
Characterization of Avian Pathogenic Escherichia coli Isolated from Broiler Breeders with Colibacillosis in Mississippi
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. E. coli Isolation and Identification
2.2. DNA Isolation
2.3. Serotyping
2.4. Phylogenetic Classification
2.5. Screening for Virulence Genes
2.6. Antimicrobial Resistance (AMR) and Heavy-Metal Resistance Patterns
2.6.1. Screening for Antimicrobial and Heavy-Metal Resistance Genes
2.6.2. Antimicrobial and Heavy-Metal Susceptibility Testing
Antibiotic Susceptibility Testing
QAC and Heavy-Metal Susceptibility Testing
3. Results
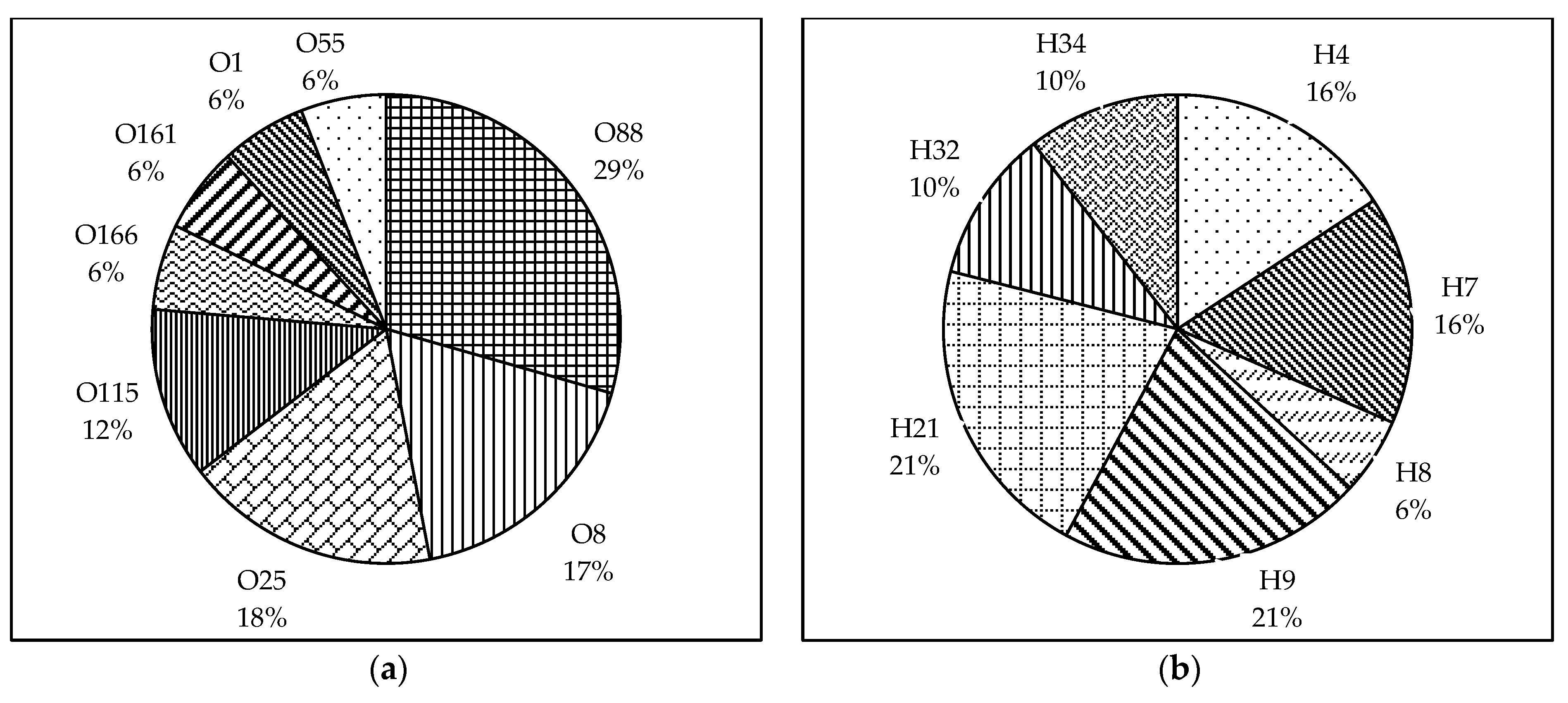
3.1. Serotyping
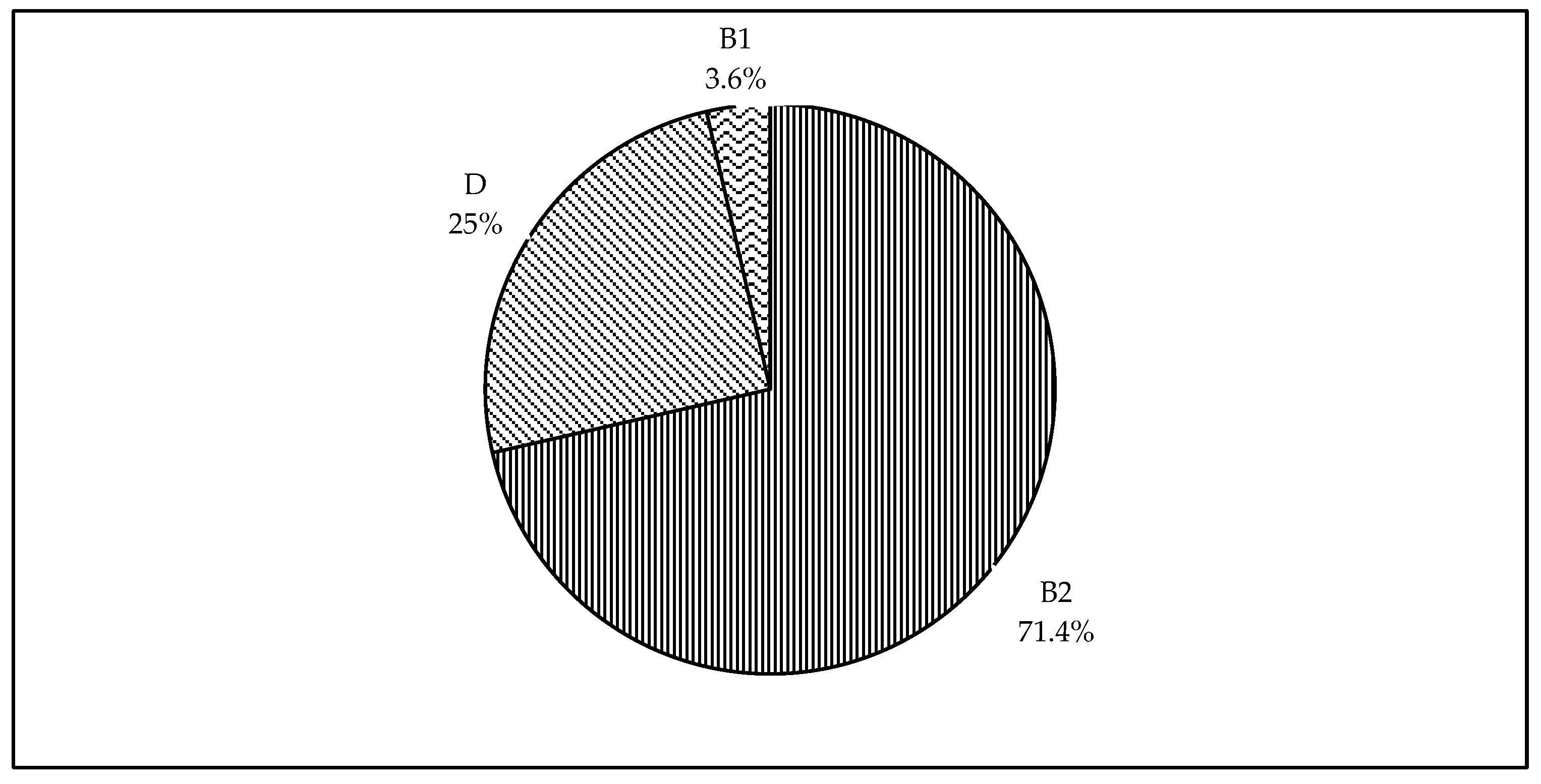
3.2. Phylogenetic Classification
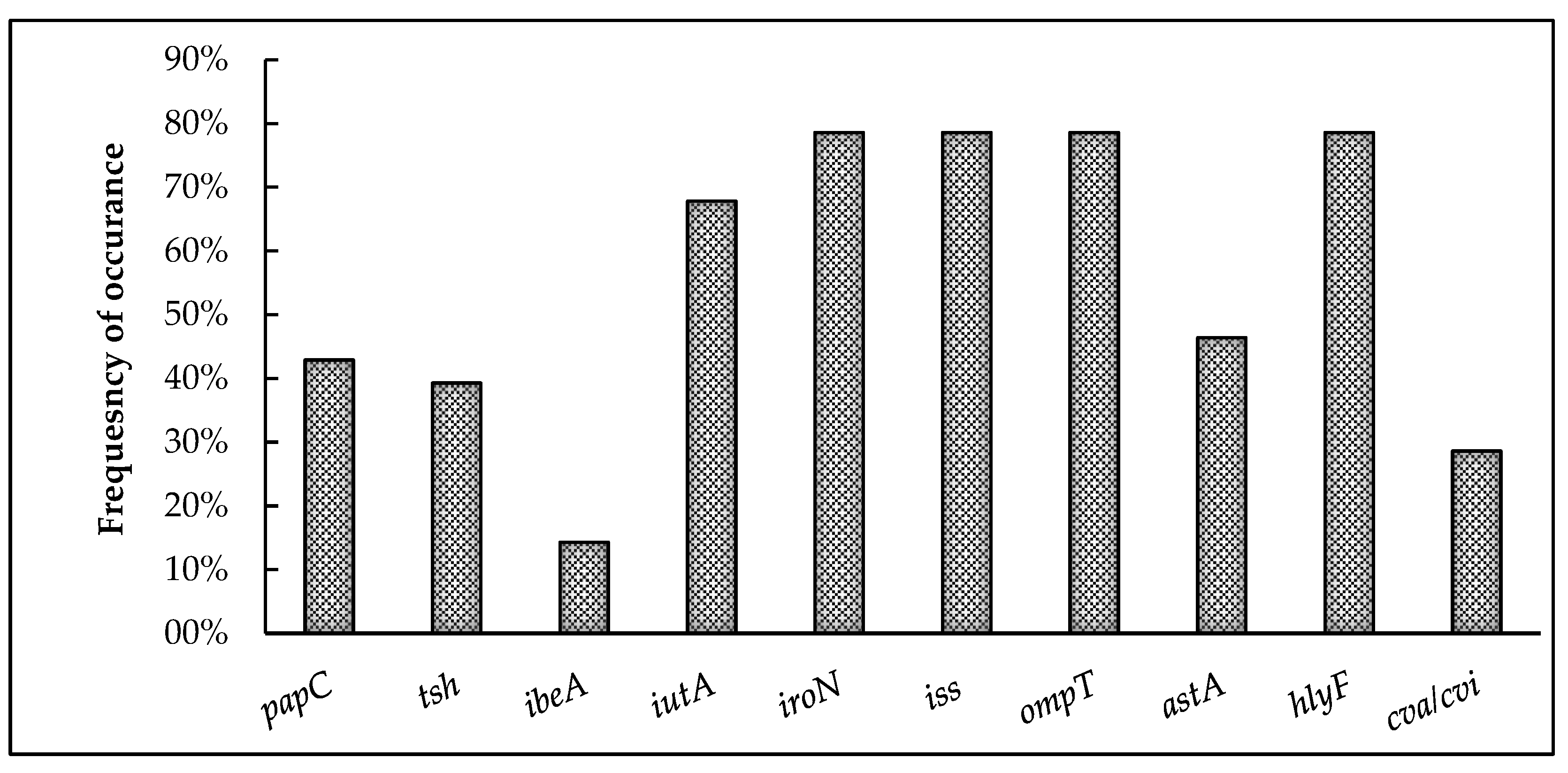
3.3. Screening for Virulence Genes
3.4. AMR and Heavy-Metal Resistance Patterns
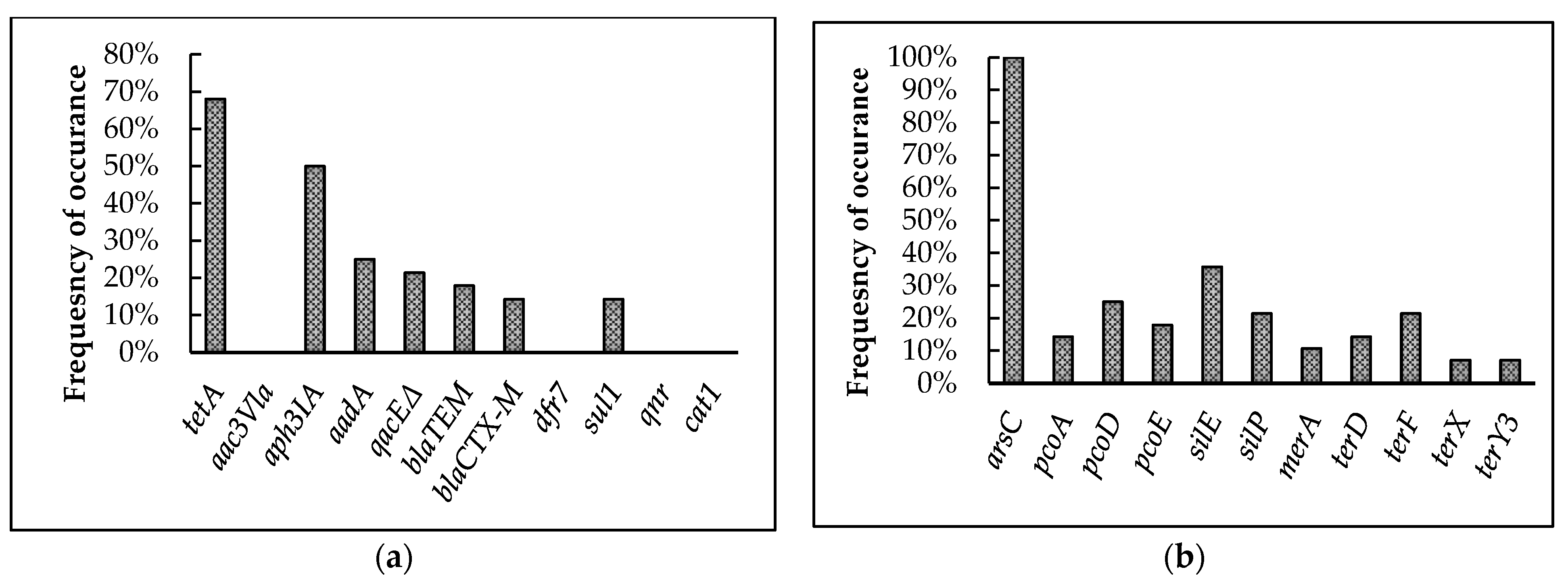
3.4.1. Screening for Antimicrobial and Heavy-Metal Resistance Genes
3.4.2. Antimicrobial and Heavy-Metal Susceptibility Testing
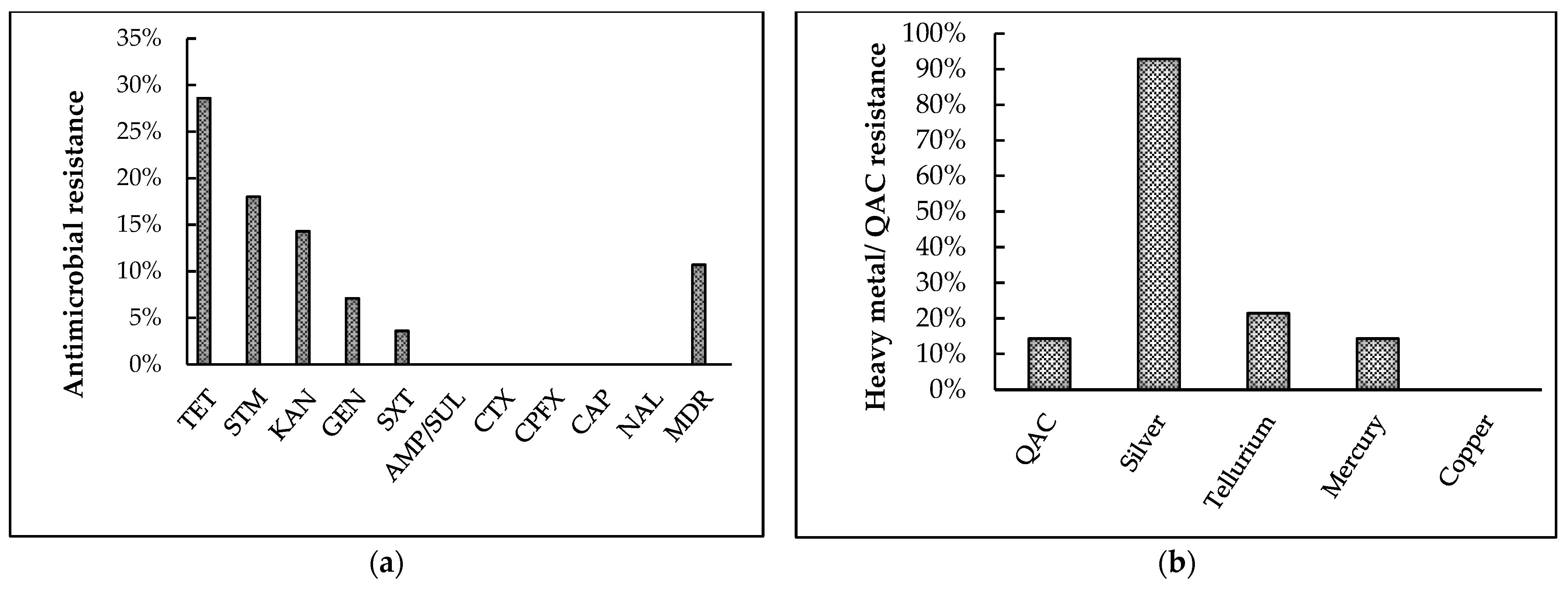
Antibiotic-Susceptibility Testing
QAC and Heavy-Metal Susceptibility Testing
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sannes, M.R.; Kuskowski, M.A.; Owens, K.; Gajewski, A.; Johnson, J.R. Virulence factor profiles and phylogenetic background of Escherichia coli isolates from veterans with bacteremia and uninfected control subjects. J. Infect. Dis. 2004, 190, 2121–2128. [Google Scholar] [CrossRef] [Green Version]
- Vila, J.; Aez-L’Opez, E.S.; Johnson, J.R.; Omling, U.R.; Dobrindt, U.; Cantón, R.; Cantón, C.; Giske, C.G.; Naas, T.; Carattoli, A.; et al. Escherichia coli: An old friend with new tidings. FEMS Microbiol. Rev. 2016, 5, 437–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dho-Moulin, M.; Fairbrother, J.M. Avian pathogenic Escherichia coli (APEC). Vet. Res. 1999, 30, 299–316. [Google Scholar] [PubMed]
- Kathayat, D.; Lokesh, D.; Ranjit, S.; Rajashekara, G. Avian Pathogenic Escherichia coli (APEC): An Overview of Virulence and Pathogenesis Factors, Zoonotic Potential, and Control Strategies. Pathogens 2021, 10, 467. [Google Scholar] [CrossRef] [PubMed]
- Dziva, F.; Stevens, M. Colibacillosis in poultry: Unravelling the molecular basis of virulence of avian pathogenic Escherichia coli in their natural hosts. Avian Pathol. 2008, 37, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.J.; Wannemuehler, Y.; Doetkott, C.; Johnson, S.J.; Rosenberger, S.C.; Nolan, L.K. Identification of Minimal Predictors of Avian Pathogenic Escherichia coli Virulence for Use as a Rapid Diagnostic Tool. J. Clin. Microbiol. 2008, 46, 3987–3996. [Google Scholar] [CrossRef] [Green Version]
- Fancher, C.A.; Zhang, L.; Kiess, A.S.; Adhikari, P.A.; Dinh, T.T.N.; Sukumaran, A.T. Avian Pathogenic Escherichia coli and Clostridium perfringens: Challenges in No Antibiotics Ever Broiler Production and Potential Solutions. Microorganisms 2020, 8, 1533. [Google Scholar] [CrossRef] [PubMed]
- Ghunaim, H.; Abu-Madi, M.A.; Kariyawasam, S. Advances in vaccination against avian pathogenic Escherichia coli respiratory disease: Potentials and limitations. Vet. Microbiol. 2014, 172, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Mellata, M. Human and avian extraintestinal pathogenic Escherichia coli: Infections, zoonotic risks, and antibiotic resistance trends. Foodborne Pathog. Dis. 2013, 10, 916–932. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, H.; Yonehara, K.; Murase, T. Virulence of Escherichia coli Isolates Obtained from Layer Chickens with Colibacillosis Associated with Pericarditis, Perihepatitis, and Salpingitis in Experimentally Infected Chicks and Embryonated Eggs. Avian Dis. 2018, 62, 233–236. [Google Scholar] [CrossRef]
- Giovanardi, D.; Campagnari, E.; Sperati Ruffoni, L.; Pesente, P.; Ortali, G.; Furlattini, V. Avian pathogenic Escherichia coli transmission from broiler breeders to their progeny in an integrated poultry production chain. Avian Pathol. 2005, 34, 313–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yassin, H.; Velthuis, A.G.J.; Boerjan, M.; van Riel, J.V. Field study on broilers’ first-week mortality. Poult. Sci. J. 2009, 88, 798–804. [Google Scholar] [CrossRef]
- Poulsen, L.L.; Thøfner, I.; Bisgaard, M.; Christensen, J.P.; Olsen, R.H.; Christensen, H. Longitudinal study of transmission of Escherichia coli from broiler breeders to broilers. Vet. Microbiol. 2017, 207, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Collingwood, C.; Kemmett, K.; Williams, N.; Wigley, P.; Tellez, G. Is the concept of avian pathogenic Escherichia coli as a single pathotype fundamentally flawed? Front Vet. Sci. 2014, 1, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolan, L.K.; Barnes, H.; Jean Pirre, V.; Tahseen, A.A.; Catherine, M.L. Diseases of Poultry, 13th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 751–785. [Google Scholar]
- Newman, D.M.; Barbieri, N.L.; de Oliveira, A.L.; Willis, D.; Nolan, L.K.; Logue, C.M. Characterizing avian pathogenic Escherichia coli (APEC) from colibacillosis cases. PeerJ. 2021, 9, e11025. [Google Scholar] [CrossRef]
- Jiang, F.; An, Y.; Bao, X.; Zhao, R.; Jernigan, L.; Lithio, A.; Nettleton, D.; Li, L.; Wurtele, E.S.; Nolan, L.K.; et al. ArcA Controls Metabolism, Chemotaxis, and Motility Contributing to the Pathogenicity of Avian Pathogenic Escherichia coli. Infect. Immun. 2015, 83, 3545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-PLoSkonska, G.; Choroszy-Krol, I. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 2019, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.J.; Wannemuehler, Y.; Kariyawasam, S.; Johnson, J.R.; Logue, C.M.; Nolan, L.K. Prevalence of avian-pathogenic Escherichia coli strain O1 Genomic islands among extraintestinal and commensal E. Coli isolates. J. Bacteriol. 2012, 194, 2846–2853. [Google Scholar] [CrossRef] [Green Version]
- Barbieri, N.L.; De Oliveira, A.L.; Tejkowski, T.M.; Pavanelo, D.B.; Rocha, D.A.; Matter, L.B.; Callegari-Jacques, S.M.; De Brito, B.G.; Horn, F. Genotypes and pathogenicity of cellulitis isolates reveal traits that modulate APEC virulence. PLoS ONE 2013, 8, e72322. [Google Scholar] [CrossRef] [Green Version]
- Schouler, C.; Schaeffer, B.; Brée, A.; Mora, A.; Dahbi, G.; Biet, F.; Oswald, E.; Mainil, J.; Blanco, J.; Moulin-Schouleur, M. Diagnostic strategy for identifying avian pathogenic Escherichia coli based on four patterns of virulence genes. J. Clin. Microbiol. 2012, 50, 1673–1678. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, D.R.A.; Octavia, S.; Lan, R. Population structure and virulence content of avian pathogenic Escherichia coli isolated from outbreaks in Sri Lanka. Vet. Microbiol. 2014, 168, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Guabiraba, R.; Schouler, C. Avian colibacillosis: Still many black holes. FEMS Microbiol. Lett. 2015, 362, 118. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.E.; Blanco, M.; Mora, A.; Jansen, W.H.; García, V.; Vázquez, M.L.; Blanco, J. Serotypes of Escherichia coli isolated from septicaemic chickens in Galicia (northwest Spain). Vet. Microbiol. 1998, 61, 229–235. [Google Scholar] [CrossRef]
- Joensen, K.G.; Tetzschner, A.M.M.; Iguchi, A.; Aarestrup, F.M.; Scheutz, F. Rapid and easy in silico serotyping of Escherichia coli isolates by use of whole-genome sequencing data. J. Clin. Microbiol. 2015, 53, 2410–2426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lía Padola, N.; Martin Dozois, C.; Delannoy, S.; Fleiss, A.; Beutin, L.; Mariani-Kurkdjian, P.; Bonacorsi, S.; Fach, P. The Escherichia coli Serogroup O1 and O2 Lipopolysaccharides Are Encoded by Multiple O-antigen Gene Clusters. Front Cell Infect. Microbiol. 2017, 7, 30. [Google Scholar]
- Rodriguez-Siek, K.E.; Giddings, C.W.; Doetkott, C.; Johnson, T.J.; Nolan, L.K. Characterizing the APEC pathotype. Vet. Res. 2005, 36, 241–256. [Google Scholar] [CrossRef] [Green Version]
- Knöbl, T.; Moreno, A.M.; Paixão, R.; Gomes, T.A.; Vieira, M.A.; da Silva Leite, D.; Blanco, J.E.; Ferreira, A.J. Prevalence of avian pathogenic Escherichia coli (APEC) clone harboring sfa gene in Brazil. Sci. World J. 2012, 2012, 437342. [Google Scholar] [CrossRef] [Green Version]
- Ronco, T.; Stegger, M.; Olsen, R.H. Spread of avian pathogenic Escherichia coli ST117 O78:H4 in Nordic broiler production. BMC Genom. 2017, 18, 13. [Google Scholar] [CrossRef] [Green Version]
- Doyle, M.P.; Schoeni, J.L. Isolation of Escherichia coli O157: H7 from retail fresh meats and poultry. Appl. Environ. Microbiol. 1987, 53, 2394–2396. [Google Scholar] [CrossRef] [Green Version]
- Cortés, P.; Blanc, V.; Mora, A.; Dahbi, G.; Blanco, J.E.; Blanco, M.; López, C.; Andreu, A.; Navarro, F.; Alonso, M.P.; et al. Isolation and characterization of potentially pathogenic antimicrobial-resistant Escherichia coli strains from chicken and pig farms in Spain. Appl. Environ. Microbiol. 2010, 76, 2799–2805. [Google Scholar] [CrossRef] [Green Version]
- Ishii, N.; Nakahigashi, K.; Baba, T.; Robert, M.; Soga, T.; Kanai, A.; Hirasawa, T.; Naba, M.; Hirai, K.; Hoque, A.; et al. Multiple high-throughput analyses monitor the response of E. coli to perturbations. Science 2007, 316, 593–597. [Google Scholar] [CrossRef]
- EscobarPáramo, P.; LeMenac’h, A.; LeGall, T.; Amorin, C.; Gouriou, S.; Picard, B.; Skurnik, D.; Denamur, E. Identification of forces shaping the commensal Escherichia coli genetic structure by comparing animal and human isolates. Environ. Microbiol. 2006, 8, 1975–1984. [Google Scholar] [CrossRef] [Green Version]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [Green Version]
- Pires-dos-Santos, T.; Bisgaard, M.; Christensen, H. Genetic diversity and virulence profiles of Escherichia coli causing salpingitis and peritonitis in broiler breeders. Vet. Microbiol. 2013, 162, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Cordoni, G.; Woodward, M.J.; Wu, H.; Alanazi, M.; Wallis, T.; La Ragione, R.M. Comparative genomics of European avian pathogenic E. coli. (APEC). BMC Genom. 2016, 17, 960. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.B.; Yoon, M.Y.; Ha, J.S.; Seo, K.W.; Noh, E.B.; Son, S.H.; Lee, Y.J. Molecular characterization of avian pathogenic Escherichia coli from broiler chickens with colibacillosis. Poult. Sci. J. 2020, 99, 1088–1095. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.I.; McQuillan, J.; Taiwo, M.; Parks, R.; Stenton, C.A.; Morgan, H.; Mowlem, M.C.; Lees, D.N. A highly specific Escherichia coli qPCR and its comparison with existing methods for environmental waters. Water Res. 2017, 126, 101–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Oliveira, A.L.; Newman, D.M.; Sato, Y.; Noel, A.; Rauk, B.; Nolan, L.K.; Barbieri, N.L.; Logue, C.M. Characterization of Avian Pathogenic Escherichia coli (APEC) Associated with Turkey Cellulitis in Iowa. Front Vet. Sci. 2020, 7, 380. [Google Scholar] [CrossRef]
- Iguchi, A.; Iyoda, S.; Seto, K.; Morita-Ishihara, T.; Scheutz, F.; Ohnishi, M. Escherichia coli O-Genotyping PCR: A Comprehensive and Practical Platform for Molecular O Serogrouping. J. Clin. Microbiol. 2015, 53, 2427–2432. [Google Scholar] [CrossRef] [Green Version]
- Banjo, M.; Iguchi, A.; Seto, K.; Kikuchi, T.; Harada, T.; Scheutz, F.; Iyoda, S. Escherichia coli H-Genotyping PCR: A complete and practical platform for molecular h typing. J. Clin. Microbiol. 2018, 56, 190–208. [Google Scholar] [CrossRef] [Green Version]
- Subedi, M.; Luitel, H.; Devkota, B.; Bhattarai, R.K.; Phuyal, S.; Panthi, P.; Shrestha, A.; Chaudhary, D.K. Antibiotic resistance pattern and virulence genes content in avian pathogenic Escherichia coli (APEC) from broiler chickens in Chitwan, Nepal. BMC Vet. Res. 2018, 14, 113. [Google Scholar] [CrossRef]
- Li, T.; Castañeda, C.D.; Miotto, J.; McDaniel, C.; Kiess, A.S.; Zhang, L. Effects of in ovo probiotic administration on the incidence of avian pathogenic Escherichia coli in broilers and an evaluation on its virulence and antimicrobial resistance properties. Poult. Sci. J. 2021, 100, 100903. [Google Scholar] [CrossRef]
- Logue, C.M.; Wannemuehler, Y.; Nicholson, B.A.; Doetkott, C.; Barbieri, N.L.; Nolan, L.K. Comparative analysis of phylogenetic assignment of human and avian ExPEC and fecal commensal Escherichia coli using the (previous and revised) clermont phylogenetic typing methods and its impact on avian pathogenic Escherichia coli (APEC) classification. Front Microbiol. 2017, 8, 283. [Google Scholar] [CrossRef] [Green Version]
- Batchelor, M.; Clifton-Hadley, F.A.; Stallwood, A.D.; Paiba, G.A.; Davies, R.H.; Liebana, E. Detection of multiple cephalosporin-resistant Escherichia coli from a cattle fecal sample in Great Britain. Microb. Drug Resist. 2005, 1, 58–61. [Google Scholar] [CrossRef]
- Van, T.T.; Chin, J.; Chapman, T.; Tran, L.T.; Coloe, P.J. Safety of raw meat and shellfish in Vietnam: An analysis of Escherichia coli isolations for antibiotic resistance and virulence genes. Int. J. Food Microbiol. 2008, 10, 217–223. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 32nd ed.; CLSI: Wayne, PA, USA, 2022; p. 34. [Google Scholar]
- Deus, D.; Krischek, C.; Pfeifer, Y.; Sharifi, A.R.; Fiegen, U.; Reich, F.; Klein, G.; Kehrenberg, C. Comparative analysis of the susceptibility to biocides and heavy metals of extended-spectrum β-lactamase-producing Escherichia coli isolates of human and avian origin, Germany. Diagn. Microbiol. Infect. Dis. 2017, 88, 88–92. [Google Scholar] [CrossRef]
- Vajiheh, K.; Naser, B.; Giti, E. Antimicrobial, heavy metal resistance and plasmid profile of coliforms isolated from nosocomial infections in a hospital in Isfahan, Iran. Afr. J. Biotechnol. 2003, 2, 379–383. [Google Scholar] [CrossRef]
- Vrionis, H.A.; Wang, S.; Haslam, B.; Turner, R.J. Selenite protection of tellurite toxicity toward Escherichia coli. Front Mol. Biosci. 2015, 2, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herisse, M.; Duverger, Y.; Martin-Verstraete, I.; Barras, F.; Ezraty, B. Silver potentiates aminoglycoside toxicity by enhancing their uptake. Mol. Microbiol. 2017, 105, 115–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McPeake, S.J.; Smyth, J.A.; Ball, H.J. Characterisation of avian pathogenic Escherichia coli (APEC) associated with colisepticaemia compared to faecal isolates from healthy birds. Vet. Microbiol. 2005, 110, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Solà-Ginés, M.; Cameron-Veas, K.; Badiola, I.; Dolz, R.; Majó, N.; Dahbi, G. Diversity of Multi-Drug Resistant Avian Pathogenic Escherichia coli (APEC) Causing Outbreaks of Colibacillosis in Broilers during 2012 in Spain. PLoS ONE. 2015, 10, e0143191. [Google Scholar] [CrossRef] [Green Version]
- Someya, A.; Otsuki, K.; Murase, T. Characterization of Escherichia coli strains obtained from layer chickens affected with colibacillosis in a commercial egg-producing farm. J. Vet. Med. Sci. 2007, 69, 1009–1014. [Google Scholar] [CrossRef] [Green Version]
- Naundrup, T.I.C.; Poulsen, L.L.; Bisgaard, M.; Christensen, H.; Olsen, R.H.; Christensen, J.P. Longitudinal study on causes of mortality in Danish broiler breeders. Avian Dis. 2019, 63, 400–410. [Google Scholar] [CrossRef]
- Christensen, H.; Bachmeier, J.; Bisgaard, M. New strategies to prevent and control avian pathogenic Escherichia coli (APEC). Avian Pathol. 2021, 50, 370–381. [Google Scholar] [CrossRef] [PubMed]
- Monroy, M.A.R.; Knöbl, T.; Bottino, J.A.; Astolfi Ferreira, C.S.; Ferreira, A.J.P. Virulence characteristics of Escherichia coli isolates obtained from broiler breeders with salpingitis. Comp. Immunol. Microbiol. Infect. Dis. 2005, 28, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Koutsianos, D.; Athanasiou, L.V.; Mossialos, D.; Franzo, G.; Cecchinato, M.; Koutoulis, K.C. Investigation of Serotype Prevalence of Escherichia coli Strains Isolated from Layer Poultry in Greece and Interactions with Other Infectious Agents. Vet. Sci. 2022, 9, 152. [Google Scholar] [CrossRef] [PubMed]
- Awad, A.M.; El-shall, N.A.; Khalil, D.S.; El-hack, M.E.A.; Swelum, A.A.; Mahmoud, A.H.; Ebaid, H.; Komany, A.; Sammour, R.H.; Sedeik, M.E. Incidence, Pathotyping, and Antibiotic Susceptibility of Avian Pathogenic Escherichia coli among Diseased Broiler Chicks. Pathogens 2020, 9, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolan, L.K.; Vaillancourt, J.P.; Barbieri, N.L.; Logue, C.M. Diseases of Poultry, 14th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 770–830. [Google Scholar]
- Oliveira, E.S.; Cardozo, M.V.; Borzi, M.M.; Borges, C.A.; Guastalli, E.A.L.; Ávila, F.A. Highly Pathogenic and Multidrug Resistant Avian Pathogenic Escherichia Coli in Free-Range Chickens from Brazil. Braz. J. Poult. Sci. 2019, 21, 1–8. [Google Scholar] [CrossRef]
- Lozica, L.; Kabalin, A.E.; Dolenčić, N.; Vlahek, M.; Gottstein, Ž. Phylogenetic characterization of avian pathogenic Escherichia coli strains longitudinally isolated from broiler breeder flocks vaccinated with autogenous vaccine. Poult. Sci. J. 2021, 100, e101079. [Google Scholar] [CrossRef]
- Johnson, T.J.; Johnson, S.J.; Nolan, L.K. Complete DNA sequence of a ColBM plasmid from avian pathogenic Escherichia coli suggests that it evolved from closely related ColV virulence plasmids. J. Bacteriol. 2006, 188, 5975–5983. [Google Scholar] [CrossRef] [Green Version]
- Varga, C.; Brash, M.L.; Slavic, D.; Boerlin, P.; Ouckama, R.; Weis, A.; Petrik, M.; Philippe, C.; Barham, M.; Guerin, M.T. Evaluating Virulence-Associated Genes and Antimicrobial Resistance of Avian Pathogenic Escherichia coli Isolates from Broiler and Broiler Breeder Chickens in Ontario, Canada. Avian Dis. 2018, 62, 291–299. [Google Scholar] [CrossRef] [PubMed]
- De Carli, S.; Ikuta, N.; Lehmann, F.K.M.; da Silveira, V.P.; de Melo Predebon, G.; Fonseca, A.S.K.; Lunge, V.R. Virulence gene content in Escherichia coli isolates from poultry flocks with clinical signs of colibacillosis in Brazil. Poult. Sci. J. 2015, 94, 2635–2640. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, A.L.; Rocha, D.A.; Finkler, F.; de Moraes, L.B.; Barbieri, N.L.; Pavanelo, D.B.; Winkler, C.; Grassotti, T.T.; de Brito, K.C.; de Brito, B.G.; et al. Prevalence of ColV plasmid-linked genes and in vivo pathogenicity of avian strains of Escherichia coli. Foodborne Pathog. Dis. 2015, 12, 679–685. [Google Scholar] [CrossRef]
- Ahmed, A.M.; Shimamoto, T.; Shimamoto, T. Molecular characterization of multidrug-resistant avian pathogenic Escherichia coli isolated from septicemic broilers. Int. J. Med. Microbiol. 2013, 303, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Aarestrup, F.M. Veterinary Drug Usage and Antimicrobial Resistance in Bacteria of Animal Origin. Basic Clin. Pharmacol. Toxicol. 2005, 96, 271–281. [Google Scholar] [CrossRef]
- Mazel, D. Integrons: Agents of bacterial evolution. Nat. Rev. Microbiol. 2006, 4, 608–620. [Google Scholar] [CrossRef] [PubMed]
- Agyare, C.; Boamah, V.E.; Zumbi, C.N.; Osei, F.B. Antimicrobial Resistance—A Global Threat, 1st ed.; Intechopen Limited: London, UK, 2018; pp. 33–51. [Google Scholar]
- Indrawati, A.; Khoirani, K.; Setiyaningsih, S.; Affif, U.; Safika; Ningrum, S.G. Detection of Tetracycline Resistance Genes among Escherichia coli Isolated from Layer and Broiler Breeders in West Java, Indonesia. Trop. J. Anim. Sci. 2021, 44, 267–272. [Google Scholar] [CrossRef]
- Thomrongsuwannakij, T.; Blackall, P.J.; Djordjevic, S.P.; Cummins, M.L.; Chansiripornchai, N. A comparison of virulence genes, antimicrobial resistance profiles and genetic diversity of avian pathogenic Escherichia coli (APEC) isolates from broilers and broiler breeders in Thailand and Australia. Avian Pathol. 2020, 49, 457–466. [Google Scholar] [CrossRef]
- Kim, S.W.; Kim, K.; JuLee, Y. Comparative analysis of antimicrobial resistance and genetic characteristics of Escherichia coli from broiler breeder farms in Korea. Can. J. Anim. Sci. 2022, 102, 342–351. [Google Scholar] [CrossRef]
- Ibrahim, R.A.; Cryer, T.L.; Lafi, S.Q.; Basha, E.A.; Good, L.; Tarazi, Y.H. Identification of Escherichia coli from broiler chickens in Jordan, their antimicrobial resistance, gene characterization and the associated risk factors. BMC Vet. Res. 2019, 15, 159. [Google Scholar] [CrossRef] [Green Version]
- Heuer, H.; Krögerrecklenfort, E.; Wellington, E.M.H.; Egan, S.; van Elsas, J.D.; van Overbeek, L.; Collard, J.M.; Karagouni, G.A.D.; Nikolakopoulou, T.L.; Smalla, K. Gentamicin resistance genes in environmental bacteria: Prevalence and transfer. FEMS Microbiol. Ecol. 2002, 42, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Benameur, Q.; Tali-Maamar, H.; Assaous, F.; Guettou, B.; Rahal, K.; Ben-Mahdi, M.H. Detection of multidrug resistant Escherichia coli in the ovaries of healthy broiler breeders with emphasis on extended-spectrum β-lactamases producers. Comp. Immunol. Micrbiol. Infect. Dis. 2019, 64, 163–167. [Google Scholar] [CrossRef] [PubMed]
- Zurfluh, K.; Wang, J.; Klumpp, J.; Nüesch-Inderbinen, M.; Fanning, S.; Stephan, R. Vertical transmission of highly similar blaCTX-M-1-harbouring IncI1 plasmids in Escherichia coli with different MLST types in the poultry production pyramid. Front. Microbiol. 2014, 5, 519. [Google Scholar] [CrossRef]
- Oikarainen, P.E.; Pohjola, L.K.; Pietola, E.S.; Heikinheimo, A. Direct vertical transmission of ESBL/pAmpC-producing Escherichia coli limited in poultry production pyramid. Vet. Microbiol 2019, 231, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, O.; Börjesson, S.; Landén, A.; Bengtsson, B. Vertical transmission of Escherichia coli carrying plasmid-mediated AmpC (pAmpC) through the broiler production pyramid. J. Antimicrob. Chemother. 2014, 69, 1497–1500. [Google Scholar] [CrossRef]
- Cyoia, P.S.; Koga, V.L.; Nishio, E.K.; Houle, S.; Dozois, C.M.; de Brito, K.C.T.; de Brito, B.G.; Nakazato, G.; Kobayashi, R.K.T. Distribution of ExPEC Virulence Factors, blaCTX-M, fosA3, and mcr-1 in Escherichia coli Isolated from Commercialized Chicken Carcasses. Front. Microbiol. 2019, 14, 3254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afayibo, D.J.A.; Zhu, H.; Zhang, B.; Yao, L.; Abdelgawad, H.A.; Tian, M.; Qi, J.; Liu, Y.; Wang, S. Isolation, Molecular Characterization, and Antibiotic Resistance of Avian Pathogenic Escherichia coli in Eastern China. Vet. Sci. 2022, 25, 319. [Google Scholar] [CrossRef]
- Neil, W.; Jane, F.T.; David, M.L. Multiresistant Gram-negative bacteria: The role of high-risk clones in the dissemination of antibiotic resistance. FEMS Microbiol. Rev. 2011, 35, 736–755. [Google Scholar]
- Ashraf, A.A.T.; Soad, A.N.; Fatma, I.E.H.; Ola, A.I. Prevalence of eaeA and qacEΔ1 genes in Escherichia coli isolated from omphalitis in baby chicks. Benha. Vet. Med. J. 2017, 32, 184–192. [Google Scholar]
- Maertens, H.; Demeyere, K.; De Reu, K. Effect of subinhibitory exposure to quaternary ammonium compounds on the ciprofloxacin susceptibility of Escherichia coli strains in animal husbandry. BMC Microbiol. 2020, 20, 155. [Google Scholar] [CrossRef]
- Liu, Q.; Peng, H.; Lu, X.; Zuidhof, M.J.; Li, X.F.; Le, X.C. Arsenic Species in Chicken Breast: Temporal Variations of Metabolites, Elimination Kinetics, and Residual Concentrations. Environ. Health Perspect. 2016, 124, 1174–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Deng, W.; Liu, S.; Yu, X.; Mustafa, G.R.; Chen, S.; He, L.; Ao, X.; Yang, Y.; Zhou, K.; et al. Presence of heavy metal resistance genes in Escherichia coli and Salmonella isolates and analysis of resistance gene structure in E. coli E308. J. Glob. Antimicrob. Resist. 2020, 21, 420–426. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Description | Size (bp) | Primer Sequence (5′-3′) | Annealing Temperature (°C) | References |
|---|---|---|---|---|---|
| Virulence genes | |||||
| iroN | Iron acquisition | 553 | F: AATCCGGCAAAGAGACGAACCGCCT R: GTTCGGGCAACCCCTGCTTTGACTTT | 63 | [6] |
| iutA | 302 | F: GGCTGGACATCATGGGAACTGG R: CGTCGGGAACGGGTAGAATCG | 63 | [6] | |
| ompT | Protectins | 496 | F: TCATCCCGGAAGCCTCCCTCACTACTAT R: TAGCGTTTGCTGCACTGGCTTCTGATAC | 63 | [6] |
| iss | 323 | F: CAGCAACCCGAACCACTTGATG R: AGCATTGCCAGAGCGGCAGAA | 63 | [6] | |
| hlyF | Toxins | 450 | F: GGCCACAGTCGTTTAGGGTGCTTACC R: GGCGGTTTAGGCATTCCGATACTCAG | 63 | [6] |
| astA | 116 | F: TGCCATCAACACAGTATATCC R: TCAGGTCGCGAGTGACGGC | 57 | [42] | |
| papC | Adhesins | 501 | F: TGATATCACGCAGTCAGTAGC R: CCGGCCATATTCACATAA | 60 | [42] |
| tsh | 824 | F: ACTATTCTCTGCAGGAAGTC R: CTTCCGATGTTCTGAACGT | 60 | [42] | |
| ibeA | Invasins | 171 | F: AGGCAGGTGTGCGCCGCGTAC R: TGGTGCTCCGGCAAACCATGC | 63 | [39] |
| cva/cvi | Colicin V operon | 1181 | F: TGGTAGAATGTGCCAGAGCAAG R: GAGCTGTTTGTAGCGAAGCC | 60 | [42] |
| Gene | Description | Size (bp) | Primer Sequence (5′-3′) | Annealing Temperature (°C) | References |
|---|---|---|---|---|---|
| Antimicrobial Resistance Genes | |||||
| blaTEM | β-lactamase inhibitor | 558 | F: ATGTGCGCGGAACCCCTATTTGTTTA R: AAAAAGCGGTTAGCTCCTTCGGTCCT | 55 | [44] |
| blaCTX-M | 3rd Generation Cephalosporins | 585 | F: CGATGTGCAGTACCAGTAA R: TTAGTGACCAGAATCAGCGG | 56 | [45] |
| aac3Vla | Aminoglycosides | 502 | F: GGCACCCGCGACGCCCTGGTCCAAAAG R: GGGCCCGGCGCCGATCGACAGGATTT | 55 | [44] |
| aph3IA | 378 | F: TCGGGCAATCAGGTGCGACAATCTA R: TGCCAGCGCATCAACAATATTTTCACC | 55 | [44] | |
| aadA | 365 | F: TAACGGCGCAGTGGCGGTTTTCA R: AAGCTCGCCGCGTTGTTTCATCAAG | 55 | [6] | |
| tetA | Tetracyclines | 372 | F: CGGGGCGACTGGGGCGGTAGC R: CAAAGCGCGGCCGGCACCTGT | 55 | [19] |
| dfr7 | Sulfonamides | 214 | F: TCTTTAAAGCGCTCACATATAATCAGTG R: ATTTGACCGCCACCAGAGACA | 55 | [44] |
| sul1 | 462 | F: CGCCGCTCTTAGACGCCCTGTCC R: CGCCGCTCTTAGACGCCCTGTCC | 55 | [19] | |
| qnr | Quinolones | 440 | F: TCGCCGCTGCCGCTTTTATCAGT R: GCCAACAGTCGCGGGAGAAGGTG | 55 | [45] |
| cat1 | Phenicols | 547 | F: AGTTGCTCAATGTACCTATAACC R: TTGTAATTCATTAAGCATTCTGCC | 56 | [46] |
| Heavy-Metal Resistance Genes | |||||
| terD | Tellurite | 231 | F: CCACTGCGCGGAATTTCCACTCACCAT R: ACGCCGTCCCGTCTGATGTTGACAAG | 55 | [43] |
| terX | 576 | F: ATGCGCCGCCTGCCTGTTTACCTTGTTA R: CGCGCTTGTGCTGCCGGAAGACA | 55 | [43] | |
| terF | 428 | F: CCGACAAACTTCCAGAAGATGGGGTAGT R: GAGGCAGCGGTTGCATTTGTACTTGACG | 55 | [43] | |
| terY3 | 302 | F: CCTGGGGCCGTCAGCGGACCTG R: TCCTTGCTGGTGGCCGTTCATACTTCAT | 55 | [43] | |
| pcoA | Copper | 507 | F: ATCCGGAAGGTCAGCACCGTCCATAGAC R: GACCTCGCGGATGTCAGTGGCTACACCT | 55 | [43] |
| pcoD | 502 | F: GGCGCCCAGAATGATAATCGCAACA R: GGGCGTGGCGCTGGCTACACTT | 55 | [43] | |
| pcoE | 385 | F: GTGGGGCAGCTTTTGCTCAGTCCAGTGA R: CGAAGCTTTCTTGCCTGCGTCTGATGTG | 55 | [43] | |
| arsC | Arsenic | 268 | F: ACCCGCTTCATCAACCACTT R: TGCCGATATGGGGATTTCCG | 55 | [43] |
| silP | Silver | 603 | F: ACACCCCGGCCTGGGCTCCTT R: GTGCGGGCACGGGAACAAACCTC | 55 | [43] |
| merA | Mercury | 250 | F: GATCCGCGCCGCCCATATCGCCCATCTG R: CACGCGCTCGCCGCCGTCGTTGAGTTG | 56 | [43] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joseph, J.; Jennings, M.; Barbieri, N.; Zhang, L.; Adhikari, P.; Ramachandran, R. Characterization of Avian Pathogenic Escherichia coli Isolated from Broiler Breeders with Colibacillosis in Mississippi. Poultry 2023, 2, 24-39. https://doi.org/10.3390/poultry2010004
Joseph J, Jennings M, Barbieri N, Zhang L, Adhikari P, Ramachandran R. Characterization of Avian Pathogenic Escherichia coli Isolated from Broiler Breeders with Colibacillosis in Mississippi. Poultry. 2023; 2(1):24-39. https://doi.org/10.3390/poultry2010004
Chicago/Turabian StyleJoseph, Jiddu, Madalyn Jennings, Nicolle Barbieri, Li Zhang, Pratima Adhikari, and Reshma Ramachandran. 2023. "Characterization of Avian Pathogenic Escherichia coli Isolated from Broiler Breeders with Colibacillosis in Mississippi" Poultry 2, no. 1: 24-39. https://doi.org/10.3390/poultry2010004