
Cell Factory for Phenylnaphthacenoid Polyketide Production



1
Department of Biology and Environmental Science, College of Science, University of the Philippines Cebu, Lahug, Cebu City 6000, Philippines
2
Marine Biodiscovery Center, Department of Chemistry, University of Aberdeen, Meston Walk, Aberdeen AB24 3UE, Scotland, UK
*
Authors to whom correspondence should be addressed.
SynBio 2023, 1(1), 89-102; https://doi.org/10.3390/synbio1010007
Submission received: 5 January 2023
/
Revised: 19 January 2023
/
Accepted: 19 January 2023
/
Published: 24 January 2023
Abstract
:Covering 2009–2022. Phenylnaphthacenoid polyketides have gained significant interest in recent years owing to their potent antibacterial and anticancer activities. Notably, more than 100 members of this class of natural products have been discovered from various Streptomyces species by different research groups including ours over the last 13 years. This review summarizes the current knowledge of the discovery, chemical diversity, and bioactivity of phenylnaphthacenoid polyketides. The current review also highlights the cell factory for phenylnaphthacenoid production: (1) native strains, (2) mutant strains, (3) heterologous expression, and (4) biocatalytic halogenations. Furthermore, current challenges and future opportunities are also presented as a guide for researchers to explore them more purposefully.
1. Introduction
Phenylnaphthacenoids belong to a class of rare aromatic polyketides containing a core gem-dimethyl-naphthacenone scaffold (A–B–C–D ring) connected to a phenyl moiety (D ring) [1,2]. These natural polyketides, which are usually substituted with monochloride or multi-chlorides, show potent activity against Gram-positive bacteria including clinically relevant pathogens MRSA and VRE [3,4]. Some congeners also exhibit excellent activities against Gram-negative bacteria and are cytotoxic against several cancer cell lines [5,6]. Furthermore, they also inhibit Poly(ADP-ribose) polymerase-1 (PARP1) [7] and protein tyrosine phosphatase-1B (PTP-1B) [8].
To date, more than 100 members of phenylnaphthacenoid polyketides (PNPs) have been identified (Figure 1). The first disclosed members of this class of natural products were reported in the patent literature from Streptomyces sp. KB-3346-5 in 2009 [9]. Thereafter, several other strains of Streptomyces were shown to produce PNPs [2,6,10,11]. With the recent advances in engineering microbial cell factories, many more PNPs with diverse structures were discovered that would have been otherwise inaccessible. Notably, more than three-fourths of the identified PNPs to date were discovered over the last 5 years, including naphthacemycins, fasamycins, formicamycins, accramycins, streptovertimycins, and formicapyridines, among others (Figure 1, Table 1) [1,2,3,11,12,13].
Tremendous progress in the field of PNPs has been made by many different research groups including ours [1,2,3,11,12]. Hence, this review summarizes the current knowledge of the discovery, structural diversity, bioactivity, and methods of promoting the expression of PNPs covering the period spanning 2009 to 2022. Furthermore, the current challenges and future opportunities in the field of PNPs are also presented and discussed, thereby providing a guide for researchers to explore them more purposefully.
2. Chemical Diversity of PNPs
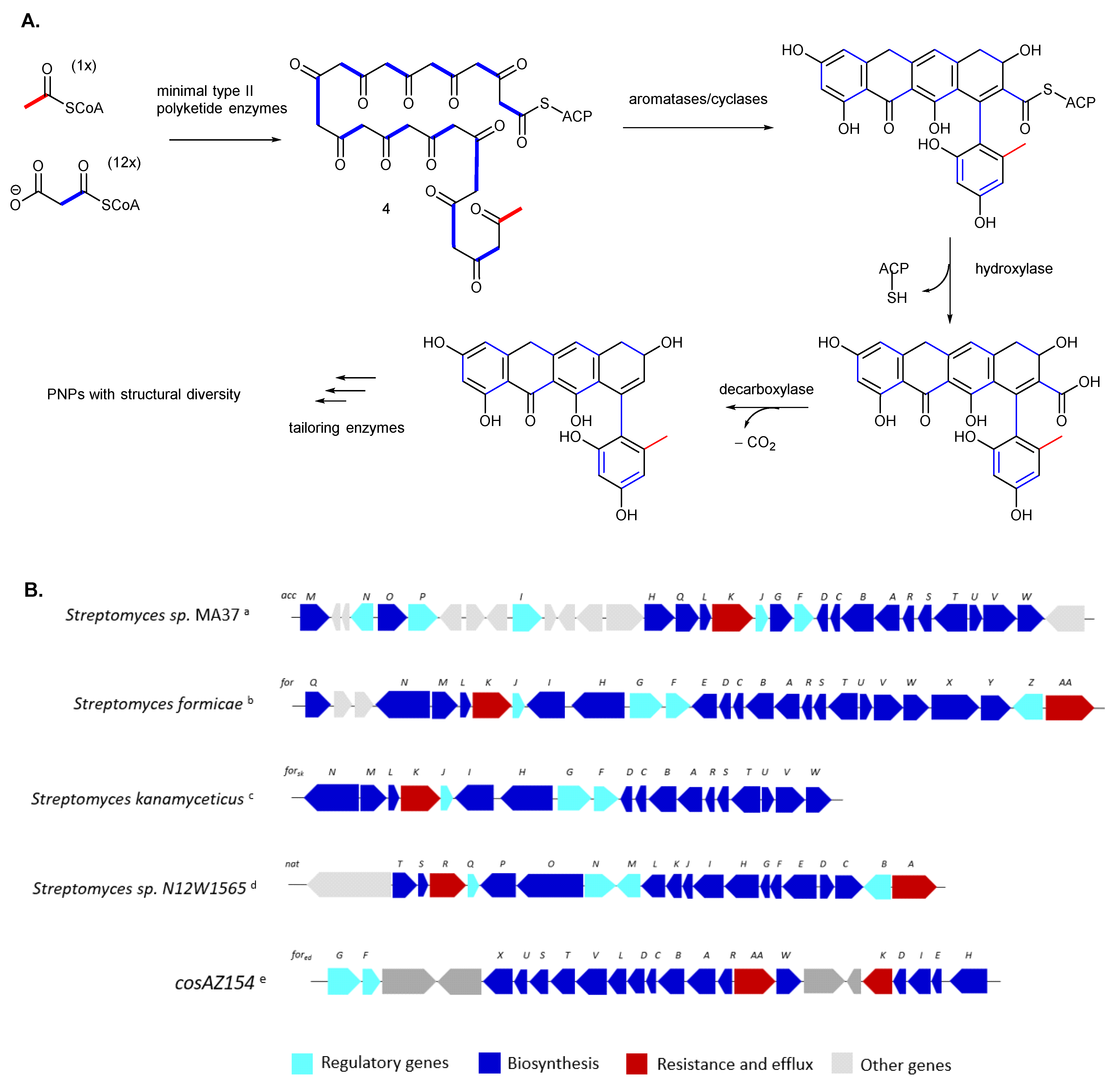
The biosynthesis of PNP natural products originates from a conserved type II polyketide synthase (PKS) system to generate the phenylnaphthacenoid scaffold (Figure 2A). In brief, a linear polyketide chain is assembled by Claisen condensation of short-chain acyl-CoA, which is catalyzed by the minimal PKS, consisting of the ketosynthase KSα, a chain-length factor (CLF, or KSβ), and the acyl carrier protein (ACP) [14]. To date, several biosynthetic pathways encoding the PNPs have been reported, including the accramycin biosynthetic gene cluster (BGC) from Streptomyces sp. MA37 [1,2,15], fasamycin BGC from environmental DNA (eDNA)(cosAZ154) [3], fasamycin BGC from Streptomyces kanamyceticus [16], formicamycin BGC from S. formicae [12,17], and naphthacemycin BGC from Streptomyces sp. N12W1565 [10] (Figure 2B). Consistent with the very similar structures of fasamycins, accramycins, and naphthacemycin type-B compounds, their biosynthetic genes showed very high sequence homology [1,2,10,12,15,17]. Post-PKS tailoring enzymes are also involved to introduce various structural modifications to the polyketide core, including O-methylation and halogenation, plus oxidative and reductive modifications to generate a plethora of PNPs with great structural diversity [1,2,3,12,15,16,17]. On the other hand, the NatY enzyme responsible for the installation of the sugar moiety to the phenolic hydroxyl group of the naphthacemycin core was identified in vitro and is located outside the nat gene cluster [10].
Structure variants with differences in the oxidation state of ring C are commonly encountered in the isolation of PNPs (Figure 1). Accramycins, fasamycins, streptovertimycins, and naphthacemycin B compounds feature a dehyroxyquinol [1,2,4], naphthacemycin A compounds have a quinone motif [6,18], naphthacemycin C molecules consist of a semiquinone-like structure, formicamycins bear a partially oxidized core, and the Baeyer–Villiger intermediates depict a lactone moiety [17]. Streptovertimycin I features a quinone motif in ring E [11]. Naphthacemycins D1 and D2 have a glucose moiety linked to ring D via an O-glycosidic bond [10]. Other structural variations observed in ring D include a cleaved ring, such as in streptovertidione, and a rare pyridine moiety in formicapyridines and streptovertidines [6,13]. It is noteworthy that the axial chirality at C10–C21 of all reported streptovertimycin, accramycin, and fasamycin congeners to date was in the P (or Sa) configuration, though they were obtained from different microbial sources [11,12,16]. This stereo configuration is likely a common characteristic of PNP natural products.
3. Cell Factory for Phenylnaphthacenoid Production
Typically, bioactive compounds are generated by the cell factory of the producing microorganism [19]. Several PNPs discovered to date were obtained from the wild-type strains of various Streptomyces species (Table 1) [2,6,11,12]. Moreover, the recent advances in microbial cell factory engineering techniques gave rise to a larger number of PNPs with diverse structures and improved product titers [1,17,19]. The next section summarizes the methods for promoting the expression of PNPs (i.e., native strain, mutant strain, heterologous expression, and biocatalytic halogenations) along with their biological activities.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Naturally occurring phenylnaphthacenoid polyketide natural products obtained from various Streptomyces species and their bioactivity.
Table 1.
Naturally occurring phenylnaphthacenoid polyketide natural products obtained from various Streptomyces species and their bioactivity.
| Compound | Producing Organism | Production Method/s | Bioactivity | Ref | |
|---|---|---|---|---|---|
| 1 | Accramycin A | S. sp. MA37 S. morookaense | WT, mutant | G+ MRSA, VRE, MSSA, VSE, A549, HeLa, HepG2, MCF-7, Vero | [1,2,6] |
| 2 | Accramycin B | S. sp. MA37 S. kanamyceticus | mutant heterologous expression | G+ | [1,2,16] |
| 3 | Accramycin D, E, G, I | S. sp. MA37 S. morookaense | mutant WT | G+ MRSA, VRE, MSSA, VSE, A549, HeLa, HepG2, MCF-7, Vero | [1,2,6] |
| 4 | Accramycin C, F, H, J, K | S. sp. MA37 | mutant | G+ | [1,2] |
| 5 | Baeyer–Villiger lactone intermediates | S. formicae | mutant | NT | [17] |
| 6 | Fasamycin A, B | S. albus | heterologous expression | MRSA, VRE | [3,4] |
| 7 | Fasamycin C | S. formicae S. sp. KIB-1414 S. sp. MA37 S. kanamyceticus | WT mutant heterologous expression | G+ MRSA, VRE | [12] [5] [1] [16] |
| 8 | Fasamycin D | S. formicae | WT | G+ MRSA, VRE | [12] |
| 9 | Fasamycin E | S. formicae S. sp. KIB-1414 | WT | G+ MRSA, VRE | [12] [5] |
| 10 | Fasamycin F | S. formicae | mutant | NT | [13] |
| 11 | Fasamycin G, I, K | S. sp. KIB-1414 | WT | MRSA | [5] |
| 12 | Fasamycin H | S. sp. KIB-1414 S. kanamyceticus | WT Heterologous expression | G+ MRSA | [5] [16] |
| 13 | Fasamycin J | S. sp. KIB-1414 S. kanamyceticus | WT Enzymatic halogenation | MRSA G+ | [5] [16] |
| 14 | Fasamycin L–Q | S. formicae | mutant | MRSA, MSSA | [20] |
| 15 | Fasamycin R–T | S. kanamyceticus | Heterologous expression | G+ | [16] |
| 16 | Fasamycin U–Y | S. kanamyceticus | Enzymatic halogenation | G+ | [16] |
| 17 | Formicamycin A–C, E–G | S. formicae S. sp. KIB-1414 | WT | MRSA, VRE | [12] [5] |
| 18 | Formicamycin D, H–M | S. formicae | WT | MRSA, VRE | [12] |
| 19 | Formicamycin N–Q | S. sp. KIB-1414 | WT | MRSA | [5] |
| 20 | Formicamycin R, S | S. formicae | mutant | MRSA, MSSA | [20] |
| 21 | Formicapyridine A–I | S. formicae | WT mutant | NA | [13] |
| 22 | Naphthacemycin A1–A11 | S. sp. KB-3346-5 | WT | MRSA, MSSA | [18,21] |
| 23 | Naphthacemycin B1 | S. sp. KB-3346-5S. sp. MA37 | WT mutant | MRSA | [18,21,22] |
| 24 | Naphthacemycin B2 | S. sp. KB-3346-5S. sp. N12W1565 | WT | MRSA, MSSA | [8,18,21] |
| 25 | Naphthacemycin B3, B4 | S. sp. KB-3346-5 | WT | MRSA, MSSA | [21] |
| 26 | Naphthacemycin B5–B13 | S. sp. N12W1565 | WT | PTP1B | [8] |
| 27 | Naphthacemycin C1, C2 | S. sp. KB-3346-5 | WT | MRSA | [21] |
| 28 | Naphthacemycin D1, D2 | S. sp. N12W1565 | WT | MRSA | [10] |
| 29 | Streptovertidine A B | S. morookaense | WT | A549, HeLa, HepG2, MCF-7, Vero | [6] |
| 30 | Streptovertidione | S. morookaense | WT | A549, MCF-7 | [6] |
| 31 | Streptovertimycin A | S. morookaense S. kanamyceticus | WT heterologous expression | MRSA, VRE, MSSA, VSE, A549, HeLa, HepG2, MCF-7, Vero | [6,11,16] |
| 32 | Streptovertimycin B–T | S. morookaense | WT | MRSA, VRE, MSSA, VSE, A549, HeLa, HepG2, MCF-7, Vero | [6,11] |
WT: wild type; G+: Gram positive; NA: no activity against Bacillus subtilis 168; PTP1B: protein-tyrosine phosphatase 1B; MRSA: methicillin-resistant Staphylococcus aureus; MSSA: methicillin-susceptible Staphylococcus aureus; VRE: vancomycin-resistant Enterococci; VSE: vancomycin-susceptible Enterococci.
3.1. Native Strains for Phenylnaphthacenoid Polyketide Production
A myriad array of PNPs have been isolated from the wild-type (WT) strains of various Streptomyces bacteria and were given alternative names, such as naphthacemycins, fasamycins, formicamycins, streptovertimycins, and accramycins (Figure 1, Table 1). The earliest identified PNPs were the KB-3346-5 substances reported in the patent literature by Omura et al. in 2009 [9] and then renamed as naphthacemycins in 2017 [18,21]. A total of seventeen naphthacemycins (A1–A11, B1–B4, and C1–C2) were isolated from the culture broth of Streptomyces sp. KB-3346-5, all of which showed activity against clinically isolated MRSA (MIC = 8–64 μg·mL−1). Notably, naphthacemycins A8 and A9 displayed comparable anti-MRSA activities to vancomycin antibiotics, and they also inhibited the growth of linezolid-resistant MRSA. Combining naphthacemycins with an imipenem antibiotic enhanced antibacterial activity by 100–500× against β-lactam-resistant MRSA [18,21]. The addition of naphthacemycins reduced the MIC of imipenem from 32 μg·mL−1 to 0.06–0.25 μg·mL−1, breathing new life into old antibiotics that were weakened by the growing antibiotic-resistant strains. Such drug combinations demonstrate an immense potential to treat multidrug-resistant bacterial infections [23]. In 2020, nine new congeners, naphthacemycins B5-B13, were isolated from the culture of Streptomyces sp. N12W1565. All of the compounds inhibited protein-tyrosine phosphatase 1B (PTP1B) with IC50 values of less than 10 μM, indicating that naphthacemycin type-B compounds are promising anti-PTP1B inhibitors [8]. It is noteworthy that the pentachlorinated naphthacemycin B13 showed 3.2× more potent activity (IC50 = 0.34 ± 0.0.06 μM) than the positive control, Na3VO4 (IC50 = 1.10 ± 0.14 μM).
Fasamycins C–E and formicamycins A–M, some of which contain up to four halogen substituents, were identified from Streptomyces formicae, isolated from the tree-associated Tetraponera penzigi ants [12]. The brominated congeners, formicamycins K–M, were produced by supplementing the culture medium (MS agar) with NaBr elicitor. The compounds effectively inhibited the growth of MRSA, VRE, and B. subtilis, with no observed propensity towards resistance in vitro. The potency of the compounds increased with an increasing number of chlorine atoms in the PNP scaffold. Formicamycin congeners with a nonaromatic C ring also appear to be more active than the structurally related fasamycins. Notably, the bromination of formicamycins substantially improved their intrinsic biological activity. The biological effect of halogen atoms on the increased potency of PNPs is probably due to increased lipophilicity resulting in an enhanced ability to cross the bacterial cell membrane [24].
Fasamycins G–K and formicamycins N–Q were discovered from the soil-derived Streptomyces sp. KIB-1414. All compounds were active against MRSA, S. aureus, B. subtilis, and E. coli strains (MIC = 0.20–50.00 μg·mL−1) [5]. Streptovertimycins A–H were identified from the wheat-grown cultures of Streptomyces morookaense (formerly Streptoverticillium morookaense) SC1169 strain [11]. When S. morookaense was cultivated on rice, a diverse array of PNP congeners was produced, including streptovertidines A and B, streptovertidione, and streptovertimycins I–T, some of which showed excellent activity against human lung carcinoma (A549), human cervical carcinoma (HeLa), human hepatocellular carcinoma (HepG2), and human breast carcinoma (MCF-7) cell lines [6].
The naphthacemycin repertoire has recently been expanded with two glycosylated molecules, naphthacemycins D1 and D2 isolated from Streptomyces sp. N12W1565 [10]. Both compounds showed moderate activity against MRSA and B. subtilis (MIC = 16.8–24.4 μg·mL−1) but enhanced aqueous solubility up to 20× higher than non-glycosylated PNPs. Glycosylation has been a useful approach to enhance the solubility and bioavailability of poorly water-soluble drugs [25].
Formicapyridines are pyridine-containing trace-level metabolites identified from S. formicae via a manual curation and bespoke dereplication approach [13]. Their poor productivity in the WT strain indicates that they are shunt products caused by an aberrant derailment in the cyclization step of the fasamycin/formicamycin machinery. Compared to other PNP congeners, formicapyridines are inactive against B. subtilis, suggesting that the pyridine moiety diminished its activity.
Accramycin A was isolated from the Ghanaian soil bacterium Streptomyces sp. MA37 [2]. Two other accramycin analogs were detected in the extract by MS, UV, and molecular network analyses but could not be isolated due to very low levels of production. Moreover, the single PNP accramycin A isolated from the native MA37 strain was only produced at 0.2 mg·L−1 titer.
3.2. Mutant Strains for Phenylnaphthacenoid Polyketides Production
Streptomyces is the largest antibiotic-producing genus to date [26]. However, native strains usually produce these antibiotics in minute quantities, and thus further chemical investigations remain challenging and are a bottleneck to the molecules being advanced for clinical trials. Their production is controlled by several factors, such as nutrition (carbon and nitrogen sources), biosynthetic efficiency, and the complex regulatory mechanisms in producing strains [27].
Microbial cell factories have been engineered to identify new metabolic pathways and design efficient strains to produce pharmaceutically valuable PNPs. This has been achieved in three ways: (1) by manipulating regulatory genes, (2) by engineering PKS biosynthetic enzymes, and (3) by combining mutations in regulatory and biosynthetic genes.
Four regulatory genes are present in the accramycin cluster, including LuxR (AccF), MarR (AccJ), LysR (AccI), and MerR (AccP) transcriptional regulators [1]. These genes were hypothesized to control the productivity of accramycins in the native microbial cultures. Hence, the roles of the four regulatory genes were investigated by inactivating these genes in MA37 via a double crossover homologous recombination approach. One variant (ΔaccJ) resulted in the overproduction of accramycin A up to 330-fold higher than WT levels, indicating that the accJ gene is a negative regulator in accramycin biosynthesis. Furthermore, the ΔaccJ engineered strain produced new accramycin congeners (B–K) and other small polyketide molecules that were identified for the first time from bacteria [15]. MarR (multiple antibiotic resistance)-family regulators, such as accJ, repress the transcription of their target genes by binding specifically to DNA sequences within promoter regions [28]. All accramycin congeners displayed potent activity against Gram-positive bacteria, including Enterococcus faecium (K59-68 and K60-39) and Staphylococcus haemolyticus clinical isolates (MIC = 3.1–12.5 μg·mL−1). ForJ, a homolog of AccJ, in formicamycin biosynthesis was also shown to repress the production of formicamycins [20]. Deletion of forJ switched biosynthesis from solid agar to liquid cultures, increased formicamycin titers, and induced the production of new congeners (formicamycins R and S, each bearing five chlorine atoms) in engineered S. formicae strains. Both molecules exhibited potent antibacterial activity against MSSA and MRSA (MIC = 2 μg·mL−1).
PKS tailoring enzymes have been engineered to understand metabolic pathways and expand the chemical space of PNPs. Formicamycin BGC has an additional set of four grouped unique genes (forX, forY, for Z, and forAA) compared to other PNP-producing gene clusters (Figure 2B) [12]. Deletion of the flavin-dependent monooxygenase ForX abrogated formicamycin production and lead to the accumulation of fasamycin E [17]. Deletion of the flavin-dependent reductase ForY also abolished formicamycin biosynthesis and led to the accumulation of new Baeyer–Villiger (BV)-derived lactone intermediates (Figure 1). Therefore, fasamycin molecules are biosynthetic precursors of the formicamycins, and ForX and ForY enzymes are required for this two-step conversion which proceeds via ring expansion to form the BV intermediates followed by a unique reductive ring contraction Favorskii-type reaction.
The production of the shunt metabolites, formicapyridines, increased approximately 25-fold relative to the WT by deletion of the antibiotic biosynthesis monooxygenase (ABM) ForS enzyme [13]. Meanwhile, the production titer of formicamycins in the ΔforS mutant was reduced to about one-third that of the WT strain. The ΔforS mutant also produced a minor congener, Fasamycin F, which harbors a carboxylic acid group at C-11, indicating that the deletion of ForS in S. formicae did not abolish polyketide production (Figure 1). This congener was not observed in any WT cultures. Hence, ForS is not involved in ring-D (pyridine) cyclization as initially perceived nor in the biosynthesis of the for BGC polyketide backbone but rather in the maintenance of pathway productivity and fidelity. ForS acts as a chaperone or fidelity factor that ensures that the biosynthetic mechanism proceeds efficiently.
Combining mutations in genes encoding the biosynthetic machinery and regulation resulted in the biosynthesis of additional PNP precursors. Deletion of the halogenase forV gene in ΔforJ mutant in S. formicae (ΔforJΔforV) leads to the accumulation of nonhalogenated fasamycin congeners. Particularly, fasamycin C production is elevated 90-fold higher than the total fasamycins produced by the WT strain [20]. On the other hand, deletion of the forX gene involved in the ring expansion during the transformation of fasamycins to formicamycins in ΔforJ mutant (ΔforJΔforX) abolished formicamycin production but accumulated chlorinated fasamycins including six new congeners (fasamycins L-Q) (Figure 1). All these congeners showed potent antibacterial activity against MSSA and MRSA (MIC = 2–4 μg·mL−1). Furthermore, the product titers on solid cultures were increased to approximately 120 times higher than the WT titers.
Two-component systems (TCSs), consisting of sensor kinase and response regulator proteins, are one of the major ways in which bacteria respond and adapt to ever-changing environmental conditions [29]. The large genomes of Streptomyces harbor a high number of TCSs compared to other bacterial genera, and the majority have been shown to control secondary metabolism and morphological development [29,30]. The deletion of the TCS ForGF in S. formicae diminished fasamycin and formicamycin production, indicating that ForGF is required for the transcription of the for BGC [20]. On the other hand, overexpression of ForGF in a ΔforJ mutant improved formicamycin yield on solid cultures 10-fold compared to the WT strain. Therefore, cluster-situated TCSs, such as the ForGF, represent a promising target for overexpression to activate silent or cryptic BGCs and obtain higher efficiencies in secondary metabolite production.
3.3. Heterologous Expression for Phenylnaphthacenoid Polyketide Production
Nature holds an enormous amount of microbial diversity with remarkable biosynthetic potential; however, the majority remains recalcitrant to conventional culturing methods. With the recent advances in culture-independent approaches and metagenomics, a large number of unculturable or poorly culturable microbes hitherto became accessible for functional analysis [31]. Functional metagenomics has been used to isolate and characterize small molecules and proteins from soil environmental DNA (e-DNA) metagenomes. Among the rare skeletons isolated using this strategy were fasamycins A and B obtained by the heterologous expression of clones from soil-derived PKS e-DNA cosmid (cosAZ154) in Streptomyces albus (Figure 1, Table 1) [3].
The structure of fasamycin B differs from A in the presence of an additional chlorine substituent at carbon-3. Fasamycins A and B showed activity against MRSA and VRE by inhibiting FabF during type II fatty acid elongation [3,4]. The mono-chlorinated fasamycin A (MIC = 0.8 μg·mL−1) is almost eight times more active than the dichlorinated fasamycin B (MIC = 6.25 μg·mL−1) against VRE, indicating that the specific halogenation pattern is critical for potent antimicrobial efficacy. Both compounds, however, were inactive against yeast or Gram-negative bacteria at the highest concentration tested (50 μg·mL−1) [3].
In another study, heterologous expression was used to awaken silent or cryptic biosynthetic pathways to produce medically useful compounds. Three new fasamycin congeners (R–T) along with four known fasamycin-type analogs were obtained via the overexpression of two phosphopantetheinyl transferases (PPtases) in S. kanamyceticus [16]. PPtases are responsible for the installation of a 4′-phosphopantetheine arm from Coenzyme A to various carrier proteins (CPs), an essential step in the biosynthesis of PKS, NRPS, and fatty acids (FAs) [32]. All congeners showed potent activity against B. subtilis and S. aureus (MIC = 1.6–12.5 μg·mL−1) [16].
3.4. Biocatalytic Halogenations of the Phenylnaphthacenoid Scaffold
Biocatalytic halogenations have been important and valuable reactions in organic synthesis owing to their regio- and stereoselectivity, and they are carried out by enzymes called halogenases or haloperoxidases [33,34]. Halogenases enable the selective installation of halogen atoms in chemical scaffolds of varying complexity under mild reaction conditions. The introduction of a halogen atom into a molecule can effectively alter its properties (e.g., bioactivity and pharmacokinetic profile), yielding bioactive compounds of industrial and pharmacological significance [34,35].
The fasamycin BGC in S. kanamyceticus encodes a single halogenase FasVsk, which shows high homology to FasVed from the fasamycin BGC of eDNA (59.8%), AccV of the accramycin BGC (67.5%), and ForV of the formicamycin BGC (92.8%). The FasVsk was expressed in Escherichia coli and its catalytic efficiency was tested in vitro using fasamycin C, fasamycin R, and streptovertimycin R as substrates (Figure 3). Similarly, the halogenase Abx(−)H29 in anthrabenzoxocinone (ABX) biosynthesis, which has been shown to introduce halogen modifications into the ABX scaffolds was also investigated in vitro [36]. Both halogenases FasVsk and Abx(−)H29 specifically installed the halogen substituent into the C-13 and/or C-25 positions of fasamycins, different from the specific halogenation patterns observed in formicamycins or accramycins [16]. The enzymatic halogenation reactions yielded six new fasamycin congeners (fasamycins J and U-Y), further expanding the bioactive repertoire of PNP scaffolds. An antimicrobial assay of the fasamycins showed that the halogenation profoundly enhanced their antibiotic activity, among which the dibrominated fasamycin V is four times more potent (MIC = 1.6 μg·mL−1) than the non-halogenated fasamycin C (MIC = 6.3 μg·mL−1) against B. subtilis and S. aureus.
In another study, a chemoenzymatic approach was used to introduce the chlorine substituent in naphthacemycin B1 to form halogenated PNPs [37]. The retrosynthesis of naphthacemycin B1 was carried out in a convergent manner by combining two fragments in a Sammes annulation strategy followed by the installation of the pendant phenyl ring by Suzuki coupling. Naphthacemycin B1 was then subjected to an enzymatic halogenation reaction using FasV and ForV halogenases in combination with five FAD reductases, namely Fre, SfFre (a Fre homolog from S. formicae), SsuE, CtcQ, and HpaC. Mass spectrometric analysis of the reaction mixture revealed monochlorinated and dichlorinated PNPs with fasamycin A as the major product as confirmed by comparison with an authentic standard. Among the combinations tested, the FasV/CtcQ pair showed the highest conversion to fasamycin A though the overall chlorination yield was very low.
4. Challenges and Future Opportunities
Phenylnaphthacenoid polyketides possess potent antibacterial and anticancer activities with a high barrier to the development of resistance, which demonstrates immense potential as lead candidates for investigation as a new structural class of antibiotics. However, the inadequate supply of naturally derived antimicrobials has been a major limiting factor in their preclinical and clinical development. In another case, the expression of PNPs in liquid cultures has been a significant hurdle in S. formicae [20]. Indeed, this has been a bottleneck since the majority of industrial antibiotic production is produced in bioreactors in liquid cultures rather than in solid media [38]. Engineering microbial cell factories offer tremendous potential in inducing biosynthesis in liquid cultures, improving product titers, and expanding the chemical space. The deletion of MarR regulatory genes, accJ and forJ, in accramycin and formicamycin biosynthesis, respectively, enabled the production of PNPs in higher yields and the expression of new analogs [1,2,20]. In addition, engineered S. formicae strains produced PNPs not only in solid but also in liquid cultures.
Refactoring of complex biosynthetic pathways in heterologous hosts has also enabled the generation of cell factories that are optimized for producing new PNP congeners [3,16]. Biocatalysts, such as halogenases, have produced new chlorinated and brominated PNP derivatives although, in some cases, the yield is very low. Future study is therefore geared towards engineering enzymes and understanding their optimum reaction conditions. It is noteworthy that the bromination (and in most cases chlorination) of PNP molecules rendered them more bioactive [6,12]. Hence, the characterization of halogenases that can selectively introduce a halogen atom into desired positions in the PNP scaffold is of significant interest to access antimicrobials with increased potencies.
Two is better than one as demonstrated in the combined activity of naphthacemycin and imipenem against β-lactam-resistant MRSA [18,21]. Such drug combinations offer better efficacy than single agents and are therefore an avenue for further exploration to combat multidrug-resistant superbugs. Further research is also needed to understand the molecular mechanisms underlying them. In the face of the growing threat of AMR with a dry antibiotic pipeline, it is more critical than ever to find other treatment options [23,39].
Looking forward, further characterization of PKS-encoding genes involved in PNP pathways will uncover new antibiotics, pave the way for a greater understanding of their complex biosynthetic mechanisms, and establish new strategies for metabolic engineering. The characterization of TCS genes could unveil cryptic antibiotics that are not expressed under standard laboratory conditions. Understanding the novel mechanism involved in reductive ring contraction catalyzed by ForY in formicamycin biosynthesis could produce further improved molecules with potent bioactivities.
The biosynthesis of nanoparticles in Streptomyces is an emerging trend in bionanotechnology [40,41]. Nanoparticles have wide applications in therapeutics and medicine, drug and gene delivery, bioimaging and biosensors, cancer nanotechnology, and the chemical industry [42,43]. Because Streptomyces is the largest antibiotic-producing genus to date [44], it comprises efficient candidates for the biological synthesis of metal nanoparticles [40]. Exploiting the biosynthetic potential of PNP-producing Streptomyces to produce diverse nanomaterials is therefore an exciting field to explore! We expect that with the recent advances in science and technology, there will be increasing momentum in the discovery and development of nanoparticle systems in the coming years.
5. Conclusions
Taken together, the discovery, structural diversity, bioactivity, and cell factory for metabolite production highlighted in this review demonstrate the huge potential of the phenylnaphthacenoid class of natural products in modern medicine. PNPs could prove invaluable in the fight against resistant superbugs. More than 100 members of PNPs with diverse structures and biological activities have been isolated from various Streptomyces species to date. These compounds have been obtained from native strains, mutant strains, heterologous expression, and biocatalytic modifications. There are exciting opportunities ahead!
Author Contributions
Conceptualization, F.M. and H.D. formal analysis, F.M.; investigation, F.M.; data curation, F.M.; writing—original draft preparation, F.M.; writing—review and editing, F.M., H.D. All authors have read and agreed to the published version of the manuscript.
Funding
F.M. is grateful to the Department of Science and Technology (DOST)—National Research Council of the Philippines (NRCP) and the DOST through the Science for Change Program (S4CP) Collaborative Research and Development to Leverage Philippine Economy (CRADLE) Project no. 8647. H.D. is thankful for the financial support of the Leverhulme Trust-Royal Society Africa award (AA090088) and the jointly funded UK Medical Research Council–UK Department for International Development (MRC/DFID) Concordat Agreement African Research Leaders Award (MR/S00520X/1).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Maglangit, F.; Zhang, Y.; Kyeremeh, K.; Deng, H. Discovery of new antibacterial accramycins from a genetic variant of the soil bacterium, Streptomyces sp. MA37. Biomolecules 2020, 10, 1464. [Google Scholar] [CrossRef] [PubMed]
- Maglangit, F.; Fang, Q.; Leman, V.; Soldatou, S.; Ebel, R.; Kyeremeh, K.; Deng, H. Accramycin A, a new aromatic polyketide, from the soil bacterium, Streptomyces sp. MA37. Molecules 2019, 24, 3384. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Kallifidas, D.; Brady, S.F. Functional analysis of environmental DNA-derived type II polyketide synthases reveals structurally diverse secondary metabolites. Proc. Natl. Acad. Sci. USA 2011, 108, 12629–12634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Z.; Chakraborty, D.; Dewell, S.B.; Reddy, B.V.B.; Brady, S.F. Environmental DNA-encoded antibiotics fasamycins A and B inhibit FabF in type II fatty acid biosynthesis. J. Am. Chem. Soc. 2012, 134, 2981–2987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Wang, L.; Ren, J.; Huang, J.P.; Yu, M.; Tang, J.; Yan, Y.; Yang, J.; Huang, S.X. Antibacterial Pentacyclic Polyketides from a Soil-Derived Streptomyces. J. Nat. Prod. 2020, 83, 1919–1924. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, P.; Li, H.; Xue, J.; Xu, H.; Wei, X. Antibacterial and Cytotoxic Phenyltetracenoid Polyketides from Streptomyces morookaense. J. Nat. Prod. 2021, 84, 1806–1815. [Google Scholar] [CrossRef]
- Shen, W.; Lu, X.; Zhu, J.; Mu, Y.; Xu, Y.; Gao, J.; Zhang, X.; Zheng, Z. Discovery of naphthacemycins as a novel class of PARP1 inhibitors. Bioorganic Med. Chem. Lett. 2019, 29, 1904–1908. [Google Scholar] [CrossRef]
- Huo, C.; Zheng, Z.; Xu, Y.; Ding, Y.; Zheng, H.; Mu, Y.; Niu, Y.; Gao, J.; Lu, X. Naphthacemycins from a Streptomyces sp. as Protein-Tyrosine Phosphatase Inhibitors. J. Nat. Prod. 2020, 83, 1394–1399. [Google Scholar] [CrossRef] [PubMed]
- Omura, S.; Takahashi, Y.; Kim, Y.-P.; Hanaki, H.; Koda, H.; Suzuki, M.; Shiomi, K. New KB-3346-5 Substance and Method for Producing the Same. JP2009046404A, 2009. [Google Scholar]
- Gao, Y.H.; Nie, Q.Y.; Hu, Y.; Lu, X.; Xiang, W.; Wang, X.; Tang, G.L. Discovery of glycosylated naphthacemycins and elucidation of the glycosylation. Biochem. Biophys. Res. Commun. 2022, 622, 122–128. [Google Scholar] [CrossRef]
- Yang, L.; Li, X.; Wu, P.; Xue, J.; Xu, L.; Li, H.; Wei, X. Streptovertimycins A–H, new fasamycin-type antibiotics produced by a soil-derived Streptomyces morookaense strain. J. Antibiot. (Tokyo) 2020, 73, 283–289. [Google Scholar] [CrossRef]
- Qin, Z.; Munnoch, J.T.; Devine, R.; Holmes, N.A.; Seipke, R.F.; Wilkinson, K.A.; Wilkinson, B.; Hutchings, M.I. Formicamycins, antibacterial polyketides produced by Streptomyces formicae isolated from African Tetraponera plant-ants. Chem. Sci. 2017, 8, 3218–3227. [Google Scholar] [CrossRef] [Green Version]
- Qin, Z.; Devine, R.; Hutchings, M.I.; Wilkinson, B. A role for antibiotic biosynthesis monooxygenase domain proteins in fidelity control during aromatic polyketide biosynthesis. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Khosla, A.D. and C. Biosynthesis of Aromatic Polyketides in Bacteria. Acc. Chem. Res. 2009, 42, 631–639. [Google Scholar] [CrossRef] [Green Version]
- Maglangit, F.; Kyeremeh, K.; Deng, H. Deletion of the accramycin pathway-specific regulatory gene accJ activates the production of unrelated polyketide metabolites. Nat. Prod. Res. 2022, 1–6. [Google Scholar] [CrossRef]
- Jiang, K.; Yan, X.; Deng, Z.; Lei, C.; Qu, X. Expanding the Chemical Diversity of Fasamycin Via Genome Mining and Biocatalysis. J. Nat. Prod. 2022, 85, 943–950. [Google Scholar] [CrossRef]
- Qin, Z.; Devine, R.; Booth, T.J.; Farrar, E.H.E.; Grayson, M.N.; Hutchings, M.I.; Wilkinson, B. Formicamycin biosynthesis involves a unique reductive ring contraction. Chem. Sci. 2020, 11, 8125–8131. [Google Scholar] [CrossRef]
- Fukumoto, A.; Kim, Y.P.; Iwatsuki, M.; Hirose, T.; Sunazuka, T.; Hanaki, H.; Omura, S.; Shiomi, K. Naphthacemycins, novel circumventors of β-lactam resistance in MRSA, produced by Streptomyces sp. KB-3346-5. II. Structure elucidation. J. Antibiot. (Tokyo) 2017, 70, 568–573. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, J. Cell factory engineering for improved production of natural products. Nat. Prod. Rep. 2019, 36, 1233–1236. [Google Scholar] [CrossRef] [Green Version]
- Devine, R.; McDonald, H.P.; Qin, Z.; Arnold, C.J.; Noble, K.; Chandra, G.; Wilkinson, B.; Hutchings, M.I. Re-wiring the regulation of the formicamycin biosynthetic gene cluster to enable the development of promising antibacterial compounds. Cell Chem. Biol. 2021, 28, 515–523. [Google Scholar] [CrossRef]
- Fukumoto, A.; Kim, Y.P.; Matsumoto, A.; Takahashi, Y.; Suzuki, M.; Onodera, H.; Tomoda, H.; Matsui, H.; Hanaki, H.; Iwatsuki, M.; et al. Naphthacemycins, novel circumventors of β-lactam resistance in MRSA, produced by Streptomyces sp. KB-3346-5. I. The taxonomy of the producing strain, and the fermentation, isolation and antibacterial activities. J. Antibiot. (Tokyo) 2017, 70, 562–567. [Google Scholar] [CrossRef]
- Maglangit, F.; Fang, Q.; Kyeremeh, K.; Sternberg, J.M.; Ebel, R.; Deng, H. A Co-Culturing Approach Enables Discovery and Biosynthesis of a Bioactive Indole Alkaloid Metabolite. Molecules 2020, 25, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyers, M.; Wright, G.D. Drug combinations: A strategy to extend the life of antibiotics in the 21st century. Nat. Rev. Microbiol. 2019, 17, 141–155. [Google Scholar] [CrossRef]
- Naumann, K. Influence of Chlorine Substituents on Biological Activity of Chemicals: A Review. Adv. Synth. Catal. 1999, 341, 417–435. [Google Scholar] [CrossRef]
- Moradi, S.V.; Hussein, W.M.; Varamini, P.; Simerska, P.; Toth, I. Glycosylation, an effective synthetic strategy to improve the bioavailability of therapeutic peptides. Chem. Sci. 2016, 7, 2492–2500. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, F.; Hou, Y.; Zhang, H.; Chu, Y.; Xia, H.; Tian, Y. Regulatory genes and their roles for improvement of antibiotic biosynthesis in Streptomyces. 3 Biotech 2017, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Grove, A. MarR family transcription factors. Curr. Biol. 2013, 23, R142–R143. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, H.; Rico, S.; Díaz, M.; Santamaría, R.I. Two-component systems in Streptomyces: Key regulators of antibiotic complex pathways. Microb. Cell Fact. 2013, 12, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hutchings, M.I.; Hoskisson, P.A.; Chandra, G.; Buttner, M.J. Sensing and responding to diverse extracellular signals? Analysis of the sensor kinases and response regulators of Streptomyces coelicolor A3(2). Microbiology 2004, 150, 2795–2806. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M.C.; Piel, J. Metagenomic approaches for exploiting uncultivated bacteria as a resource for novel biosynthetic enzymology. Chem. Biol. 2013, 20, 636–647. [Google Scholar] [CrossRef]
- Beld, J.; Sonnenschein, E.C.; Vickery, C.R.; Noel, J.P.; Burkart, M.D. The Phosphopantetheinyl Transferases: Catalysis of a Posttranslational Modification Crucial for Life. Nat. Prod. Rep. 2014, 31, 61–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, V.; Miles, Z.D.; Winter, J.M.; Eustáquio, A.S.; El Gamal, A.A.; Moore, B.S. Enzymatic Halogenation and Dehalogenation Reactions: Pervasive and Mechanistically Diverse. Chem. Rev. 2017, 117, 5619–5674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Maglangit, F.; Deng, H. Fluorine biocatalysis. Curr. Opin. Chem. Biol. 2020, 55, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Büchler, J.; Papadopoulou, A.; Buller, R. Recent Advances in Flavin-Dependent Halogenase Biocatalysis: Sourcing, Engineering, and Application. Catalysts 2019, 9, 1030. [Google Scholar] [CrossRef] [Green Version]
- Mei, X.; Yan, X.; Zhang, H.; Yu, M.; Shen, G.; Zhou, L.; Deng, Z.; Lei, C.; Qu, X. Expanding the Bioactive Chemical Space of Anthrabenzoxocinones through Engineering the Highly Promiscuous Biosynthetic Modification Steps. ACS Chem. Biol. 2018, 13, 200–206. [Google Scholar] [CrossRef]
- Li, J.; Renata, H. Concise Chemoenzymatic Synthesis of Fasamycin A. J. Org. Chem. 2021, 86, 11206–11211. [Google Scholar] [CrossRef] [PubMed]
- Manteca, Á.; Yagüe, P. Streptomyces differentiation in liquid cultures as a trigger of secondary metabolism. Antibiotics 2018, 7, 41. [Google Scholar] [CrossRef] [Green Version]
- Maglangit, F.; Yu, Y.; Deng, H. Bacterial pathogens: Threat or treat (a review on bioactive natural products from bacterial pathogens. Nat. Prod. Rep. 2021, 38, 782–821. [Google Scholar] [CrossRef]
- Kolahalam, L.A.; Kasi Viswanath, I.V.; Diwakar, B.S.; Govindh, B.; Reddy, V.; Murthy, Y.L.N. Review on nanomaterials: Synthesis and applications. Mater. Today Proc. 2019, 18, 2182–2190. [Google Scholar] [CrossRef]
- Subbaiya, R.; Saravanan, M.; Priya, A.R.; Shankar, K.R.; Selvam, M.; Ovais, M.; Balajee, R.; Barabadi, H. Biomimetic synthesis of silver nanoparticles from Streptomyces atrovirens and their potential anticancer activity against human breast cancer cells. IET Nanobiotechnology 2017, 11, 965–972. [Google Scholar] [CrossRef]
- Stark, W.J.; Stoessel, P.R.; Wohlleben, W.; Hafner, A. Industrial applications of nanoparticles. Chem. Soc. Rev. 2015, 44, 5793–5805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Gu, F.X.; Chan, J.M.; Wang, A.Z.; Langer, R.S.; Farokhzad, O.C. Nanoparticles in medicine: Therapeutic applications and developments. Clin. Pharmacol. Ther. 2008, 83, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Genilloud, O. Actinomycetes: Still a source of novel antibiotics. Nat. Prod. Rep. 2017, 34, 1203–1232. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(A) Structures of accramycins A–K isolated from Streptomyces sp. MA37; (B) naphthacemycins A1–A11 from Streptomyces sp. KB-3346-5; (C) naphthacemycins B1–B4 from Streptomyces sp. KB-3346-5 and naphthacemycins B5-B13 from Streptomyces sp. N12W1565; (D) naphthacemycins C1–C2 from Streptomyces sp. KB-3346-5; (E) naphthacemycins D1–D2 from Streptomyces sp. N12W15; (F) Baeyer–Villiger lactone intermediates A–E from Streptomyces formicae ΔforY mutant; (G) fasamycins A–F, L–Q from Streptomyces formicae, fasamycins G–K from Streptomyces sp. KIB-1414, and fasamycins R–Y from Streptomyces kanamyceticus; (H) formicamycins A–M, R–S from Streptomyces formicae, formicamycins N–Q from Streptomyces sp. KIB-1414, (I) formicapyridines A–I from Streptomyces formicae; (J) streptovertimycins A–T from Streptomyces morookaense; (K) streptovertimycin I from Streptomyces morookaense; (L) streptovertidines A-B from Streptomyces morookaense; and (M) streptovertidione from Streptomyces morookaense.
Figure 1.
(A) Structures of accramycins A–K isolated from Streptomyces sp. MA37; (B) naphthacemycins A1–A11 from Streptomyces sp. KB-3346-5; (C) naphthacemycins B1–B4 from Streptomyces sp. KB-3346-5 and naphthacemycins B5-B13 from Streptomyces sp. N12W1565; (D) naphthacemycins C1–C2 from Streptomyces sp. KB-3346-5; (E) naphthacemycins D1–D2 from Streptomyces sp. N12W15; (F) Baeyer–Villiger lactone intermediates A–E from Streptomyces formicae ΔforY mutant; (G) fasamycins A–F, L–Q from Streptomyces formicae, fasamycins G–K from Streptomyces sp. KIB-1414, and fasamycins R–Y from Streptomyces kanamyceticus; (H) formicamycins A–M, R–S from Streptomyces formicae, formicamycins N–Q from Streptomyces sp. KIB-1414, (I) formicapyridines A–I from Streptomyces formicae; (J) streptovertimycins A–T from Streptomyces morookaense; (K) streptovertimycin I from Streptomyces morookaense; (L) streptovertidines A-B from Streptomyces morookaense; and (M) streptovertidione from Streptomyces morookaense.



Figure 2.
(A) The biosynthetic model of type II polyketide backbone formation leading to the ring systems of PNP, and (B) comparison of the fasamycin, accramycin, formicamycin, and naphthacemycin biosynthetic gene clusters (a from accramycins produced by Streptomyces sp. MA37, b from formicamycins produced by Streptomyces formicae, c from fasamycins produced by Streptomyces kanamyceticus, d naphthacemycin BGC from Streptomyces sp. N12W1565, e from environmental DNA, containing the fasamycin BGC).
Figure 2.
(A) The biosynthetic model of type II polyketide backbone formation leading to the ring systems of PNP, and (B) comparison of the fasamycin, accramycin, formicamycin, and naphthacemycin biosynthetic gene clusters (a from accramycins produced by Streptomyces sp. MA37, b from formicamycins produced by Streptomyces formicae, c from fasamycins produced by Streptomyces kanamyceticus, d naphthacemycin BGC from Streptomyces sp. N12W1565, e from environmental DNA, containing the fasamycin BGC).

Figure 3.
Enzymatic halogenation of fasamycins to produce new congeners.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Maglangit, F.; Deng, H. Cell Factory for Phenylnaphthacenoid Polyketide Production. SynBio 2023, 1, 89-102. https://doi.org/10.3390/synbio1010007
AMA Style
Maglangit F, Deng H. Cell Factory for Phenylnaphthacenoid Polyketide Production. SynBio. 2023; 1(1):89-102. https://doi.org/10.3390/synbio1010007
Chicago/Turabian StyleMaglangit, Fleurdeliz, and Hai Deng. 2023. "Cell Factory for Phenylnaphthacenoid Polyketide Production" SynBio 1, no. 1: 89-102. https://doi.org/10.3390/synbio1010007