
Diagnosing Infectious Diseases in Poultry Requires a Holistic Approach: A Review

Clinic for Poultry and Fish Medicine, University of Veterinary Medicine Vienna, Veterinaerplatz 1, 1210 Vienna, Austria
*
Author to whom correspondence should be addressed.
Poultry 2023, 2(2), 252-280; https://doi.org/10.3390/poultry2020020
Submission received: 8 March 2023
/
Revised: 17 April 2023
/
Accepted: 20 April 2023
/
Published: 25 April 2023
(This article belongs to the Special Issue Poultry Infectious Diseases)
Abstract
:Controlling infectious diseases is vital for poultry health and diagnostic methods are an indispensable feature to resolve disease etiologies and the impact of infectious agents on the host. Although the basic principles of disease diagnostics have not changed, the spectrum of poultry diseases constantly expanded, with the identification of new pathogens and improved knowledge on epidemiology and disease pathogenesis. In parallel, new technologies have been devised to identify and characterize infectious agents, but classical methods remain crucial, especially the isolation of pathogens and their further characterization in functional assays and studies. This review aims to highlight certain aspects of diagnosing infectious poultry pathogens, from the farm via the diagnostic laboratory and back, in order to close the circle. By this, the current knowledge will be summarized and future developments will be discussed in the context of applied state-of-the-art techniques. Overall, a common challenge is the increasing demand for infrastructure, skills and expertise. Divided into separate chapters, reflecting different disciplines, daily work implies the need to closely link technologies and human expertise in order to improve bird health, the production economy and to implement future intervention strategies for disease prevention.
Keywords:
chicken; flock health; infectious diseases; virology; molecular biology; bacteriology; parasitology; serology1. Introduction
Infectious diseases pose a constant risk on poultry health and production, with substantial consequences on welfare and economy. This is addressed in various book chapters focusing on infectious diseases, together with numerous review articles. Likewise, dedicated books and research articles suggest specific diagnostic techniques to diagnose infectious diseases in poultry.
The first successful example combining pathogen detection with initial characterization followed by the implementation of a screening program was the control of Pullorum disease. The concentrated action of different stakeholders paved the way for the National Poultry Improvement Plan (NPIP) in the USA and is a role model for successful cooperation between scientists, government and industry as reviewed in detail by Schat et al. [1]. Whereas some countries successfully eradicated the disease, others still rely on vaccination, which was developed in the mid of last century [2]. Interrupting vertical transmission as a main route of pathogen spread was also successful for eradication of avian leukosis J virus from primary broiler breeder flocks in some countries [3], again highlighting the close interaction of stakeholders. However, eradication is much less successful on a broad scale and poultry health is confronted with old and new diseases, sometimes summarized as (re-)emergent, today. As farms and flocks become bigger, economic losses of any disease outbreak are constantly increasing, although detailed figures, especially for endemic diseases, are very often not available [4]. Furthermore, in some areas, ethical concerns about husbandry (cage vs. free range), management practices or breed of birds (e.g., fast- vs. slow-growing broilers) influence the epidemiology of certain pathogens and/or disease prevalence. Finally, despite the fact that backyard poultry is very often not covered by legislation which is applied to commercial poultry, diagnostic procedures should be performed to clarify disease etiology considering the importance of birds raised under loose biosecurity for pathogen spread [5,6].
Overall, in most cases, diagnosing infectious diseases follows an established procedure, starting at the farm with the collection of data on disease history. This is frequently combined with initial post mortems. Further investigations require a laboratory environment to proceed with different techniques and disciplines. This review aims to deliver an overview of standard procedures applied to diagnose infectious poultry diseases and to highlight new developments keeping in mind that each chapter requires a review on its own.
2. Diagnostic Activities on Farm
In poultry medicine, the diagnostic process originally shifted from the traditional veterinarian approach centered on individual animals to the health assessment of entire flocks. Flocks are commonly classified as “healthy” if they perform according to their genetic potential and are considered free from clinical disease. On-farm, diagnostic activities comprise routine sampling and investigations in line with health control programs; nationally and/or internationally adopted control programs for certain Mycoplasma and Salmonella species represent examples of paramount importance [7,8]. Samples may be investigated immediately on site (e.g., rapid antigen test for avian influenza) or sent for further processing to a laboratory (e.g., ELISA and PCR). Field veterinarians further implement diagnostic surveillance in order to provide epidemiological data for flock management purposes. The periodical collection of samples (e.g., feces, serum samples, and swabs from mucosal surfaces) is primarily used to confirm the infection (free) status of a flock or to monitor vaccine response. Altogether, generated data facilitate objective judgment and decision making in order to optimize flock health and production.
In the field, diagnostic procedures are initiated as soon as flock health is compromised, using morbidity and/or mortality as initial indicators. In such a scenario, investigations start with the compilation of a case history pertaining to relevant flock, management and infection/disease characteristics (e.g., bird type and origin, age, routine medications, vaccination program, previous diseases, husbandry system, standard operation procedures such as feeding and watering systems, ventilation, lighting program, hygiene and biosecurity processes, production parameters, morbidity and mortality data, duration of signs/problems and epidemiological links to other production sites). On the farm, diagnostics starts with the clinical examination of flocks, individual birds in various stages of the disease and their products (e.g., feces and eggs) by experienced poultry workers and veterinarians thoroughly familiar with the appearance of a healthy flock and the environment. Clinical examinations are time consuming and labor intensive and can regrettably fail to detect diseases; especially subclinical diseases can be challenging to be accurately diagnosed. The manifestation of an infectious disease can vary from subclinical to severe clinical illness, depending on various etiological factors and influences such as the causative agent, host and/or environment altogether complicating diagnosis [9]. Clinical signs comprise non-specific, general signs (e.g., apathy, ruffled feathers, and inappetence), which can be associated with a wide range of diseases, often together with more specific signs indicative of a certain disorder (e.g., enteric, respiratory, and neurologic) or even pathognomonic for a specific disease (e.g., histomonosis in turkeys) [10]. Diagnostic procedures continue with post mortem investigations, on farm or in the laboratory, which serve to identify gross pathologic changes in organs and tissues in order to further specify a tentative cause of impaired performance and clinical signs. For instance, high mortality in a turkey flock coinciding with sulfur-colored droppings paired with necropsy results of typhlitis and hepatitis indicates an infection with Histomonas meleagridis; a presumptive diagnose, which might be confirmed by histology or even molecular diagnostics [11]. In the absence of mortality, euthanasia of sick birds might become a necessity to gain samples for further investigations. Altogether, a comprehensive case history together with the accurate assessment of clinical signs and thorough post mortem investigations narrows the range of presumptive diagnoses. This provides the basis to select appropriate laboratory methods as highlighted below in the outlook.
At the beginning, many infectious poultry diseases can be characterized by shortfalls in performance [12]. Those might be present long before clinical signs occur or even if clinical signs remain absent at all in infected flocks [4]. Consequently, production parameters (e.g., weight gain, feed-conversion rate, and egg production) of flocks are assessed and compared with established breed and/or company standards to detect anomalies. Very often production parameters are only available with certain delay (e.g., hatchability, chick quality or slaughter results) and can only be analyzed retrospectively with the purpose to enhance understanding of a diseases’ impact and the implementation of corrective and preventive actions. Thus, financial losses due to subclinical disease outbreaks of necrotic enteritis have been calculated [13] and production losses as a result of emerging diseases have been detailed [14,15]. Similarly, the outcome and effectiveness of treatment plans have been evaluated analyzing historical flock data (e.g., coccidiosis) [16]. For diagnostic purposes, prompt evaluation of production efficiency and quality is essential. Modern poultry farming already uses precision livestock farming (PLF) technologies such as environmental sensors or platform scales to measure, predict and analyze various factors related to production in real time directly from within poultry houses [17]. So far, this is used mainly to optimize systems for feeding and drinking or to regulate environmental settings (heating and ventilation). Many characteristics of poultry production (e.g., closed housing system and large-scale integrations) provide a good opportunity to adopt PLF strategies to assess bird health and for early disease detection. Recently, modern technologic advancements utilizing video surveillance, audio recordings and/or wearable sensors have focused on behavior and behavioral changes in individual birds and entire flocks [18,19]. Thereby, bird activity, posture and/or vocalizations have been investigated in order to automate the detection of (early) infections and to identify disease outbreaks such as avian influenza or Newcastle Disease (Table 1). Likewise, fecal changes indicative of coccidiosis or salmonellosis have been assessed by imaging technologies to facilitate AI-based diagnostic services [20].
It seems obvious that sensor- or computer-based, automated systems will be part of the future of disease diagnostics, both in the field and in laboratories. However, the commonly rural location of farms might challenge field application of new diagnostic technologies and devices, which generally require a stable power supply and internet connection to function and provide data in real time [21]. Prospectively, as farm-generated data gains importance in poultry production and management, data security must be considered carefully to maintain company safety and integrity. Finally, the use of automated monitoring systems in intensive poultry production with a possible effect on the human–animal relationship may raise ethical concerns related to the objectification of animals [22].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Precision livestock farming research focusing on infectious agents and related poultry diseases.
Table 1.
Precision livestock farming research focusing on infectious agents and related poultry diseases.
| Disease/Pathogen | Method (Variables Measured) | Type of Study * | Reference |
|---|---|---|---|
| Avian influenza | wearable sensor (body temperature) | experimental setting | [23] |
| Avian influenza | wearable sensor (activity, body temperature) | experimental setting | [24] |
| Avian influenza + Infectious bronchitis + Newcastle disease | sound analysis (vocalizations) | experimental setting | [25] |
| Avian influenza | imaging (posture) | experimental setting | [26] |
| Avian influenza | sound analysis (vocalizations) | experimental setting | [27] |
| Avian influenza | sound analysis (vocalizations) | experimental setting | [28] |
| Avian influenza | Imaging (thermal images) | experimental setting | [29] |
| Campylobacter jejuni | imaging (flock movement—optical flow) | dataset from broiler buildings | [30] |
| Clostridium perfringens | sound analysis (vocalizations) | experimental setting | [31] |
| Coccidiosis | sensor (volatile organic compounds) | experimental setting + broiler building | [32] |
| Coccidiosis | sensor (volatile organic compounds) | dataset from broiler buildings | [33] |
| Coccidiosis + Salmonella spp. | imaging (feces) | dataset of images | [20] |
| Ektoparasites | wearable sensor (activity) | dataset from poultry building | [34] |
| Infectious bronchitis | sound analysis (rales) | experimental setting | [35] |
| Infectious bronchitis | sound analysis (rales) | experimental setting | [36] |
| Infectious bronchitis + Newcastle disease | sound analysis (vocalizations) | experimental setting | [37] |
| Newcastle disease | sound analysis (sneezes) | experimental setting | [38] |
| Newcastle disease | imaging (posture and mobility) | experimental setting | [39] |
| Newcastle disease | sound analysis (vocalizations) | experimental setting | [40] |
| Non-specific, clinical signs | imaging (feces) | dataset of images | [41] |
| Non-specific, clinical signs | imaging and sound analysis | dataset of audio samples | [42] |
| Non-specific, clinical signs | imaging (feces) | dataset from broiler building | [43] |
| Non-specific, clinical signs | imaging (head motion, appearance) | experimental setting | [44] |
| Non-specific, clinical signs | imaging (posture, appearance) | experimental setting | [45] |
| Non-specific, clinical signs | sound analysis (abnormal respiratory sounds) | dataset from broiler building | [46] |
| Non-specific, clinical signs | imaging (posture, appearance) | dataset of images | [47] |
| Pasteurella spp. | imaging (thermal images) | experimental setting | [48] |
* All studies were perfomed with chickens, except Noh et al. [29] who also used ducks.
3. Bacteriology
In recent years, the extension of diagnostic tools in clinical microbiology laboratories had a major impact on bacteriological investigations. In general, a certain trend to introduce molecular techniques and proteomics that complement or substitute classical bacteriology can be observed [49]. The following chapter intends to highlight the value of classical bacteriology but also the contribution of new techniques to broaden the knowledge on relevant bacteria in poultry medicine.
Very often, the clinical picture and post mortem findings cannot be attributed to a specific bacteria or fungi. Therefore, one of the main goals of bacteriological and fungal examination in diagnostics is the isolation of the pathogen. In addition to disease diagnosis, culturing of bacteria is also obligatory for some surveillance programs, such as that of Salmonella, which are based on the isolation of live bacteria [50,51].
For culturing, material can be taken directly from affected organs or the environment. Either the streak plate procedure with subsequent dilutions to gain single colonies is applied or material will firstly be transferred into broth cultures and afterwards streaked out on solid media [52]. The culture media widely used in diagnostic laboratories have to be supplemented with diverse nutrients and components to enable or facilitate the growth of particular bacteria/fungi. To isolate fastidious bacteria blood agar containing different concentrations of sheep/horse blood is used which is also suited to assess the hemolytic ability [53]. MacConkey agar supplemented with bile salts and crystal violet inhibiting the growth of most Gram-positive bacteria is a well-established media for the isolation of Gram-negative Enterobacterales [54]. More specific media, such as modified semisolid Rappaport Vassiliadis agar (MSRV) and modified charcoal-cefoperazone-deoxycholate agar (mCCDA) containing specific components and antibiotics, are used to isolate Salmonella species and thermophilic Campylobacter, respectively [51,55]. For cultivation of fungi and yeasts different variations of Sabouraud-glucose agars are mainly used [56]. In general, bacteria and fungi of interest have their growth optimum between 37.0 °C and 41.5 °C [52]. Some bacterial pathogens have special atmosphere requirements. For example, strict microaerobic conditions are needed for the cultivation of Avibacterium paragallinarum [57]. In contrary, most Brachyspira and Clostridium species will strictly grow under anaerobic conditions only [58,59]. Some bacteria are difficult to cultivate, relying on complex media and longer incubation time to achieve sufficient growth, e.g., Mycoplasma species, Avibacterium paragallinarum, Campylobacter hepaticus and Mycobacterium avium [57,60,61,62]. In addition to suitable media and cultivation conditions, classical bacteriological cultivation is also limited by the appearance of viable but non-culturable (VBNC) bacteria which are known to develop under unfavorable environmental conditions reported for Campylobacter jejuni and Escherichia coli [63,64].
The classical cultivation techniques do not only provide information if pure or mixed bacterial cultures are present, it also enables further typing, the application of antimicrobial susceptibility tests, production of autogenous vaccines and execution of functional studies. Traditional typing methods comprise the determination of colony morphology, hemolysis, staining and phenotypic identification tests. Based on colony morphology (size, shape, color, margin, surface) unusual growth variants of bacterial species can be detected as previously shown for Avibacterium paragallinarum and Ornithobacterium rhinotracheale [65,66]. Hemolysis is used to group bacteria into non-hemolytic such as Pasteurella multocida, partial-hemolytic (α-hemolysis) such as Erysipelothrix rhusiopathiae and complete-hemolytic (ß-hemolysis) such as Staphylococcus aureus. Most often used staining method is the Gram-stain differentiating bacteria by chemical and physical properties of their cell walls into Gram-positive and Gram-negative. The Ziehl-Neelsen stain is used to identify acid-fast organisms such as Mycobacterium avium, and the lactophenol-blue stain is applied to illustrate fungi and yeasts. Phenotypic identification tests comprise diverse biochemical reactions for which commercial systems are available, e.g., analytical profile index (API) systems [67]. Such tests can also be applied to discriminate subspecies. In Pasteurella multocida, the classification of the three subspecies, subsp. multocida, septica, or gallicida, is based on the ability to ferment sorbitol and dulcitol [68]. However, these biochemical methods are time consuming and resource intensive. Furthermore, variable characteristics among members of the same species or the lack to ferment most of the carbohydrates results in a heterogeneous or insufficient outcome of testing as shown for Gallibacterium anatis or Riemerella anatipestifer [69,70]. Worth to mention that also commercially available phenotypic databases sometimes lack updated bacterial classification data hindering proper and reliable identification as shown for Ornithobacterium rhinotracheale [71]. Furthermore, serotyping schemes are widely used based on antibodies raised against certain bacterial components such as capsules, somatic structures or flagella. Autoagglutination, cross-reactions and non-typeable bacterial strains are regularly found, altogether limiting the approach [71,72,73,74].
Live bacteria are also needed to determine antibiotic resistance profiles in order to implement a targeted treatment and to collect data for long-term assessment of antibiotic resistance [75,76,77,78,79]. Most common in routine diagnostics is the application of the disc diffusion method [80], which is still the golden standard to test antimicrobial susceptibility. The use of the broth microdilution method, for which commercially available semi-automated systems are available, is also popular [81]. The evaluation of the outcomes from such testing is based on standard documents, e.g., EUCAST [82] and CLSI [83].
Due to limitations in cultivating and typing, molecular techniques and proteomics has become more popular in recent years in routine bacteriological diagnostics [84,85]. For example, different PCR assays are widely used for the detection of Mycoplasma gallisepticum and Mycoplasma synoviae [86], Chlamydia spp. [87], Gallibacterium anatis [88], Avibacterium paragallinarum [89] or Salmonella spp. [90]. Specific PCR assays are also applied for species identification and further typing of isolates, e.g., defining virulence-associated genes in Escherichia coli or the Type C neurotoxin (BoNTC) gene in Clostridium botulinum [91,92]. In addition to, sequencing based on 16S and 23S ribosomal RNA (rRNA) regions as well as next-generation sequencing (NGS)/whole-genome sequencing (WGS) are increasingly applied. Sequence analysis of rRNA has been used for example for species identification of Campylobacter jejuni and Campylobacter coli [93], or Avibacterium paragallinarum [94]. NGS/WGS proved valuable in gaining more data regarding characterization of Morganella morganii [95], Ornithobacterium rhinotracheale [96], E. coli [97] and Salmonella Infantis [98]. These methods can also be applied for direct identification of bacterial communities as shown for samples from poultry carcasses in slaughterhouses [99]. A new area of application for such culture-independent assays is the screening of gut microbiota, but also environmental samples for antibiotic resistance genes to obtain a better understanding of the antibiotic resistome [100,101,102,103,104].
In addition to genetic methods, proteomics found its way into routine diagnostics mainly due to the implementation of matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS), which is based on ribosomal protein profiles obtained from bacterial isolates [84,105]. The availability of benchtop instruments makes it easier to develop in-house databases to be used in addition to commercially available ones. With this, a broad range of bacteria relevant in poultry medicine can successfully be identified to the species level, as shown for Gallibacterium [106], Riemerella [107], Mycoplasma [108], and Aspergillus [109,110], whereas limitations for others such as Avibacterium were reported [111]. Investigations beyond the species level were able to define clonal lineages for Gallibacterium anatis [112] or to discriminate environmental source specific isolates of Escherichia coli [113]. MALDI-TOF MS also has several applications to detect specific feature of antibiotic resistance, e.g., enzymatic activity (carbapenemases) or direct analysis of bacterial extracts (vancomycin-resistant enterococci) [114], with the advantage that results are available within a few hours.
Both molecular methods and proteomics are suited to determine phylogenetic relationships of isolates [112,115,116]. This feature proved of value in surveillance programs as shown for Campylobacter and Salmonella spp. [117,118]. It also highlights the importance of cultivating bacteria, which, in our opinion, remains irreplaceable despite modern technologies. It is crucial to perform animal trials and to investigate host–pathogen interactions. Live and well-defined bacteria also provide the basis for the production of autogenous vaccines, an important aspect of disease prevention [119,120,121,122].
However, with the help of more advanced technologies such as PCR and MALDI-TOF MS, the direct detection of pathogens, especially those which are difficult to cultivate, as well as routine identification of bacteria and fungi have become more rapid and easier during the last decade. Furthermore, NGS is currently transitioning from research to diagnostics, becoming more common in diagnostic laboratories. In regard to antimicrobial susceptibility testing, phenotypic methods will still remain crucial and its replacement by other techniques in the near future will not be the case. Finally, serological tests implemented in the field will still remain valuable tools for official monitoring programs as outlined below in Section 7.
4. Parasitology
Diagnoses of poultry parasites depends particularly on macroscopical and microscopical examination [123]. Ectoparasites include insects and arachnids that can be observed during clinical examination and necropsy on the feathers or the skin of infected poultry. Changes in the integument and its structures display a first suspicion that can be further specified by careful examination with the naked eye [124]. However, to determine the level of infestation, traps positioned in various areas of the poultry house are needed [125]. Internal parasites of poultry can colonize different organs or the blood [126,127].
The growing trend of raising laying hens in alternative housing systems with less biosecurity favors the prevalence of parasites [128,129]. Hence, Jung et al. [130] showed that in such systems, the majority of flocks excreted more than 200 ascarid eggs per gram feces. Accurate necropsy can reveal the presence of larvae and adult stages of helminths; however, eggs and protozoan parasites have to be identified by microscopic examination. For intestinal parasites, scrapings of the mucosa, intestinal content or feces are used and assessed for the presence of eggs. The identification of parasites and/or eggs can be achieved considering their morphological characteristics and depends on the applied technique and the experience of the investigator [131]. The applied diagnostic method is of high importance. The flotation method in combination with post mortem investigations is recommended for the detection of intestinal helminths to reduce false negatives [132]. Recently, fecal egg count techniques have been evaluated and it was demonstrated that McMaster is more accurate but less precise than mini-FLOTAC, underlining the need to evaluate diagnostic techniques [133]. Flotation and enumeration can also be applied for intestinal protozoans such as Eimeria spp. [134]. Other intestinal protozoans, such as flagellates, can be diagnosed by microscopic detection from cecal content or following isolation in culture medium but this requires fresh sample material [10]. Alternatively, molecular detection tools such as PCR, immunohistochemistry and in situ hybridization, as comprehensively described in the respective chapter of this review, have been developed for a sensitive and specific diagnosis of parasitic diseases of poultry.
In addition to conventional techniques, PCR can be applied to increase sensitivity and specificity to detect parasites with the advantage of combing it with typing as shown for Eimeria spp. [135,136]. In addition to tissue samples, environmental samples might be screened by PCR for parasite DNA even though clinical relevance seems less clear [137,138,139]. Furthermore, indirect detection by serology can be applied, which is helpful to generate results from high numbers of birds. Experimental ELISA systems were developed to determine antibodies against Ascaridia galli and Histomonas meleagridis [140,141,142]. Although there are a few research studies demonstrating the value of the assays, none of them are commercially available, limiting their use as monitoring tools.
5. Virology
Poultry is the host of numerous viruses, with some of them infecting chickens, turkeys, waterfowl, spreading to wild birds and even humans. The most important examples are avian influenza viruses with a broad host range and substantial variation in disease pathogenesis depending on host–virus interaction. Diagnosis of viral infections is a paradigm of a multifaceted approach as PCR has become a method of choice enabling fast detection of the nucleic acid (see below Section 6). However, to resolve functional aspects of an infection or to gain antigen for vaccine production, very often isolation is needed. However, multiplication of viruses relies on a certain substrate but the lack of broadly susceptible cell lines is very obvious in a poultry diagnostic laboratory. In general, initial isolation of a virus is somehow different to adaptation and propagation of a pathogen already available. In this context, poultry virology is still dominated by embryonated specific pathogen-free (SPF) chicken eggs and cells obtained from those embryos or hatched SPF birds. In a diagnostic laboratory, SPF eggs are very often at hand and the broad range of susceptibility is a substantial surplus, albeit additional work is needed to characterize and type the isolate which might be considered as a second step.
The first epithelial cell line from fowl was established from the liver of carcinogen treated chickens [143]. In one of the first studies, this hepatocellular carcinoma cell line (LMH) was capable of supporting multiplication of infectious laryngotracheitis virus (ILTV) although initial isolation was not reported [144]. Chicken astrovirus (CAstV) was successfully isolated on LMH cells, inducing a marked CPE after 3–5 passages [145,146]. In an earlier study, the susceptibility of the cell line to multiplicate Avian Nephritis Virus (ANV) and duck astroviruses (DAstV) was demonstrated as well. Isolation of DAstV-1 was described later on, again with CPE at the 5th passage [147,148]. The same number of passages was reported to induce visible changes in LMH cells during isolation of goose astrovirus [149]. With the recent appearance of virulent fowl adenovirus (FAdV) serotype 4 in China, LMH cells have become attractive to substitute SPF embryos for virus isolation [150]. However, in a very comprehensive epidemiological study, out of 2210 PCR positive samples, only 877 FAdV isolates were obtained despite 6 blind passages on LMH cells [151]. Although the difference might be explained by the absence of live virus, it could also well be that some serotypes/strains are more easily isolated and/or LMH cells might be a less suitable substrate compared with primary cells and/or SPF embryos. Except the initial study about the growth of FAdV-9 on LMH cells, no additional serotypes are investigated in detail, although cells are widely used in recent studies on virulent FAdV-4 in China [152,153]. Isolation of avian reoviruses from chickens, turkeys, pheasants and guinea fowl was very successful on LMH cells with CPE already detected after 24 h [154]. Similarly, a recombinant ARV isolate from geese with high sequence homology to chicken isolates could be isolated using LMH cells [155]. A single report described the isolation of avian Hepatitis E Virus (aHEV) on LMH cells which contradicts an earlier report that cell to cell spread of the virus is impossible following transfection of LMH cells with an infectious aHEV clone [156,157].
Several cell lines were established from chemically induced fibrosarcoma of Japanese quails [158]. On one of those, QT-35 was able to induce a cytopathogenic effect by testing a broad range of avian viruses, although variations among virus strains were noticed [159]. Strain variations have become obvious as none of the tested adenoviruses induced a CPE but later on the attenuation of virulent FAdV-4 on QT-35 cells was reported [160]. QT-35 cells also support initial replication of ILTV but continuous passaging was not possible [144,159]. These quail cells also support the induction of a CPE by reticuloendotheliosis virus (REV) [161]. Being equipped with a high number of α2,3-gal receptors, avian influenza viruses from different birds grew best on QT-6 cells, compared with DF-1 and MDCK cells [162].
Contrary to those reports on multiplication of already isolated viruses, QT-35 cells were used to isolate an orthoreovirus from tendons of turkeys suffering from arthritis after several passages [163]. Similarly, avian metapneumovirus subtype C, either from commercial turkeys or wild Canada goose, was successfully isolated on QT-35 cells. Success for other aMPVs was not reported although multiplication was demonstrated [164,165,166,167].
A cell line with susceptibility for different viruses is the chick embryo related (CER) cell line, a mixture of chicken embryo fibroblasts and baby hamster kidney (BHK21) cells [168]. It was used to isolate avian metapneumovirus (aMPV) from turkeys and it was later shown to support growth of infectious bursal disease virus (IBDV) and infectious bronchitis virus [169,170,171].
The DF-1 cell line is a fibroblast cell line with the special feature of carrying no endogenous avian sarcoma and leucosis virus [172]. With this feature, the cell line is well suited for isolating avian leucosis viruses [173]. An infectious bronchitis virus that causes kidney lesions and respiratory signs can induce a CPE in DF-1 cells, but without being passaged that is different to the observation in QT-35 cells, in which passaging could be achieved albeit with low titers [159,174]. Similarly, higher titers were obtained with primary chicken embryo fibroblasts as compared to DF-1 cells, even though both cell culture systems multiplied IBDV and NDV [175].
Frequent studies describe the isolation of viruses infecting poultry on mammalian cell lines, especially kidney cells. Following initial isolation in tracheal organ culture and adaption of aMPV on Vero cells such cells were later also used for primary virus isolation [164,176,177,178,179]. Vero cells are also described to be susceptible for IB, following primary isolation in SPF embryos [180]. First successful isolation of avian group A rotaviruses in Rhesus monkey kidney (MA-104) cells was reported from feces of diseased turkeys [181]. The Madin–Darby canine kidney (MDCK) cell line is widely described for isolation and propagation of influenza A virus but QT-6 and DF-1 cells might be favorable for propagating isolates from poultry due to high presence of sialic acid α2,3-gal linked receptors [162,182]. However, routinely, SPF eggs remain a gold standard for isolating AIV as indicated in the relevant chapter of the OIE manual [183]. Vero cells, DF-1, baby hamster kidney cells (BHK) and other vertebrate cell lines might be used to isolate Arboviruses (West Nile Virus, or Eastern Equine Encephalitis Virus) keeping in mind that some viruses, e.g., Turkey meningoencephalitis virus, rely on primary chicken cells [184].
Of high importance for avian virology are tumor cell lines established in the course of Marek’s disease. One of those cell lines, MDTC-RP19, established from turkey B cells, can be used to isolate and propagate hemorrhagic enteritis virus of turkeys (Turkey adenovirus 3 (TAdV-3) [185]. Similarly, chicken anemia virus (CAV) can be isolated and propagated in a MDV transformed T-cell line (MDCC-MSB1) established from a tumor in chickens [186].
A spontaneously immortalized turkey turbinate cell line displayed good susceptibility for avian metapneumovirus with higher titers than in commonly used Vero cells, albeit with delayed replication [187]. No other viruses were tested on this cell line although quite some potential would be available considering the number of pathogens targeting the respiratory tract as mentioned by the authors.
Despite the ability of cell lines reported above there are severe limitations for certain viruses to be isolated and the best examples are members of the Picornaviridae for which isolation is restricted to host or chicken embryos. This includes avian encephalomyelitis virus (AEV) but also turkey hepatitis virus (THV), and duck hepatitis A virus (DHAV). Similarly, isolation of turkey coronavirus (TCoV), chicken parvovirus or proventricular necrosis virus (CPNV), a member of the Birnaviridae, rely on SPF embryos [188,189]. Different to avian group A rotaviruses which can be isolated in MA104 cells success to isolate members of groups D, F and G from poultry species is very much limited [190,191]. Except CAV, circoviruses from waterfowl such as ducks (DuCV) or geese (GoCV) are isolated in respective embryos which are also used for isolating parvoviruses. Similarly, for goose parvovirus (Derzy’s disease) or Muscovy Duck parvovirus, both inducing hepatitis, isolation relies on embryos or primary cultures. A recently described goose pegivirus (GPgV), family Flaviviridae, was isolated in goose embryos and transferred to goose embryo fibroblast albeit with only a low increase in titer and no CPE was noticed [192].
In summary, it is very obvious that SPF eggs or primary cells are still an indispensable feature in a diagnostic laboratory. The susceptibility for numerous viruses is a clear surplus but the preparation is time consuming and costly. In addition, it needs a constant supply, and the procedures are difficult to standardize. Furthermore, there remain ethical concerns with regard to the use of primary cells and SPF embryos. However, it is difficult to believe that this situation will change in the future, since activities on established poultry cell lines are neglectable as no major achievements to establish cells lines from poultry species are reported in recent years.
6. Molecular Diagnostics
Molecular diagnostics comprises technologies identifying a disease or a pathogen by examining molecules, such as nucleic acids and proteins, in biological materials and environmental samples. In poultry medicine, molecular diagnostics is utilized to detect and analyze pathogen-specific DNA and/or RNA molecules. Although successful detection indicates the presence of the pathogen’s nucleic acid, it does not provide information if the detected pathogen was alive. Compared to conventional virological, bacteriological and parasitological investigations, molecular diagnostics offers a fast and specific alternative to detect an infectious agent without requirement of a live pathogen. In this respect the application of molecular diagnostics is superior, because samples can be inactivated before shipping, which simplifies their transport to a diagnostic laboratory. A good example is the use of Flinders Technology Associates (FTA) cards®, an envelope-like covering made of filter paper treated with a patented chemical mix that lyses cells, denatures proteins and preserves nucleic acids. In this way, the applied sample remains suitable for molecular diagnostics without the risk of spreading an infectious agent [193]. Nonetheless, the possibility that some infectious material might remain on the FTA card argues for careful handling since some studies reported detection of viable viruses or bacteria after their application on the FTA card [194,195]. In addition to use the intentionally inactivated material, the advantage of molecular diagnostics lies in the capacity to analyze a great variety of sample material such as all kinds of tissues and organs including formaldehyde-fixed paraffin-embedded (FFPE) material, body fluids, eggs, feathers, swabs and environmental samples such as dust, soil or litter.
In poultry medicine, the term molecular diagnostics generally considers various PCR-based methods. Even though strictly taken, methods such as in situ hybridization, immuno-staining or Enzyme-Linked Immunosorbent Assay (ELISA) also directly analyze molecules such as nucleic acids or proteins, they are considered as part of histology or serology and will be discussed therein. Due to easiness, speed and at the same time high specificity of PCR diagnostics detection of viral poultry pathogens is currently almost entirely based on this method [196]. In contrast to this, the detection of bacterial and parasitic pathogens is very often still carried out by classical bacteriology or parasitology. Exceptions are special situations in which the cultivation of the pathogen is either time consuming and/or complex such as for example the cultivation of Mycoplasma strains or impossible due the inherence of the starting material as FTA card or FFPE tissues [193,197]. Some molecular diagnostic assays are qualitative by nature assessing the presence or absence of the pathogen’s nucleic acids. Both conventional and real-time PCR tests are employed, although the prevailing tendency goes toward the fluorescently labelled oligonucleotide probe-based real-time PCR tests. This is mainly due to their greater specificity provided by the presence of a sequence-specific oligonucleotide probe as compared to conventional PCR or SYBR Green-based real-time PCRs, which lack this feature [198]. Furthermore, the capacity to specifically detect different targets within a single reaction, so called multiplexing, provides an additional surplus. Multiplexing is readily employed in PCR tests using an internal positive control, in which pathogen’s nucleic acid is detected in parallel to an independent nucleic acid target, either an endogenous internal control such as host-specific target or artificially added target DNA or RNA representing an exogenous internal control [199,200]. Inclusion of an internal control in a PCR assay is a requirement for assays accredited by ISO17025 to determine the success of the PCR within a given matrix [201]. Noteworthy is the implementation of multiplex PCR assays in i) detection of more than one pathogen in a course of a single PCR, or ii) for pathogen typing. Various assays for simultaneous detection of different pathogens have been described, such as protocols based on multiplex real-time PCR in detection of fowl poxvirus and reticuloenotheliosis virus [202,203] or detection of Mycoplasma gallisepticum and Mycoplasma synoviae [204] or conventional assays such as simultaneous detection of picorna-, astro- and caliciviruses [205] or Campylobacter, Arcobacter and Helicobacter species [206]. Similarly, diverse multiplex PCR assays are available for detecting and typing a pathogen within the same assay, and avian influenza viruses subjected to multiplex real-time PCR are a good example [207,208]. Similarly, rapid serotyping of Erysipelothrix rhusiopathiae by conventional multiplex PCR can be mentioned [209,210].
The multiplexing scheme is also included in the high-throughput methods using devices such as Genome Lab Gene Expression Profiler (GeXP) or Luminex in analysis of PCR products. The GeXP system uses capillary electrophoresis for fine separation of PCR products, detecting length differences of just few base pairs which enables the detection of up to 30 different targets [211]. The use of GeXP-based PCR methods was reported for both (i) concurrent detection of different pathogens and (ii) for pathogen typing [212,213,214,215,216]. Another application involving multiplexing is the alliance of PCR with the Luminex system, in which the detection of different targets is based on measuring beads that are coupled with target specific nucleic acids [217]. Similar to the GeXP system, bead-based multiplex assays are applicable for high throughput and can simultaneously analyze up to 50–500 different targets within a single sample depending on the instrument and assay applied. In diagnosing poultry infectious agents bead-based multiplex PCRs were used in (i) parallel detection of different pathogens and (ii) typing of pathogens [218,219,220,221,222]. The clear advantage of both GeXP-analyzer- and Luminex-based methods is the much higher number of targets that can be detected in a single reaction as compared to real-time PCR or conventional PCR, which speeds up the diagnostics process. However, the high price of devices which are essential for such analyses (GeXP-analyzer or Luminex), challenges the justification for these methodologies, especially if it is not used at full capacity.
Specific application of PCR is a DIVA (differentiating infected from vaccinated animals)-PCR, a method used for screening a vaccinated flock for infections with a filed/virulent strain, as for example in Marek’s disease virus (MDV), Mycoplasma gallisepticum, Mycoplasma synoviae or turkey meningoencephalitis virus [223,224,225,226]. In order to accomplish functionality, DIVA tests should be able to distinguish between a vaccine and a field strain even in situations when multiple strains are present within the sample. In some cases, such as MDV Rispens or CVI988 vaccine and virulent/oncogenic MDV strain, difference between the vaccine strain and wild-type/virulent strain is often minor, which introduces challenges in methodology. In the context of differentiating vaccinated and unvaccinated birds, method Mismatch Amplification Mutation Assay PCR (MAMA-PCR) has been applied for MDV, Mycoplasma gallisepticum and Mycoplasma synoviae [227,228,229]. The approach is based on the presence of a single nucleotide polymorphism (SNP) between vaccine and field/virulent strain, which is incorporated at 3′-end of one of the primers. The incorporated SNP causes mismatching of the primer with the template, which should ultimately result in the abolition of amplification. However, the stringency of this process is not high and very often one of the specific primers demonstrates a certain level of cross-reaction with the opposite template detecting the unwanted strain/type, such as CVI988 vaccine-specific primers detecting MDV oncogenic strains [227]. Therefore, a set of good controls for evaluating the MAMA test system is needed.
In addition to sole PCR-based assays, pathogen typing often includes a combination of conventional PCR with sequence analysis of the PCR product. The advantage of such an approach is a much broader detection range of strains/types/variants, as for viral pathogens with high diversity such as FAdV, infectious bronchitis virus (IBV), IBDV or Newcastle disease virus (NDV) [230,231,232,233,234,235,236,237]. Targets used for typing/classification are in general genes coding for surface proteins which display high variation among different strains/variants as compared to pathogen’s proteins not exposed to the host. This characteristic is directly related to the immune response of the host and frequently also to an earlier classification method such as serotyping, patho-typing, or protecto-typing. Sequence variations often cluster in specific gene regions that are flanked by a more conserved sequence, the aspect used for designing the primers to amplify as many variants/strains as possible. Classification employs a phylogenetic analysis of obtained sequences integrating them with available data from reference strains.
In recent years the implementation of NGS and third-generation sequencing (TGS) in diagnosing infectious diseases in poultry has gained its importance, as rapid advancements in sequencing technologies, bioinformatics and computational tools have made these approaches broadly available [238]. Both methodologies enable random sequencing of all nucleic acids in the sample, detecting potentially all microbes and viruses. Such assumption-free procedure enables the discovery of (un)expected or novel pathogens (re)defining the etiology [239]. This is especially valuable in cases that have exhausted the available diagnostic analyses, as for example, in disclosure of a etiological agents of fulminating enteritis of French guinea fowl or hepatitis in pheasants, which were only possible with metagenomic NGS of samples from diseased birds [240,241]. Since metagenomic analyses produce vast amount of data, scrutiny of all available clinical, diagnostic and epidemiological results is required for accurate evaluation of the respective case. Metagenomic analyses are currently also very popular in epidemiological investigations of various health issues and frequently result in finding novel species [239,242].
However, proper study designs and functional knowledge about microbes and viruses in a certain host are a prerequisite for an accurate evaluation of microbial species involved in clinically relevant cases. The NGS or TGS of clinical samples have also been shown beneficial for detecting pathogens with high mutation rate such as RNA viruses, which due to their error-prone genome replication and re-assortment of genome segments demonstrate increased genetic variation [243,244,245]. Since classical molecular detection using PCR methods is based on specific targets in pathogen’s genome, introduction of new mutations might hinder their proper detection [246]. In addition to the fact that the application of NGS and TGS opens powerful modern strategies in molecular diagnostics, several drawbacks or bottlenecks limit these methodologies from being a tool of choice. Pathogen detection is more expensive and labor intensive compared with classical molecular diagnostics.
In particular, the generation of a large quantity of raw data and their complex analysis demand high-performance computers and extensive bioinformatics analysis [247]. Furthermore, untargeted sequencing of total DNA and/or RNA from clinical samples results in very low amount of microbes and especially viral nucleic acids as compared to the host cellular DNA and RNA; therefore, special sample preparation such as depletion of host nucleic acids and /or enrichment of viral DNA/RNA is frequently required [248,249,250,251,252,253]. However, considering the speed of the development of techniques, protocols and tools focused on NGS and TGS, one can foresee that these technologies will soon emerge as the tool of choice in molecular diagnostics.
7. Serology
After contact with an infectious agent, the poultry immune system frequently reacts with the production of antibodies. Typically, serology uses specific antigen–antibody reactions for the detection of bacteria, viruses or parasites that may be difficult to detect by other methods. In contrast to the isolation or detection of infectious agents, especially viruses, serological testing is fast and easy to perform with minimal laboratory requirements. Primarily, it focuses on the presence, absence or level of specific antibodies in the serum. In practice, antibodies can be detected in a number of body fluids such as egg, tears, saliva, and mucus secretions [254,255].
Many of the classical serological techniques (e.g., the agar gel precipitation test, plate agglutination tests (RSA), and hemagglutination inhibition tests (HI)) have been employed for decades; standardized reference sera and antigens for many poultry pathogens are commercially available worldwide. Considering the high flexibility, quick turnaround and the large sample size to be processed at a time, ELISA assays have become routine procedures for flock health monitoring and for poultry diagnostics alike, with commercial systems available for the majority of poultry pathogens.
In addition to direct pathogen detection described above in Section 3 and Section 6, some national/international poultry health control programs for certain infections/diseases (Mycoplasma gallisepticum, Mycoplasma meleagridis, Salmonella Gallinarum/Pullorum, and avian influenza) employ sero-surveillance to provide evidence for circulating infections within poultry populations [256,257,258,259]. Office International des Epizooties (OIE) standard serological tests—RSA and ELISA—are recommended for flock screening [50,86]. However, variations in specificity and sensitivity are known for Mycoplasma species and need to be considered. Such variations were mainly reported in contexts with the application of inactivated vaccines against different other bacterial agents [260,261,262,263,264]. Thus, reactors have to be confirmed by HI and suspected flocks are generally investigated with molecular methods as well. Over the last century, successful control of Salmonella Gallinarum/Pullorum has been achieved by applying RSA in the field [50,265]. Furthermore, a recent study showed the high specificity of this test system even in flocks vaccinated against Salmonella Enteritidis which was often seen as a problem due to cross-reactions [266].
Different to this, the majority of ELISA systems are used for vaccination control or even to predict the time point of vaccination as shown for infectious bursal disease [267]. However, ELISA kits from different manufacturers may vary in specificity and/or sensitivity, resulting in conflicting titer profiles [268], which needs to be considered when interpreting results and for decision making. Archived serum samples are helpful to retrospectively trace pathogen transmission as shown for vertically induced adenovirus gizzard erosion (AGE) [15].
So far, single assays are very widespread, with the need to perform individual tests for each pathogen on independent samples. Moreover, the majority of commonly applied serological assays do not allow the differentiation of antigenically related organisms with consequences to differentiate between vaccination response or natural infection. Furthermore, it is not possible to discriminate serological response against a specific serotype for which more cumbersome test systems, such as hemagglutination inhibition or neutralization assays, have to be applied. Recent developments based upon recombinant antigens and proteins have improved assay specificity and facilitated standardization of interpretation. Recent developments enable the differentiation between birds being vaccinated with chemically inactivated viruses or with viral vector vaccines and birds being infected with field virus, as shown for, e.g., avian influenza virus, NDV, ILTV, IBDV and FAdV [269,270,271,272].
Recently, microsphere immunoassays were developed, enabling the measurement and discrimination of numerous analytes with the power of automation. Such bead-based systems were developed to detect antibodies against multiple pathogens or subtypes simultaneously as shown for influenza A virus, NDV, IBV, ILTV, FAdV and/or aHEV [273,274,275,276,277,278]. With this, much more data are obtained in comparison to a single component. The large number of analytes to be measured offers high potential and can combine several pathways of host response. This might not only include immunological response characterized by antibodies, but can also include broader physiological parameters to assess the well-being of animals such as heat shock proteins or biomarkers for gut heath. Finally, for pathogen detection, biosensors gained certain attractiveness due to their application in the field [279]. Practicability simply depends on the technical composition of the system. Those systems gained importance especial to detect avian influenza virus on site as reviewed by Astill et al. [19].
8. Histology
Microscopic changes in organs and tissue structures caused by infectious agents are important features for diagnosis, even though the occurrence of pathognomonic lesions is exceptional and differences in the anatomy of birds and mammals or diseases affecting particular bird species may cause difficulties in the interpretation of histological changes [280]. However, the exact description of microscopic changes in tissues can be a significant contribution for a presumptive diagnosis and it is even more vital for endemic pathogens with substantial variation in pathogenicity.
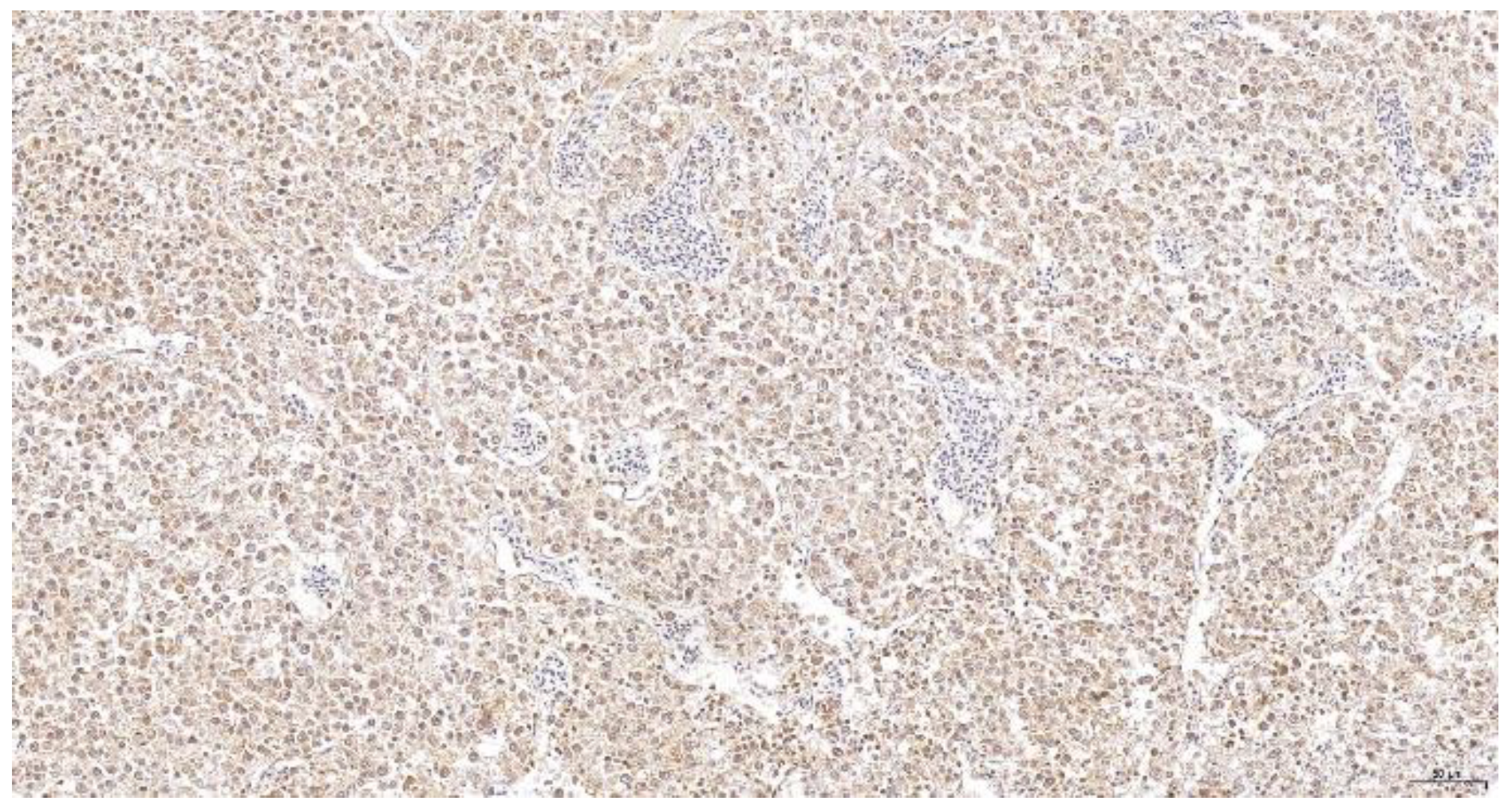
For histopathology, different tissue preparations can be processed to evaluate changes in host organs, on the organ or cellular levels. Most commonly, tissue samples are fixed in formalin, dehydrated, paraffin-embedded and stained with hematoxylin-eosin staining [281]. In recent decades, different techniques for the specific detection of molecular structures in cells have been developed [282]. These include tools such as immunohistochemistry and in situ hybridization using specific antibodies and nucleic acid probes. This is of particular relevance for the detection of pathogens that cannot be identified or determined by morphological characteristics. Some diseases, such as Marek’s disease, do not always cause typical histological changes and require further diagnostic confirmation. The presumptive diagnosis of Marek’s disease relies on identifying the involved tissues (inner organs, eye, skin and nerves) and the main type of involved lymphoid cells (T cells) to discriminate it from lymphoid leucosis (Figure 1) [283].
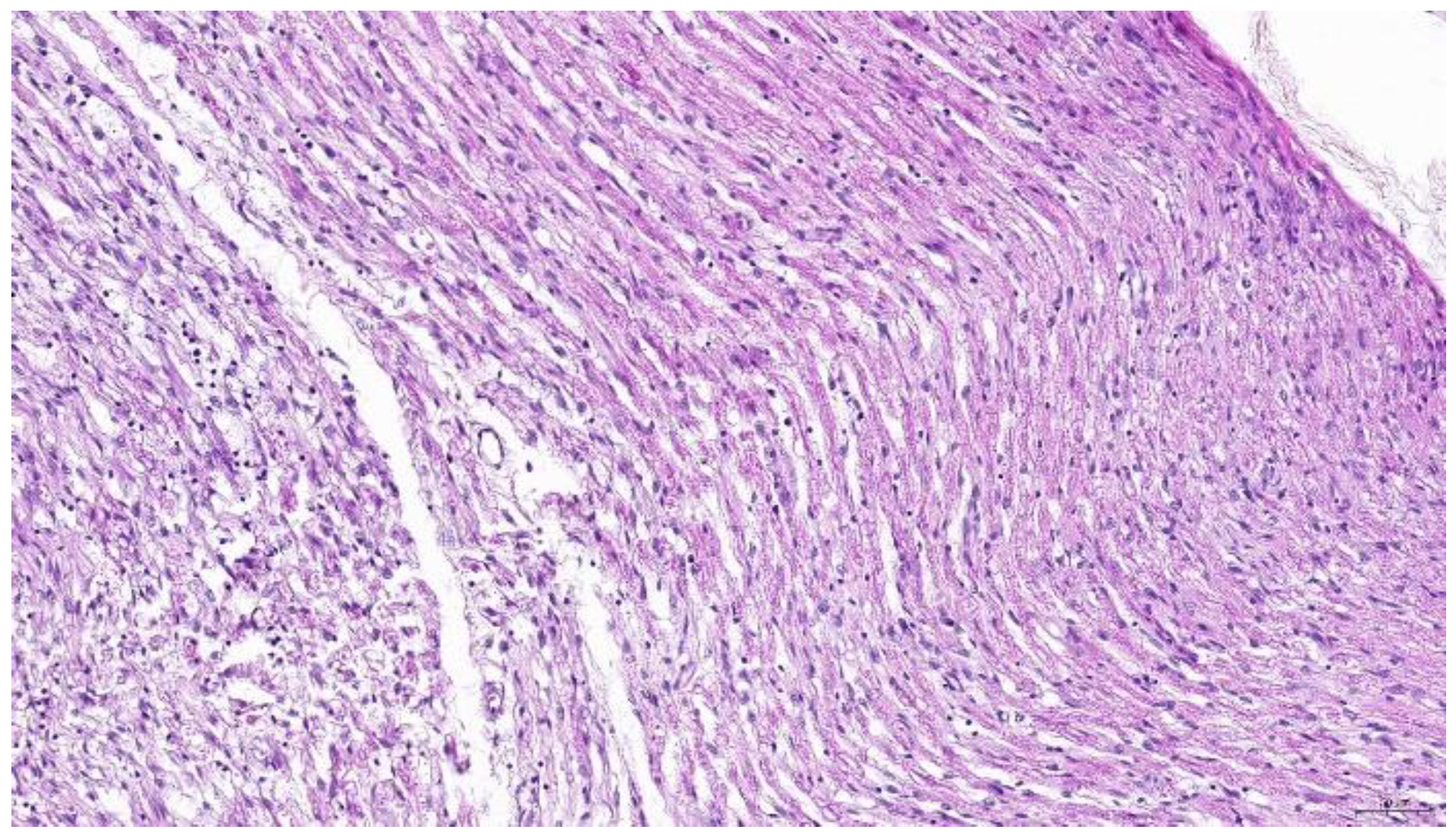
However, reticuloendotheliosis can transform T cells, potentially infiltrating nerve tissue [284,285]. Furthermore, peripheral neuropathy results in similar lesions compared to Type B lesions in nerves reported for Marek’s disease (Figure 2) [286].
Therefore, the presumptive diagnosis of Marek’s disease must be confirmed by the detection of the virus in the tissue with the Meq antigen being constantly expressed in MDV tumors [287]. Consequently, recent advances using histological tools for the detection of MDV involve the application of polyclonal and monoclonal antibodies against Meq that can be used for the specific diagnosis of the disease in paraffin embedded tissues [288,289].
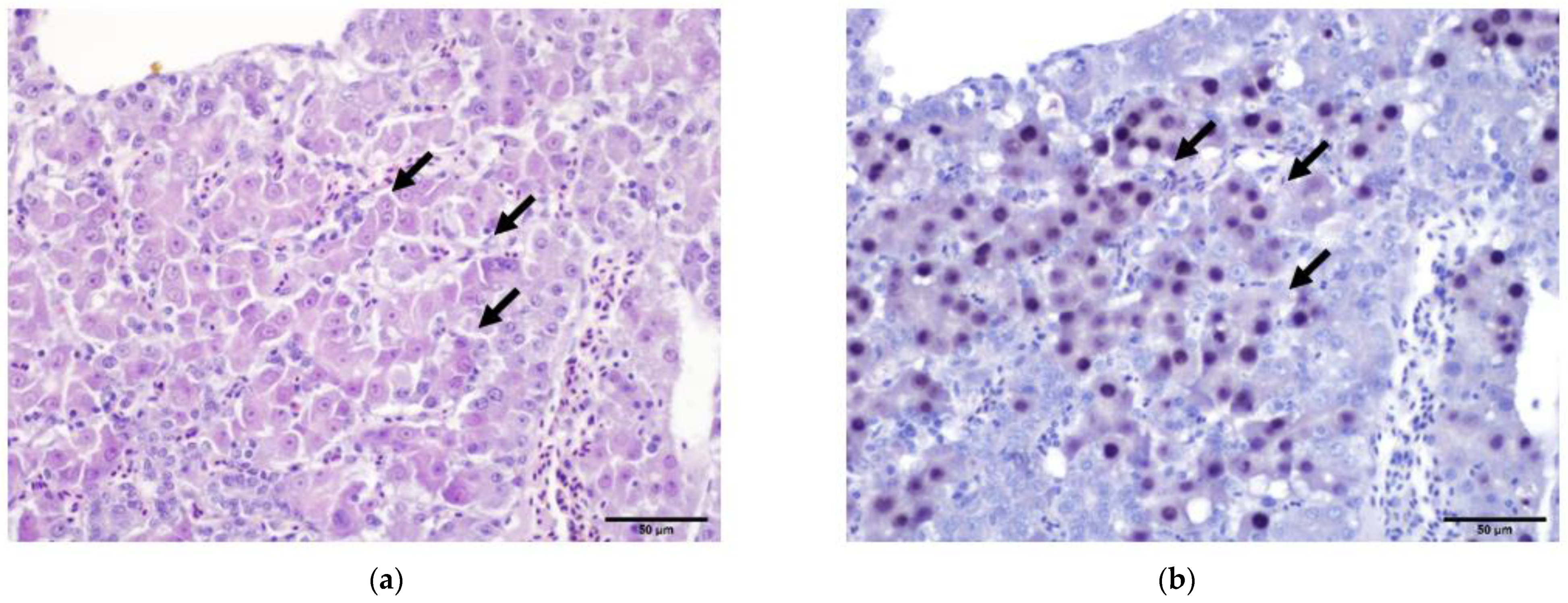
For other viral diseases, inclusion bodies in host cells might be pathognomonic in histopathological findings due to their specific characteristics, such as the location in an infected cell (nucleus, cytoplasm) as well as the staining affinity using HE. Based on such parameters, Kato et al. [290] differentiated intra-cytoplasmatic accumulations in Type A and Type B inclusions, both observed in cells infected with fowlpox. Similarly, intranuclear hepatic inclusions in the course of viral hepatitis must be well examined for a valid presumptive diagnosis as the causative virus may be tentatively determined by the structure of inclusion bodies [291]. In a recent case of hepatitis in pheasants, histopathology clearly revealed inclusion bodies [241]. Inclusion body hepatitis (IBH) caused by certain fowl adenoviruses (FAdVs) of the genus Aviadenovirus is not described in pheasants, although such birds can suffer from marble spleen disease (MSDV), a member of the genus Siadenovirus, characterized by inclusions within mononuclear cells of the spleen [292,293]. However, based on the histopathological changes and the features of the intranuclear inclusions, showing amphophilic to acidophilic prominent inclusion bodies that filled the nucleoplasm of hepatocytes (Figure 3a), a parvovirus was suggested to be the etiological agent. The use of specific probes against the parvovirus DNA confirmed the presence of this virus in the nuclei of cells (Figure 3b).
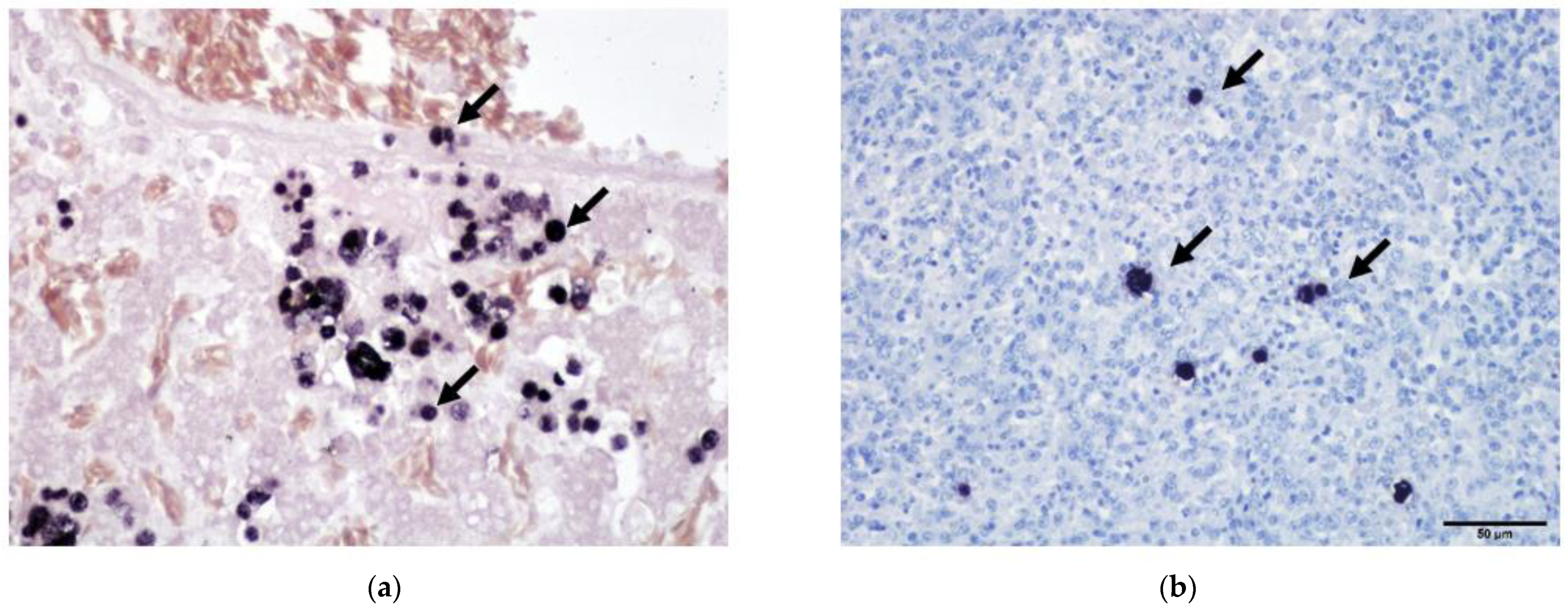
Advanced histological techniques are also crucial in diagnosing pathogens which are microscopically visible. The protozoan Histomonas meleagridis is a parasite of poultry that infiltrates different organs and can cause high morbidity and mortality in infected flocks [10]. The parasite is observed in tissue samples stained by hematoxylin-eosin (HE) or the Periodic acid-Schiff method [294,295] but low numbers in tissues without inflammation can easily be missed. To increase the sensitivity of conventional staining protocols, in situ hybridization and immunohistochemistry were developed [296,297]. By this, a single parasite can be detected and differentiation to other flagellates is possible. Furthermore, Histomonas meleagridis causes similar lesions in liver and ceca of poultry such as other trichomonads [298]. Conventional staining reveals the presence of the protozoa (Figure 4a) but in situ hybridization allows the differentiation of Histomonas meleagridis and Tetratrichomonas gallinarum (Figure 4b) [299]. This can furthermore be necessary for the diagnosis of other infectious agents such as bacteria to localize and identify the relevant pathogenic species of co-infected birds as previously reported [300].
In the future, digitalization of histological preparations is expected to be more and more implemented [301]. This implies automatic detection of tissue structures such as different chicken leukocytes in digital images of histological sections and its quantification without human bias as recently demonstrated [302]. Furthermore, computerized quantification can be combined with the localization of a pathogen and specific immune cells in the same tissue to identify relevant immune traits [303]. Digital pictures of microscopic lesions are useful for documentation of cases and can be stored in an archive and data repositories. By this, pictures with metadata can be made accessible worldwide to be used for different studies and applications. Computer-aided diagnosis can support the examiner in evaluating microscopic changes. The use of artificial intelligence (AI) enables the development of algorithms for specific histopathological changes. AI is already being much used in the medical field, including infectious disease diagnostics [304]. However, there are still certain limitations of AI-based computer-assisted pathological diagnosis, including a lack of confidence in diagnostic results, inconvenience in practical use and simplicity of function. [305]. Nevertheless, it can be expected that digitalization of histopathology and the application of AI is a promising approach that will improve diagnosing poultry diseases.
9. Outlook and Challenges
The sustainability of commercial poultry production depends on quick and accurate detection of impaired flock health in order to set up interventions and to adjust prophylactic strategies. Considering animal welfare, targeted treatment and efficacious prophylaxis, together with the extent and quality of diagnostic procedures, have to be constantly improved keeping in mind that the lack of efficacious drugs and chemicals is a serious limitation. Standardization of laboratory techniques is an important aspect of disease diagnosis and modern laboratories possess an externally validated quality assurance system. This includes numerous features, one of which is the regular participation at interlaboratory comparisons to evaluate the performance of a certain procedure. We already demonstrated the value of such comparisons for accurate detection of Mycoplasma gallispeticum and Mycoplasma synoviae by PCR approximately 20 years ago [306]. In the field of PCR or serology these comparisons are very common today, but they remain exceptional in disciplines such as histology, where personal experience and skills dominate the technology, a characteristic of bioinformatics as well. This highlights the importance of linking expertise with technologies to develop a holistic approach as a basic strategy to diagnose (infectious) diseases in poultry (Figure 5).
In this concept, diagnosing poultry diseases starts at the farm with precise recoding and detailed anamnesis. To date, PLF tools for early detection and recognition of diseases on farm are not routinely available, as most data have been acquired under experimental conditions and farm equipment has to be updated. Nevertheless, it can be expected that with additional research, leading to progress in technology, the validation of behavioral tracking and analyses of flock data collected in poultry house environments, PLF developments offer great potential for the future of disease diagnostics in poultry. In the laboratory, diagnostic methods will be further developed and bioinformatic tools will become more and more important, replacing conventional methods, with molecular techniques versus pathogen isolation being at the forefront. This will coincide with more demanding equipment and technologies, although numerous conventional methods will remain, and pathogen isolation would be a good example to study functional aspects but also to produce certain types of vaccines.
Consequently, in the future, more information technology will be applied to connect diagnostic investigations with the farm environment. Modern laboratories already use a database accessible via a mobile app to trace samples during investigations and to follow up records. This indicates the need for close interaction between all people involved in diagnostic investigations keeping in mind that specialization is ongoing, and skills of experts will diverge even more. In such a scenario, it remains challenging to interlink people and infrastructure in order to generate a surplus as outlined in Figure 5. This is more easily established in integrated production systems with the downside that data are less accessible for research and science.
Author Contributions
Conceptualization, introduction and conclusions, D.L., I.B., B.G., C.H. and M.H.; diagnostic activities on farm and serology, B.G.; bacteriology, C.H.; parasitology and histology, D.L.; molecular diagnostics, I.B.; virology and outlook, I.B. and M.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Figure 5 was created with BioRender.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schat, K.A.; Nagaraja, K.V.; Saif, Y.M. Pullorum Disease: Evolution of the Eradication Strategy. Avian Dis. 2021, 65, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Wigley, P. Salmonella enterica serovar Gallinarum: Addressing fundamental questions in bacteriology sixty years on from the 9R vaccine. Avian Pathol. 2017, 46, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Payne, L.N.; Nair, V. The long view: 40 years of avian leukosis research. Avian Pathol. 2012, 41, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.J.; Niemi, J.; Christensen, J.-P.; Tranter, R.B.; Bennett, R.M. A review of the financial impact of production diseases in poultry production systems. Anim. Prod. Sci. 2019, 59, 1585. [Google Scholar] [CrossRef]
- Ayala, A.J.; Yabsley, M.J.; Hernandez, S.M. A review of pathogen transmission at the backyard chicken-wild bird interface. Front. Vet. Sci. 2020, 7, 539925. [Google Scholar] [CrossRef]
- Whitehead, M.L.; Roberts, V. Backyard poultry: Legislation, zoonoses and disease prevention. J. Small Anim. Pract. 2014, 55, 487–496. [Google Scholar] [CrossRef]
- National Poultry Improvement Plan. Available online: https://www.poultryimprovement.org/default.cfm (accessed on 9 February 2023).
- EU. Commission Regulation 2019/268 of 15 February 2019 Amending Regulations (EU) No 200/2010, (EU) No 517/2011, (EU) No 200/2012 and (EU) No 1190/2012 as regards Certain Methods for Salmonella Testing and Sampling in Poultry. L46, 18.02.2019, 11-16. 2019. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32019R0268&rid=7 (accessed on 21 April 2023).
- Hess, M. Commensal or pathogen—A challenge to fulfil Koch’s Postulates. Br. Poult. Sci. 2017, 58, 1–12. [Google Scholar] [CrossRef]
- Hess, M.; Liebhart, D.; Bilic, I.; Ganas, P. Histomonas meleagridis—New insights into an old pathogen. Vet. Parasitol. 2015, 208, 67–76. [Google Scholar] [CrossRef]
- Liebhart, D.; Hess, M. Spotlight on Histomonosis (blackhead disease): A re-emerging disease in turkeys and chickens. Avian Pathol. 2020, 49, 1–4. [Google Scholar] [CrossRef]
- Collett, S.R.; Smith, J.A.; Boulianne, M.; Owen, R.L.; Gingerich, E.; Singer, R.S.; Johnson, T.J.; Hofacre, C.L.; Berghaus, R.D.; Stewart-Brown, B. Principles of Disease Prevention, Diagnosis, and Control. In Diseases of Poultry; Swayne, D.E., Boulianne, M., Logue, C.M., McDougald, L.R., Nair, V., Suarez, D.L., Wit, S., Grimes, T., Johnson, D., Kromm, M., et al., Eds.; Wiley: Hoboken, NJ, USA, 2020; pp. 1–78. ISBN 9781119371168. [Google Scholar]
- Skinner, J.T.; Bauer, S.; Young, V.; Pauling, G.; Wilson, J. An economic analysis of the impact of subclinical (mild) necrotic enteritis in broiler chickens. Avian Dis. 2010, 54, 1237–1240. [Google Scholar] [CrossRef]
- Long, K.E.; Hastie, G.M.; Ojkić, D.; Brash, M.L. Economic impacts of white chick syndrome in Ontario, Canada. Avian Dis. 2017, 61, 402–408. [Google Scholar] [CrossRef]
- Grafl, B.; Aigner, F.; Liebhart, D.; Marek, A.; Prokofieva, I.; Bachmeier, J.; Hess, M. Vertical transmission and clinical signs in broiler breeders and broilers experiencing adenoviral gizzard erosion. Avian Pathol. 2012, 41, 599–604. [Google Scholar] [CrossRef]
- Haug, A.; Gjevre, A.-G.; Skjerve, E.; Kaldhusdal, M. A survey of the economic impact of subclinical Eimeria infections in broiler chickens in Norway. Avian Pathol. 2008, 37, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Berckmans, D. General introduction to precision livestock farming. Anim. Front. 2017, 7, 6–11. [Google Scholar] [CrossRef]
- Neethirajan, S. Automated tracking systems for the assessment of farmed poultry. Animals 2022, 12, 232. [Google Scholar] [CrossRef] [PubMed]
- Astill, J.; Dara, R.A.; Fraser, E.D.G.; Sharif, S. Detecting and predicting emerging disease in poultry with the implementation of new technologies and big data: A focus on avian influenza virus. Front. Vet. Sci. 2018, 5, 263. [Google Scholar] [CrossRef]
- Mbelwa, H.; Mbelwa, J.; Machuve, D. Deep convolutional neural network for chicken diseases detection. Int. J. Adv. Comput. Sci. Appl. 2021, 12, 295. [Google Scholar] [CrossRef]
- Guarino, M.; Norton, T.; Berckmans, D.; Vranken, E.; Berckmans, D. A blueprint for developing and applying precision livestock farming tools: A key output of the EU-PLF project. Anim. Front. 2017, 7, 12–17. [Google Scholar] [CrossRef]
- Schillings, J.; Bennett, R.; Rose, D.C. Exploring the potential of precision livestock farming technologies to help address farm animal welfare. Front. Anim. Sci. 2021, 2, 639678. [Google Scholar] [CrossRef]
- Okada, H.; Suzuki, K.; Itoh, T.; Tsukamoto, K. Wireless sensor system for detection of avian influenza outbreak farms at an early stage. In Proceedings of the 8th Annual IEEE Conference on Sensors, Christchurch, New Zealand, 25–28 October 2009; pp. 1374–1377, ISBN 978-1-5090-4545-7. [Google Scholar]
- Okada, H.; Suzuki, K.; Kenji, T.; Itoh, T. Applicability of wireless activity sensor network to avian influenza monitoring system in poultry farms. J. Sens. Technol. 2014, 4, 18–23. [Google Scholar] [CrossRef]
- Banakar, A.; Sadeghi, M.; Shushtari, A. An intelligent device for diagnosing avian diseases: Newcastle, infectious bronchitis, avian influenza. Comput. Electron. Agric. 2016, 127, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Bi, M.; Guo, J.; Wu, S.; Zhang, T. Development of an early warning algorithm to detect sick broilers. Comput. Electron. Agric. 2018, 144, 102–113. [Google Scholar] [CrossRef]
- Huang, J.; Wang, W.; Zhang, T. Method for detecting avian influenza disease of chickens based on sound analysis. Biosyst. Eng. 2019, 180, 16–24. [Google Scholar] [CrossRef]
- Cuan, K.; Zhang, T.; Huang, J.; Fang, C.; Guan, Y. Detection of avian influenza-infected chickens based on a chicken sound convolutional neural network. Comput. Electron. Agric. 2020, 178, 105688. [Google Scholar] [CrossRef]
- Noh, J.-Y.; Kim, K.-J.; Lee, S.-H.; Kim, J.-B.; Kim, D.-H.; Youk, S.; Song, C.-S.; Nahm, S.-S. Thermal image scanning for the early detection of fever induced by highly pathogenic avian influenza virus infection in chickens and ducks and its application in farms. Front. Vet. Sci. 2021, 8, 616755. [Google Scholar] [CrossRef]
- Colles, F.M.; Cain, R.J.; Nickson, T.; Smith, A.L.; Roberts, S.J.; Maiden, M.C.J.; Lunn, D.; Dawkins, M.S. Monitoring chicken flock behaviour provides early warning of infection by human pathogen Campylobacter. Proc. Biol. Sci. 2016, 283, 20152323. [Google Scholar] [CrossRef]
- Sadeghi, M.; Banakar, A.; Khazaee, M.; Soleimani. An intelligent procedure for the detection and classification of chickens infected by Clostridium perfringens based on their vocalization. Rev. Bras. Cienc. Avic. 2015, 17, 537–544. [Google Scholar] [CrossRef]
- Grilli, G.; Borgonovo, F.; Tullo, E.; Fontana, I.; Guarino, M.; Ferrante, V. A pilot study to detect coccidiosis in poultry farms at early stage from air analysis. Biosyst. Eng. 2018, 173, 64–70. [Google Scholar] [CrossRef]
- Borgonovo, F.; Ferrante, V.; Grilli, G.; Pascuzzo, R.; Vantini, S.; Guarino, M. A data-driven prediction method for an early warning of coccidiosis in intensive livestock systems: A preliminary study. Animals 2020, 10, 747. [Google Scholar] [CrossRef]
- Ahmed, G.; Malick, R.A.S.; Akhunzada, A.; Zahid, S.; Sagri, M.R.; Gani, A. An approach towards IoT-based predictive service for early detection of diseases in poultry chickens. Sustainability 2021, 13, 13396. [Google Scholar] [CrossRef]
- Carroll, B.T.; Anderson, D.; Daley, W.; Harbert, S.; Britton, D.; Jackwood, M. Detecting symptoms of diseases in poultry through audio signal processing. In Proceedings of the 2014 IEEE Global Conference on Signal and Information Processing (GlobalSIP 2014), Atlanta, GA, USA, 3–5 December 2014; pp. 1132–1135, ISBN 978-1-5090-4545-7. [Google Scholar]
- Rizwan, M.; Carroll, B.T.; Anderson, D.V.; Daley, W.; Harbert, S.; Britton, D.F.; Jackwood, M.W. Identifying rale sounds in chickens using audio signals for early disease detection in poultry. In Proceedings of the 2016 IEEE Global Conference on Signal and Information Processing (GlobalSIP), Washington, DC, USA, 7–9 December 2016; pp. 55–59, ISBN 978-1-5090-4545-7. [Google Scholar]
- Mahdavian, A.; Minaei, S.; Marchetto, P.M.; Almasganj, F.; Rahimi, S.; Yang, C. Acoustic features of vocalization signal in poultry health monitoring. Appl. Acoust. 2021, 175, 107756. [Google Scholar] [CrossRef]
- Carpentier, L.; Vranken, E.; Berckmans, D.; Paeshuyse, J.; Norton, T. Development of sound-based poultry health monitoring tool for automated sneeze detection. Comput. Electron. Agric. 2019, 162, 573–581. [Google Scholar] [CrossRef]
- Okinda, C.; Lu, M.; Liu, L.; Nyalala, I.; Muneri, C.; Wang, J.; Zhang, H.; Shen, M. A machine vision system for early detection and prediction of sick birds: A broiler chicken model. Biosyst. Eng. 2019, 188, 229–242. [Google Scholar] [CrossRef]
- Cuan, K.; Zhang, T.; Li, Z.; Huang, J.; Ding, Y.; Fang, C. Automatic Newcastle disease detection using sound technology and deep learning method. Comput. Electron. Agric. 2022, 194, 106740. [Google Scholar] [CrossRef]
- Aziz, N.A.; Bin Othman, M.F. Binary classification using SVM for sick and healthy chicken based on chicken’s excrement Image. Pertanika J. Sci. Technol. 2017, 25, 315–324. [Google Scholar]
- Alex, A.G.R.; Joseph, G.J. Real-time poultry health identification using IoT test setup, optimization and results. In Advances in Signal Processing and Intelligent Recognition Systems; Thampi, S.M., Marques, O., Krishnan, S., Li, K.-C., Ciuonzo, D., Kolekar, M.H., Eds.; Springer: Singapore, 2019; pp. 30–40. ISBN 978-981-13-5757-2. [Google Scholar]
- Wang, J.; Shen, M.; Liu, L.; Xu, Y.; Okinda, C. Recognition and classification of broiler droppings based on deep convolutional neural network. J. Sens. 2019, 2019, 3823515. [Google Scholar] [CrossRef]
- Xiao, L.; Ding, K.; Gao, Y.; Rao, X. Behavior-induced health condition monitoring of caged chickens using binocular vision. Comput. Electron. Agric. 2019, 156, 254–262. [Google Scholar] [CrossRef]
- Zhuang, X.; Zhang, T. Detection of sick broilers by digital image processing and deep learning. Biosyst. Eng. 2019, 179, 106–116. [Google Scholar] [CrossRef]
- Liu, L.; Li, B.; Zhao, R.; Yao, W.; Shen, M.; Yang, J. A novel method for broiler abnormal sound detection using WMFCC and HMM. J. Sens. 2020, 2020, 2985478. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, C. Design of sick chicken automatic detection system based on improved residual network. In Proceedings of the 2020 IEEE 4th Information Technology, Networking, Electronic and Automation Control Conference (ITNEC 2020), Chongqing, China, 12–14 June 2020. [Google Scholar]
- Xiong, X.; Lu, M.; Yang, W.; Duan, G.; Yuan, Q.; Shen, M.; Norton, T.; Berckmans, D. An automatic head surface temperature extraction method for top-view thermal image with individual broiler. Sensors 2019, 19, 5286. [Google Scholar] [CrossRef]
- Hrabak, J.; Bitar, I.; Papagiannitsis, C.C. Combination of mass spectrometry and DNA sequencing for detection of antibiotic resistance in diagnostic laboratories. Folia Microbiol. 2020, 65, 233–243. [Google Scholar] [CrossRef]
- OIE Terrestrial Manual. 2. Fowl Typhoid and Pullorum Disease. Chapter 2.3.11. 2018. Available online: https://www.woah.org/fileadmin/Home/eng/Health_standards/tahm/2.03.11_FOWL_TYPHOID.pdf (accessed on 21 April 2023).
- ISO 6579-1:2017/AMD 1:2020; Microbiology of the Food Chain—Horizontal Method for the Detection and Enumeration and Serotyping of Salmonella—Part 1: Detection of Salmonella spp.—Amendment 1: Broader Range of Incubation Temperatures, Amendment to the Status of Annex D, and Correction of the Composition of MSRV and SC. ISO: Geneva, Switzerland, 2020.
- Valentin-Weigand, P. Teil II Allgemeine Bakteriologie. In Tiermedizinische Mikrobiologie, Infektions- und Seuchenlehre; Selbitz, H.J., Truyen, U., Valentin-Weigand, P., Eds.; Enke Verlag: Erlangen, Germany, 2015; pp. 96–136. ISBN 978-3-8304-1262-5. [Google Scholar]
- Buxton, R. Blood Agar Plates and Hemolysis Protocol. American Society for Microbiology. 2005. Available online: https://asm.org/getattachment/7ec0de2b-bb16-4f6e-ba07-2aea25a43e76/protocol-28 (accessed on 21 April 2022).
- Jung, B.; Hoilat, G.J. MacConkey Medium. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- ISO 10272-1:2017; Microbiology of the Food Chain—Horizontal Method for Detection and Enumeration of Campylobacter spp. ISO: Geneva, Switzerland, 2017.
- Odds, F.C. Sabouraud(’s) agar. J. Med. Vet. Mycol. 1991, 29, 355–359. [Google Scholar] [CrossRef]
- Blackall, P.J.; Soriano-Vargas, E. Infectious Coryza and related bacterial infections. In Diseases of Poultry, 14th ed.; Swayne, D.E., Bouillane, M., Logue, C.M., McDougald, L.R., Nair, V., Suarez, D.L., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2020; pp. 890–906. [Google Scholar]
- Mappley, L.J.; La Ragione, R.M.; Woodward, M.J. Brachyspira and its role in avian intestinal spirochaetosis. Vet. Microbiol. 2014, 168, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Wang, H.; Liu, L.; Miao, Z.; Huo, Y.; Zhong, Z. Prevalence and characterization of Clostridium perfringens isolated from different chicken farms in China. Anaerobe 2021, 72, 102467. [Google Scholar] [CrossRef]
- Feberwee, A.; Mekkes, D.R.; de Wit, J.J.; Hartman, E.G.; Pijpers, A. Comparison of culture, PCR, and different serologic tests for detection of Mycoplasma gallisepticum and Mycoplasma synoviae infections. Avian Dis. 2005, 49, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Crawshaw, T. A review of the novel thermophilic Campylobacter, Campylobacter hepaticus, a pathogen of poultry. Transbound. Emerg. Dis. 2019, 66, 1481–1492. [Google Scholar] [CrossRef] [PubMed]
- Sattar, A.; Zakaria, Z.; Abu, J.; Aziz, S.A.; Rojas-Ponce, G. Isolation of Mycobacterium avium and other nontuberculous mycobacteria in chickens and captive birds in peninsular Malaysia. BMC Vet. Res. 2021, 17, 13. [Google Scholar] [CrossRef]
- Ding, T.; Suo, Y.; Xiang, Q.; Zhao, X.; Chen, S.; Ye, X.; Liu, D. Significance of viable but nonculturable Escherichia coli: Induction, detection, and control. J. Microbiol. Biotechnol. 2017, 27, 417–428. [Google Scholar] [CrossRef]
- Jackson, D.N.; Davis, B.; Tirado, S.M.; Duggal, M.; van Frankenhuyzen, J.K.; Deaville, D.; Wijesinghe, M.A.K.; Tessaro, M.; Trevors, J.T. Survival mechanisms and culturability of Campylobacter jejuni under stress conditions. Antonie Van Leeuwenhoek 2009, 96, 377–394. [Google Scholar] [CrossRef] [PubMed]
- Zahra, M.; Ferreri, M.; Alkasir, R.; Yin, J.; Han, B.; Su, J. Isolation and characterization of small-colony variants of Ornithobacterium rhinotracheale. J. Clin. Microbiol. 2013, 51, 3228–3236. [Google Scholar] [CrossRef]
- Falconi-Agapito, F.; Saravia, L.E.; Flores-Pérez, A.; Fernández-Díaz, M. Naturally occurring β-nicotinamide adenine dinucleotide-independent Avibacterium paragallinarum isolate in Peru. Avian Dis. 2015, 59, 341–343. [Google Scholar] [CrossRef] [PubMed]
- Dousse, F.; Thomann, A.; Brodard, I.; Korczak, B.M.; Schlatter, Y.; Kuhnert, P.; Miserez, R.; Frey, J. Routine phenotypic identification of bacterial species of the family Pasteurellaceae isolated from animals. J. Vet. Diagn. Investig. 2008, 20, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Hunt Gerardo, S.; Citron, D.M.; Claros, M.C.; Fernandez, H.T.; Goldstein, E.J. Pasteurella multocida subsp. multocida and P. multocida subsp. septica differentiation by PCR fingerprinting and alpha-glucosidase activity. J. Clin. Microbiol. 2001, 39, 2558–2564. [Google Scholar] [CrossRef]
- Bisgaard, M.; Korczak, B.M.; Busse, H.-J.; Kuhnert, P.; Bojesen, A.M.; Christensen, H. Classification of the taxon 2 and taxon 3 complex of Bisgaard within Gallibacterium and description of Gallibacterium melopsittaci sp. nov., Gallibacterium trehalosifermentans sp. nov. and Gallibacterium salpingitidis sp. nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 735–744. [Google Scholar] [CrossRef] [PubMed]
- Christensen, H.; Bisgaard, M. Phylogenetic relationships of Riemerella anatipestifer serovars and related taxa and an evaluation of specific PCR tests reported for R. anatipestifer. J. Appl. Microbiol. 2010, 108, 1612–1619. [Google Scholar] [CrossRef]
- van Empel, P.C.; Hafez, H.M. Ornithobacterium rhinotracheale: A review. Avian Pathol. 1999, 28, 217–227. [Google Scholar] [CrossRef]
- Paudel, S.; Stessl, B.; Hess, C.; Zloch, A.; Hess, M. High genetic diversity among extraintestinal Escherichia coli isolates in pullets and layers revealed by a longitudinal study. BMC Vet. Res. 2016, 12, 221. [Google Scholar] [CrossRef]
- Smith, E.; Miller, E.; Aguayo, J.M.; Figueroa, C.F.; Nezworski, J.; Studniski, M.; Wileman, B.; Johnson, T. Genomic diversity and molecular epidemiology of Pasteurella multocida. PLoS ONE 2021, 16, e0249138. [Google Scholar] [CrossRef]
- Omaleki, L.; Blackall, P.J.; Bisgaard, M.; Turni, C. Molecular and serological characterization of Riemerella isolates associated with poultry in Australia. Avian Pathol. 2021, 50, 31–40. [Google Scholar] [CrossRef]
- Peña-Vargas, E.R.; Vega-Sánchez, V.; Morales-Erasto, V.; Trujillo-Ruíz, H.H.; Talavera-Rojas, M.; Soriano-Vargas, E. Serotyping, genotyping, and antimicrobial susceptibility of Ornithobacterium rhinotracheale isolates from Mexico. Avian Dis. 2016, 60, 669–672. [Google Scholar] [CrossRef]
- Roth, N.; Käsbohrer, A.; Mayrhofer, S.; Zitz, U.; Hofacre, C.; Domig, K.J. The application of antibiotics in broiler production and the resulting antibiotic resistance in Escherichia coli: A global overview. Poult. Sci. 2019, 98, 1791–1804. [Google Scholar] [CrossRef]
- Hess, C.; Grafl, B.; Bagheri, S.; Kaesbohrer, A.; Zloch, A.; Hess, M. Antimicrobial resistance profiling of Gallibacterium anatis from layers reveals high number of multiresistant strains and substantial variability even between isolates from the same organ. Microb. Drug Resist. 2020, 26, 169–177. [Google Scholar] [CrossRef]
- Oikarainen, P.E.; Pohjola, L.K.; Pietola, E.S.; Heikinheimo, A. Direct vertical transmission of ESBL/pAmpC-producing Escherichia coli limited in poultry production pyramid. Vet. Microbiol. 2019, 231, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Benrabia, I.; Hamdi, T.M.; Shehata, A.A.; Neubauer, H.; Wareth, G. Methicillin-resistant Staphylococcus aureus (MRSA) in poultry species in Algeria: Long-term study on prevalence and antimicrobial resistance. Vet. Sci. 2020, 7, 54. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Benkova, M.; Soukup, O.; Marek, J. Antimicrobial susceptibility testing: Currently used methods and devices and the near future in clinical practice. J. Appl. Microbiol. 2020, 129, 806–822. [Google Scholar] [CrossRef] [PubMed]
- EUCAST. The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 13.0. 2023. Available online: http://www.eucast.org (accessed on 21 April 2023).
- CLSI. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 5th ed.; CLSI Supplement VET01S; Clinical and Laboratory Standards Institute: Berwyn, PA, USA, 2020. [Google Scholar]
- Hess, C.; Alispahic, M.; Hess, M. Application of MALDI-TOF MS in Veterinary and Food Microbiology. In MALDI-TOF Mass Spectrometry in Microbiology; Caister Academic Press: Poole, UK, 2016; pp. 109–126. ISBN 9781910190418. [Google Scholar]
- Austin, B. The value of cultures to modern microbiology. Antonie Van Leeuwenhoek 2017, 110, 1247–1256. [Google Scholar] [CrossRef]
- OIE Terrestrial Manual. Avian Mycoplasmosis (Mycoplasma gallisepticum, M. synoviae) Chapter 3.3.5. 2018. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.03.05_%20AVIAN_MYCO.pdf (accessed on 8 March 2022).
- OIE Terrestrial Manual. Avian Chlamydiosis. Chapter 2.3.1. 2018. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/2.03.01_AVIAN_CHLAMYD.pdf (accessed on 8 March 2022).
- Wang, C.; Robles, F.; Ramirez, S.; Riber, A.B.; Bojesen, A.M. Culture-independent identification and quantification of Gallibacterium anatis (G. anatis) by real-time quantitative PCR. Avian Pathol. 2016, 45, 538–544. [Google Scholar] [CrossRef]
- Kuchipudi, S.V.; Yon, M.; Surendran Nair, M.; Byukusenge, M.; Barry, R.M.; Nissly, R.H.; Williams, J.; Pierre, T.; Mathews, T.; Walner-Pendleton, E.; et al. A highly sensitive and specific probe-based real-time PCR for the detection of Avibacterium paragallinarum in clinical samples from poultry. Front. Vet. Sci. 2021, 8, 609126. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.; Patel, V.; Hickey, T.; Maguire, C.; Greig, D.R.; Lee, W.; Godbole, G.; Grant, K.; Chattaway, M.A. Real-time PCR assay for differentiation of typhoidal and nontyphoidal Salmonella. J. Clin. Microbiol. 2019, 57, e00167-19. [Google Scholar] [CrossRef]
- Ewers, C.; Janssen, T.; Kiessling, S.; Philipp, H.-C.; Wieler, L.H. Rapid detection of virulence-associated genes in avian pathogenic Escherichia coli by multiplex polymerase chain reaction. Avian Dis. 2005, 49, 269–273. [Google Scholar] [CrossRef]
- Lindberg, A.; Skarin, H.; Knutsson, R.; Blomqvist, G.; Båverud, V. Real-time PCR for Clostridium botulinum type C neurotoxin (BoNTC) gene, also covering a chimeric C/D sequence--application on outbreaks of botulism in poultry. Vet. Microbiol. 2010, 146, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, I.M.; Hsieh, Y.-H.; Simpson, S. Species identification of Campylobacter jejuni and Campylobacter coli isolates from raw poultry products by MALDI-TOF MS and rRNA sequence analysis. J. AOAC Int. 2020, 103, 197–204. [Google Scholar] [CrossRef]
- Luna-Castrejón, L.P.; Buter, R.; Pantoja-Nuñez, G.I.; Acuña-Yanes, M.; Ceballos-Valenzuela, K.; Talavera-Rojas, M.; Salgado-Miranda, C.; Heuvelink, A.; de Wit, S.; Soriano-Vargas, E.; et al. Identification, HPG2 sequence analysis, and antimicrobial susceptibility of Avibacterium paragallinarum isolates obtained from outbreaks of infectious coryza in commercial layers in Sonora State, Mexico. Avian Dis. 2021, 65, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Palmieri, N.; Hess, C.; Hess, M.; Alispahic, M. Sequencing of five poultry strains elucidates phylogenetic relationships and divergence in virulence genes in Morganella morganii. BMC Genom. 2020, 21, 579. [Google Scholar] [CrossRef] [PubMed]
- Alispahic, M.; Endler, L.; Hess, M.; Hess, C. Ornithobacterium rhinotracheale: MALDI-TOF MS and whole genome sequencing confirm that serotypes K, L and M deviate from well-known reference strains and numerous field isolates. Microorganisms 2021, 9, 1006. [Google Scholar] [CrossRef]
- Chen, X.; Liu, W.; Li, H.; Yan, S.; Jiang, F.; Cai, W.; Li, G. Whole genome sequencing analysis of avian pathogenic Escherichia coli from China. Vet. Microbiol. 2021, 259, 109158. [Google Scholar] [CrossRef]
- Kürekci, C.; Sahin, S.; Iwan, E.; Kwit, R.; Bomba, A.; Wasyl, D. Whole-genome sequence analysis of Salmonella Infantis isolated from raw chicken meat samples and insights into pESI-like megaplasmid. Int. J. Food Microbiol. 2021, 337, 108956. [Google Scholar] [CrossRef] [PubMed]
- Kunert-Filho, H.C.; Furian, T.Q.; Sesterhenn, R.; Chitolina, G.Z.; Willsmann, D.E.; Borges, K.A.; Salle, C.T.P.; Moraes, H.L.d.S.; do Nascimento, V.P. Bacterial community identification in poultry carcasses using high-throughput next generation sequencing. Int. J. Food Microbiol. 2022, 364, 109533. [Google Scholar] [CrossRef]
- Di Francesco, C.E.; Smoglica, C.; Profeta, F.; Farooq, M.; Di Giannatale, E.; Toscani, T.; Marsilio, F. Research Note: Detection of antibiotic-resistance genes in commercial poultry and turkey flocks from Italy. Poult. Sci. 2021, 100, 101084. [Google Scholar] [CrossRef]
- Juricova, H.; Matiasovicova, J.; Kubasova, T.; Cejkova, D.; Rychlik, I. The distribution of antibiotic resistance genes in chicken gut microbiota commensals. Sci. Rep. 2021, 11, 3290. [Google Scholar] [CrossRef]
- Savin, M.; Alexander, J.; Bierbaum, G.; Hammerl, J.A.; Hembach, N.; Schwartz, T.; Schmithausen, R.M.; Sib, E.; Voigt, A.; Kreyenschmidt, J. Antibiotic-resistant bacteria, antibiotic resistance genes, and antibiotic residues in wastewater from a poultry slaughterhouse after conventional and advanced treatments. Sci. Rep. 2021, 11, 16622. [Google Scholar] [CrossRef]
- Wang, X.-R.; Lian, X.-L.; Su, T.-T.; Long, T.-F.; Li, M.-Y.; Feng, X.-Y.; Sun, R.-Y.; Cui, Z.-H.; Tang, T.; Xia, J.; et al. Duck wastes as a potential reservoir of novel antibiotic resistance genes. Sci. Total Environ. 2021, 771, 144828. [Google Scholar] [CrossRef]
- Zhu, T.; Chen, T.; Cao, Z.; Zhong, S.; Wen, X.; Mi, J.; Ma, B.; Zou, Y.; Zhang, N.; Liao, X.; et al. Antibiotic resistance genes in layer farms and their correlation with environmental samples. Poult. Sci. 2021, 100, 101485. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Rolain, J.-M. Use of MALDI-TOF mass spectrometry for identification of bacteria that are difficult to culture. J. Microbiol. Methods 2013, 92, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Alispahic, M.; Christensen, H.; Hess, C.; Razzazi-Fazeli, E.; Bisgaard, M.; Hess, M. Identification of Gallibacterium species by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry evaluated by multilocus sequence analysis. Int. J. Med. Microbiol. 2011, 301, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Rubbenstroth, D.; Ryll, M.; Hotzel, H.; Christensen, H.; Knobloch, J.K.-M.; Rautenschlein, S.; Bisgaard, M. Description of Riemerella columbipharyngis sp. nov., isolated from the pharynx of healthy domestic pigeons (Columba livia f. domestica), and emended descriptions of the genus Riemerella, Riemerella anatipestifer and Riemerella columbina. Int. J. Syst. Evol. Microbiol. 2013, 63, 280–287. [Google Scholar] [CrossRef]
- Spergser, J.; Hess, C.; Loncaric, I.; Ramírez, A.S. Matrix-assisted laser desorption ionization-time of flight mass spectrometry is a superior diagnostic tool for the identification and differentiation of Mycoplasmas isolated from animals. J. Clin. Microbiol. 2019, 57, e00316-19. [Google Scholar] [CrossRef]
- Salah, H.; Kolecka, A.; Rozaliyani, A.; Wahyuningsih, R.; Taj-Aldeen, S.J.; Boekhout, T.; Houbraken, J. A new filter based cultivation approach for improving Aspergillus identification using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS). Mycopathologia 2022, 187, 39–52. [Google Scholar] [CrossRef]
- Panda, A.; Ghosh, A.K.; Mirdha, B.R.; Xess, I.; Paul, S.; Samantaray, J.C.; Srinivasan, A.; Khalil, S.; Rastogi, N.; Dabas, Y. MALDI-TOF mass spectrometry for rapid identification of clinical fungal isolates based on ribosomal protein biomarkers. J. Microbiol. Methods 2015, 109, 93–105. [Google Scholar] [CrossRef]
- Alispahic, M.; Christensen, H.; Bisgaard, M.; Hess, M.; Hess, C. MALDI-TOF mass spectrometry confirms difficulties in separating species of the Avibacterium genus. Avian Pathol. 2014, 43, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Alispahic, M.; Christensen, H.; Hess, C.; Razzazi-Fazeli, E.; Bisgaard, M.; Hess, M. MALDI-TOF mass spectrometry confirms clonal lineages of Gallibacterium anatis between chicken flocks. Vet. Microbiol. 2012, 160, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Siegrist, T.J.; Anderson, P.D.; Huen, W.H.; Kleinheinz, G.T.; McDermott, C.M.; Sandrin, T.R. Discrimination and characterization of environmental strains of Escherichia coli by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF-MS). J. Microbiol. Methods 2007, 68, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Hrabák, J.; Chudácková, E.; Walková, R. Matrix-assisted laser desorption ionization-time of flight (maldi-tof) mass spectrometry for detection of antibiotic resistance mechanisms: From research to routine diagnosis. Clin. Microbiol. Rev. 2013, 26, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Lozica, L.; Kazazić, S.P.; Gottstein, Ž. High phylogenetic diversity of Gallibacterium anatis is correlated with low biosecurity measures and management practices on poultry farms. Avian Pathol. 2020, 49, 467–475. [Google Scholar] [CrossRef]
- Jung, A.; Metzner, M.; Ryll, M. Comparison of pathogenic and non-pathogenic Enterococcus cecorum strains from different animal species. BMC Microbiol. 2017, 17, 33. [Google Scholar] [CrossRef]
- Oakeson, K.F.; Wagner, J.M.; Rohrwasser, A.; Atkinson-Dunn, R. Whole-genome sequencing and bioinformatic analysis of isolates from foodborne illness outbreaks of Campylobacter jejuni and Salmonella enterica. J. Clin. Microbiol. 2018, 56, e00161-18. [Google Scholar] [CrossRef]
- Whitehouse, C.A.; Young, S.; Li, C.; Hsu, C.-H.; Martin, G.; Zhao, S. Use of whole-genome sequencing for Campylobacter surveillance from NARMS retail poultry in the United States in 2015. Food Microbiol. 2018, 73, 122–128. [Google Scholar] [CrossRef]
- Chukiatsiri, K.; Sasipreeyajan, J.; Neramitmansuk, W.; Chansiripornchai, N. Efficacy of autogenous killed vaccine of Avibacterium paragallinarum. Avian Dis. 2009, 53, 382–386. [Google Scholar] [CrossRef]
- Li, L.; Thøfner, I.; Christensen, J.P.; Ronco, T.; Pedersen, K.; Olsen, R.H. Evaluation of the efficacy of an autogenous Escherichia coli vaccine in broiler breeders. Avian Pathol. 2017, 46, 300–308. [Google Scholar] [CrossRef]
- Gallardo, R.A.; Da Silva, A.P.; Egaña-Labrin, S.; Stoute, S.; Kern, C.; Zhou, H.; Cutler, G.; Corsiglia, C. Infectious Coryza: Persistence, Genotyping, and Vaccine Testing. Avian Dis. 2020, 64, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Lozica, L.; Kabalin, A.E.; Dolenčić, N.; Vlahek, M.; Gottstein, Ž. Phylogenetic characterization of avian pathogenic Escherichia coli strains longitudinally isolated from broiler breeder flocks vaccinated with autogenous vaccine. alispahic. Poult. Sci. 2021, 100, 101079. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, J. Parasites of poultry. In Parasitic Infections of Domestic Animals; Softcover Reprint of the Original 1st Edition 1996; Springer: Basel, Switzerland, 2012; pp. 337–393. ISBN 9783034876681. [Google Scholar]
- Hinkle, N.C.; Corrigan, R.M. External Parasites and Poultry Pests. In Diseases of Poultry, 14th ed.; Swayne, D.E., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 1137–1156. [Google Scholar]
- Decru, E.; Mul, M.; Nisbet, A.J.; Vargas Navarro, A.H.; Chiron, G.; Walton, J.; Norton, T.; Roy, L.; Sleeckx, N. Possibilities for IPM strategies in European laying hen farms for improved control of the poultry red mite (Dermanyssus gallinae): Details and state of affairs. Front. Vet. Sci. 2020, 7, 565866. [Google Scholar] [CrossRef] [PubMed]
- McDougald, L.R. Internal parasites. In Diseases of Poultry, 14th ed.; Swayne, D.E., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 1157–1191. [Google Scholar]
- Hess, M.; McDougald, L.R. Histomoniasis (histomonosis, blackhead disease). In Diseases of Poultry, 14th ed.; Swayne, D.E., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 1223–1230. [Google Scholar]
- Shifaw, A.; Feyera, T.; Walkden-Brown, S.W.; Sharpe, B.; Elliott, T.; Ruhnke, I. Global and regional prevalence of helminth infection in chickens over time: A systematic review and meta-analysis. Poult. Sci. 2021, 100, 101082. [Google Scholar] [CrossRef]
- Zloch, A.; Kuchling, S.; Hess, M.; Hess, C. Influence of alternative husbandry systems on postmortem findings and prevalence of important bacteria and parasites in layers monitored from end of rearing until slaughter. Vet. Rec. 2018, 182, 350. [Google Scholar] [CrossRef]
- Jung, L.; Brenninkmeyer, C.; Niebuhr, K.; Bestman, M.; Tuyttens, F.A.M.; Gunnarsson, S.; Sørensen, J.T.; Ferrari, P.; Knierim, U. Husbandry conditions and welfare outcomes in organic egg production in eight European countries. Animals 2020, 10, 2102. [Google Scholar] [CrossRef]
- Taylor, M.A.; Coop, R.L.; Wall, R.L. Laboratory diagnosis of parasitism. In Veterinary Parasitology; Taylor, M.A., Coop, R.L., Wall, R.L., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 259–312. ISBN 9781119073680. [Google Scholar]
- Zloch, A.; Kuchling, S.; Hess, M.; Hess, C. In addition to birds’ age and outdoor access, the detection method is of high importance to determine the prevalence of gastrointestinal helminths in laying hens kept in alternative husbandry systems. Vet. Parasitol. 2021, 299, 109559. [Google Scholar] [CrossRef]
- Daş, G.; Klauser, S.; Stehr, M.; Tuchscherer, A.; Metges, C.C. Accuracy and precision of McMaster and Mini-FLOTAC egg counting techniques using egg-spiked faeces of chickens and two different flotation fluids. Vet. Parasitol. 2020, 283, 109158. [Google Scholar] [CrossRef]
- Blake, D.; Liebhart, D. Advances in understanding parasite infections of poultry: Focus on protozoa and the red mite. In Optimising Poultry Flock Health; de Witt, S., Ed.; In Press; Burleigh Dodds Science Publishing: London, UK, 2022. [Google Scholar]
- Carvalho, F.S.; Wenceslau, A.A.; Teixeira, M.; Matos Carneiro, J.A.; Melo, A.D.B.; Albuquerque, G.R. Diagnosis of Eimeria species using traditional and molecular methods in field studies. Vet. Parasitol. 2011, 176, 95–100. [Google Scholar] [CrossRef]
- Cantacessi, C.; Riddell, S.; Morris, G.M.; Doran, T.; Woods, W.G.; Otranto, D.; Gasser, R.B. Genetic characterization of three unique operational taxonomic units of Eimeria from chickens in Australia based on nuclear spacer ribosomal DNA. Vet. Parasitol. 2008, 152, 226–234. [Google Scholar] [CrossRef]
- Cupo, K.L.; Beckstead, R.B. PCR detection of Heterakis gallinarum in environmental samples. Vet. Parasitol. 2019, 271, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Sulejmanovic, T.; Turblin, V.; Bilic, I.; Jaskulska, B.; Hess, M. Detection of Histomonas meleagridis DNA in dust samples obtained from apparently healthy meat turkey flocks without effect on performance. Avian Pathol. 2019, 48, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Ahaduzzaman, M.; Keerqin, C.; Kumar, A.; Musigwa, S.; Morgan, N.; Kheravii, S.K.; Sharpe, S.; Williamson, S.; Wu, S.-B.; Walkden-Brown, S.W.; et al. Detection and Quantification of Clostridium perfringens and Eimeria spp. in Poultry Dust Using Real-Time PCR Under Experimental and Field Conditions. Avian Dis. 2021, 65, 77–85. [Google Scholar] [CrossRef]
- Grafl, B.; Liebhart, D.; Windisch, M.; Ibesich, C.; Hess, M. Seroprevalence of Histomonas meleagridis in pullets and laying hens determined by ELISA. Vet. Rec. 2011, 168, 160. [Google Scholar] [CrossRef]
- van der Heijden, H.M.J.F.; Stegeman, A.; Landman, W.J.M. Development of a blocking-ELISA for the detection of antibodies against Histomonas meleagridis in chickens and turkeys. Vet. Parasitol. 2010, 171, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Martín-Pacho, J.R.; Montoya, M.N.; Arangüena, T.; Toro, C.; Morchón, R.; Marcos-Atxutegi, C.; Simón, F. A coprological and serological survey for the prevalence of Ascaridia spp. in laying hens. J. Vet. Med. B Infect. Dis. Vet. Public Health 2005, 52, 238–242. [Google Scholar] [CrossRef]
- Kawaguchi, T.; Nomura, K.; Hirayama, Y.; Kitagawa, T. Establishment and characterization of a chicken hepatocellular carcinoma cell line, LMH. Cancer Res. 1987, 47, 4460–4464. [Google Scholar]
- Schnitzlein, W.M.; Tripathy, D.N. Replication of infectious laryngotracheitis virus in a quail cell line, QT-35. Avian Dis. 1995, 39, 528–531. [Google Scholar] [CrossRef]
- Baxendale, W.; Mebatsion, T. The isolation and characterisation of astroviruses from chickens. Avian Pathol. 2004, 33, 364–370. [Google Scholar] [CrossRef]
- Kang, K.-I.; Linnemann, E.; Icard, A.H.; Durairaj, V.; Mundt, E.; Sellers, H.S. Chicken astrovirus as an aetiological agent of runting-stunting syndrome in broiler chickens. J. Gen. Virol. 2018, 99, 512–524. [Google Scholar] [CrossRef]
- Yin, L.; Zhou, Q.; Mai, K.; Huang, J.; Yan, Z.; Wei, X.; Shen, H.; Li, Q.; Chen, L.; Zhou, Q. Isolation and characterization of a novel chicken astrovirus in China. Poult. Sci. 2021, 100, 101363. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Yang, J.; Wang, Y.; Chen, H.; Diao, Y.; Tang, Y. Isolation and characterization of a duck-origin goose astrovirus in China. Emerg. Microbes Infect. 2020, 9, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Wang, Y.; Wang, Q.; Yang, J.; Jiang, X.; He, D.; Diao, Y.; Tang, Y. The isolation and characterization of Duck astrovirus type-1 remerging in China. Transbound. Emerg. Dis. 2021, 69, 2890–2897. [Google Scholar] [CrossRef]
- Pan, Q.; Liu, L.; Gao, Y.; Liu, C.; Qi, X.; Zhang, Y.; Wang, Y.; Li, K.; Gao, L.; Wang, X.; et al. Characterization of a hypervirulent fowl adenovirus 4 with the novel genotype newly prevalent in China and establishment of reproduction infection model of hydropericardium syndrome in chickens. Poult. Sci. 2017, 96, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Niu, D.; Feng, J.; Duan, B.; Shi, Q.; Li, Y.; Chen, Z.; Ma, L.; Liu, H.; Wang, Y. Epidemiological survey of avian adenovirus in China from 2015 to 2021 and the genetic variability of highly pathogenic Fadv-4 isolates. Infect. Genet. Evol. 2022, 101, 105277. [Google Scholar] [CrossRef]
- Alexander, H.; Huber, P.; Cao, J.; Krell, P.; Nagy, É. Growth characteristics of fowl adenovirus type 8 in a chicken hepatoma cell line. J. Virol. Methods 1998, 74, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhao, J. Pathogenesis of Hypervirulent Fowl Adenovirus Serotype 4: The Contributions of Viral and Host Factors. Viruses 2019, 11, 741. [Google Scholar] [CrossRef]
- Lu, H.; Tang, Y.; Dunn, P.A.; Wallner-Pendleton, E.A.; Lin, L.; Knoll, E.A. Isolation and molecular characterization of newly emerging avian reovirus variants and novel strains in Pennsylvania, USA, 2011–2014. Sci. Rep. 2015, 5, 14727. [Google Scholar] [CrossRef]
- Yang, J.; Tian, J.; Chen, L.; Tang, Y.; Diao, Y. Isolation and genomic characterization of a novel chicken-orign orthoreovirus causing goslings hepatitis. Vet. Microbiol. 2018, 227, 69–77. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, H.; Chi, Z.; Cui, Z.; Chang, S.; Wang, Y.; Zhao, P. Isolation, identification and genome analysis of an avian hepatitis E virus from white-feathered broilers in China. Poult. Sci. 2022, 101, 101633. [Google Scholar] [CrossRef]
- Huang, F.F.; Pierson, F.W.; Toth, T.E.; Meng, X.J. Construction and characterization of infectious cDNA clones of a chicken strain of hepatitis E virus (HEV), avian HEV. J. Gen. Virol. 2005, 86, 2585–2593. [Google Scholar] [CrossRef]
- Moscovici, C.; Moscovici, M.; Jimenez, H.; Lai, M.M.; Hayman, M.J.; Vogt, P.K. Continuous tissue culture cell lines derived from chemically induced tumors of Japanese quail. Cell 1977, 11, 95–103. [Google Scholar] [CrossRef]
- Cowen, B.S.; Braune, M.O. The propagation of avian viruses in a continuous cell line (QT35) of Japanese quail origin. Avian Dis. 1988, 32, 282. [Google Scholar] [CrossRef]
- Schonewille, E.; Singh, A.; Göbel, T.W.; Gerner, W.; Saalmüller, A.; Hess, M. Fowl adenovirus (FAdV) serotype 4 causes depletion of B and T cells in lymphoid organs in specific pathogen-free chickens following experimental infection. Vet. Immunol. Immunopathol. 2008, 121, 130–139. [Google Scholar] [CrossRef]
- Cho, B.R. Cytopathic effects and focus formation by reticuloendotheliosis viruses in a quail fibroblast cell line. Avian Dis. 1983, 27, 261. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-W.; Jung, K.; Jadhao, S.J.; Suarez, D.L. Evaluation of chicken-origin (DF-1) and quail-origin (QT-6) fibroblast cell lines for replication of avian influenza viruses. J. Virol. Methods 2008, 153, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Mor, S.K.; Sharafeldin, T.A.; Porter, R.E.; Ziegler, A.; Patnayak, D.P.; Goyal, S.M. Isolation and characterization of a turkey arthritis reovirus. Avian Dis. 2013, 57, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Goyal, S.M.; Chiang, S.J.; Dar, A.M.; Nagaraja, K.V.; Shaw, D.P.; Halvorson, D.A.; Kapur, V. Isolation of avian pneumovirus from an outbreak of respiratory illness in Minnesota turkeys. J. Vet. Diagn. Investig. 2000, 12, 166–168. [Google Scholar] [CrossRef]
- Bennett, R.S.; McComb, B.; Shin, H.-J.; Njenga, M.K.; Nagaraja, K.V.; Halvorson, D.A. Detection of avian pneumovirus in wild canada geese (Branta canadensis) and blue-winged teal (Anas discors). Avian Dis. 2002, 46, 1025–1029. [Google Scholar] [CrossRef]
- Chiang, S.J.; Dar, A.; Goyal, S.M.; Nagaraja, K.V.; Halvorson, D.; Kapur, V. Isolation of avian pneumovirus in QT-35 cells. Vet. Rec. 1998, 143, 596. [Google Scholar]
- Sabara, M.I.; Larence, J.E. Evaluation of a Japanese quail fibrosarcoma cell line (QT-35) for use in the propagation and detection of metapneumovirus. J. Virol. Methods 2002, 102, 73–81. [Google Scholar] [CrossRef]
- Smith, A.L.; Tignor, G.H.; Mifune, K.; Motohashi, T. Isolation and assay of rabies serogroup viruses in CER cells. Intervirology 1977, 8, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Hafez, H.M.; Weiland, F. Isolierung des Virus der Rhinotracheitis der Puten (TRT). Tierärztliche Umsch. 1990, 45, 103–111. [Google Scholar]
- Cardoso, T.C.; Rahal, P.; Pilz, D.; Teixeira, M.C.; Arns, C.W. Replication of classical infectious bursal disease virus in the chicken embryo related cell line. Avian Pathol. 2000, 29, 213–217. [Google Scholar] [CrossRef]
- Ferreira, H.L.; Pilz, D.; Mesquita, L.G.; Cardoso, T. Infectious bronchitis virus replication in the chicken embryo related cell line. Avian Pathol. 2003, 32, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Himly, M.; Foster, D.N.; Bottoli, I.; Iacovoni, J.S.; Vogt, P.K. The DF-1 chicken fibroblast cell line: Transformation induced by diverse oncogenes and cell death resulting from infection by avian leukosis viruses. Virology 1998, 248, 295–304. [Google Scholar] [CrossRef]
- Schaefer-Klein, J.; Givol, I.; Barsov, E.V.; Whitcomb, J.M.; VanBrocklin, M.; Foster, D.N.; Federspiel, M.J.; Hughes, S.H. The EV-O-derived cell line DF-1 supports the efficient replication of avian leukosis-sarcoma viruses and vectors. Virology 1998, 248, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Liu, Y.; Zou, C.; Tan, Y.; Han, Z.; Xue, C.; Cao, Y. A highly pathogenic recombinant infectious bronchitis virus with adaptability in cultured cells. Virus Res. 2021, 292, 198229. [Google Scholar] [CrossRef] [PubMed]
- Anam, S.; Rahman, S.U.; Ali, S.; Saeed, M.; Goyal, S.M. Comparative growth kinetic study of Newcastle disease virus, infectious bursal disease virus and avian influenza virus in chicken embryo fibroblast and DF-1 cell lines. Pol. J. Vet. Sci. 2021, 24, 287–292. [Google Scholar] [CrossRef]
- Buys, S.B.; Du Preez, J.H.; Els, H.J. The isolation and attenuation of a virus causing rhinotracheitis in turkeys in South Africa. Onderstepoort J. Vet. Res. 1989, 56, 87–98. [Google Scholar]
- Cook, J.K.A.; Cavanagh, D. Detection and differentiation of avian pneumoviruses (metapneumoviruses). Avian Pathol. 2002, 31, 117–132. [Google Scholar] [CrossRef] [PubMed]
- Patnayak, D.P.; Tiwari, A.; Goyal, S.M. Growth of vaccine strains of avian pneumovirus in different cell lines. Avian Pathol. 2005, 34, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Kwon, J.-S.; Lee, H.-J.; Jeong, S.-H.; Park, J.-Y.; Hong, Y.-H.; Lee, Y.-J.; Youn, H.-S.; Lee, D.-W.; Do, S.-H.; Park, S.-Y.; et al. Isolation and characterization of avian metapneumovirus from chickens in Korea. J. Vet. Sci. 2010, 11, 59–66. [Google Scholar] [CrossRef]
- Alonso-Caplen, F.V.; Matsuoka, Y.; Wilcox, G.E.; Compans, R.W. Replication and morphogenesis of avian coronavirus in Vero cells and their inhibition by monensin. Virus Res. 1984, 1, 153–167. [Google Scholar] [CrossRef]
- Kang, S.Y.; Nagaraja, K.V.; Newman, J.A. Primary isolation and identification of avian rotaviruses from turkeys exhibiting signs of clinical enteritis in a continuous MA 104 cell line. Avian Dis. 1986, 30, 494–499. [Google Scholar] [CrossRef]
- Reina, J.; Fernandez-Baca, V.; Blanco, I.; Munar, M. Comparison of Madin-Darby canine kidney cells (MDCK) with a green monkey continuous cell line (Vero) and human lung embryonated cells (MRC-5) in the isolation of influenza A virus from nasopharyngeal aspirates by shell vial culture. J. Clin. Microbiol. 1997, 35, 1900–1901. [Google Scholar] [CrossRef]
- OIE World Organisation for Animal Health. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals Chapter 3.3.4: Chapter 3.3.4 Avian Influenza (Including Infection with High Pathogencity Avian Infleunza Viruses); OIE World Organisation for Animal Health: Paris, France, 2021. [Google Scholar]
- Davidson, I.; Grinberg, R.; Malkinson, M.; Mechani, S.; Pokamunski, S.; Weisman, Y. Diagnosis of turkey meningoencephalitis virus infection in field cases by RT-PCR compared to virus isolation in embryonated eggs and suckling mice. Avian Pathol. 2000, 29, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Nazerian, K.; Fadly, A.M. Propagation of virulent and avirulent turkey hemorrhagic enteritis virus in cell culture. Avian Dis. 1982, 26, 816–827. [Google Scholar] [CrossRef]
- Yuasa, N. Propagation and infectivity titration of the Gifu-1 strain of chicken anemia agent in a cell line (MDCC-MSB1) derived from Marek’s disease lymphoma. Natl. Inst. Anim. Health Q. 1983, 23, 13–20. [Google Scholar]
- Kong, B.-W.; Foster, L.K.; Foster, D.N. Establishment of an immortal turkey turbinate cell line suitable for avian metapneumovirus propagation. Virus Res. 2007, 127, 106–115. [Google Scholar] [CrossRef]
- Kisary, J. Experimental infection of chicken embryos and day-old chickens with parvovirus of chicken origin. Avian Pathol. 1985, 14, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Guy, J.S.; Barnes, H.J.; Smith, L.; Owen, R.; Fuller, F.J. Partial characterization of an adenovirus-like virus isolated from broiler chickens with transmissible viral proventriculitis. Avian Dis. 2005, 49, 344–351. [Google Scholar] [CrossRef] [PubMed]
- McNulty, M.S.; Allan, G.M.; Todd, D.; McFerran, J.B.; McCracken, R.M. Isolation from chickens of a rotavirus lacking the rotavirus group antigen. J. Gen. Virol. 1981, 55, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Takase, K.; Nonaka, F.; Sakaguchi, M.; Yamada, S. Cytopathic avian rotavirus isolated from duck faeces in chicken kidney cell cultures. Avian Pathol. 1986, 15, 719–730. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Wu, Y.; Zhang, W.; Merits, A.; Simmonds, P.; Wang, M.; Jia, R.; Zhu, D.; Liu, M.; Zhao, X.; et al. The first nonmammalian pegivirus demonstrates efficient in vitro replication and high lymphotropism. J. Virol. 2020, 94, e01150-20. [Google Scholar] [CrossRef]
- Wannaratana, S.; Thontiravong, A.; Pakpinyo, S. Comparison of three filter paper-based devices for safety and stability of viral sample collection in poultry. Avian Pathol. 2021, 50, 78–84. [Google Scholar] [CrossRef]
- Maw, M.T.; Yamaguchi, T.; Kasanga, C.J.; Terasaki, K.; Fukushi, H. A practical tissue sampling method using ordinary paper for molecular detection of infectious bursal disease virus RNA by RT-PCR. Avian Dis. 2006, 50, 556–560. [Google Scholar] [CrossRef]
- Rajendram, D.; Ayenza, R.; Holder, F.M.; Moran, B.; Long, T.; Shah, H.N. Long-term storage and safe retrieval of DNA from microorganisms for molecular analysis using FTA matrix cards. J. Microbiol. Methods 2006, 67, 582–592. [Google Scholar] [CrossRef]
- OIE Terrestrial Manual. Manual for Diagnostic Tests and Vaccines for Terrestrial Animals 2021, Section 3.3. 2021. Available online: https://www.woah.org/fileadmin/Home/eng/Health_standards/tahm/A_summry.htm (accessed on 21 April 2023).
- Ferguson-Noel. Mycoplasmosis. In Diseases of Poultry, 14th ed.; Swayne, D.E., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 907–965. [Google Scholar]
- Navarro, E.; Serrano-Heras, G.; Castaño, M.J.; Solera, J. Real-time PCR detection chemistry. Clin. Chim. Acta 2015, 439, 231–250. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Patrick, K.; Li, M.; Gong, J.; Xu, B.; Shen, Q.; Yang, Y.; Wei, L.; Zhang, Y.; et al. Hydroxymethylbilane synthase (HMBS) gene-based endogenous internal control for avian species. AMB Express 2020, 10, 181. [Google Scholar] [CrossRef]
- Hoffmann, B.; Depner, K.; Schirrmeier, H.; Beer, M. A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J. Virol. Methods 2006, 136, 200–209. [Google Scholar] [CrossRef] [PubMed]
- ISO/IEC 17025:2017; General Requirements for the Competence of Testing and Calibration Laboratories. ISO: Geneva, Switzerland, 2017.
- Matos, M.; Bilic, I.; Palmieri, N.; Mitsch, P.; Sommer, F.; Tvarogová, J.; Liebhart, D.; Hess, M. Epidemic of cutaneous fowlpox in a naïve population of chickens and turkeys in Austria: Detailed phylogenetic analysis indicates co-evolution of fowlpox virus with reticuloendotheliosis virus. Transbound. Emerg. Dis. 2022, 69, 2913–2923. [Google Scholar] [CrossRef]
- Hauck, R.; Prusas, C.; Hafez, H.M.; Lüschow, D. Quantitative PCR as a tool to determine the reticuloendotheliosis virus-proviral load of fowl poxvirus. Avian Dis. 2009, 53, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Sprygin, A.V.; Andreychuk, D.B.; Kolotilov, A.N.; Volkov, M.S.; Runina, I.A.; Mudrak, N.S.; Borisov, A.V.; Irza, V.N.; Drygin, V.V.; Perevozchikova, N.A. Development of a duplex real-time TaqMan PCR assay with an internal control for the detection of Mycoplasma gallisepticum and Mycoplasma synoviae in clinical samples from commercial and backyard poultry. Avian Pathol. 2010, 39, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-R.; Kwon, Y.-K.; Jang, I.; Bae, Y.-C. Viral metagenomic analysis of chickens with runting-stunting syndrome in the Republic of Korea. Virol. J. 2020, 17, 53. [Google Scholar] [CrossRef]
- Neubauer, C.; Hess, M. Detection and identification of food-borne pathogens of the genera Campylobacter, Arcobacter and Helicobacter by multiplex PCR in poultry and poultry products. J. Vet. Med. B Infect. Dis. Vet. Public Health 2006, 53, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Qin, T.; Meng, F.; Chen, S.; Peng, D.; Liu, X. Development of a multiplex probe combination-based one-step real-time reverse transcription-PCR for NA subtype typing of avian influenza virus. Sci. Rep. 2017, 7, 13455. [Google Scholar] [CrossRef]
- Yang, F.; Dong, D.; Wu, D.; Zhu, L.; Liu, F.; Yao, H.; Wu, N.; Ye, C.; Wu, H. A multiplex real-time RT-PCR method for detecting H5, H7 and H9 subtype avian influenza viruses in field and clinical samples. Virus Res. 2022, 309, 198669. [Google Scholar] [CrossRef] [PubMed]
- Shiraiwa, K.; Ogawa, Y.; Nishikawa, S.; Eguchi, M.; Shimoji, Y. Identification of serovar 1a, 1b, 2, and 5 strains of Erysipelothrix rhusiopathiae by a conventional gel-based PCR. Vet. Microbiol. 2018, 225, 101–104. [Google Scholar] [CrossRef]
- Shimoji, Y.; Shiraiwa, K.; Tominaga, H.; Nishikawa, S.; Eguchi, M.; Hikono, H.; Ogawa, Y. Development of a multiplex PCR-based assay for rapid serotyping of Erysipelothrix species. J. Clin. Microbiol. 2020, 58, e00315-20. [Google Scholar] [CrossRef]
- SCIEX. Available online: www.sciex.com/products/capillary-electrophoresis/genomelab-gexp-genetic-analysis-system (accessed on 26 January 2023).
- Xie, Z.; Luo, S.; Xie, L.; Liu, J.; Pang, Y.; Deng, X.; Xie, Z.; Fan, Q.; Khan, M.I. Simultaneous typing of nine avian respiratory pathogens using a novel GeXP analyzer-based multiplex PCR assay. J. Virol. Methods 2014, 207, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Zeng, T.; Xie, Z.; Xie, L.; Deng, X.; Xie, Z.; Luo, S.; Huang, L.; Huang, J. Simultaneous detection of eight immunosuppressive chicken viruses using a GeXP analyser-based multiplex PCR assay. Virol. J. 2015, 12, 226. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-F.; Xie, Z.-X.; Xie, L.-J.; Deng, X.-W.; Xie, Z.-Q.; Luo, S.-S.; Huang, L.; Huang, J.-L.; Zeng, T.-T. GeXP analyzer-based multiplex reverse-transcription PCR assay for the simultaneous detection and differentiation of eleven duck viruses. BMC Microbiol. 2015, 15, 247. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xie, Z.; Xie, Z.; Liu, J.; Xie, L.; Deng, X.; Luo, S.; Fan, Q.; Huang, L.; Huang, J.; et al. Simultaneous detection of eight avian influenza A virus subtypes by multiplex reverse transcription-PCR using a GeXP analyser. Sci. Rep. 2018, 8, 6183. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Xie, Z.; Huang, J.; Xie, Z.; Xie, L.; Zhang, M.; Li, M.; Wang, S.; Li, D.; Zeng, T.; et al. Simultaneous differentiation of the N1 to N9 neuraminidase subtypes of avian influenza virus by a GeXP analyzer-based multiplex reverse transcription PCR assay. Front. Microbiol. 2019, 10, 1271. [Google Scholar] [CrossRef]
- Christopher-Hennings, J.; Araujo, K.P.C.; Souza, C.J.H.; Fang, Y.; Lawson, S.; Nelson, E.A.; Clement, T.; Dunn, M.; Lunney, J.K. Opportunities for bead-based multiplex assays in veterinary diagnostic laboratories. J. Vet. Diagn. Investig. 2013, 25, 671–691. [Google Scholar] [CrossRef]
- Laamiri, N.; Fällgren, P.; Zohari, S.; Ben Ali, J.; Ghram, A.; Leijon, M.; Hmila, I. Accurate detection of avian respiratory viruses by use of multiplex PCR-based luminex suspension microarray assay. J. Clin. Microbiol. 2016, 54, 2716–2725. [Google Scholar] [CrossRef]
- Periyannan Rajeswari, P.K.; Soderberg, L.M.; Yacoub, A.; Leijon, M.; Andersson Svahn, H.; Joensson, H.N. Multiple pathogen biomarker detection using an encoded bead array in droplet PCR. J. Microbiol. Methods 2017, 139, 22–28. [Google Scholar] [CrossRef]
- Cong, F.; Zhu, Y.; Liu, X.; Li, X.; Chen, M.; Huang, R.; Guo, P. Development of an xTAG-multiplex PCR array for the detection of four avian respiratory viruses. Mol. Cell. Probes 2018, 37, 1–5. [Google Scholar] [CrossRef]
- Gand, M.; Mattheus, W.; Roosens, N.; Dierick, K.; Marchal, K.; Bertrand, S.; de Keersmaecker, S.C.J. A genoserotyping system for a fast and objective identification of Salmonella serotypes commonly isolated from poultry and pork food sectors in Belgium. Food Microbiol. 2020, 91, 103534. [Google Scholar] [CrossRef]
- Gand, M.; Mattheus, W.; Roosens, N.H.C.; Dierick, K.; Marchal, K.; de Keersmaecker, S.C.J.; Bertrand, S. A multiplex oligonucleotide ligation-PCR method for the genoserotyping of common Salmonella using a liquid bead suspension assay. Food Microbiol. 2020, 87, 103394. [Google Scholar] [CrossRef]
- Baigent, S.J.; Nair, V.K.; Le Galludec, H. Real-time PCR for differential quantification of CVI988 vaccine virus and virulent strains of Marek’s disease virus. J. Virol. Methods 2016, 233, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Raviv, Z.; Callison, S.A.; Ferguson-Noel, N.; Kleven, S.H. Strain differentiating real-time PCR for Mycoplasma gallisepticum live vaccine evaluation studies. Vet. Microbiol. 2008, 129, 179–187. [Google Scholar] [CrossRef]
- Dijkman, R.; Feberwee, A.; Landman, W.J.M. Development, validation and field evaluation of a quantitative real-time PCR able to differentiate between field Mycoplasma synoviae and the MS-H-live vaccine strain. Avian Pathol. 2017, 46, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Davidson, I.; Raibstein, I.; Altory-Natour, A.; Simanov, M.; Khinich, Y. Development of duplex dual-gene and DIVA real-time RT-PCR assays and use of feathers as a non-invasive sampling method for diagnosis of Turkey Meningoencephalitis Virus. Avian Pathol. 2017, 46, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Gimeno, I.M.; Dunn, J.R.; Cortes, A.L.; El-Gohary, A.E.-G.; Silva, R.F. Detection and differentiation of CVI988 (Rispens vaccine) from other serotype 1 Marek’s disease viruses. Avian Dis. 2014, 58, 232–243. [Google Scholar] [CrossRef]
- Sulyok, K.M.; Kreizinger, Z.; Bekő, K.; Forró, B.; Marton, S.; Bányai, K.; Catania, S.; Ellis, C.; Bradbury, J.; Olaogun, O.M.; et al. Development of molecular methods for rapid differentiation of Mycoplasma gallisepticum vaccine strains from field isolates. J. Clin. Microbiol. 2019, 57, e01084-18. [Google Scholar] [CrossRef]
- Kreizinger, Z.; Sulyok, K.M.; Pásztor, A.; Erdélyi, K.; Felde, O.; Povazsán, J.; Kőrösi, L.; Gyuranecz, M. Rapid, simple and cost-effective molecular method to differentiate the temperature sensitive (ts+) MS-H vaccine strain and wild-type Mycoplasma synoviae isolates. PLoS ONE 2015, 10, e0133554. [Google Scholar] [CrossRef]
- Hess, M. Detection and differentiation of avian adenoviruses: A review. Avian Pathol. 2000, 29, 195–206. [Google Scholar] [CrossRef]
- Meulemans, G.; Couvreur, B.; Decaesstecker, M.; Boschmans, M.; van den Berg, T.P. Phylogenetic analysis of fowl adenoviruses. Avian Pathol. 2004, 33, 164–170. [Google Scholar] [CrossRef]
- Marek, A.; Günes, A.; Schulz, E.; Hess, M. Classification of fowl adenoviruses by use of phylogenetic analysis and high-resolution melting-curve analysis of the hexon L1 gene region. J. Virol. Methods 2010, 170, 147–154. [Google Scholar] [CrossRef]
- de Wit, J.J. Detection of infectious bronchitis virus. Avian Pathol. 2000, 29, 71–93. [Google Scholar] [CrossRef]
- Valastro, V.; Holmes, E.C.; Britton, P.; Fusaro, A.; Jackwood, M.W.; Cattoli, G.; Monne, I. S1 gene-based phylogeny of infectious bronchitis virus: An attempt to harmonize virus classification. Infect. Genet. Evol. 2016, 39, 349–364. [Google Scholar] [CrossRef]
- Tucciarone, C.M.; Franzo, G.; Legnardi, M.; Fortin, A.; Valastro, V.; Lazzaro, E.; Terregino, C.; Cecchinato, M. Effect of assay choice, viral concentration and operator interpretation on infectious bronchitis virus detection and characterization. Avian Pathol. 2021, 50, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, D.J.; Schat, K.A.; Michel, L.O.; de Wit, S. A proposed nomenclature for infectious bursal disease virus isolates. Avian Pathol. 2018, 47, 576–584. [Google Scholar] [CrossRef]
- Dimitrov, K.M.; Abolnik, C.; Afonso, C.L.; Albina, E.; Bahl, J.; Berg, M.; Briand, F.-X.; Brown, I.H.; Choi, K.-S.; Chvala, I.; et al. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infect. Genet. Evol. 2019, 74, 103917. [Google Scholar] [CrossRef] [PubMed]
- Athanasopoulou, K.; Boti, M.A.; Adamopoulos, P.G.; Skourou, P.C.; Scorilas, A. Third-generation sequencing: The spearhead towards the radical transformation of modern genomics. Life 2021, 12, 30. [Google Scholar] [CrossRef] [PubMed]
- Kwok, K.T.T.; Nieuwenhuijse, D.F.; Phan, M.V.T.; Koopmans, M.P.G. Virus metagenomics in farm animals: A systematic review. Viruses 2020, 12, 107. [Google Scholar] [CrossRef]
- Liais, E.; Croville, G.; Mariette, J.; Delverdier, M.; Lucas, M.-N.; Klopp, C.; Lluch, J.; Donnadieu, C.; Guy, J.S.; Corrand, L.; et al. Novel avian coronavirus and fulminating disease in guinea fowl, France. Emerg. Infect. Dis. 2014, 20, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Matos, M.; Bilic, I.; Tvarogová, J.; Palmieri, N.; Furmanek, D.; Gotowiecka, M.; Liebhart, D.; Hess, M. A novel genotype of avian hepatitis E virus identified in chickens and common pheasants (Phasianus colchicus), extending its host range. Sci. Rep. 2022, 12, 21743. [Google Scholar] [CrossRef]
- Lagan Tregaskis, P.; Devaney, R.; Smyth, V.J. The first whole genome sequence and characterisation of avian nephritis virus genotype 3. Viruses 2021, 13, 235. [Google Scholar] [CrossRef] [PubMed]
- Flint, J. Synthesis of RNA from RNA templates. In Principles of Virology, 4th ed.; Volume 1: Molecular Biology; ASM Press: Washington, DC, USA, 2015; pp. 157–184. [Google Scholar]
- Asif, K.; O’Rourke, D.; Sabir, A.J.; Shil, P.; Noormohammadi, A.H.; Marenda, M.S. Characterisation of the whole genome sequence of an avian hepatitis E virus directly from clinical specimens reveals possible recombination events between European and USA strains. Infect. Genet. Evol. 2021, 96, 105095. [Google Scholar] [CrossRef] [PubMed]
- Butt, S.L.; Taylor, T.L.; Volkening, J.D.; Dimitrov, K.M.; Williams-Coplin, D.; Lahmers, K.K.; Miller, P.J.; Rana, A.M.; Suarez, D.L.; Afonso, C.L.; et al. Rapid virulence prediction and identification of Newcastle disease virus genotypes using third-generation sequencing. Virol. J. 2018, 15, 179. [Google Scholar] [CrossRef] [PubMed]
- Read, S.J.; Burnett, D.; Fink, C.G. Molecular techniques for clinical diagnostic virology. J. Clin. Pathol. 2000, 53, 502–506. [Google Scholar] [CrossRef]
- Ibrahim, B.; McMahon, D.P.; Hufsky, F.; Beer, M.; Deng, L.; Le Mercier, P.; Palmarini, M.; Thiel, V.; Marz, M. A new era of virus bioinformatics. Virus Res. 2018, 251, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Briand, F.-X.; Schmitz, A.; Ogor, K.; Le Prioux, A.; Guillou-Cloarec, C.; Guillemoto, C.; Allée, C.; Le Bras, M.-O.; Hirchaud, E.; Quenault, H.; et al. Emerging highly pathogenic H5 avian influenza viruses in France during winter 2015/16: Phylogenetic analyses and markers for zoonotic potential. Euro. Surveill. 2017, 22, 30473. [Google Scholar] [CrossRef]
- Reddy, V.R.A.P.; Theuns, S.; Roukaerts, I.D.M.; Zeller, M.; Matthijnssens, J.; Nauwynck, H.J. Genetic characterization of the Belgian nephropathogenic infectious bronchitis virus (NIBV) reference strain B1648. Viruses 2015, 7, 4488–4506. [Google Scholar] [CrossRef]
- Djikeng, A.; Halpin, R.; Kuzmickas, R.; Depasse, J.; Feldblyum, J.; Sengamalay, N.; Afonso, C.; Zhang, X.; Anderson, N.G.; Ghedin, E.; et al. Viral genome sequencing by random priming methods. BMC Genom. 2008, 9, 5. [Google Scholar] [CrossRef]
- Dimitrov, K.M.; Sharma, P.; Volkening, J.D.; Goraichuk, I.V.; Wajid, A.; Rehmani, S.F.; Basharat, A.; Shittu, I.; Joannis, T.M.; Miller, P.J.; et al. A robust and cost-effective approach to sequence and analyze complete genomes of small RNA viruses. Virol. J. 2017, 14, 72. [Google Scholar] [CrossRef]
- Flageul, A.; Courtillon, C.; Allée, C.; Leroux, A.; Blanchard, Y.; Deleforterie, Y.; Grasland, B.; Brown, P.A. Extracting turkey coronaviruses from the intestinal lumen of infected turkey embryos yields full genome data with good coverage by NGS. Avian Pathol. 2022, 51, 291–294. [Google Scholar] [CrossRef]
- Kubacki, J.; Fraefel, C.; Bachofen, C. Implementation of next-generation sequencing for virus identification in veterinary diagnostic laboratories. J. Vet. Diagn. Investig. 2021, 33, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Burns, R.B. Specific antibody production against a soluble antigen in the Harderian gland of the domestic fowl. Clin. Exp. Immunol. 1976, 26, 371–374. [Google Scholar]
- Aitken, I.D.; Parry, S.H. Local immunity in the respiratory tract of the chicken: I. Transudation of circulationg antibody in normal and virus-infected birds. Immunology 1976, 31, 33. [Google Scholar]
- EFSA. Assessment of listing and categorization of animal diseases within the framework of the Animal Health Law (Regulation (EU) No 2016/429): Avian mycoplasmosis (Mycoplasma gallisepticum, M. meleagridis). Sci. Opin. 2017, 15, e04953. [Google Scholar] [CrossRef]
- EU. Commission Implementing Regulation (EU) 2018/1882 of 3 December 2018 on the Application of Certain Disease Prevention and Control Rules to Categories of Listed Diseases and Establishing a List of Species and Groups of Species Posing a Considerable Risk for the Spread of Those Listed Diseases; Official Journal of the European Union; European Union: Brussels, Belgium, 2018. [Google Scholar]
- EU. Commission Decision 2011/214/EU of 1 April 2011 Amending Annexes II to IV to Council Directive 2009/158/EC on Animal Health Conditions Governing Intra-Community Trade in, and Imports from Third Countries of, Poultry and Hatching Eggs; Official Journal of the European Communities, L90, 06.04.2011, 27–49; European Union: Brussels, Belgium, 2011. [Google Scholar]
- EU. Commission Decision 2010/367/EU of 25 June 2010 on the Implementation by Member States of Surveillance Programmes for Avian Influenza in Poultry and Wild Birds; Official Journal of the European Union, L 166, 01.07.2010, 22–32; European Union: Brussels, Belgium, 2010. [Google Scholar]
- Glisson, J.R.; Dawe, J.F.; Kleven, S.H. The effect of oil-emulsion vaccines on the occurrence of nonspecific plate agglutination reactions for Mycoplasma gallisepticum and M. synoviae. Avian Dis. 1984, 28, 397–405. [Google Scholar] [CrossRef]
- Yoder, H.W. Nonspecific reactions to Mycoplasma serum plate antigens induced by inactivated poultry disease vaccines. Avian Dis. 1989, 33, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Cummins, D.R.; Reynolds, D.L. Use of an avidin-biotin enhanced dot-immunobinding assay to detect antibodies for avian mycoplasma in sera from Iowa market turkeys. Avian Dis. 1990, 34, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Ben Abdelmoumen, B.; Roy, R.S. Antigenic relatedness between seven avian mycoplasma species as revealed by western blot analysis. Avian Dis. 1995, 39, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Ewing, M.L.; Lauerman, L.H.; Kleven, S.H.; Brown, M.B. Evaluation of diagnostic procedures to detect Mycoplasma synoviae in commercial multiplier-breeder farms and commercial hatcheries in Florida. Avian Dis. 1996, 40, 798–806. [Google Scholar] [CrossRef]
- Barrow, P.A.; Freitas Neto, O.C. Pullorum disease and fowl typhoid--new thoughts on old diseases: A review. Avian Pathol. 2011, 40, 1–13. [Google Scholar] [CrossRef]
- ter Veen, C.; Feberwee, A.; Augustijn, M.; de Wit, S. High specificity of the Salmonella Pullorum/Gallinarum rapid plate agglutination test despite vaccinations against Salmonella Enteritidis and Salmonella Typhimurium. Avian Pathol. 2022, 51, 19–25. [Google Scholar] [CrossRef]
- de Wit, J.J. Gumboro disease: Estimation of optimal time of vacination by the Deventer formula. Pol. Vet. J. 1998, 3, 19–22. [Google Scholar]
- de Wit, J.J.; Heijmans, J.F.; Mekkes, D.R.; van Loon, A.A. Validation of five commercially available ELISAs for the detection of antibodies against infectious bursal disease virus (serotype 1). Avian Pathol. 2001, 30, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Godoy, A.; Icard, A.; Martinez, M.; Mashchenko, A.; García, M.; El-Attrachea, J. Detection of infectious laryngotracheitis virus antibodies by glycoprotein-specific ELISAs in chickens vaccinated with viral vector vaccines. Avian Dis. 2013, 57, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Luo, S.; Fan, Q.; Xie, L.; Liu, J.; Xie, Z.; Pang, Y.; Deng, X.; Wang, X. Detection of antibodies specific to the non-structural proteins of fowl adenoviruses in infected chickens but not in vaccinated chickens. Avian Pathol. 2013, 42, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Zhang, E.; Li, Q.; Xu, Q.; Ou, J.; Yin, H.; Li, K.; Wang, L.; Zhao, X.; Niu, X.; et al. A plant-produced recombinant fusion protein-based Newcastle disease subunit vaccine and rapid differential diagnosis platform. Vaccines 2020, 8, 122. [Google Scholar] [CrossRef]
- Bortolami, A.; Donini, M.; Marusic, C.; Lico, C.; Drissi Touzani, C.; Gobbo, F.; Mazzacan, E.; Fortin, A.; Panzarin, V.M.; Bonfante, F.; et al. Development of a novel assay based on plant-produced infectious bursal disease virus VP3 for the differentiation of infected from vaccinated animals. Front. Plant Sci. 2021, 12, 786871. [Google Scholar] [CrossRef]
- Watson, D.S.; Reddy, S.M.; Brahmakshatriya, V.; Lupiani, B. A multiplexed immunoassay for detection of antibodies against avian influenza virus. J. Immunol. Methods 2009, 340, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Lupiani, B.; Mozisek, B.; Mason, P.W.; Lamichhane, C.; Reddy, S.M. Simultaneous detection of avian influenza virus NP and H5 antibodies in chicken sera using a fluorescence microsphere immunoassay. Avian Dis. 2010, 54, 668–672. [Google Scholar] [CrossRef]
- Pinette, M.M.; Rodriguez-Lecompte, J.C.; Pasick, J.; Ojkic, D.; Leith, M.; Suderman, M.; Berhane, Y. Development of a duplex Fluorescent Microsphere Immunoassay (FMIA) for the detection of antibody responses to influenza A and newcastle disease viruses. J. Immunol. Methods 2014, 405, 167–177. [Google Scholar] [CrossRef]
- Gerber, P.F.; Trampel, D.W.; Willinghan, E.M.; Billam, P.; Meng, X.-J.; Opriessnig, T. Subclinical avian hepatitis E virus infection in layer flocks in the United States. Vet. J. 2015, 206, 304–311. [Google Scholar] [CrossRef]
- Feichtner, F.; Schachner, A.; Berger, E.; Hess, M. Fiber-based fluorescent microsphere immunoassay (FMIA) as a novel multiplex serodiagnostic tool for simultaneous detection and differentiation of all clinically relevant fowl adenovirus (FAdV) serotypes. J. Immunol. Methods 2018, 458, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Grund, C.; Beer, M.; Wang, G.; Harder, T.C. Tetraplex fluorescent microbead-based immunoassay for the serodiagnosis of Newcastle disease virus and avian influenza viruses in poultry sera. Pathogens 2022, 11, 1059. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Xie, K.; Dong, X.; Wang, L.; Huang, C.; Xu, F.; Xiao, W.; Jin, M.; Huang, B.; Tang, Y. Ultrasensitive detection of avian influenza A (H7N9) virus using surface-enhanced Raman scattering-based lateral flow immunoassay strips. Anal. Chim. Acta 2019, 1053, 139–147. [Google Scholar] [CrossRef]
- Randall, C.J.; Reece, R.L. Color Atlas of Avian Histopathology; Mosby-Wolfe: London, UK, 1996; ISBN 0723420874. [Google Scholar]
- Alturkistani, H.A.; Tashkandi, F.M.; Mohammedsaleh, Z.M. Histological stains: A literature review and case study. Glob. J. Health Sci. 2015, 8, 72–79. [Google Scholar] [CrossRef]
- Titford, M.; Bowman, B. What may the future hold for histotechnologists? Lab. Med. 2012, 43, e5–e10. [Google Scholar] [CrossRef]
- Neumann, U.; Witter, R.L. Differential diagnosis of lymphoid leukosis and Marek’s disease by tumor-associated criteria I. Studies on experimentally infected chickens. Avian Dis. 1979, 23, 417. [Google Scholar] [CrossRef]
- Witter, R.L.; Purchase, H.G.; Burgoyne, G.H. Peripheral nerve lesions similar to those of Marek’s disease in chickens inoculated with reticuloendotheliosis virus. J. Natl. Cancer Inst. 1970, 45, 567–577. [Google Scholar] [CrossRef]
- Herzog, N.K.; Bose, H.R. Expression of the oncogene of avian reticuloendotheliosis virus in Escherichia coli and identification of the transforming protein in reticuloendotheliosis virus T-transformed cells. Proc. Natl. Acad. Sci. USA 1986, 83, 812–816. [Google Scholar] [CrossRef]
- Payne, L.N.; Biggs, P.M. Studies on Marek’s disease. II. Pathogenesis. J. Natl. Cancer Inst. 1967, 39, 281–302. [Google Scholar]
- Gimeno, I.M.; Witter, R.L.; Fadly, A.M.; Silva, R.F. Novel criteria for the diagnosis of Marek’s disease virus-induced lymphomas. Avian Pathol. 2005, 34, 332–340. [Google Scholar] [CrossRef]
- Kurokawa, A.; Yamamoto, Y. Development of monoclonal antibodies specific to Marek disease virus-EcoRI-Q (Meq) for the immunohistochemical diagnosis of Marek disease using formalin-fixed, paraffin-embedded samples. J. Vet. Diagn. Investig. 2022, 34, 10406387221080444. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, H.; Mays, J.; Kiupel, M.; Dunn, J.R. Development of reliable techniques for the differential diagnosis of avian tumour viruses by immunohistochemistry and polymerase chain reaction from formalin-fixed paraffin-embedded tissue sections. Avian Pathol. 2018, 47, 364–374. [Google Scholar] [CrossRef]
- Kato, S.; Takahashi, M.; Kameyama, S.; Kamahora, J. A study of new inclusion bodies of cowpox virus. Bikens J. J. Res. 1959, 2, 93–96. [Google Scholar]
- Abdul-Aziz, T.; Fletcher, O.J.; Barnes, H.J.; Shivaprasad, H.L.; Swayne, D.E. Avian Histopathology, 4th ed.; The American Association of Avian Pathologists: Jacksonville, FL, USA, 2016; ISBN 9780978916367. [Google Scholar]
- Hess, M. Aviadenovirus Infections. In Diseases of Poultry, 14th ed.; Swayne, D.E., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 322–332. [Google Scholar]
- Rautenschlein, S.; Mahsoub, H.M.; Fitzgerald, S.D.; Pierson, F.W. Hemorrhagic enteritis and related infections. In Diseases of Poultry, 14th ed.; Swayne, D.E., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 339–347. [Google Scholar]
- Kemp, R.L.; Reid, W.M. Staining techniques for differential diagnosis of histomoniasis and mycosis in domestic poultry. Avian Dis. 1966, 10, 357. [Google Scholar] [CrossRef]
- Tyzzer, E.E. Developmental phases of the protozoon of “blackhead” in turkeys. J. Med. Res. 1919, 40, 1–30.3. [Google Scholar]
- Liebhart, D.; Weissenböck, H.; Hess, M. In-situ hybridization for the detection and identification of Histomonas meleagridis in tissues. J. Comp. Pathol. 2006, 135, 237–242. [Google Scholar] [CrossRef]
- Singh, A.; Weissenböck, H.; Hess, M. Histomonas meleagridis: Immunohistochemical localization of parasitic cells in formalin-fixed, paraffin-embedded tissue sections of experimentally infected turkeys demonstrates the wide spread of the parasite in its host. Exp. Parasitol. 2008, 118, 505–513. [Google Scholar] [CrossRef]
- Allen, E.A. A pentatrichomonas associated with certain cases of enterohepatitis or “blackhead” of poultry. Trans. Am. Microsc. Soc. 1936, 55, 315. [Google Scholar] [CrossRef]
- Liebhart, D.; Neale, S.; Garcia-Rueda, C.; Wood, A.M.; Bilic, I.; Wernsdorf, P.; Jaskulska, B.; Hess, M. A single strain of Tetratrichomonas gallinarum causes fatal typhlohepatitis in red-legged partridges (Alectoris rufa) to be distinguished from histomonosis. Avian Pathol. 2014, 43, 473–480. [Google Scholar] [CrossRef]
- Paudel, S.; Ruhnau, D.; Wernsdorf, P.; Liebhart, D.; Hess, M.; Hess, C. Presence of Avibacterium paragallinarum and histopathologic lesions corresponds with clinical signs in a co-infection model with Gallibacterium anatis. Avian Dis. 2017, 61, 335–340. [Google Scholar] [CrossRef] [PubMed]
- Jansen, I.; Lucas, M.; Savci-Heijink, C.D.; Meijer, S.L.; Marquering, H.A.; de Bruin, D.M.; Zondervan, P.J. Histopathology: Ditch the slides, because digital and 3D are on show. World J. Urol. 2018, 36, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Paudel, S.; Hess, C.; Wernsdorf, P.; Käser, T.; Meitz, S.; Jensen-Jarolim, E.; Hess, M.; Liebhart, D. The systemic multiplication of Gallibacterium anatis in experimentally infected chickens is promoted by immunosuppressive drugs which have a less specific effect on the depletion of leukocytes. Vet. Immunol. Immunopathol. 2015, 166, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Kidane, F.A.; Mitra, T.; Wernsdorf, P.; Hess, M.; Liebhart, D. Allocation of interferon gamma mRNA positive cells in caecum hallmarks a protective trait against histomonosis. Front. Immunol. 2018, 9, 1164. [Google Scholar] [CrossRef] [PubMed]
- Agrebi, S.; Larbi, A. Use of artificial intelligence in infectious diseases. In Artificial Intelligence in Precision Health: From Concept to Applications; Barh, D., Ed.; Elsevier: London, UK; Academic Press: San Diego, CA, USA, 2020; pp. 415–438. ISBN 9780128171332. [Google Scholar]
- Nam, S.; Chong, Y.; Jung, C.K.; Kwak, T.-Y.; Lee, J.Y.; Park, J.; Rho, M.J.; Go, H. Introduction to digital pathology and computer-aided pathology. J. Pathol. Transl. Med. 2020, 54, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Hess, M.; Neubauer, C.; Hackl, R. Interlaboratory comparison of ability to detect nucleic acid of Mycoplasma gallisepticum and Mycoplasma synoviae by polymerase chain reaction. Avian Pathol. 2007, 36, 127–133. [Google Scholar] [CrossRef]
Figure 1.
Immunohistochemistry for the detection of T cells (brown stained cells) in lymphoma caused by Marek’s disease virus.
Figure 1.
Immunohistochemistry for the detection of T cells (brown stained cells) in lymphoma caused by Marek’s disease virus.

Figure 2.
Diffuse infiltration of pleomorphic lymphoid cells mainly indicating peripheral neuropathy, Marek’s disease or reticuloendotheliosis.
Figure 2.
Diffuse infiltration of pleomorphic lymphoid cells mainly indicating peripheral neuropathy, Marek’s disease or reticuloendotheliosis.

Figure 3.
(a) Intranuclear inclusions indicating viral hepatitis in the liver of a pheasant (arrow). (b) Specific identification of a parvovirus by in situ hybridization (arrow).
Figure 3.
(a) Intranuclear inclusions indicating viral hepatitis in the liver of a pheasant (arrow). (b) Specific identification of a parvovirus by in situ hybridization (arrow).

Figure 4.
In situ hybridization for the specific detection and localization of (a) Histomonas meleagridis and (b) Tetratrichomonas gallinarum (arrow).
Figure 4.
In situ hybridization for the specific detection and localization of (a) Histomonas meleagridis and (b) Tetratrichomonas gallinarum (arrow).

Figure 5.
Diagram to demonstrate the holistic approach for diagnosing infectious poultry diseases (created with BioRender.com).
Figure 5.
Diagram to demonstrate the holistic approach for diagnosing infectious poultry diseases (created with BioRender.com).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liebhart, D.; Bilic, I.; Grafl, B.; Hess, C.; Hess, M. Diagnosing Infectious Diseases in Poultry Requires a Holistic Approach: A Review. Poultry 2023, 2, 252-280. https://doi.org/10.3390/poultry2020020
AMA Style
Liebhart D, Bilic I, Grafl B, Hess C, Hess M. Diagnosing Infectious Diseases in Poultry Requires a Holistic Approach: A Review. Poultry. 2023; 2(2):252-280. https://doi.org/10.3390/poultry2020020
Chicago/Turabian StyleLiebhart, Dieter, Ivana Bilic, Beatrice Grafl, Claudia Hess, and Michael Hess. 2023. "Diagnosing Infectious Diseases in Poultry Requires a Holistic Approach: A Review" Poultry 2, no. 2: 252-280. https://doi.org/10.3390/poultry2020020