
Vitamin E and A Availability in Goose Embryos and Goslings and Improvement of Reproduction Traits Depending on the Starting Temperature Regime of Egg Incubation
 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
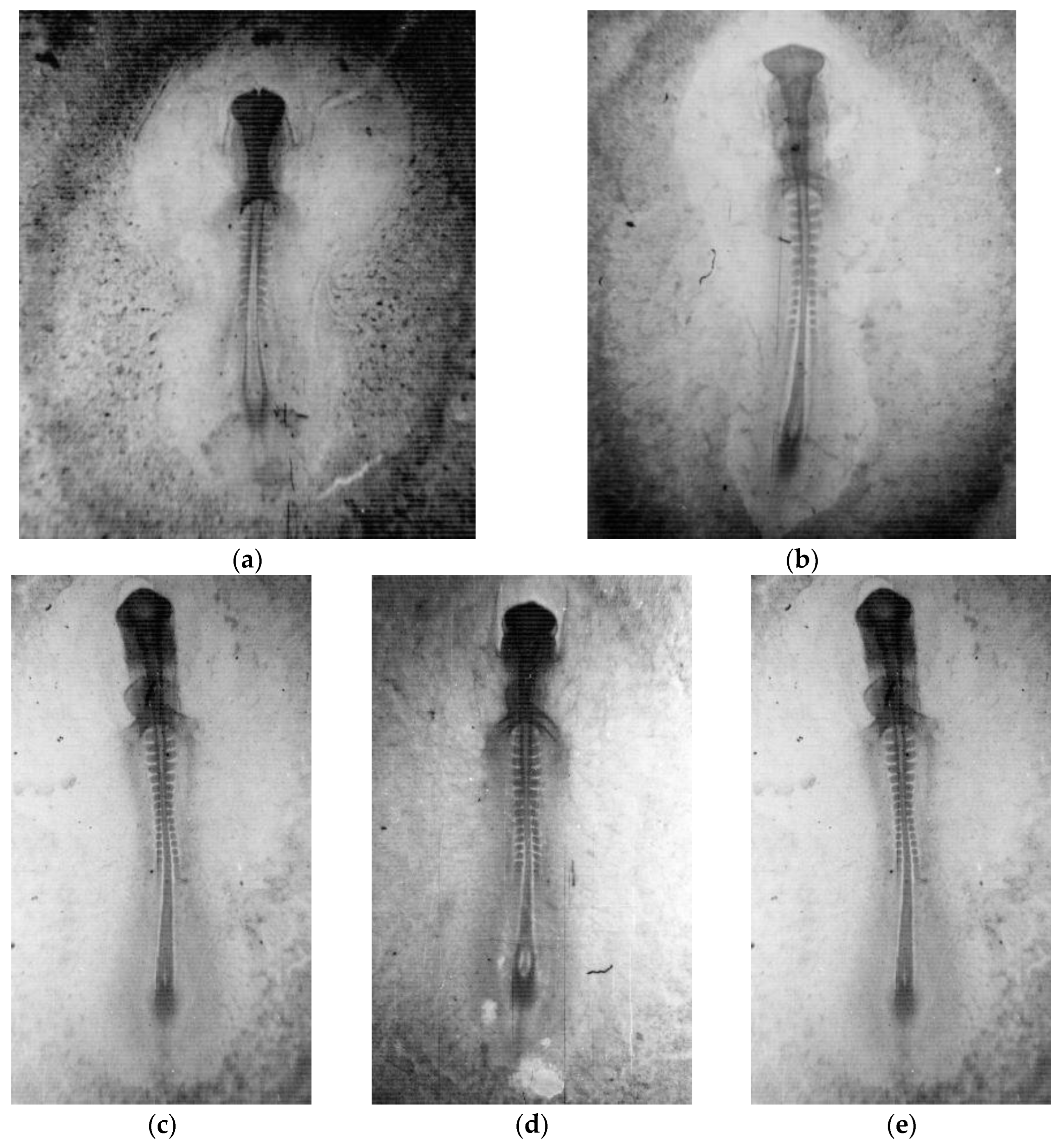
2. Materials and Methods
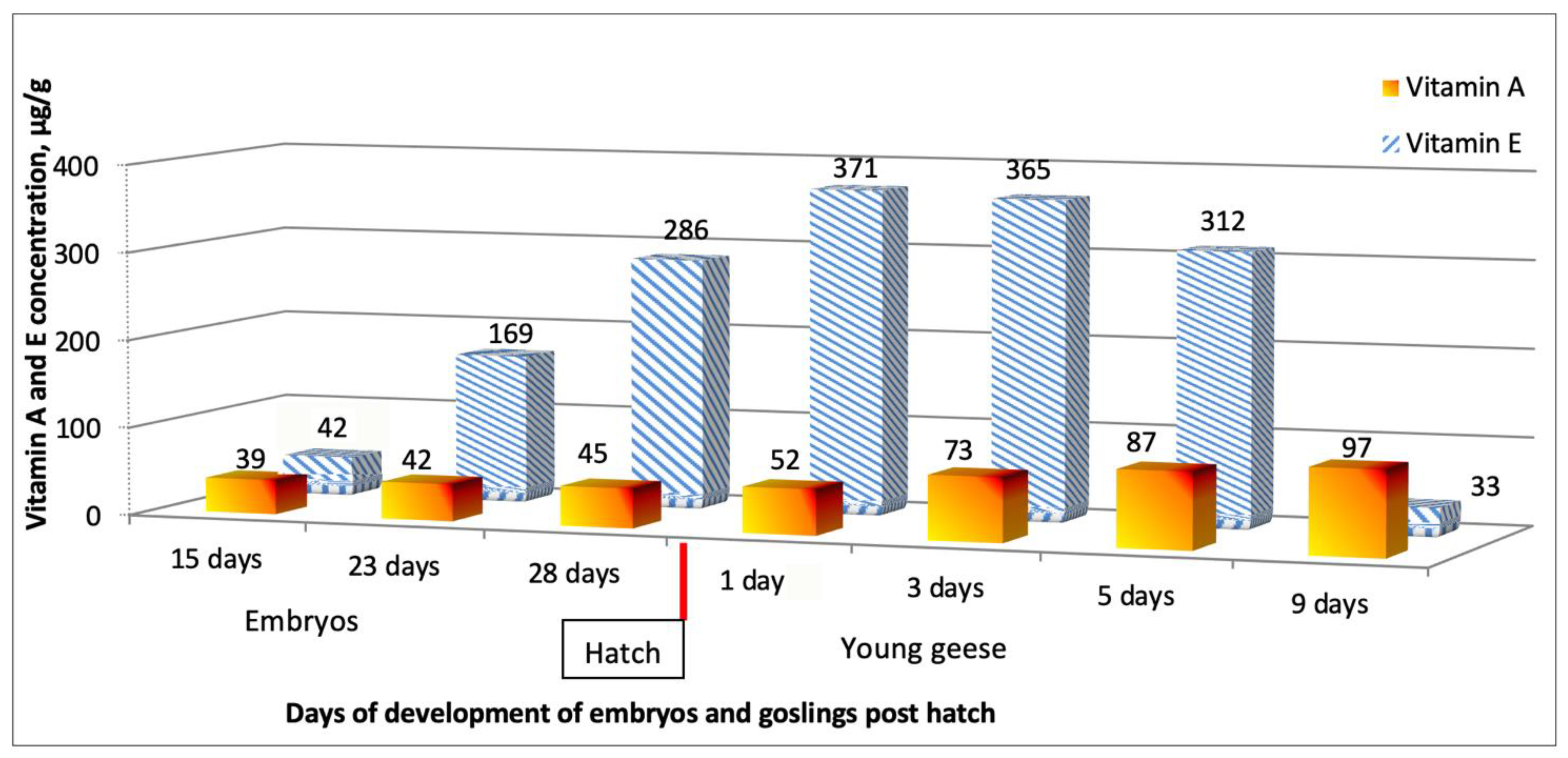
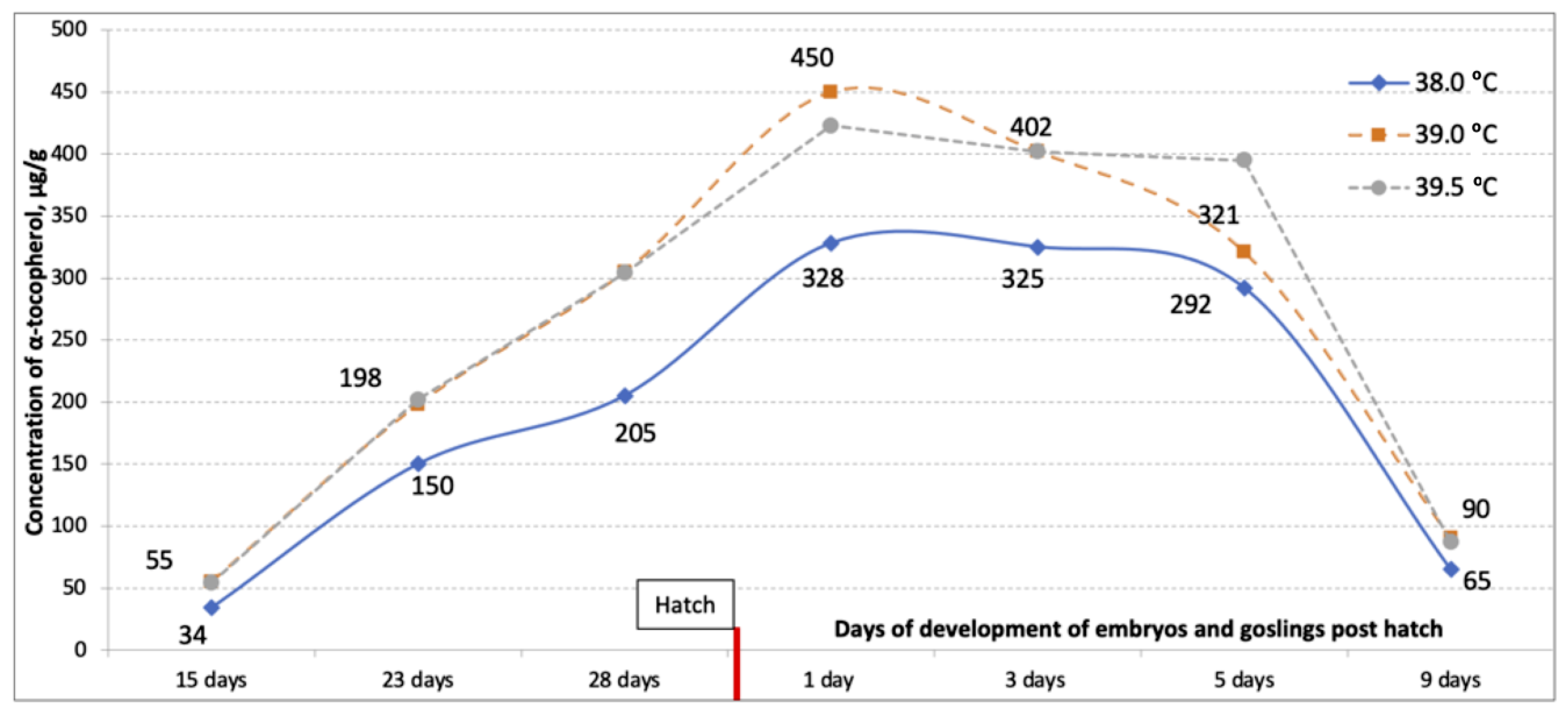
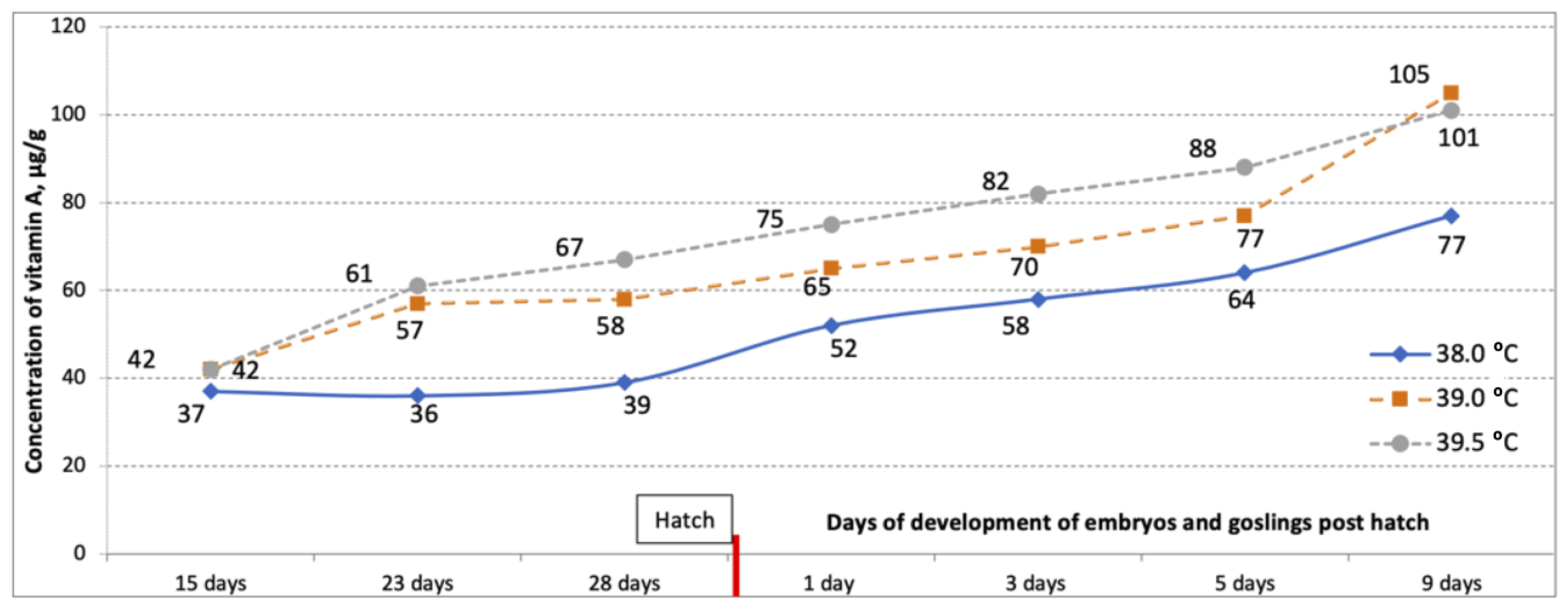
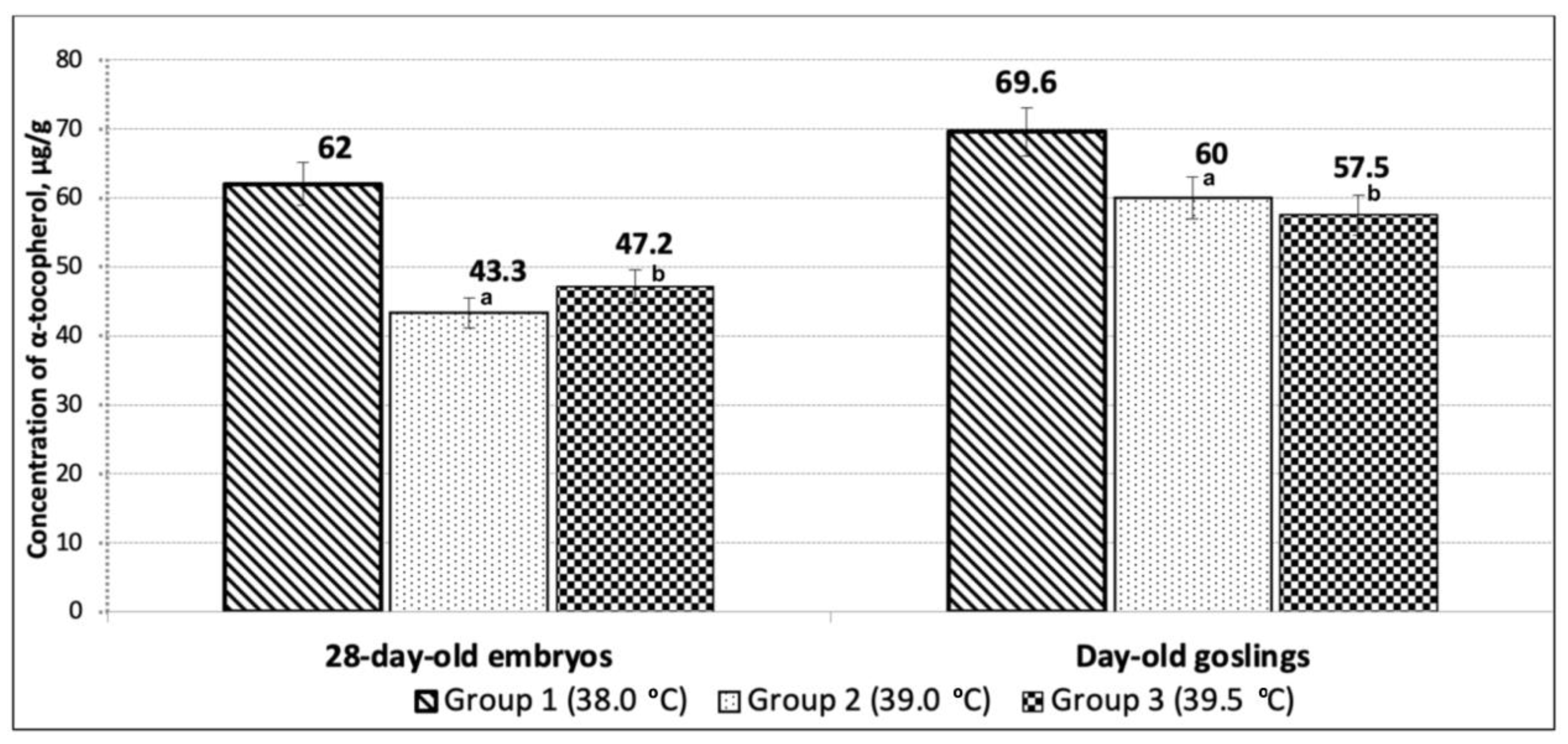
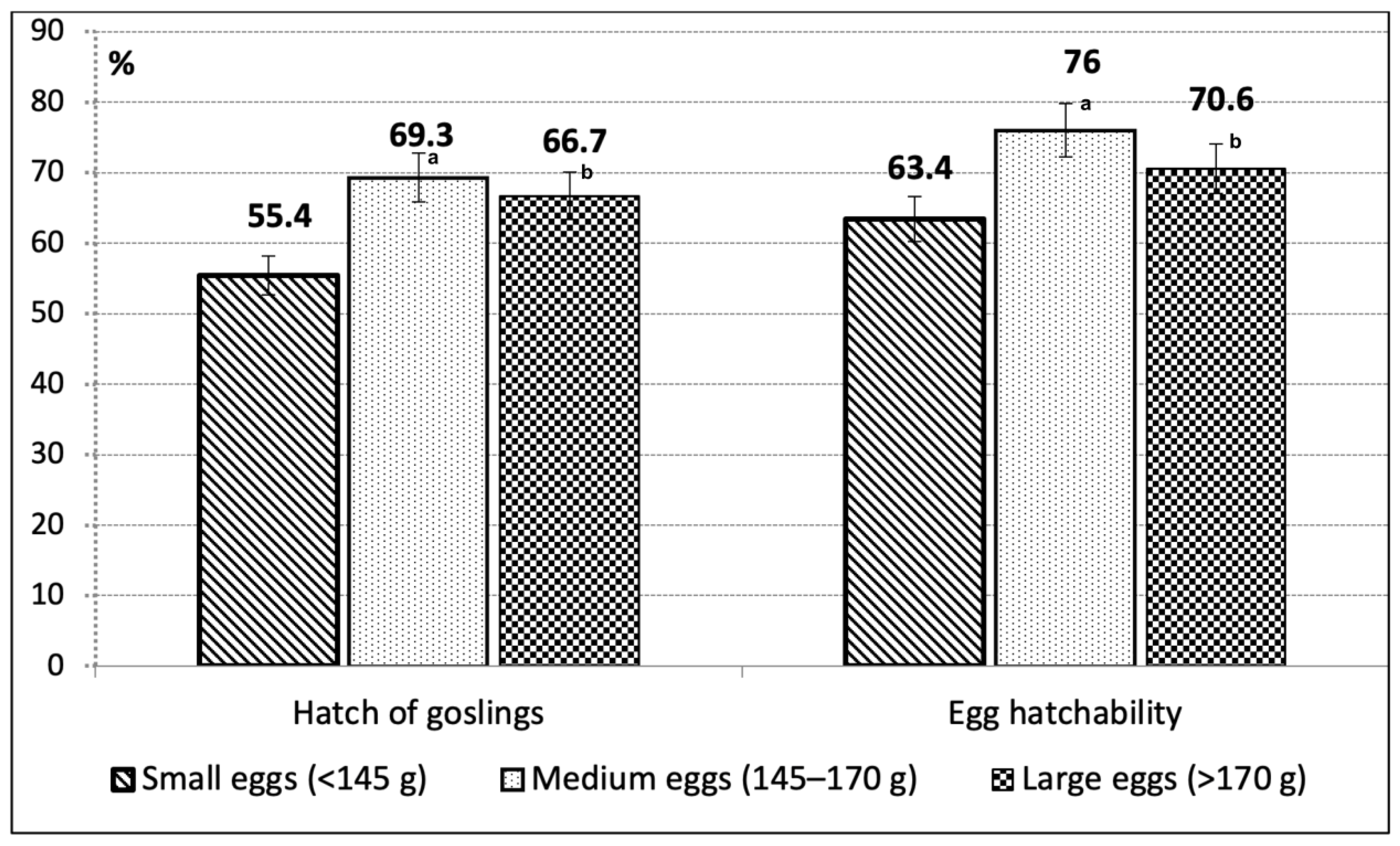
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Romanov, M.N. Evolution of domestic geese. Centres of domestication and dispersion. In New Investigations on Palearctic Geese; Gudina, A.N., Ed.; Zaporizhya Branch of the Ukrainian Ornithological Society, Zaporizhya State University: Zaporizhya, Ukraine, 1995; pp. 120–126. [Google Scholar]
- Romanov, M.N. Geese. In Encyclopedia of Animal Science, Two-Volume Set, 2nd ed.; Baer, C.K., Ullrey, D.E., Pond, W.G., Eds.; Taylor & Francis Inc.: London, UK; CRC Press Inc.: Boca Raton, FL, USA, 2018; Volume 1, pp. 487–489. [Google Scholar] [CrossRef]
- Tereshchenko, A.V.; Mikityuk, D.N.; Ryabokon, Y.A.; Ivko, I.I.; Melnik, V.A.; Ryabinina, E.V.; Khvostik, V.P.; Gorbanev, A.P. Geese on the Farm and in the Courtyard, 2nd ed.; Tereshchenko, A.V., Ed.; Poultry Research Institute, Ukrainian Academy of Agrarian Sciences: Borky, Ukraine, 2008. (In Russian) [Google Scholar]
- Bondarenko, Yu.V.; Zharkova, I.P.; Romanov, M.N. Study of down colour genotype in the collection flock geese at the All-Union Poultry Research and Technological Institute. Naučno-Teh. Bûll.—Ukr. Naučno-Issled. Inst. Pticevod. (Ukr. Poult. Res. Inst. Kharkiv Ukr.) 1986, 21, 3–7. (In Russian) [Google Scholar]
- Bondarenko, Y.V.; Tereshchenko, O.V.; Tkachyk, T.E.; Zakharchenko, O.P.; Hadiuchko, O.T. Recommendations on the Use of Modern Methods for Determining the Sex of Young Birds of Poultry (Theory and Practice); Kharkiv, Ukraine, 2007. (In Ukrainian) [Google Scholar]
- Sakhatsky, N.I.; Tereshchenko, A.V.; Artemenko, A.B. An express-method of estimation of fertilizing ability of freezing-thawing of poultry spermatozoa. Sel’skokhozyaistvennaya Biol. (Agric. Biol.) 1987, 22, 77–80. (In Russian) [Google Scholar]
- Romanov, M.N. Population-morphological Studies on Domestic Geese. In Proceedings of the Ukrainian Conference of Young Scientists and Postgraduates on Poultry Problems, Borky, Kharkiv, Ukraine; Poultry Research Institute, Ukrainian Academy of Agrarian Sciences: Kharkiv, Ukraine, 1992; pp. 8–9. (In Ukrainian) [Google Scholar]
- Romanov, M.N. Genetic Studies on Geese of Ukraine and Russia. 1. Historical Preface. In Proceedings of the 9th International Symposium on Waterfowl, Pisa, Italy, 16–18 September 1992; World’s Poultry Science Association: Pisa, Italy, 1992; pp. 102–104. [Google Scholar]
- Romanov, M.N.; Veremeyenko, R.P.; Bondarenko, Y.V. Conservation of Waterfowl Germplasm in Ukraine. In Proceedings of the 10th International Symposium on Waterfowl, Halle (Saale), Germany, 26–31 March 1995; World’s Poultry Science Association: Halle (Saale), Germany, 1995; pp. 401–414. [Google Scholar]
- Romanov, M.N.; Weigend, S.; Bondarenko, Y.V.; Podstreshny, A.P.; Kutnyuk, P.I.; Sakhatsky, N.I. Studies on Poultry Germplasm Diversity and Conservation in Ukraine. In Proceedings of the Poultry Genetics Symposium, Mariensee, Germany, 6–8 October 1999; Working Group 3 of WPSA: Mariensee, Germany, 1999; p. 140. [Google Scholar]
- Romanov, M.N.; Betuel, A.M.; Chemnick, L.G.; Ryder, O.A.; Kulibaba, R.O.; Tereshchenko, O.V.; Payne, W.S.; Delekta, P.C.; Dodgson, J.B.; Tuttle, E.M.; et al. Widely Applicable PCR Markers for Sex Identification in Birds. Russ. J. Genet. 2019, 55, 220–231. [Google Scholar] [CrossRef]
- Gadyuchko, O.T.; Sakhatsky, N.I.; Tereshchenko, A.V.; Anisimov, V.I.; Kislaya, E.D.; Svinarev, V.F. Genetic potential of breeds and populations of geese in Ukraine. Ptakhivnytstvo (Poult. Farming) 2003, 53, 54–62. (In Russian) [Google Scholar]
- Tagirov, M.T.; Tereshchenko, L.V.; Tereshchenko, A.V. Substantiation of the possibility of using primary germ cells as material for the preservation of poultry genetic resources. Ptakhivnytstvo (Poult. Farming) 2006, 58, 464–473. (In Russian) [Google Scholar]
- Tagirov, M.T.; Ogurtsova, N.S.; Tereshchenko, A.V. Analysis of hatchability problems for incubated eggs. Ptakhivnytstvo (Poult. Farming) 2009, 63, 199–215. (In Russian) [Google Scholar]
- Narushin, V.G.; Chausov, M.G.; Shevchenko, L.V.; Pylypenko, A.P.; Davydovych, V.A.; Romanov, M.N.; Griffin, D.K. Shell, a naturally engineered egg packaging: Estimated for strength by non-destructive testing for elastic deformation. Biosyst. Eng. 2021, 210, 235–246. [Google Scholar] [CrossRef]
- Narushin, V.G.; Romanov, M.N.; Griffin, D.K. Non-destructive evaluation of the volumes of egg shell and interior: Theoretical approach. J. Food Eng. 2021, 300, 110536. [Google Scholar] [CrossRef]
- Tagirov, M.T.; Shomina, N.V.; Artemenko, A.B.; Tkachenko, S.N.; Baydevlyatova, O.N.; Tereshchenko, A.B.; Sakhatsky, N.I. Incubation of Poultry Eggs: Guidelines; Poultry Research Institute, Ukrainian Academy of Agrarian Sciences: Kharkiv, Ukraine, 2009. (In Russian) [Google Scholar]
- Khvostyk, V.P.; Bondarenko, Y.V. Genetic polymorphism of egg white proteins of geese of different breeds of domestic selection. Ef. Ptakhivnytstvo (Eff. Poult. Farming) 2008, 12, 48. (In Ukrainian) [Google Scholar]
- Surai, P.F.; Ionov, I.A.; Bondarenko, Y.V. The content of carotenoids and vitamin a in the yolk of poultry eggs. Naučno-Teh. Bûll.—Inst. Pticevod. (Poult. Res. Inst. Ukr. Acad. Agrar. Sci. Kharkiv Ukr.) 1992, 33, 28–33. (In Russian) [Google Scholar]
- Bratyshko, N.I.; Haviley, O.V.; Tereshchenko, O.V.; Prytulenko, O.V. The effect of triticale on the vitamin content of eggs of chickens of different breeds. Ptakhivnytstvo (Poult. Farming) 2010, 65, 93–100. (In Ukrainian) [Google Scholar]
- Kokoszyński, D. Chapter 4—Guinea fowl, goose, turkey, ostrich, and emu eggs. In Egg Innovations and Strategies for Improvements; Hester, P.Y., Ed.; Academic Press: Cambridge, MA, USA; Elsevier Inc.: London, UK, 2017; pp. 33–43. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant defence systems and oxidative stress in poultry biology: An update. Antioxidants 2019, 8, 235. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Romanov, M.N.; Griffin, D.K. Nutritional modulation of the antioxidant capacities in poultry: The case of vitamin E. Poult. Sci. 2019, 98, 4030–4041. [Google Scholar] [CrossRef] [PubMed]
- Ionov, I.A. Physiological Status of the Birds in Embryogenesis and Postnatal Ontogenesis Depending on Its A-, E- and K-Vitamin Availability. Ph.D. Thesis, Poultry Research Institute, Ukrainian Academy of Agrarian Sciences, Kharkiv, Ukraine, 1997. (In Ukrainian). [Google Scholar]
- Surai, P.F.; Fisinin, V.I.; Karadas, F. Antioxidant systems in chick embryo development. Part 1. Vitamin E, carotenoids and selenium. Anim. Nutr. 2016, 2, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Yan, L.; Lei, M.; Dai, Z.; Shi, Z. Wider angle egg turning during incubation enhances yolk utilization and promotes goose embryo development. Animals 2021, 11, 2485. [Google Scholar] [CrossRef] [PubMed]
- Kucharska-Gaca, J.; Adamski, M.; Biesek, J. Effect of parent flock age on hatching, growth rate, and features of both sexes goose carcasses. Poult. Sci. 2022, 101, 101920. [Google Scholar] [CrossRef] [PubMed]
- Kucharska-Gaca, J.; Adamski, M.; Biesek, J. The age of the geese from the parent flock and the laying period affect the features of the eggs. Poult. Sci. 2022, 101, 102094. [Google Scholar] [CrossRef]
- Guo, B.B.; Dai, Z.C.; Ren, Y.H.; Zhu, H.X.; Shao, X.B.; Sun, A.D.; Shi, Z.D. Improvement of goose embryonic and muscular developments by wider angle egg turning during incubation and the regulatory mechanisms. Poult. Sci. 2021, 100, 101477. [Google Scholar] [CrossRef]
- Kochish, I.I.; Titov, V.Y.; Nikonov, I.N.; Brazhnik, E.A.; Vorobyov, N.I.; Korenyuga, M.V.; Myasnikova, O.V.; Dolgorukova, A.M.; Griffin, D.K.; Romanov, M.N. Unraveling signatures of chicken genetic diversity and divergent selection in breed-specific patterns of early myogenesis, nitric oxide metabolism and post-hatch growth. Front. Genet. 2023, 13, 1092242. [Google Scholar] [CrossRef]
- Kochish, I.I.; Brazhnik, E.A.; Vorobyov, N.I.; Nikonov, I.N.; Korenyuga, M.V.; Myasnikova, O.V.; Griffin, D.K.; Surai, P.F.; Romanov, M.N. Features of fractal conformity and bioconsolidation in the early myogenesis gene expression and their relationship to the genetic diversity of chicken breeds. Animals 2023, 13, 521. [Google Scholar] [CrossRef]
- Guo, B.; Dai, Z.; Chen, R.; Liu, J.; Shi, Z. Enhancing gosling growth and secretion of somatotrophic and thyrotrophic axis hormones through egg turning during incubation. Br. Poult. Sci. 2022, 64, 122–128. [Google Scholar] [CrossRef]
- Bondarenko, Yu.V.; Kovalenko, A.T.; Kutnyuk, P.I. Efficiency of modal selection in poultry populations. Naučno-Teh. Bûll.—Ukr. Naučno-Issled. Inst. Pticevod. (Ukr. Poult. Res. Inst. Kharkiv Ukr.) 1979, 7, 3–7. (In Russian) [Google Scholar]
- Bondarenko, Yu.V. The study of the comparative variability of the weight of eggs and day-old chicks of nine poultry species. Naučno-Teh. Bûll.—Ukr. Naučno-Issled. Inst. Pticevod. (Ukr. Poult. Res. Inst. Kharkiv Ukr.) 1989, 26, 6–10. (In Russian) [Google Scholar]
- Artemenko, O.B.; Tereshchenko, O.V.; Shomina, N.V.; Baidevlyatova, O.M.; Gavilei, O.V. Study of the Effect of Circadian Temperature Stimuli on the Results of Egg Incubation. In Proceedings of the Materials of the XIII International Conference “Poultry Industry 2017”, Truskavets, Ukraine, 19–21 September 2017; pp. 83–88. (In Ukrainian). [Google Scholar]
- Lomako, D.V. Technology of incubation of goose eggs in the conditions of Myrhorod private rental agricultural hatchery and poultry enterprise. Visnyk Poltavsʹkoyi Derzhavnoyi Ahrarnoyi Akad. (Bull. Poltava State Agrar. Acad.) 2015, 4, 44–46. (In Ukrainian) [Google Scholar] [CrossRef]
- Christensen, V.L.; Bagley, L.G. Improved hatchability of turkey eggs at high altitudes due to added oxygen and increased incubation temperature. Poult. Sci. 1988, 67, 956–960. [Google Scholar] [CrossRef] [PubMed]
- Łukaszewicz, E.; Lasoń, M.; Rosenberger, J.; Kowalczyk, A.; Bakst, M. Goose embryonic development from oviposition through 16 h of incubation. Poult. Sci. 2017, 96, 1934–1938. [Google Scholar] [CrossRef] [PubMed]
- Podstreshny, O.P.; Nalyvaiko, L.I.; Bondarenko, Y.V. Genetic polymorphism in populations of the Large Gray geese. Ptakhivnytstvo (Poult. Farming) 2001, 50, 19–26. (In Ukrainian) [Google Scholar]
- Mcmurray, C.H.; Blanchflower, W.J.; Rice, D.A. Influence of Extraction Techniques on Determination of α-Tocopherol in Animal Feedstuffs. J. AOAC Int. 1980, 63, 1258–1261. [Google Scholar] [CrossRef]
- Vahl, H.A.; Klooster, A.T. Effects of excessive vitamin A levels in broiler rations. J. Anim. Physiol. Anim. Nutr. 1987, 57, 204–218. [Google Scholar] [CrossRef]
- Bellairs, R.; Osmond, M. Appendix II—Normal Table of Hamburger and Hamilton (1951; 1992). In The Atlas of Chick Development, 3rd ed.; Academic Press: Oxford, UK; Waltham, MA, USA, 2014; pp. 603–621. [Google Scholar] [CrossRef]
- Li, S.; Bai, S.; Qin, X.; Zhang, J.; Irwin, D.M.; Zhang, S.; Wang, Z. Comparison of whole embryonic development in the duck (Anas platyrhynchos) and goose (Anser cygnoides) with the chicken (Gallus gallus). Poult. Sci. 2019, 98, 3278–3291. [Google Scholar] [CrossRef]
- Liang, J.; Dai, H.; Yang, H.; Yang, Z.; Wang, Z. The effect of dietary vitamin A supplementation in maternal and its offspring on the early growth performance, liver vitamin A content, and antioxidant index of goslings. Poult. Sci. 2019, 98, 6849–6856. [Google Scholar] [CrossRef]
- Surai, P.F.; Ionov, I.A.; Kuchmistova, E.F.; Noble, R.C.; Speake, B.K. The relationship between the levels of α-tocopherol and carotenoids in the maternal feed, yolk and neonatal tissues: Comparison between the chicken, turkey, duck and goose. J. Sci. Food Agric. 1998, 76, 593–598. [Google Scholar] [CrossRef]
- Surai, P.; Kuklenko, T.; Ionov, I.; Noble, R.; Sparks, N. Effect of vitamin A on the antioxidant system of the chick during early postnatal development. Br. Poult. Sci. 2000, 41, 454–458. [Google Scholar] [CrossRef]
- Surai, P.F.; Ionov, I.A.; Kuklenko, T.V.; Kostjuk, I.A.; Acpherson, A.M.; Speake, B.K.; Noble, R.C.; Sparks, N.H. Effect of supplementing the hen's diet with vitamin A on the accumulation of vitamins A and E, ascorbic acid and carotenoids in the egg yolk and in the embryonic liver. Br. Poult. Sci. 1998, 39, 257–263. [Google Scholar] [CrossRef]
- Shirakami, Y.; Lee, S.-A.; Clugston, R.D.; Blaner, W.S. Hepatic metabolism of retinoids and disease associations. Biochim. Biophys. Acta 2012, 1821, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Senoo, H.; Mezaki, Y.; Fujiwara, M. The stellate cell system (vitamin A-storing cell system). Anat. Sci. Int. 2017, 92, 387–455. [Google Scholar] [CrossRef]
- Bozhkov, A.; Ionov, I.; Kurhuzova, N.; Novikova, A.; Katerynych, O.O.; Akzhyhitov, R. Vitamin A intake forms resistance to hypervitaminosis A and affects the functional activity of the liver. Clin. Nutr. Open Sci. 2021, 41, 82–97. [Google Scholar] [CrossRef]
- Ivchenko, V.V.; Bondarenko, Y.V.; Rozhen, T.M. Method of Selecting Goose Eggs for Incubation. Patent SU 1535505 A1, 1990. Available online: https://patents.google.com/patent/SU1535505A1/en (accessed on 26 April 2023). (In Russian).
- Zhang, J.; Huang, B.; Cai, Z.; Xu, J.; Zheng, Y.; Wang, M. Fat-soluble vitamins in different eggs and egg products in Hangzhou City. Wei Sheng Yan Jiu 2020, 49, 984–989. (In Chinese) [Google Scholar] [CrossRef] [PubMed]
- Narushin, V.G.; Romanov, M.N. Influence of egg parameters on hatchability (a research review). Ptakhivnytstvo (Poult. Farming) 2001, 51, 422–429. (In Russian) [Google Scholar]
- Narushin, V.G.; Romanov, M.N. Physical Characteristics of Chicken Eggs in Relation to Their Hatchability and Chick Weight. In Proceedings of the ASAE Annual International Meeting/CIGR World Congress, Chicago, IL, USA, 28–31 July 2002; Paper #026066. American Society of Agricultural and Biological Engineers: St. Joseph, MI, USA, 2002. [Google Scholar] [CrossRef]
- Narushin, V.G.; Romanov, M.N. Relationship between physical characteristics and results of hatching in chicken eggs. Arch. Geflugelkd. 2002, 66 (Suppl. S2), 128. [Google Scholar]
- Narushin, V.G.; Bogatyr, V.P.; Romanov, M.N. Relationship between hatchability and non-destructive physical measurements of chicken eggs. J. Agric. Sci. 2016, 154, 359–365. [Google Scholar] [CrossRef]
- Prokudina, N.A.; Duyunova, A.A.; Ogurtsova, N.S.; Ryabokon, Y.A.; Artyomenko, A.B.; Ivko, I.I.; Melnik, V.A.; Bondarenko, Y.V. Turkey Egg Incubation Guide; Poultry Research Institute, Ukrainian Academy of Agrarian Sciences: Borki, Ukraine, 2007. (In Russian) [Google Scholar]
- Yang, H.; Liang, J.; Dai, H.; Wan, X.; Wang, Z. Effects of vitamin A supplementation in the diet of breeding geese on offspring intestinal tissue morphology and immune performance. Asian-Australas. J. Anim. Sci. 2020, 33, 1463–1469. [Google Scholar] [CrossRef]
- Surai, P.; Fisinin, V. Vitagenes in poultry production: Part 1. Technological and environmental stresses. World's Poult. Sci. J. 2016, 72, 721–734. [Google Scholar] [CrossRef]
- Surai, P.; Fisinin, V. Vitagenes in poultry production: Part 2. Nutritional and internal stresses. World's Poult. Sci. J. 2016, 72, 761–772. [Google Scholar] [CrossRef]
- Surai, P.F. Antioxidants in poultry nutrition and reproduction: An update. Antioxidants 2020, 9, 105. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F.; Earle-Payne, K. Antioxidant defences and redox homeostasis in animals. Antioxidants 2022, 11, 1012. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mode of Starting Heating | 28-Day Embryos | Day-old Goslings | ||
|---|---|---|---|---|
| Content, µg/g | Total, µg | Content, µg/g | Total, µg | |
| 38.0 °C (Group 1) | 53.8 ± 1.89 | 1688.0 ± 171.4 | 57.6 ± 4.45 | 590.8 ± 116.0 |
| 39.0 °C (Group 4) | 48.7 ± 1.52 | 1290.5 ± 61.7 * | 52.8 ± 2.90 | 288.7 ± 65.6 ** |
| 39.5 °C (Group 5) | 41.2 ± 3.00 * | 903.1 ± 111.7 ** | 45.3 ± 3.20 * | 309.6 ± 73.4 ** |
| Indicators | Groups | ||||
|---|---|---|---|---|---|
| 1 (38.0 °C) | 2 (39.0 °C) | 3 (39.5 °C) | 4 (39.0 °C) | 5 (39.5 °C) | |
| 24-h Starting Heating | 36-h Starting Heating | ||||
| No. of eggs incubated | 700 | 766 | 772 | 705 | 706 |
| Egg fertility, % | 94.3 | 94.6 | 94.2 | 90.8 | 92.1 |
| No. of goslings hatched | 504 | 571 | 576 | 520 | 557 |
| Hatch of young geese, % * | 65.4 | 74.5 | 74.6 | 73.7 | 78.9 |
| Egg hatchability, % ** | 69.4 | 78.7 | 79.2 | 81.3 | 85.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ionov, I.A.; Katerinich, O.O.; Kuchmistov, V.O.; Anisimova, O.V.; Griffin, D.K.; Romanov, M.N.; Zhukova, I.O. Vitamin E and A Availability in Goose Embryos and Goslings and Improvement of Reproduction Traits Depending on the Starting Temperature Regime of Egg Incubation. Poultry 2023, 2, 305-319. https://doi.org/10.3390/poultry2020023
Ionov IA, Katerinich OO, Kuchmistov VO, Anisimova OV, Griffin DK, Romanov MN, Zhukova IO. Vitamin E and A Availability in Goose Embryos and Goslings and Improvement of Reproduction Traits Depending on the Starting Temperature Regime of Egg Incubation. Poultry. 2023; 2(2):305-319. https://doi.org/10.3390/poultry2020023
Chicago/Turabian StyleIonov, Igor A., Oleg O. Katerinich, Viktor O. Kuchmistov, Olga V. Anisimova, Darren K. Griffin, Michael N. Romanov, and Irina O. Zhukova. 2023. "Vitamin E and A Availability in Goose Embryos and Goslings and Improvement of Reproduction Traits Depending on the Starting Temperature Regime of Egg Incubation" Poultry 2, no. 2: 305-319. https://doi.org/10.3390/poultry2020023