1. Introduction
The seed–seedling transition is a central point during the plant’s life cycle. Taking this into account, this transition process has been studied from different points of view in areas such as general botany, agronomy and physiology [
1]. Nevertheless, there are still many gaps in the knowledge that need to be further studied, and polyembryony is one of the most intriguing areas of research. The presence of more than one embryo per seed, called polyembryony [
2], is a common feature in angiosperms and it seems to be present in approximately 30% of the species studied so far. It has been associated with sporophytic apomixis (unreduced embryos originated from maternal tissues) or embryo cleavage (either apomictic or, more rarely, sexual embryos) [
3]. However, only a few species have been studied in detail regarding these embryos’ origins or the ecological consequences of this reproductive phenomenon, with most studies focusing on the effects of polyembryony on the competition for nutrients and space between embryos and seedlings originated from the same seed [
4,
5,
6,
7]. This is a type of intraspecific competition [
8] but with some particularities and important effects on the species’ biology.
The competition between embryos and/or seedlings from a polyembryonic seed will be treated in this paper as gemellar competition due to particularities such as: (i) the somatic and possibly clonal origin of the embryos inside a seed; and (ii) the limited space inside a seed, which leads to a reduction in the size of the embryos and seedlings as the number of embryos per seed increases [
4,
5,
6,
7]. Additional aspects such as morphological anomalies, differences in survival rates in embryos and seedlings, and competition among sexual and asexual embryos [
6,
7,
9]—with a posterior reduction in the competition effects on the seedlings in the long term [
4]—have also been reported as particularities of this type of competition.
Some studies have pointed out that polyembryony could promote positive effects. One of them is the Allee effect, described as the mutual collaboration of seedlings while occupying patches [
10]. Alternatively, polyembryony would also promote the bet-hedging effect, since the reduction in size and possible decrease in the fitness of polyembryonic seedlings would be compensated by the possibility that at least one of them would survive during germination in stressful conditions [
5,
10,
11]. The positive effect of the increased chance of survival of at least one gemellar seedling per seed would result in greater individual seed survival [
7]. However, the effect of gemellar competition is absent from some studies, hindering the creation of models for greater generalizations or future hypotheses on germination and plant development from polyembryonic seeds.
Handroanthus chrysotrichus (Mart. ex DC.) Mattos is a polyploid, self-compatible, apomictic, and polyembryonic tree and is one of the twenty-seven known
Handroanthus species of Bignoniaceae [
7,
12,
13,
14]. The species is amply used as ornamental trees—known as “golden trumpet trees”—in urban environments due to their magnificent flowers [
7,
12,
13,
14]. Since it produces a lot of fruits and viable seeds, it seems to be a good model for studies on polyembryony. Like other native species used for ornamentation, its distribution is not clearly known given the low sampling in cities; however, polyembryony screenings are available. All plants studied so far, either natural or cultivated, are polyembryonic [
15], which could save time in population selection. Their seeds also show germinability ranging between 25 and 87%, with intraspecific variability [
16,
17,
18,
19,
20]. This species produces up to seven embryos and five seedlings per seed [
7] making it easy to compare seeds with different numbers of embryos and seedlings. Previous data have shown that the higher the number of embryos per seed, the lower the mass of each embryo and seedling and of seedling length, and the higher the mortality of individual seedlings and the frequency of atypical embryos [
7]. In addition, the mean seedling number emerged from one seed is also affected by the temperature during the emergence process [
21]. Despite this basic knowledge, various previous studies ignored the effect of polyembryony on seedling success during the seed–seedling transition [
22].
Taking this into account, it is clear that an increase in the number of gemellar embryos can affect the seed–seedling transition; however, it is not clear how and why this occurs. Therefore, we build the hypothesis that intraspecific gemellar competition affects the seed–seedling transition by affecting the dynamics, uniformity and synchrony of seed germination and of the seedling emergence processes. As a consequence, the morphometry of seedlings may be negatively affected, and early plant establishment hindered. In this context, our expectation is to provide data that will contribute to further the understanding of the impact of polyembryony on the seed biology and early plant development of Handroanthus chrysotrichus.
2. Materials and Methods
2.1. Sampling and Study Site
Handroanthus chrysotrichus is a Brazilian tree that occurs naturally in the “Mata Atlântica” and disturbed forest areas, but which is also described in the Cerrado Biome [
23], Brazil’s Neotropical Savanna [
24]. It is also widely cultivated for ornamental use in urban areas [
23]. Its origin, natural ecology, invasive and native actual distribution are unknown.
Seeds of five mother plants were collected in 2005, in an urban area of Uberlândia, Minas Gerais State, Brazil (18°53′6.64″ S, 48°15′36.23″ W). The climate region is the Aw Megathermic type according to the updated classification of Köppen–Geiger [
25], a tropical wet climate with a dry winter (April to September) and a wet summer (October to March) (see climate diagram in Ranal [
26]). The sample was deposited in the Herbarium Uberlandense (HUFU 52589). The origin and cultivation history of these mother plants are unknown. The data related to the embryo mass, number of embryos and seedlings formed per seed were reanalyzed from the original data obtained by Mendes-Rodrigues et al. [
7].
2.2. Seed Water Content and Mass
Eight replicates per mother plant were used to evaluate the water content of the seeds. Each replicate had approximately 500 mg of seeds (mean = 490.38, standard deviation = 29.46, n = 40). The number of seeds per replicate and per mother plant was 32 for mother plants four and five, 38 for mother plants one and two and 44 for mother plant three. The water content was obtained after keeping the seeds in a stove at 55 °C for three days, and then at 105 °C for 10 days. Before weighing, the seeds were kept in plastic boxes with dry silica gel until they returned to room temperature. The replicates were weighed individually on an analytical scale (AA250, precision 0.1 mg, Denver Instrument, Bohemia, NY, USA), before and after drying. The water content was calculated on a wet weight basis. These replicas were also used to estimate the fresh and dry mass of the seeds.
2.3. Seed Germination and Seedling Emergence Assays
The germination process was recorded from seeds sown on vermiculite (expansion volume of 0.1 m
3) moistened with distilled water at field capacity, and kept in germination plastic boxes in a germination chamber (MPG-2000, Seedburo Equipment Company, Batavia, USA) under continuous fluorescent light and a mean minimum temperature of 24.53 ± 0.24 °C (mean ± standard deviation;
n = 17) and a mean maximum temperature of 25.33 ± 0.20 °C (mean ± standard deviation;
n = 17). We used this temperature as it offered the best results for most Cerrado species [
6,
7,
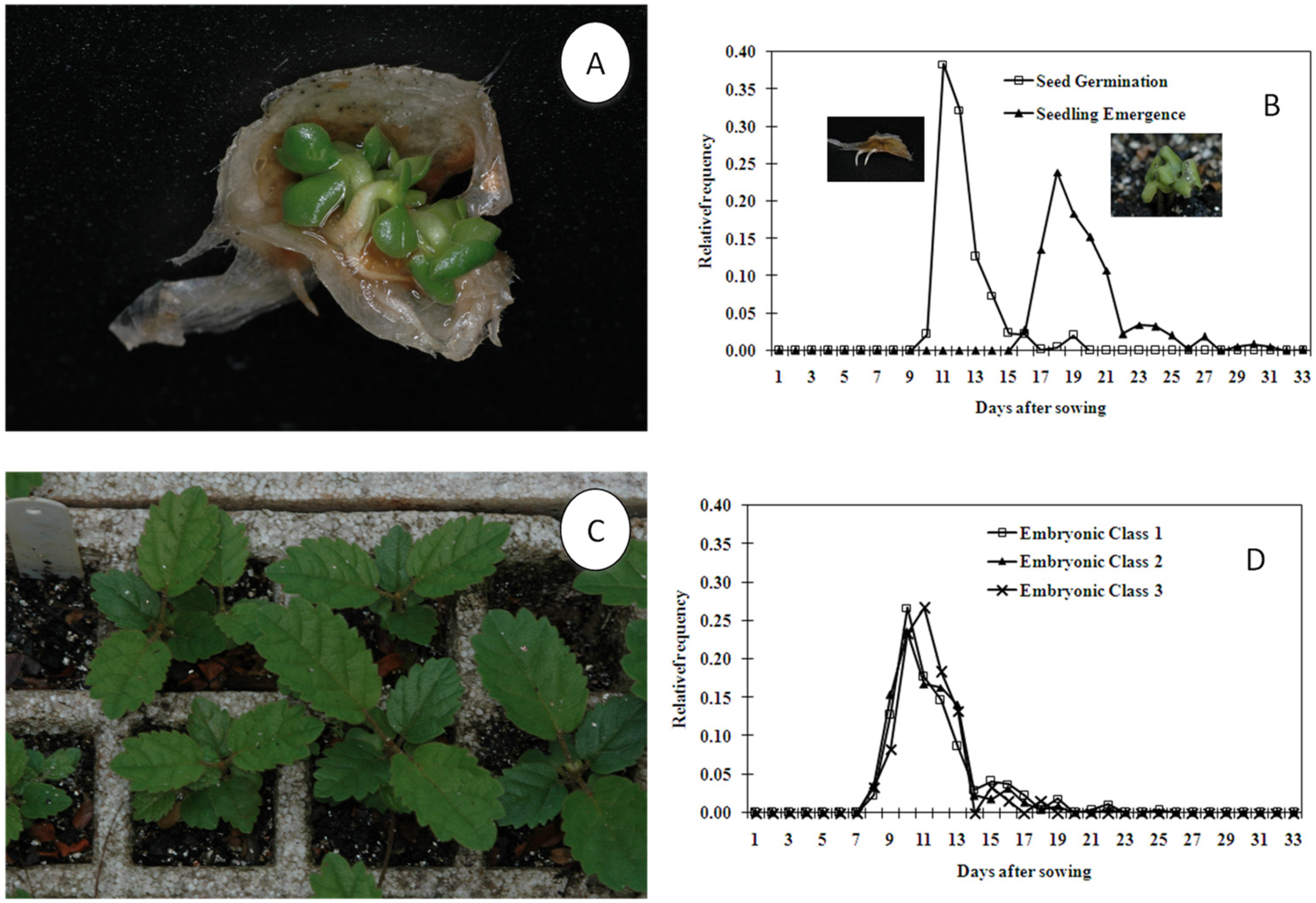
9]. The seed was considered germinated when the first radicle protruded from the seed, since the species is polyembryonic (
Figure 1A,B).
For seedling emergence, seeds were sown in polystyrene multicell boards (cell volume of 34 cm
3) with one seed per cell at ca. 0.5 cm depth, in a 1:1 mixture of commercial substrate (Plantmax
®, Eucatex Mineral Ltda, São Paulo, SP, Brazil) and vermiculite (expansion volume of 0.1 m
3). The trays were kept in an open area about 100 m away from the mother plants, under natural light conditions, and moistened at field capacity. The appearance of any part of the first seedling above the substrate’s surface was used as emergence criterion, since the species is polyembryonic (
Figure 1B,C).
Both seed germination and seedling emergence assays were conducted with small samples (
n = 32 seeds per replicate). The number of seeds germinated and seedlings emerged was recorded daily, at the same hour. No experiment was censored (break off). Observations continued until all seeds germinated or died. For each replicate, germinability or percentage of emergence, mean germination or emergence time, time to first and last seed germination or seedling emergence, coefficient of variation of germination or emergence time, uncertainty and synchrony were calculated according to Ranal and Santana [
27]. Further details of these germination measurements can be found in Ranal and Santana [
27], and Ranal et al. [
28]. The relative frequency of seed germination and seedling emergence was also calculated, grouping all 20 replicates in one sample based on the absence of intraspecific variability in seed-germination measurements [
29].
2.4. Polyembryony Indicators
Each seed germinated was dissected to count the number of embryos, as some could be hidden or did not develop completely inside the seeds. These data were used to calculate the polyembryonic seed percentage (PSP), that is, the percentage of seeds with more than one embryo, and the mean embryo number per seed (MENS). We sampled 124, 118, 120, 125, and 119 seeds for mother plants one to five, respectively. We evaluated only the germinated seeds.
To estimate the total number of seedlings emerged per seed, the observations of seedling emergence were conducted until the number of seedlings effectively emerged per each seed had stabilized. That is needed since seedlings from a polyembryonic seed have different times of emergence. In general, stabilization occurs about seven days after the emergence of the first seedling. During the first seven days, we did not observe any seedling mortality. Seedling mortality was also evaluated during the experiment (see results in Mendes-Rodrigues et al. [
7]). We conducted these observations on a daily basis because seedling mortality during the experiment could cause an underestimation of the analysis of the effective number of seedlings produced per seed (see Mendes-Rodrigues et al. [
7]). From these data, we calculated the gemellar seed percentage (GSP)—i.e., the percentage of seeds with more than one seedling emerged—and the mean seedling number per seed (MSNS). We sampled 117, 116, 91, 118, and 121 seeds for mother plants one to five, respectively. We evaluated only the seeds with emerged seedlings.
Additionally, we estimated the percentage of embryos that were converted into seedlings, treated here as Embryo Seedling Turnover Percentage (ESTP) [
7]. For this, we used the following mathematical expression: ESTP = [(MSNS × NSES)/(MENS × NSS)] × 100, where MSNS: mean seedling number per seed, NSES: number of seeds with emerged seedlings, MENS: mean embryo number per seed, and NSS: number of seeds sowed. The ESTP was calculated for each mother plant and species (independently of the mother plant).
We also classified the seeds in seed embryonic classes (SEC) and seedling gemellar classes (SGC). We found from one to seven SEC, containing from one to seven embryos per seed (SEC1 to SEC7), and found from one to five SGC containing from one to five seedlings per seed (SGC1 to SGC5). The SGCs are named differently in the literature, as singlet, doublet, triplet, quadruplet, and quintuplet, and respectively refer to SGC1 to SGC5.
After the classification of the seeds from the emergence experiment in the SGCs, we calculated the seed-emergence measurements for the seedling gemellar classes singlet, doublet and triplet (except the percentage of emergence). In this analysis, due to the low frequency of each SGC in the original replicates from the seed-emergence experiment, each mother plant was considered to be one replicate for each SGC. In this case, all seeds of each SGC of the four replicates of each mother plant were grouped. SGC4 and SGC5 did not present enough replicates for the calculations of seed-emergence measurements.
2.5. Gemellar Competition
We measured seedling mass just after germination (after the first emission of radicle—
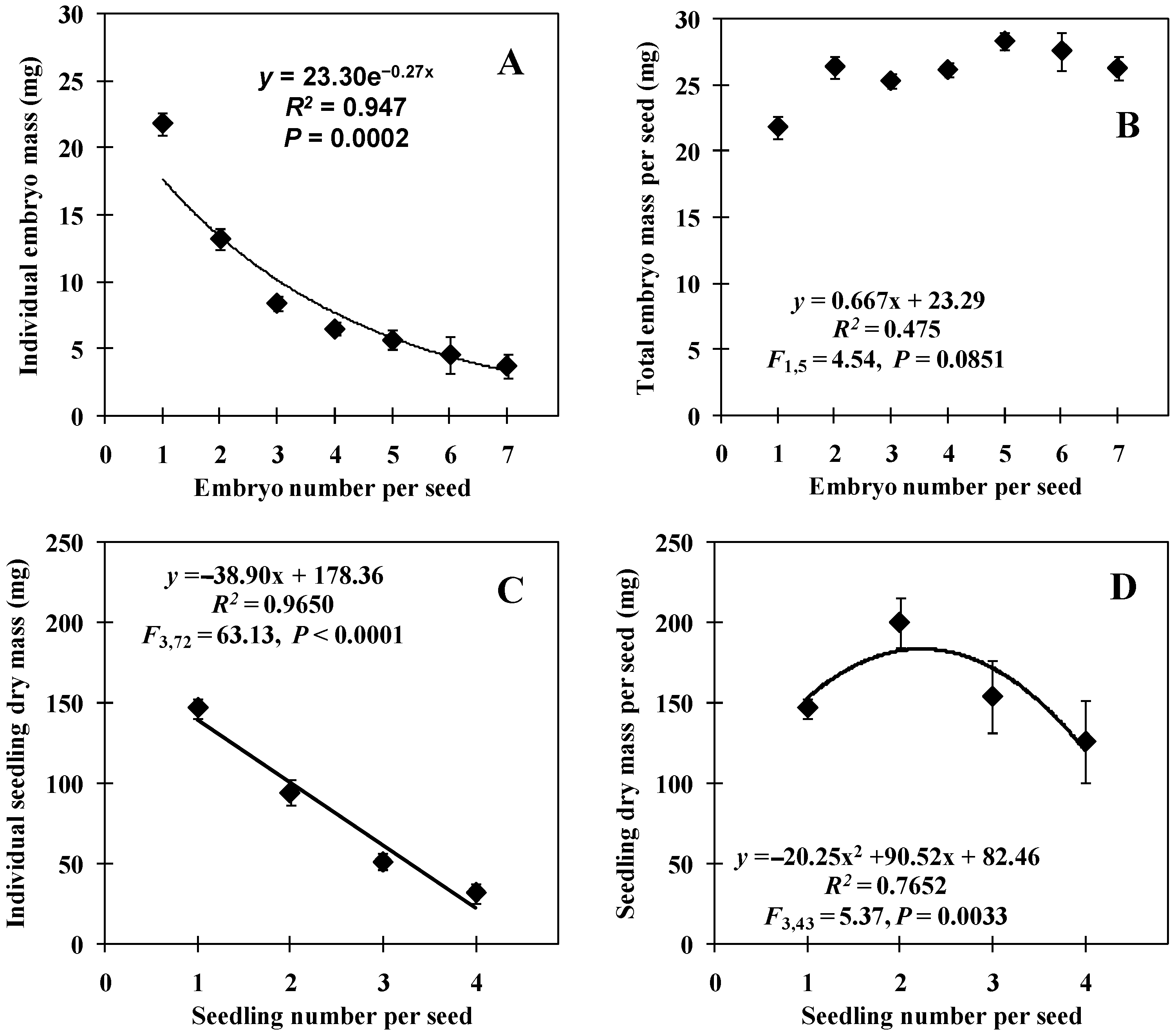
Figure 1A) as a direct estimate of embryo mass, because it was practically impossible to isolate the multiple embryos present in a non-imbibed seed without fragmenting them and causing their loss of individuality. Germinated seeds were dissected, and each embryo (now a young seedling) was weighed individually on an analytical scale (AA250, precision 0.1 mg, Denver Instrument, Bohemia, NY, USA). Sample size of each SEC was SEC1 = 50, SEC2 = 106, SEC3 = 183, SEC4 = 132, SEC5 = 65, SEC6 = 12, and SEC7 = 21. The sum of the mass of all embryos of each seed was also calculated, and the sample size of each SEC was SEC1 = 50, SEC2 = 53, SEC3 = 61, SEC4 = 33, SEC5 = 13, SEC6 = 2, and SEC7 = 3. These data were originally presented by Mendes-Rodrigues et al. [
7].
After 54 days of sowing, one sample of seedlings of each SGC was exhumed and measured. We adopted 54 days after sowing as a time marker since we did not correct growth with the time of emergence of each seedling. Moreover, after 54 days, emergence does not occur anymore and the seedling stand is homogenous. We sampled only seedlings of SGC with no mortality in a given seed up to 54 days. The survival percentages were presented by Mendes-Rodrigues et al. [
7]. At this time (54 days after sowing,
Figure 1C) the seedling size permitted hand manipulation without apparent damages. After 54 days of sowing the probability of mortality of the seedlings increased (personal observation). We measured 25 singlets, 20 doublets, 15 triplets, and 16 quadruplets. The gemellar class quintuplet was not included because only one seed from this class appeared in the sample. The analyses of seedling morphometry were not performed separately for each mother plant due to the low frequency of some SGCs at this time. The quadruplets were also not present in all mother plants analyzed.
For each seedling, we recorded the number of leaflets (NL), stem height (SH, from the collar to the apical meristem; the collar is characterized by a separation between the chlorophyllous hypocotyl and the achlorophyllous root) and stem diameter (SD, or diameter of collar). We also evaluated the aerial seedling dry mass (ASDM, or aerial mass part) above the collar, and the subterranean seedling dry mass (SSDM, or subterranean mass part) below the collar. The evaluations were performed using a digital scale (AA250, precision 0.1 mg, Denver Instrument, Bohemia, NY, USA). The seedlings were kept in paper bags at 70 °C until they reached constant dry mass (after 7 days), and 2 to 4 h afterwards in hermetic boxes with dry silica gel until they reached room temperature, before mass evaluations. We calculated the ratio between aerial seedling dry mass and the total seedling dry mass (aerial/seedling dry mass ratio, ASDMR). This ratio was used to evaluate seedling resource allocation. We also calculated the biomass production of each seed individually, treated here as the sum of the biomasses of all seedlings produced per seed, as proposed by Blanchard et al. [
5], in order to evaluate the density-dependent facilitation of polyembryonic seedlings, or Allee effects.
2.6. Mass Prediction Model for Seedlings
We applied multiple linear regression models based on the least square method to know if the seedlings’ morphometric measurements could predict the aerial and subterranean dry mass of 54-day-old seedlings. This model was proposed to estimate the seedling mass in a seedling competition experiments. We adopted the following traits as independent variables: NL, SH, SD, and density (here considered as the number of seedlings in the SGC) in a model without a constant (or intercept). We sampled these variables in three SGCs (SGC1 = 25, SGC2 = 20, SGC3 = 15) with a total of n = 60 seedlings. SGC 4 and 5 were not included due to their low occurrence in the studied sample. SGC4 also showed high seedling mortality at 54 days. The aerial and subterranean dry mass were considered to be the dependent variables.
2.7. Statistical Analysis
For the germination and emergence experiments, the assumptions of ANOVA were tested. We tested residuals normality using the Kolmogorov–Smirnov Lilliefors test and the homoscedasticity of variances using Levene’s test. When both assumptions were accepted, the data were compared using ANOVA and Tukey’s test. When at least one of the assumptions was not met, the data were compared using the Kruskall–Wallis and Dunn’s test. Data on seed mass and water content were tested with Generalized Linear Models. For seed mass and water content, we adopted the Gaussian and Gamma distribution function, respectively, and the identity link function for both. The Least Significance Difference test was used for multiple comparisons.
Polyembryonic indicators were tested with Generalized Linear Models. For the number of embryos per seed and number of seedlings per seed, we adopted the Poisson distribution function and the logarithmic link function. For the polyembryonic seed percentage, gemellar seed percentage, and embryo seedling turnover percentage, we used the Binomial distribution and identity link functions. Again, the Least Significance Difference test was used for multiple comparisons.
Initially, to test the effect of the mother plant on seedling morphometry, we performed a two-way ANOVA. For the dry-mass analysis of 54-day-old seedlings, the first factor was the mother plant and the second factor was the seedling gemellar class (singlet, doublet and triplet). In this analysis, the mother plants did not show differences (F4,59 = 0.91, p = 0.4643), the seedling gemellar classes showed differences (F2,59 = 50.87, p < 0.0001) and the interaction between these two factors was not significant (F8,59 = 1.28, p = 0.2791). After this, we did not test the effect of the mother plant in the other tested models. In addition, we found low frequency of some gemellar classes in some mother plants, which hindered their use in some of the models.
Embryo-mass data were adjusted to an exponential regression model, and the total mass data of embryos per seed were adjusted to a linear regression model by the ordinary least square method. The model significance was tested using a one-way ANOVA. The effects of initial gemellar competition in seedling morphometry were compared between embryonic classes with a one-way ANOVA. For this, the number of seedlings per seed was used as an independent factor and the seedling traits as a dependent factor. These data were adjusted to linear regression models for all traits, except for the total seedling mass per seed, which was better adjusted to a quadratic regression model. With this, the effect of polyembryony or gemellar competition was inferred by the inclination (or slope) of the linear regression model (see details in Mendes-Rodrigues et al. [
6]) whenever a linear regression was used. In these analyses, we included the embryonic classes from one to four seedlings per seed. The number of replicates for each embryonic class were SEC1 = 25, SEC 2 = 20, SEC 3 = 15, and SEC 4 = 16, except for seedling dry mass per seed (SEC1 = 25, SEC2 = 10, SEC3 = 5, and SEC4 = 4).
The density or seed gemellar class was considered a Dummy variable in the mass prediction model (a discrete quantitative variable that represents binary data and can take on only two values, 1 or zero, where 1 represents the presence of a qualitative attribute and zero the absence), since density is an integer. To adjust the mass prediction model, the presence of multicollinearity between the independent variables was tested by the VIF (variation inflation factor), and variables were excluded from the models if VIF > 4. To evaluate the contribution of each variable to the model, the Partial F test was used with a significance of 0.10. The significance of the parameter estimate was tested with Student’s t-test. The residuals of the regression model were tested for normality by the Shapiro–Wilk test. The homogeneity of variances was tested with Bartlett’s test using density as a factor of formation of the groups. The independence of the residuals was tested by the Durbin–Watson test. The tested models were evaluated without the intercept since the presence of the intercept has no biological justification in this case. Multicollinearity was observed among the predictor variables for seedling mass. The VIF value for the stem diameter was higher than four (VIF = 5.54), and the variable was excluded from the initial model. After exclusion, the multicollinearity analysis was performed again and no VIF value above four was obtained (stem height VIF = 1.44, seedling density VIF = 1.59 and number of leaflets VIF = 1.23).
For the analyses where significance was not declared, 0.05 was adopted. The analyses were performed in the Statistical Package SPSS 20.0 or in the R environment [
30].
4. Discussion
We showed here that polyembryony affects the seed–seedling transition by altering seedling morphometry and possibly vigor. This occurs more as a consequence of high intraspecific competition (gemellar competition) from the number of embryos in each seed than from intraspecific variability in the mother plants. In general, gemellar competition from two-or-more embryos per seed causes distress in the seed–seedling transition. This behavior is explained by the fact that the size of the embryos and seedlings decreased with the increase in their number per seed, which resulted in a reduction in the individual seedlings’ vigor, therefore affecting the emergence process. Only seeds that produced two seedlings per seed showed a higher total mass of seedlings per seed, evidencing a positive Allee effect.
In general, functional and physical traits are correlated in the transition from seed to seedling [
31,
32]. This is the most studied aspect of the early plant development [
33]. Thus, we were surprised when we observed that physical traits had intraspecific variability although the mother plants made no differential contribution to the functional traits. This was the first clue to how polyembryonic seeds (the majority in all samples studied) can alter the early plant development of
H. chrysotrichus. Taking this into account, we considered polyembryony could be a potential cause of distress for the species. To prove this hypothesis, we observed the way that intraspecific competition from gemellar status affects seed germination, seedling emergence and morphometry.
First, we observed that the high capacity, uniformity, and dynamics of the seed–seedling transition follows a species-specific pattern during seed germination and seedling emergence. However, seedling emergence is the key aspect of the seed–seedling transition since it is less efficient and homogeneous, as pointed out by the results. In fact, it is expected that the seed germination process is less variable than the process of seedling emergence, growth, development and establishment [
34,
35,
36,
37]. However, when more than one embryo is contained in a seed, gemellar competition can alter this expected pattern. Seed germination is an amphibolic event in which enzymatic processes and physical forces overlap, promoting embryo growth until it finally breaks through the teguments, allowing the full embryo’s protrusion [
32,
38,
39,
40,
41]. Therefore, more than one embryo in a same seed can provoke a double and conflicting effect in the seed–seedling transition process, increasing physical force to overcome teguments, but reducing reserve material for the optimal development of each embryo. From a physiological point of view, this double effect can play a central role as a trade-off, impacting the time, velocity and uniformity of protrusion and, therefore, the dynamics of seedling emergence. Therefore, the presence of polyembryony can function as a distress agent (sense Kranner et al. [
42]) in the early plant development based on the trade-off of seed germination and seedling emergence. Our findings showed this for
H. chrysotrichus. The increase in the number of embryos in a seed reduces embryo mass and, consequently, stem height and leaf number in seedlings. Those measurements are related to seedling vigor and, thus, with the seedling’s ability to establish itself in a non-optimal environment (see Finch-Savage and Bassel [
43]; Reed et al. [
44]). However, total seedling mass was optimal when two embryos were contained in a seed. Three embryos per seed led to a reduction in individual seedling mass, making the emergence process less uncertain but rapid. These are characteristics which have been identified as an escape strategy during the seed–seedling transition in order to overcome stress [
43,
44].
On the other hand, the absence of intraspecific variability in the seed germination and seedling-emergence measurements of
H. chrysotrichus may be related to the occurrence of mother plants in a restricted and homogeneous urban area. There are records of germinability of the species ranging between 20 and 87% in 17 individuals in an urban area in Jaboticabal city, São Paulo State, Brazil [
20]. Another issue that cannot be overlooked is that the apomixis present in the species can generate low genetic variability or clonality, as observed in other Cerrado species [
45]. Nevertheless, studies have demonstrated the presence of intraspecific variability in the germination measurements of apomictic species in Cerrado [
46,
47,
48], and for vegetative and reproductive characters of sexual and apomictic populations of the congeneric
H. ochraceus [
49]. However, the number of individuals needed to characterize the variability in Cerrado Bignoniaceae can be very high, variable and dependent on the reproductive system, with the sample size ranging from 7 to 212 individuals analyzed [
49,
50,
51]. In addition, even in apomictic populations with a low sampling, it is possible to detect intraspecific variability in some traits, as observed in our case.
Few studies have evaluated the type of germination measurements described here for other Cerrado Bignoniaceae [
52,
53] (
Table 7), or even for other biomes. The seed germination behavior of
H. chrysotrichus was similar to the one observed in
H. serratifolius, another polyembryonic species [
7], but distinct from that of
J. cuspidifolia, which has low polyembryony (below 5%), a low germination percentage, and a higher coefficient of variation of germination time (
Table 7).
As for seedling emergence,
H. chrysotrichus showed marked differences from
Tabebuia roseoalba (Ridl.) Sandwith, which experiences a highly synchronous germination in a short period of time, and from
J. cuspidifolia Mart, which has a low synchrony, a long germination times and dormancy—when germination takes longer than 30 days [
52,
53] (
Table 7). Both species have low polyembryony. Despite scarce data, the general emergence and germination pattern of Bignoniaceae seems to be a short germination time, a synchronous process and low variability.
Species with a low polyembryonic rate (lower than 5%) are commonly found in the Cerrado [
54] although they probably behave functionally as monoembryonic species. The role of polyembryony in these species and its relationship with seed biology has yet to be elucidated as it also occurs in Bignoniaceae [
52], which prevents further discussions. Polyembryonic individuals from the non-Cerrado Bignoniaceae species
Dolichandra unguiscati (L.) L.G. Lohmann, have a seedling emergence time of more than two months [
55], probably showing another case of dormancy in the group. Long and short germination and emergence times also occur in species with low polyembryony frequencies, as in
J. cuspidifolia (long times) and in
T. roseoalba (short times). Apparently, polyembryonic species are not associated with variations in the germination and emergence times.
Handroanthus chrysotrichus has commonly high polyembryony [
7] and does not present any atypical behavior in germination and or emergence compared to other species of the family. None of the monoembryonic populations of
H. serratifolious [
56], the monoembryonic and polyembryonic populations of
H. ochraceus [
49], or even the other polyembryonic species of the family [
57], have been studied for germination measurements. These data could elucidate the role of polyembryony in the group. Some studies have related the seed-germination measurements to polyembryonic expression or the existence of monoembryonic or polyembryonic populations [
6,
46,
49]. In Bignoniaceae, more data are still needed for the group to allow such relationships, probably requiring phylogenetic corrections or the study of populations with differences in polyembryony or its expression.
Changes in the uncertainty and emergence time of the last seed as a function of the seed’s gemellar class do not have a definitive explanation. Among the hypotheses, we think that the embryos, even the smaller ones, collaborate with each other or stimulate each other in the rupture of the tegument, which can accelerate and synchronize the germination process. Alternatively, the seeds where only one seedling emerged could have a higher expression of genetic variability because they are more likely to be monoembryonic and sexual seeds (see Mendes-Rodrigues et al. [
7]). The emergence time could thus reflect the contribution of the paternal effect as observed in some species [
58], leading to a more asynchronous and possibly slower process in seeds with only one seedling. Mendes-Rodrigues et al. [
59] showed that for the polyembryonic Malvaceae,
Eriotheca pubescens (Mart. and Zucc.) Schott and Endl, the type of pollen (auto versus crossed) may affect polyembryony, with the MENS being higher when pollen comes from another individual. This would reinforce the paternal effect on the expression of polyembryony. Further data and other examples are still necessary to test these hypotheses.
The study of germination measurements in the seeds’ embryonic or gemellar classes is hampered by some morphological aspects. In most species it is not possible to directly evaluate the number of embryos per seed before germination, except for
Ophiopogon japonicus (Thunb.) Ker Gawl. var.
umbrosus Maxim [
60], which has a translucent integument that allows the evaluation of the number of embryos per seed, even before the embryo’s protrusion. The evaluation of the ESC or GSC does not allow, for example, the estimation of germinability in our case. In these cases, the conversion of embryos into seedlings is an indirect measurement of the viability of extra embryos in the species.
The conversion of 67.22% of embryos into seedlings was high in
H. chrysotrichus. The decrease in the number of seedlings in relation to the number of embryos produced has been commonly observed [
7,
59,
61,
62]; however, the conversion rate in most species has not been measured and its determinants are unknown. It is known that temperature affects
H. chrysotrichus [
21] and temperatures between 20 and 30 °C allow the conversion of more embryos into seedlings. In some genotypes of
Citrus, about 50% of the embryos were converted into seedlings, with great variability among genotypes [
63]. In addition, the seed forehead has a strong relationship with the decrease in the conversion of embryos into seedlings, as observed in several species in which the removal of the seed tegument increased the conversion (e.g., [
61,
62]).
Monoembryonic and polyembryonic individuals within the same genus and or species could be an alternative to test these hypotheses. They could show some marked differences in the germination process and its measurements. In
Inga laurina (Sw.) Willd., a polyembryonic legume species, the process of seedling emergence is much slower than in the monoembryonic species
I. sessilis (Vell.) Mart. [
46]. In
E. pubescens, the seeds of polyembryonic individuals showed a higher coefficient of variability in the germination time (75.38%) than the seeds of monoembryonic individuals (52.66%) [
6]. These effects of monoembryony and polyembryony should be evaluated within each group of species carefully, since they may depend on other factors such as ploidy, phylogenetic and environmental conditions [
64,
65].
Monoembryonic populations of
H. chrysotrichus are not known, but if they do exist, they could help to understand the effects of polyembryony more directly. An individual of this species that does not produce fruit by manual self-pollination, indicating that it is self-incompatible, has been observed [
66], signaling the possibility that sexual plants with monoembryonic seeds occur. However, the pattern of embryony in this individual has not been described, which prevents its definition as monoembryonic based only on the reproductive system. Population studies are still needed.
Intraspecific variability was also detected in
H. chrysotrichus’s PSP (14 to 41%) [
20], but with values much lower than those observed here. The selection of individuals with no variability in the mean number of embryos and seedlings per seed would be a beneficial factor in experimental competition studies if the individual factor was not of interest to the study. The expression of polyembryony can be very variable among species, individuals, populations, and even in the same individual evaluated at different times [
7,
67]. On the other hand, the factors that determine the expression of polyembryony are still poorly studied, but there is a record of the effects of pollen type, of the presence of conspecific individuals, the nutritional status, the age of the individual, the orientation of the branch, and of fruit production [
54,
59,
68].
Studies have shown that with the increase in the number of embryos and seedlings per seed, there is a decrease in the individual morphometric measurements of embryos or seedlings [
4,
6,
7,
60]. Seedling size is a direct effect of embryo size in polyembryonic species, regardless of whether the embryos are sexual or asexual [
6,
7,
59,
69]. The reduction in embryo size and consequently seedling size should be compensated, for example, by the effects of growing together, as observed here in doublet seedlings, or it could be compensated in later stages of development. Apparently, the deleterious effects of polyembryony are more marked and stronger in the early stages of the seed–seedling transition and seedling development and may disappear over time [
4] so that an intermediate number of embryos may be more beneficial for the species [
60]. This would explain the Allee effect when there was a density of two, as it would decrease the reduction in embryo size. As an example of compensation, polyembryony increases the chances of at least one seedling surviving per seed [
7].
The nondestructive model of mass prediction of
H. chrysotrichus is suitable for estimating the seedling mass at 54 days, a phase in which manipulation can be performed without causing visible damage to the seedling. These results may allow, for example, the proposition of experimental models of nondestructive competition for the species. Models of prediction of dry mass, leaf area and other nondestructive morphometric variables have been proposed for the most diverse groups, using traditional methods of regression and image acquisition, among others [
70,
71]. These methods are fundamental for the design of experimental studies manipulating seedling growth without destruction, and with registration of the initial point of the experiment. Knowing how seedlings of different sizes respond to gemellar competition is crucial in the study of polyembryonic species; being able to predict their initial mass using nondestructive methods is also essential. The model also clearly demonstrated the impact of increasing the number of seedlings on the reduction of their individual measurements, as discussed previously.

 ,
, 


{kind=link}
{kind=link}