


Stimulating Role of Calcium and Cyclic GMP in Mediating the Effect of Magnetopriming for Alleviation of Salt Stress in Soybean Seedlings

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Treatment under Magnetic Field
2.3. Screening of Concentration of Different Modulators of cGMP and CaCl2
2.4. Early Seedling Growth Characteristics of Seedlings
2.5. Biochemical Analysis of Seeds
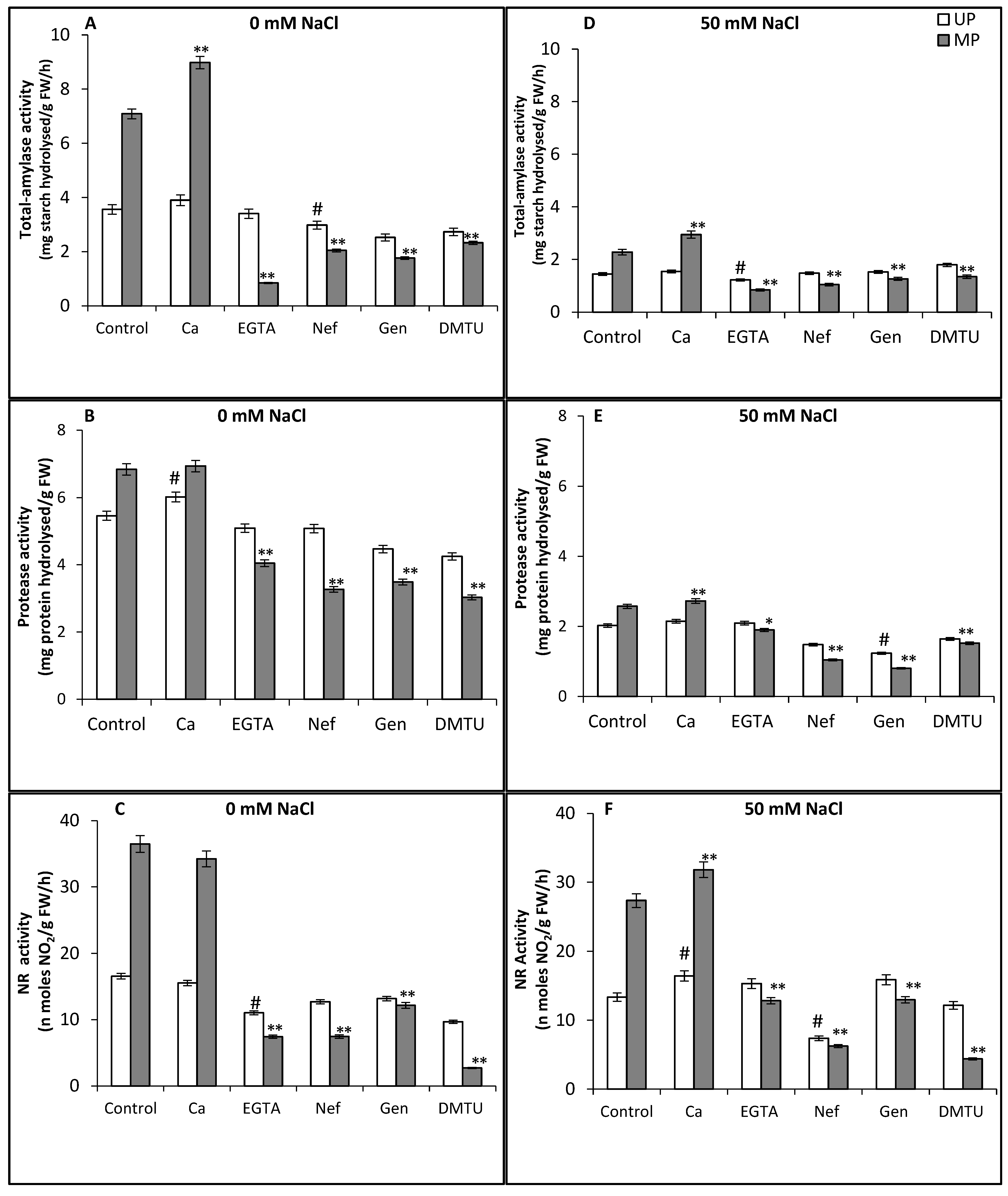
2.5.1. Total Amylase Activity
2.5.2. Protease Activity
2.5.3. Nitrate Reductase Activity
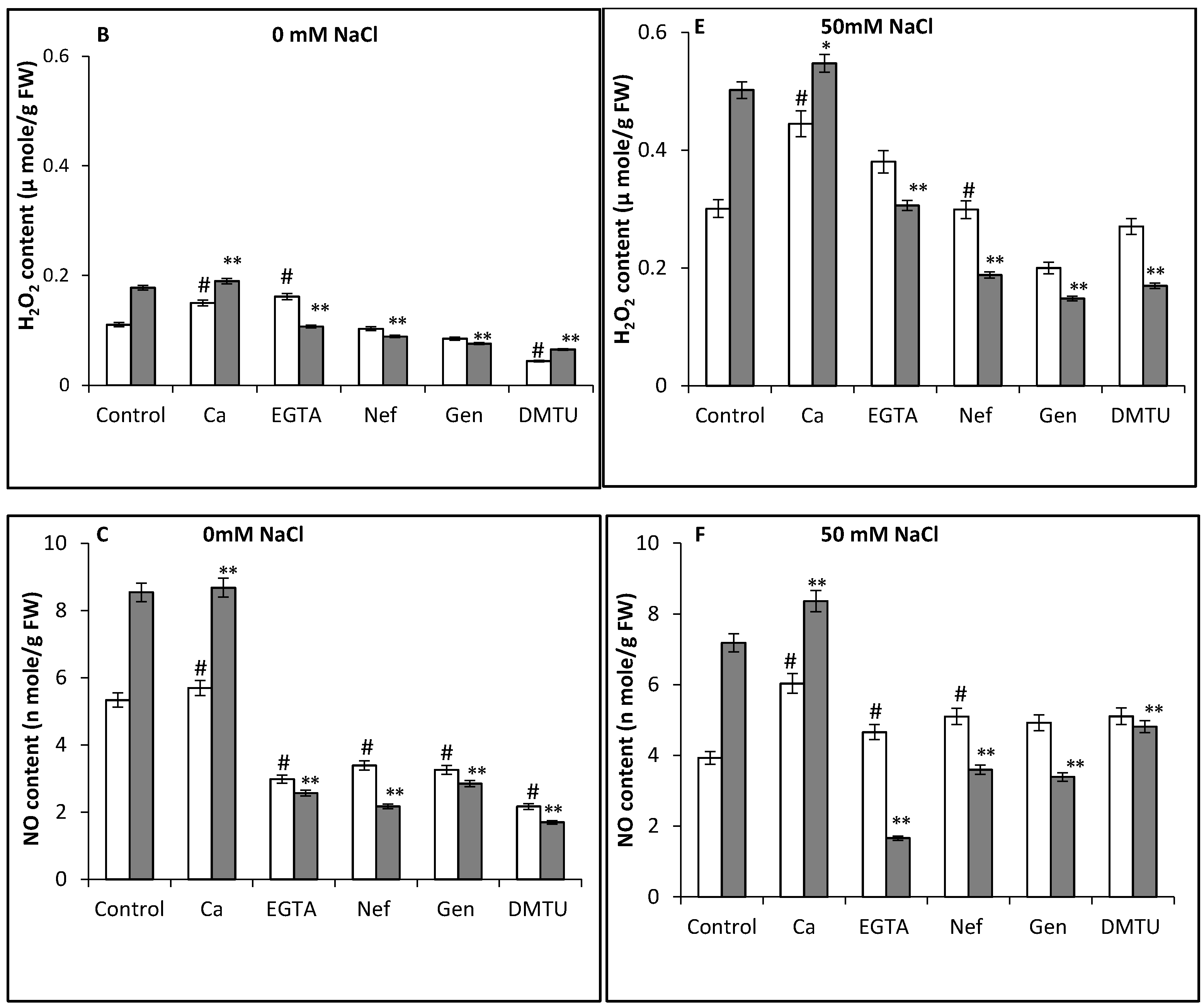
2.5.4. Estimation of Nitric Oxide
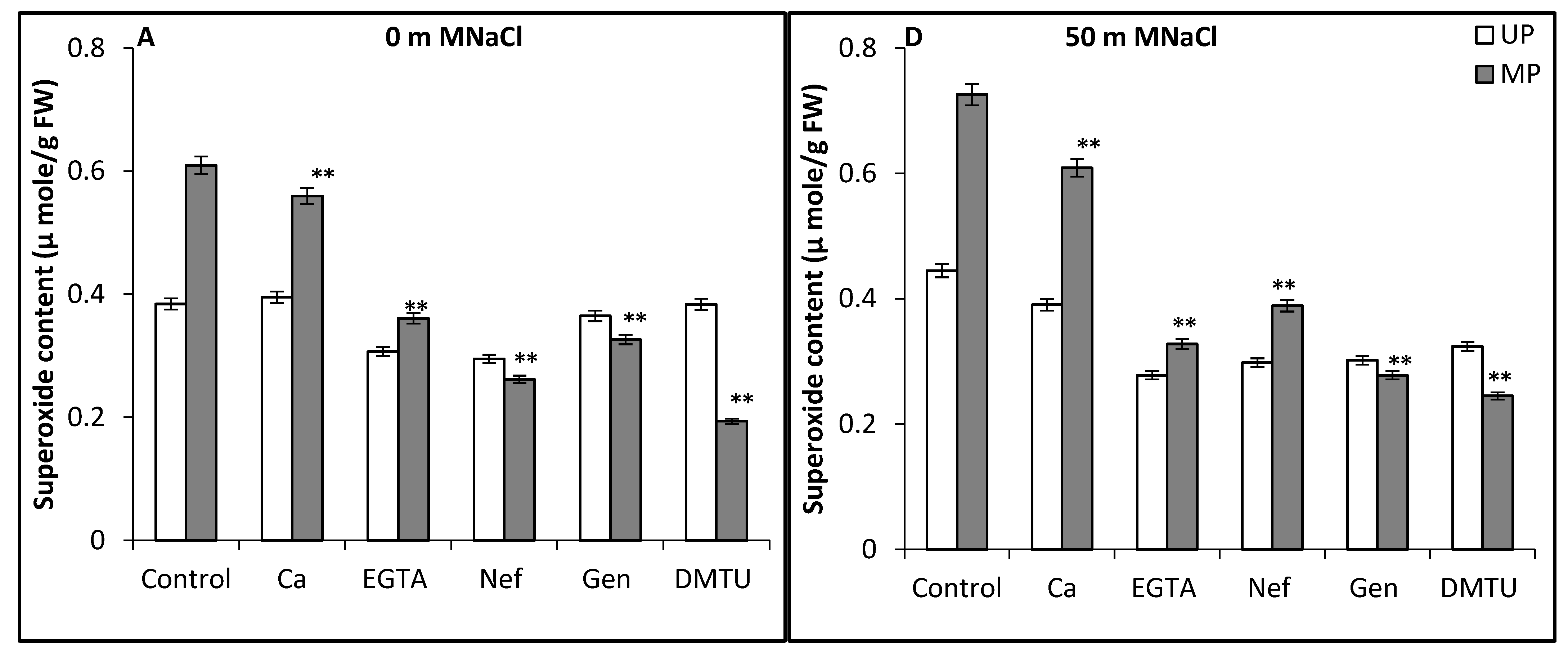
2.5.5. Estimation of ROS
Estimation of Superoxide Radical
Estimation of Hydrogen Peroxide
2.6. Statistical Analysis
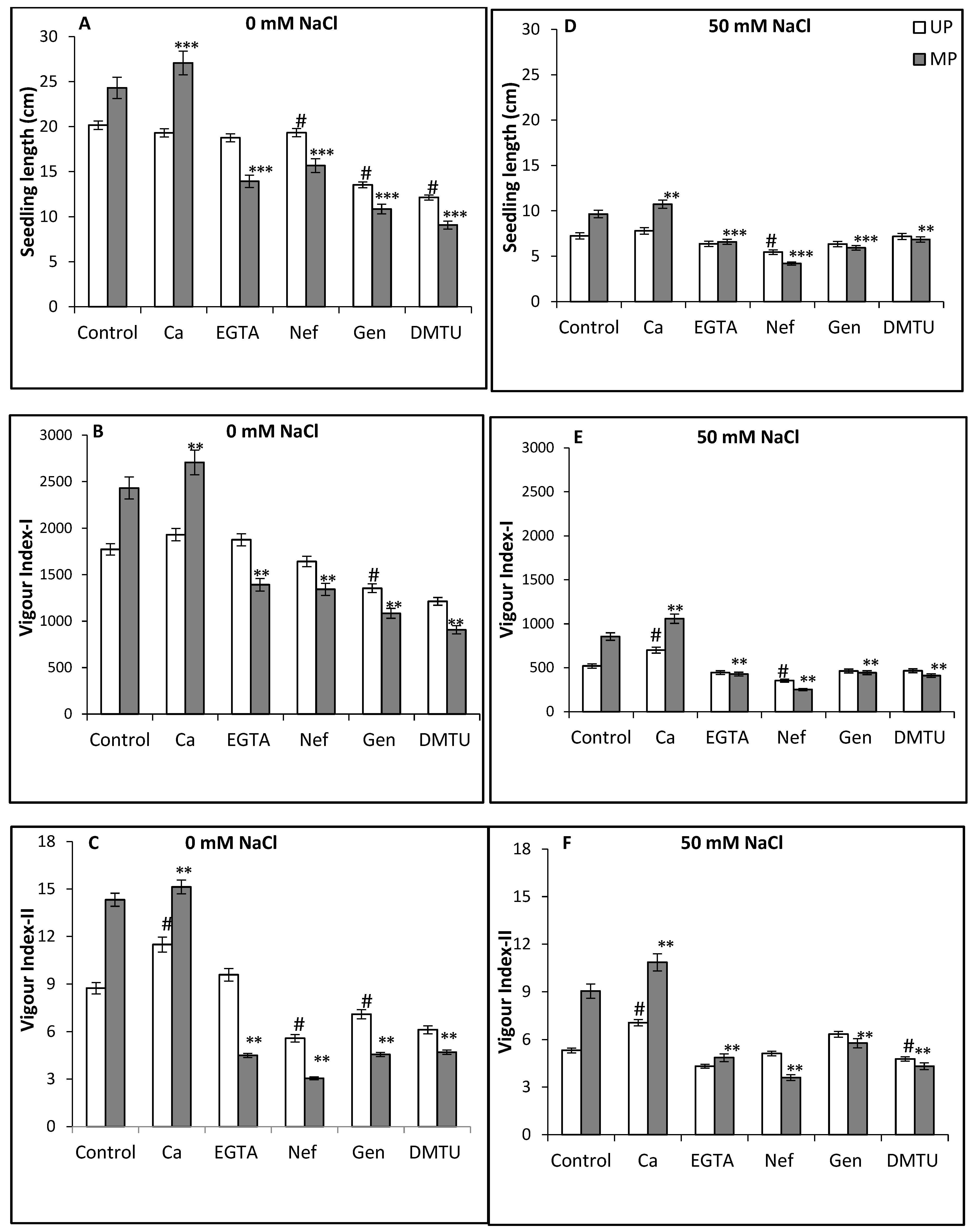
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kataria, S.; Verma, S.K. Salinity stress responses and adaptive mechanisms in major glycophytic crops: The story so far. In Salinity Responses and Tolerance in Plants: Targeting Sensory, Transport and Signaling Mechanisms; Springer: Berlin/Heidelberg, Germany, 2018; Volume 1, pp. 1–39. [Google Scholar]
- Kamran, M.; Parveen, A.; Ahmar, S.; Malik, Z.; Hussain, S.; Chattha, M.S.; Saleem, M.H.; Adil, M.; Heidari, P.; Chen, J.T. An overview of hazardous impacts of soil salinity in crops, tolerance mechanisms, and amelioration through selenium supplementation. Int. J. Mol. Sci. 2020, 21, 148. [Google Scholar] [CrossRef] [PubMed]
- Tomar, R.S.; Kataria, S.; Jajoo, A. Behind the scene: Critical role of reactive oxygen species and reactive nitrogen species in salt stress tolerance. J. Agron. Crop. Sci. 2021, 207, 577–588. [Google Scholar] [CrossRef]
- Ibrahim, E.A. Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 2016, 192, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Venâncio, C.; Pereira, R.; Lopes, I. The influence of salinization on seed germination and plant growth under mono and polyculture. Environ. Pollut. 2020, 260, 113993. [Google Scholar] [CrossRef]
- Hajihashemi, S.; Skalicky, M.; Brestic, M.; Pavla, V. Cross-talk between nitric oxide, hydrogen peroxide and calcium in salt-stressed Chenopodium quinoa Willdat seed germination stage. Plant Physiol. Biochem. 2020, 154, 657–664. [Google Scholar] [CrossRef]
- Kubala, S.; Wojtyla, Ł.; Quinet, M.; Lechowska, K.; Lutts, S.; Garnczarska, M. Enhanced expression of the proline synthesis gene P5CSA in relation to seed osmopriming improvement of Brassica napus germination under salinity stress. J. Plant Physiol. 2015, 183, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kataria, S.; Jain, M. Magnetopriming alleviates adverse effects of abiotic stresses in plants. In Plant Tolerance to Environmental Stress, 1st ed.; Hasanuzzaman, M., Fujita, M., Oku, H., Islam, T.M., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 427–442. [Google Scholar]
- Waqas, M.; Korres, N.E.; Khan, M.D.; Nizami, A.S.; Deeba, F.; Ali, I.; Hussain, H. Advances in the concept and methods of seed priming. In Priming and Pretreatment of Seeds and Seedlings: Implication in Plant Stress Tolerance and Enhancing Productivity in Crop Plants; Hasanuzzaman, M., Fotopoulos, V., Eds.; Springer: Berlin/Heidelberg, Germany, 2019; pp. 11–41. [Google Scholar]
- Prajapati, R.; Kataria, S.; Jain, M. Seed priming for alleviation of heavy metal toxicity in plants: An overview. Plant Sci. Today 2020, 7, 308–313. [Google Scholar] [CrossRef]
- Sarraf, M.; Kataria, S.; Taimourya, H.; Santos, L.O.; Menegatti, R.D.; Jain, M.; Ihtisham, M.; Liu, S. Magnetic Field (MF) Applications in Plants: An Overview. Plants 2020, 9, 1139. [Google Scholar] [CrossRef]
- Menegatti, R.D.; De Oliveira, L.O.; Da Costa, Á.V.L.; Braga, E.J.B.; Bianchi, V.J. Magnetic field and gibberellic acid as pre-germination treatments of passion fruit seeds. Rev. Ciência Agrícola 2019, 17, 15–22. [Google Scholar] [CrossRef]
- Vashisth, A.; Nagarajan, S. Exposure of seeds to static magnetic field enhances germination and early growth characteristics in chickpea (Cicer arietinum L.). BioElectroMagnetics 2008, 29, 571–578. [Google Scholar] [CrossRef]
- Bhardwaj, J.; Anand, A.; Nagarajan, S. Biochemical and biophysical changes associated with magnetopriming in germinating cucumber seeds. Plant Physiol. Biochem. 2012, 57, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Reina, F.G.; Pascual, L.A. Influence of a stationary magnetic field on water relations in lettuce seeds. Part I: Theoretical considerations. BioElectroMagnetics 2001, 22, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Vashisth, A.; Joshi, D.K. Growth characteristics of maize seeds exposed to magnetic field. Bioelectromagnetics 2017, 38, 151–157. [Google Scholar] [CrossRef]
- Poinapen, D.; Brown, D.C.; Beeharry, G.K. Seed orientation and magnetic field strength have more influence on tomato seed performance than relative humidity and duration of exposure to non-uniform static magnetic fields. J. Plant Physiol. 2013, 170, 1251–1258. [Google Scholar] [CrossRef]
- Konefał-Janocha, M.; Banaś-Ząbczyk, A.; Bester, M.; Bocak, D.; Budzik, S.; Górny, S.; Cholewa, M. The effect of stationary and variable electromagnetic fields on the germination and early growth of radish (Raphanussativus). Pol. J. Environ. Stud. 2018, 28, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Kataria, S.; Baghel, L.; Guruprasad, K. Pre-treatment of seeds with static magnetic field improves germination and early growth characteristics under salt stress in maize and soybean. Biocatal. Agric. Biotechnol. 2017, 10, 83–90. [Google Scholar] [CrossRef]
- Thomas, S.; Anand, A.; Chinnusamy, V.; Dahuja, A.; Basu, S. Magnetopriming circumvents the effect of salinity stress on germination in chickpea seeds. Acta Physiol. Plant. 2013, 35, 3401–3411. [Google Scholar] [CrossRef]
- Kataria, S.; Jain, M.; Tripathi, D.K.; Singh, V.P. Involvement of nitrate reductase-dependent nitric oxide production in magnetopriming-induced salt tolerance in soybean. Physiol. Plant. 2020, 168, 422–436. [Google Scholar] [CrossRef]
- Kataria, S.; Anand, A.; Raipuria, R.K.; Kumar, S.; Jain, M.; Watts, A.; Brestic, M. Magnetopriming actuates nitric oxide synthesis to regulate phytohormones for improving germination of soybean seeds under salt stress. Cells 2022, 11, 2174. [Google Scholar] [CrossRef]
- Jumrani, K.; Bhatia, V.S.; Kataria, S.; Alamri, S.; Siddiqui, M.H.; Rastogi, A. Inoculation with arbuscular mycorrhizal fungi alleviates the adverse effects of high temperature in soybean. Plants 2022, 11, 2210. [Google Scholar] [CrossRef]
- Kataria, S.; Baghel, L.; Jain, M.; Guruprasad, K.N. Magnetopriming regulates antioxidant defense system in soybean against salt stress. Biocatal. Agric. Biotechnol. 2019, 18, 101090. [Google Scholar] [CrossRef]
- De Souza, A.; Garcí, D.; Sueiro, L.; Gilart, F.; Porras, E.; Licea, L. Pre-sowing magnetic treatments of tomato seeds increase the growth and yield of plants. Bioelectromagnetics 2006, 4, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Li, R.; He, J.M. Magnetic field can alleviate toxicological effect induced by cadmium in mungbean seedlings. Ecotoxicology 2011, 20, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Baghel, L.; Kataria, S.; Guruprasad, K.N. Static magnetic field treatment of seeds improves carbon and nitrogen metabolism under salinity stress in soybean. Bioelectromagnetics 2016, 37, 455–470. [Google Scholar] [CrossRef] [PubMed]
- Baghel, L.; Kataria, S.; Guruprasad, K.N. Effect of static magnetic field pre-treatment on growth, photosynthetic performance and yield of soybean under water stress. Photosynthetica 2018, 56, 718–730. [Google Scholar] [CrossRef]
- Hozayn, M.; EL-Mahdy, A.A.; Zalama, M.T. Magneto-priming for improving germination, seedling attributes and field performance of barley (Hordeum vulgare L.) under salinity stress. Middle East J. Agric Res. 2018, 7, 1006–1022. [Google Scholar]
- Kataria, S.; Jain, M.; Rastogi, A.; Brestic, M. Static magnetic field treatment enhanced photosynthetic performance in soybean under supplemental ultraviolet-B radiation. Photosynth. Res. 2021, 150, 263–278. [Google Scholar] [CrossRef]
- Aliniaeifard, S.; Shomali, A.; Seifikalhor, M.; Lastochkina, O. Calcium signalling in plants under drought. In Salt and Drought Stress Tolerance in Plants; Springer: Berlin/Heidelberg, Germany, 2020; pp. 259–298. [Google Scholar]
- Seifikalhor, M.; Aliniaeifard, S.; Shomali, A.; Azad, N.; Hassani, B.; Lastochkina, O.; Li, T. Calcium signaling and salt tolerance are diversely entwined in plants. Plant Signal. Behav. 2019, 14, 1665455. [Google Scholar] [CrossRef]
- Kudla, J.; Batistič, O.; Hashimoto, K. Calcium signals: The lead currency of plant information processing. Plant Cell 2010, 22, 541–563. [Google Scholar] [CrossRef]
- Bakeer, S.M. Effect of ammonium nitrate fertilizer and calcium chloride foliar spray on fruit cracking and sunburn of Manflouty pomegranate trees. Sci. Hortic. 2016, 209, 300–308. [Google Scholar] [CrossRef]
- Choi, W.G.; Hilleary, R.; Swanson, S.J.; Kim, S.H.; Gilroy, S. Rapid, long-distance electrical and calcium signaling in plants. Annu. Rev. Plant Biol. 2016, 67, 287–307. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.H.; Wang, W.; Gao, T.; Fang, X.; Gao, X.; Li, J.; Bu, H.; Mu, J. Calcium alleviates decreases in photosynthesis under salt stress by enhancing antioxidant metabolism and adjusting solute accumulation in Calligonum mongolicum. Conserv. Physiol. 2017, 5, cox060. [Google Scholar] [CrossRef]
- Xu, T.; Niu, J.; Jiang, Z. Sensing Mechanisms: Calcium Signaling Mediated Abiotic Stress in Plants. Front. Plant Sci. 2022, 13, 925863. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Liao, W. Hydrogen peroxide signaling in plant development and abiotic responses: Crosstalk with nitric oxide and calcium. Front. Plant Sci. 2016, 4, 230. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Oku, H.; Nahar, K.; Bhuyan, M.B.; Al Mahmud, J.; Baluska, F.; Fujita, M. Nitric oxide-induced salt stress tolerance in plants: ROS metabolism, signaling and molecular interactions. Plant Biotech. Rep. 2018, 12, 77–92. [Google Scholar] [CrossRef]
- Gohari, G.; Alavi, Z.; Esfandiari, E.; Panahirad, S.; Hajihoseinlou, S.; Fotopoulos, V. Interaction between hydrogen peroxide and sodium nitroprusside following chemical priming of Ocimum basilicum L. against salt stress. Physiol. Plant. 2020, 168, 361–373. [Google Scholar]
- Liu, L.; Huang, L.; Sun, C.; Wang, L.; Jin, C.; Lin, X. Cross-talk between hydrogen peroxide and nitric oxide during plant development and responses to stress. J. Agric. Food Chem. 2021, 33, 9485–9497. [Google Scholar] [CrossRef]
- Kataria, S.; Baghel, L.; Guruprasad, K.N. Alleviation of adverse effects of ambient UV stress on growth and some potential physiological attributes in soybean (Glycine max) by seed pre-treatment with static magnetic field. J. Plant Growth Regul. 2017, 36, 550–565. [Google Scholar] [CrossRef]
- Abdul-Baki, A.A.; Anderson, J.D. Vigour determination in soybean by multiple criteria. Crop. Sci. 1973, 10, 31–34. [Google Scholar]
- Sharma, S.S.; Sharma, S.; Rai, V.K. The effect of EGTA, calcium channel blockers (Lanthanum Chloride and Nifedipine) and their interaction with abscisic acid on seed germination of Brassica juncea cv. RLM-198. Ann. Bot. 1992, 70, 295–299. [Google Scholar] [CrossRef]
- Teng, Y.; Xu, W.; Ma, M. cGMP is required for seed germination in Arabidopsis thaliana. J. Plant Physiol. 2010, 167, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Deng, B.; Kejun, Y.; Zhang, Y.; Zuotong, L. Can heavy metal pollution defend seed germination against heat stress? Effect of heavy metals (Cu2+, Cd2+ and Hg2+) on maize seed germination under high temperature. Environ. Poll. 2016, 216, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Sawhney, S.; Toky, K.L.; Nanda, K.K. Changes in amylase activity during extension growth and floral induction in Impatiens balsamina a qualitative short day plant. Indian J. Plant Physiol. 1970, 13, 198–205. [Google Scholar]
- Kunitz, M. Crystalline soybean trypsin inhibitor: II. General properties. J. Gen. Physiol. 1947, 30, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Jaworski, E.G. Nitrate reductase assay in intact plant tissues. Biochem. Biophys. Res. Commun. 1971, 43, 1274–1279. [Google Scholar] [CrossRef]
- Zhu, J.K. Regulation of ion homeostasis under salt stress. Curr. Opin. Plant Biol. 2003, 6, 441–445. [Google Scholar] [CrossRef]
- Chaitanya, K.S.K.; Naithani, S.C. Role of superoxide, lipid peroxidation and superoxide dismutase in membrane perturbations during loss of viability in seeds of Shorea robusta (Gaertn. F.). New Phytol. 1994, 126, 623–627. [Google Scholar] [CrossRef]
- Mukherjee, S.P.; Choudhari, M.A. Implications of water stressed induced changes in the levels of endogenous ascorbic acid and H2O2 in Vigna seedlings. Physiol. Plant. 1983, 58, 166–170. [Google Scholar] [CrossRef]
- Tuna, A.L.; Kaya, C.; Ashraf, M.; Altunlu, H.; Yokas, I.; Yagmur, B. The effects of calcium sulphate on growth, membrane stability and nutrient uptake of tomato plants grown under salt stress. Environ. Exp. Bot. 2007, 59, 173–178. [Google Scholar] [CrossRef]
- Marschner, P.; Rengel, Z. Nutrient availability in soils. In Marschner’s Mineral Nutrition of Higher Plants; Academic Press: Cambridge, MA, USA, 2012; pp. 315–330. [Google Scholar]
- Murillo-Amador, B.; Troyo-Diéguez, E.; García-Hernández, J.L.; López-Aguilar, R.; Ávila-Serrano, N.Y.; Zamora-Salgado, S.; Rueda-Puente, E.O.; Kaya, C. Effect of NaCl salinity in the genotypic variation of cowpea (Vigna unguiculata) during early vegetative growth. Sci. Hort. 2006, 108, 423–431. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef]
- Scrase-Field, S.A.; Knight, M.R. Calcium: Just a chemical switch? Curr. Opin. Plant Biol. 2003, 6, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Parre, E.; Ghars, M.A.; Leprince, A.S.; Thiery, L.; Lefebvre, D.; Bordenave, M.; Richard, L.; Mazars, C.; Abdelly, C.; Savouré, A. Calcium signaling via phospholipase C is essential for proline accumulation upon ionic but not nonionic hyperosmotic stresses in Arabidopsis. Plant Physiol. 2007, 144, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Anil, V.S.; Sankara, K.R. Calcium-mediated signaling during sandalwood somatic embryogenesis. Role for exogenous calcium as second messenger. Plant Physiol. 2000, 123, 1301–1311. [Google Scholar] [CrossRef]
- Patel, N.T.; Vaghela, P.M.; Patel, A.D.; Pandey, A.N. Implications of calcium nutrition on the response of Caesalpinia crista (Fabaceae) to soil salinity. Acta Ecol. Sin. 2011, 31, 24–30. [Google Scholar] [CrossRef]
- Shine, M.B.; Guruprasad, K.N.; Anand, A. Effect of stationary magnetic field strengths of 150 and 200 mT on reactive oxygen species production in soybean. BioElectroMagnetics 2012, 33, 428–437. [Google Scholar] [CrossRef]
- Gidrol, X.; Lin, W.S.; Dégousée, N.; Yip, S.F.; Kush, A. Accumulation of reactive oxygen species and oxidation of cytokinin in germinating soybean seeds. Eur. J. Biochem. 1994, 224, 21–28. [Google Scholar] [CrossRef]
- Boucelha, L.; Djebbar, R.; Abrous-Belbachir, O. Vigna unguiculata seed priming is related to redox status of plumule, radicle and cotyledons. Funct. Plant Biol. 2019, 46, 584–594. [Google Scholar] [CrossRef]
- Sachdev, S.; Ansari, S.A.; Ansari, M.I.; Fujita, M.; Hasanuzzaman, M. Abiotic stress and reactive oxygen species: Generation, signaling, and defense mechanisms. Antioxidants 2021, 10, 277. [Google Scholar] [CrossRef]
- El-Maarouf-Bouteau, H.; Bailly, C. Oxidative signaling in seed germination and dormancy. Plant Signal. Behav. 2008, 3, 175–182. [Google Scholar] [CrossRef]
- Baghel, L.; Kataria, S.; Jain, M. Mitigation of adverse effects of salt stress on germination, growth, photosynthetic efficiency and yield in maize (Zea mays L.) through magnetopriming. Acta Agrobot. 2019, 72, 1757. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Zhang, Y.; Jia, H.; Bi, Y. cGMP regulates hydrogen peroxide accumulation in calcium-dependent salt resistance pathway in Arabidopsis thaliana roots. Planta 2011, 234, 709–722. [Google Scholar] [CrossRef]
- Li, J.-H.; Liu, Y.-Q.; Lu, P.; Lin, H.-F.; Bai, Y.; Wang, X.-C.; Chen, Y.-L. A signaling pathway linking nitric oxide production to heterotrimeric g protein and hydrogen peroxide regulates extracellular calmodulin induction of stomatal closure in Arabidopsis. Plant Physiol. 2009, 150, 114–124. [Google Scholar] [CrossRef]
- Wang, W.H.; Yi, X.Q.; Han, A.D.; Liu, T.W.; Chen, J.; Wu, F.H.; Dong, X.J.; He, J.X.; Pei, Z.M.; Zheng, H.L. Calcium-sensing receptor regulates stomatal closure through hydrogen peroxide and nitric oxide in response to extracellular calcium in Arabidopsis. J. Exp. Bot. 2012, 63, 177–190. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Cabrera, M.A.; Henríquez, M.J.; Contreras, R.A.; Morales, B.; Moenne, A. Cross talk among calcium, hydrogen peroxide, and nitric oxide and activation of gene expression involving calmodulins and calcium-dependent protein kinases in Ulva compressa exposed to copper excess. Plant Physiol. 2012, 158, 1451–1462. [Google Scholar] [CrossRef] [PubMed]
- Magdalena, A.; Jolanta, F.W. Nitric oxide as a bioactive signaling molecule in plant stress responses. Plant Sci. 2007, 172, 876–887. [Google Scholar]
- Wendehenne, D.; Durner, J.; Klessig, D.F. Nitric oxide: A new player in plant signaling and defence responses. Curr. Opin. Plant Biol. 2004, 7, 449–455. [Google Scholar] [CrossRef]
- Lamotte, O.; Gould, K.; Lecourieux, D.; Sequeira-Legrand, A.; Lebrun-Garcia, A.; Durner, J.; Pugin, A.; Wendehenne, D. Analysis of nitric oxide signalling functions in tobacco cells challenged by the elicitor cryptogein. Plant Physiol. 2004, 135, 516–529. [Google Scholar] [CrossRef]
- Penson, S.P.; Schuurink, R.C.; Fath, A.; Gubler, F.; Jacobsen, J.V.; Jones, R.L. cGMP is required for gibberellic acid-induced gene expression in barley aleurone. Plant Cell 1996, 8, 2325–2333. [Google Scholar] [CrossRef]
- Wu, M.; Wang, F.; Zhang, C.; Xie, Y.; Han, B.; Huang, J.; Shen, W. Heme oxygenase-1 is involved in nitric oxide and cGMP-induced α-Amy2/54 gene expression in GA-treated wheat aleurone layers. Plant Mol. Biol. 2013, 81, 27–40. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kataria, S.; Shukla, S.; Jumrani, K.; Jain, M.; Gadre, R. Stimulating Role of Calcium and Cyclic GMP in Mediating the Effect of Magnetopriming for Alleviation of Salt Stress in Soybean Seedlings. Seeds 2023, 2, 232-245. https://doi.org/10.3390/seeds2020018
Kataria S, Shukla S, Jumrani K, Jain M, Gadre R. Stimulating Role of Calcium and Cyclic GMP in Mediating the Effect of Magnetopriming for Alleviation of Salt Stress in Soybean Seedlings. Seeds. 2023; 2(2):232-245. https://doi.org/10.3390/seeds2020018
Chicago/Turabian StyleKataria, Sunita, Shruti Shukla, Kanchan Jumrani, Meeta Jain, and Rekha Gadre. 2023. "Stimulating Role of Calcium and Cyclic GMP in Mediating the Effect of Magnetopriming for Alleviation of Salt Stress in Soybean Seedlings" Seeds 2, no. 2: 232-245. https://doi.org/10.3390/seeds2020018