form regular-shaped, round 3D aggregates within 24 h (
Figure 1E). Aggregates were cultured in the mesoderm induction medium (MIM) supplemented with Chir99021 and BMP4 for 3 days. From day 5 on, aggregates were cultured in a vascular growth medium (VGM). At day 5, vasculogenesis was stimulated by a single dose of 100 ng/mL vascular endothelial growth factor (VEGF) and organoids were cultivated for up to 20 days (
Figure 1E). To prove the successful mesoderm induction in 3D suspension culture, day 3 organoids were enzymatically dissociated into a single cell suspension and analyzed by flow cytometric analysis using a specific Brachyury antibody. The experiments revealed that 88% of the cells showed Brachyury expression confirming efficient iPSC conversion towards a mesodermal cell fate (
Figure 1F).
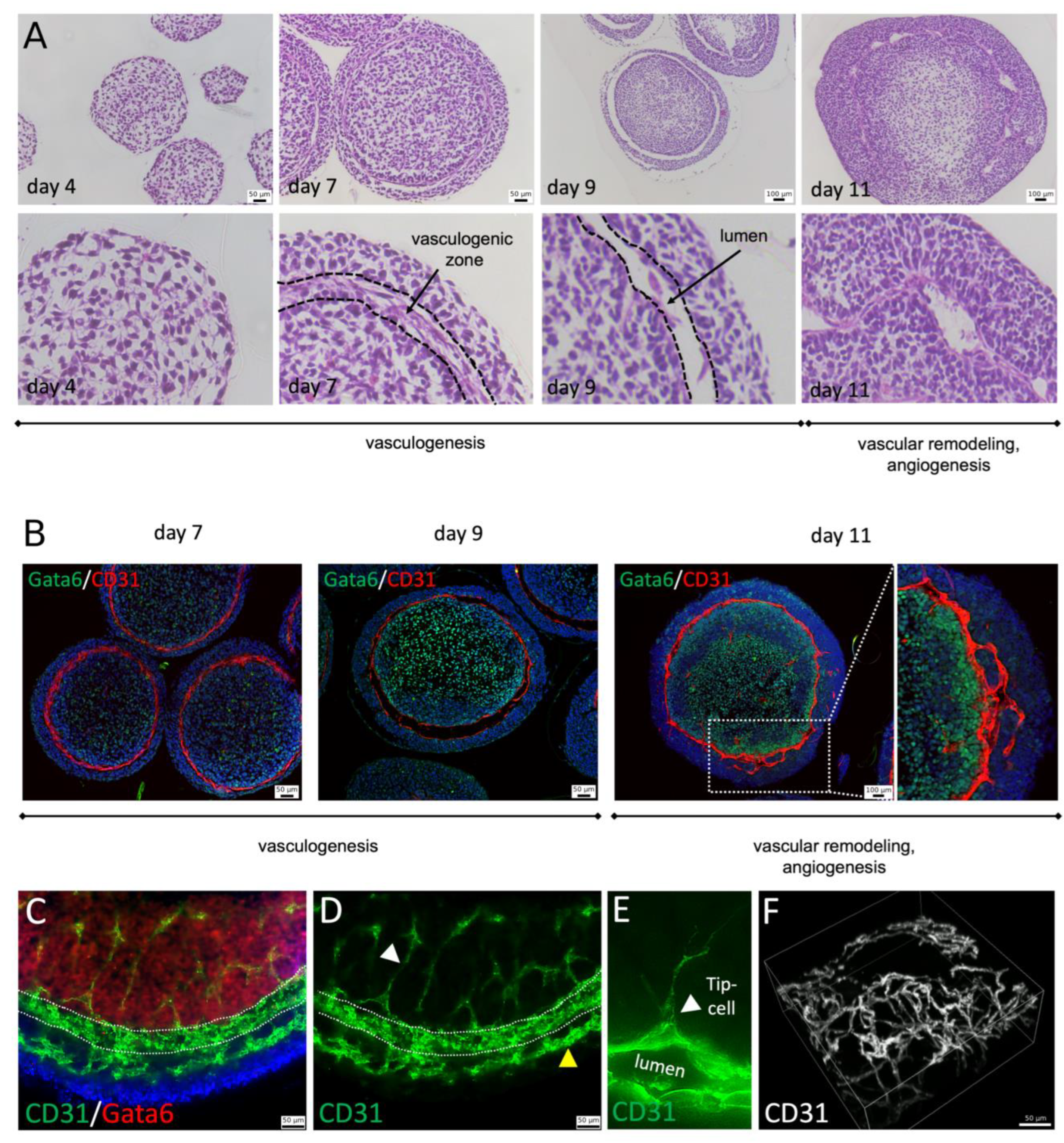
Paraffin sections of blood vessel organoids were prepared at day 4, 7, 9, and 11 of differentiation. At day 4, the organoid consists of loosely connected mesenchymal cells and displays a morphologically uniform appearance. At day 7, three tissue layers can be observed with loosely arranged mesenchymal cells in the center and more densely packed cells at the surface of the organoid. A ring-shaped vasculogenic zone forms in between these two tissue layers (
Figure 2A). At day 9, vessel lumen formation can be observed within the vasculogenic zone. At day 11, blood vessel-like structures are detectable and angiogenic sprouting into the outer and inner tissue layer is observed (
Figure 2A,B and
Figure S1A). The developing vasculogenic zone displays an expression of the endothelial cell marker CD31 (PECAM1). The same is true for cells lining the vascular lumen or cells sprouting from the vasculogenic ring into the outer and inner tissue layers confirming their endothelial cell identity (
Figure 2B–F). Interestingly, the inner tissue compartment shows an expression of GATA6, which is among others a marker for lateral plate mesoderm. The outer tissue compartment is GATA6 negative (
Figure 2B,C).
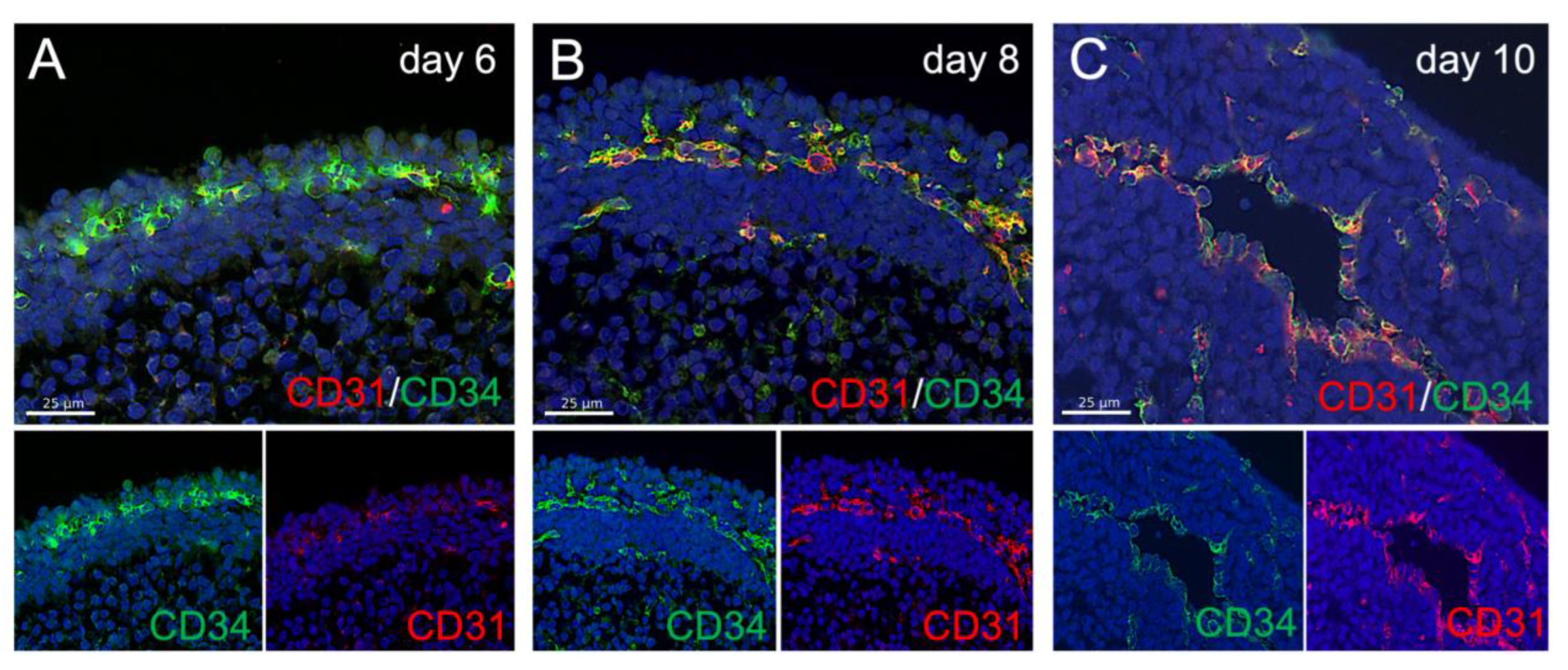
Like in the embryo, the vasculogenic zone shows an expression of the endothelial progenitor marker CD34 before the emergence of CD31 (
Figure 3A). Later, during primitive vessel formation, CD31 and CD34 are co-expressed as observed in embryonic endothelium in vivo (
Figure 3B,C).
Moreover, we produced blood vessel organoids using different numbers of starting cells. Interestingly, the size of blood vessel organoids is scalable, and no matter how many cells are used, differentiation always results in a ring-shaped vasculogenic zone (
Figure 4A,B). Different blood vessel organoids from the same experiment are highly uniform in size and shape (
Figure S2) and can be manufactured in a 96-well plate format in large quantities. Moreover, the differentiation outcome is very uniform, resulting in the formation of a ring-shaped vasculogenic zone at day 7 of differentiation (
Figure 4C). A lumen is formed at approximately day 9 and angiogenic sprouting can be observed from day 11 on (
Figure 4C,D).
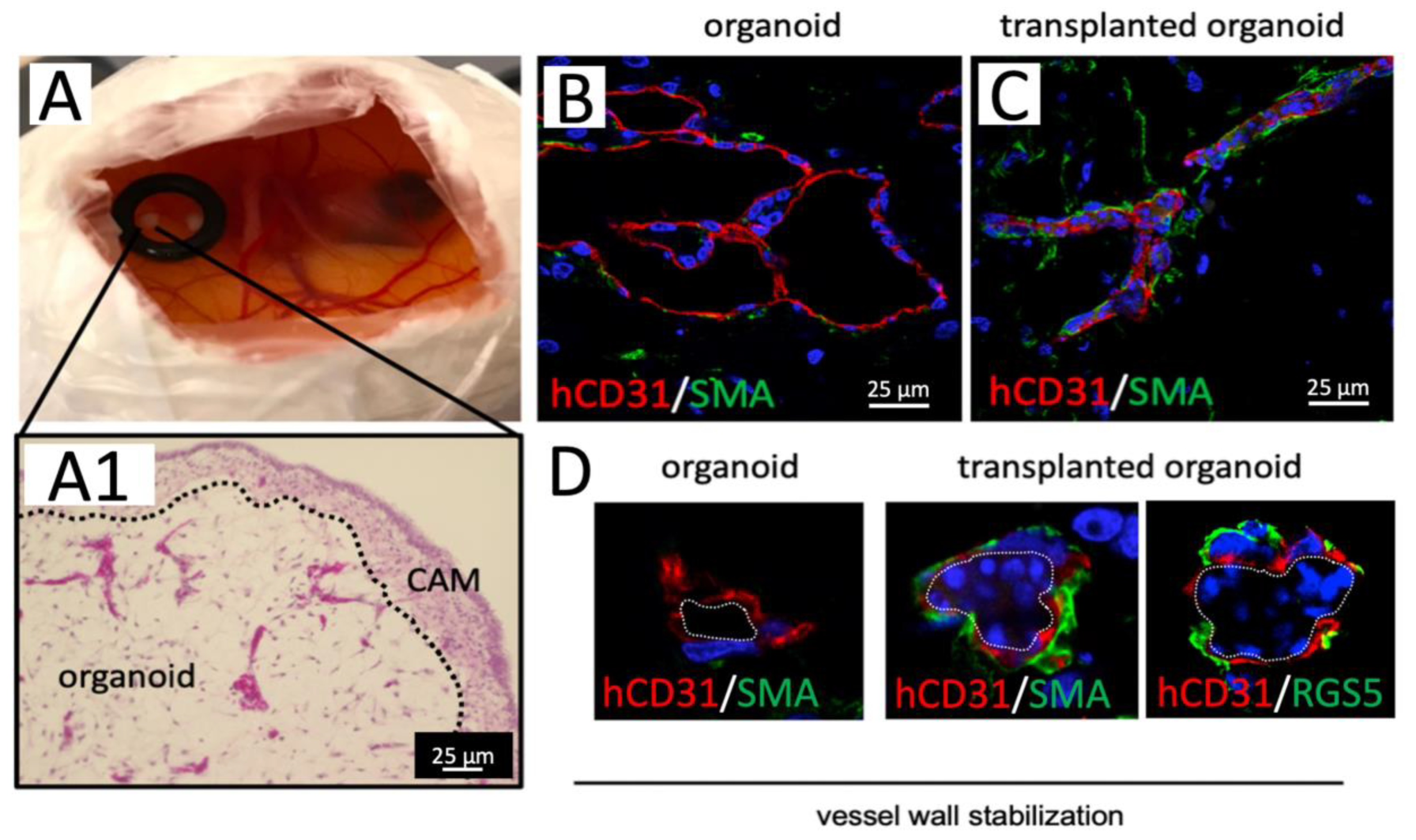
To prove blood vessel functionality, we embedded 20-days-old blood vessel organoids into basement membrane extract (BME) and transplanted them onto the chorioallantois membrane (CAM) of chicken embryos (
Figure 5A). We observed a functional connection of human blood vessels to the chicken vascular system as indicated by nucleated chicken erythrocytes and other blood cells within the human vessel lumen seven days after transplantation (
Figure 5A,C and
Figure S1B–E). A human-specific CD31 antibody was used to discriminate between chicken and human organoid vessels (
Figure 5B,C and
Figure S1D,E). We observed that blood perfusion of organoid vessels changed their morphological appearance and resulted in a reduced vessel lumen diameter and a continuous layer of smooth muscle actin (SMA) as well as RSG5 positive peri-endothelial cells [
21] around the endothelial tubes indicating vessel wall remodeling and maturation (
Figure 5B–D). SMA or RSG5 positive peri-endothelial cells are only rarely observed in non-transplanted organoids (
Figure 5B).
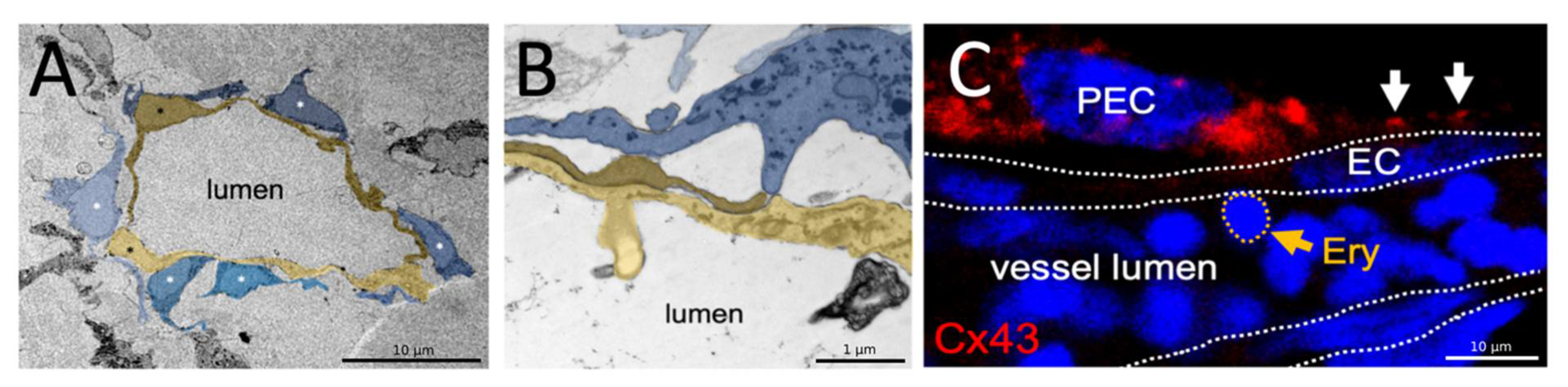
Finally, we analyzed the vascular ultrastructure using transmission electron microscopy (TEM). We observed endothelial cells lining the vascular lumen, which are closely associated with peri-endothelial cells enwrapping the endothelial tube from the outside (
Figure 6A). Higher magnification images demonstrate that peri-endothelial cells (PEC) contact the endothelial cells (EC) with finger-like processes (
Figure 6B). Immunofluorescence analyses on paraffin sections of blood perfused human organoids transplanted on the CAM revealed Connexin43 (Cx43) expression in PEC and a typical punctate Cx43 staining pattern at the EC-PEC interface indicating gap junction plaque formation, a pre-requisite for functional myo-endothelial coupling (
Figure 6C).
4. Discussion
In 2019, Wimmer and colleagues published the first paper on blood vessel organoids [
11]. The intention of their paper was the in vitro generation of a functional vascular network that gives rise to mature blood vessels following transplantation under the kidney capsule of immunodeficient mice. While the authors mainly focused on modeling vasculopathy in mature vessels, our study’s aim is to realistically mimic the early steps of blood vessel formation namely vasculogenesis, angiogenesis, and vessel wall maturation in a robust and reproducible fashion. For that reason, we added as few cytokines as possible to allow the self-organization of blood vessels by intrinsic mechanisms. In comparison to already published protocols, we left out FGF, forskolin, and the TGFb inhibitor SB43152, reduced the administration of VEGF to a single dose of 100 ng/mL on day 5, and did not embed organoids into the extracellular matrix [
11,
22,
23]. Applying this protocol, we observed that blood vessel organoids recapitulate vasculogenesis, angiogenesis, and vessel wall maturation within approximately 10 days of culture like in the early embryo. As a positive byproduct, the reduction of supplements makes the protocol easier and more cost-effective.
During vasculogenesis, angioblasts give rise to new blood vessels at distinct embryonic regions, e.g., within the lateral plate mesoderm (LPM) [
24]. In the zebra fish, a single line of etv2-expressing angioblasts can be detected in the LPM as early as the 4-somite stage [
25]. These migrate over the endoderm towards the dorsal midline and give rise to a vascular cord, which subsequently undergoes lumen formation to form the dorsal aorta [
26]. Finally, segmental arteries start to sprout from the dorsal aorta in a process termed angiogenesis [
27].
In our organoid model, we also observe a distinct zone of CD34
+ vascular progenitors which arrange in a ring-shaped pattern underneath the surface of the organoid at the interface between two distinct cell populations (
Figure 2A and
Figure 3A). Like in vivo, a CD31/CD34 double-positive vascular cord forms which subsequently undergoes lumen formation. Finally, angiogenic sprouting can be observed from the large lumenized vessel around day 10 (
Figure 3B–F and
Figure 4C). Tip cells, the leading cells of angiogenic sprouts, were detectable and identified by their characteristic filopodia (
Figure 3E) [
28].
In the last step, we transplanted the vascularized organoids on the chicken chorioallantois membrane (CAM) [
29]. The human vessels became functionally connected to the chicken’s circulatory system and underwent vessel wall maturation probably due to blood pressure and shear forces. Vessel maturation was assessed by the formation of a continuous layer of SMA/RGS5 positive cells around the endothelial tube demonstrating multi-layered vessel wall formation. For that reason, vascular organoids in combination with the CAM assay are an attractive system to investigate mechanisms of human vessel wall maturation without a need for transplantation into mice or rats. The CAM assay is inexpensive and easy to handle and can be performed in high throughput. An ethical advantage is that the CAM is not innervated at that time point of development and experiments are stopped before the emergence of brain regions associated with pain perception. In the future, we will explore the perfusion of organoids using microfluidic devices to be able to do without animal experimentation completely [
30].
We assume, that the presented organoid model recapitulates vasculogenesis and angiogenesis like in the embryo and therefore represents an in vitro 3D-cell culture platform to investigate these early developmental processes and help to reveal the mechanisms underlying early blood vessel development directly within the human tissue context. In combination with CRISPR/Cas9 mediated gene editing, the human organoids represent a powerful tool for studying human vascular developmental biology [
31]. Moreover, by utilizing blood vessel organoids the impact of toxins or drugs on vascular development can be screened to identify teratogenic substances [
32]. Finally, blood vessel organoids could be used as building blocks for biofabrication. As our organoids are scalable, very small ones could be used for bioprinting together with tissue-specific cell types and combined with larger biofabricated vessels and microfluidic systems to engineer large and functionally vascularized tissue constructs.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





