
Cardioprotective Effects of Resveratrol in the Mediterranean Diet: A Short Narrative Review



Department of Medical and Surgical Sciences, University of Foggia, Viale Pinto 1, 71122 Foggia, Italy
*
Author to whom correspondence should be addressed.
Dietetics 2023, 2(2), 174-190; https://doi.org/10.3390/dietetics2020014
Submission received: 13 December 2022
/
Revised: 18 April 2023
/
Accepted: 18 May 2023
/
Published: 26 May 2023
(This article belongs to the Special Issue Mediterranean Diet, Cardiovascular Diseases, Aging and Frailty)
Abstract
:The beneficial effects of a Mediterranean diet are due to the numerous active compounds in the food and, particularly, the high concentration of compounds with synergistically acting antioxidant properties. Resveratrol, a stilbenoid nonflavonoid phenol, is an antioxidant that is naturally produced by numerous plants as a defensive agent in response to attacks from pathogens, such as bacteria and fungi. Resveratrol has several effects on human health, including on the lipid profile, where it primarily downregulates the enzyme 3-hydroxy-3-methylglutaryl coenzyme A reductase, reducing the synthesis of cholesterol. Resveratrol also increases the expression of LDL receptors in the liver, contributing to the reduction in the LDL-cholesterol levels. This short narrative review, based on relevant articles written in English from a PubMed search, using the keywords “resveratrol”, “atherosclerosis”, “cardiovascular disease”, and “Mediterranean Diet“, focuses on the possible effects of this molecule on cardiovascular disease, lipid metabolism, and atherosclerosis.
1. Introduction
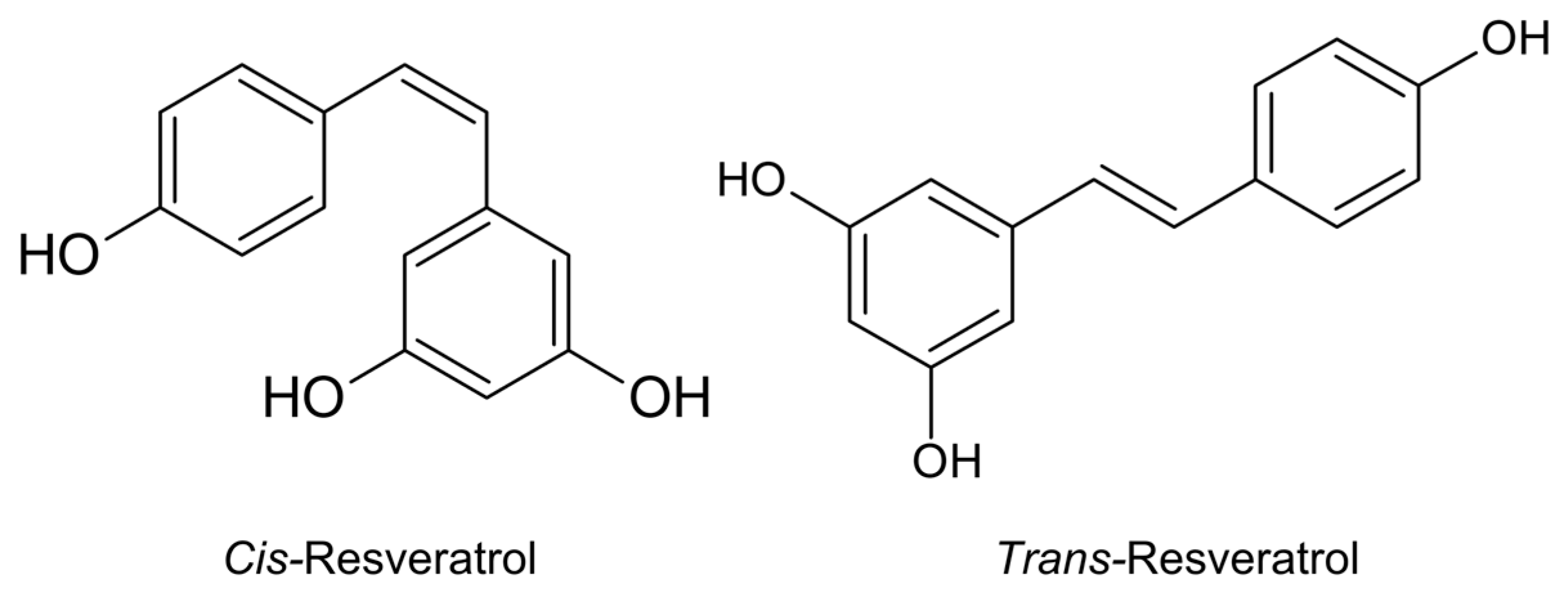
Resveratrol (RSV) is a stilbenoid, or a nonflavonoid phenol, that is naturally produced by numerous plants as a defensive agent in response to attacks from pathogens, such as bacteria or fungi. The chemical structure of RSV consists of two aromatic rings linked by a methylene bridge. RSV naturally occurs in two isomeric forms, cis and trans (Figure 1), however the trans form is the most stable and biologically active [1,2]. RSV is mostly contained in the peel of grape berries, and the other common sources of RSV are peanuts, mulberries, blueberries, bilberries, soybeans, pomegranates, cranberries, soy, dark chocolate, pistachios, and Polygonum cuspidatum [3]. Plants usually produce RSV in response to mechanical injuries and ultraviolet radiation and as a defense against viral and fungal infections. In common foods, the RSV concentrations widely varies from 19 mg/g in tomato skin [4] to 350 mg/kg in dark chocolate, 100 mg/kg in milk chocolate [5], 68 mg/100 mL in Itadori tea [6], 59–1759 mg/kg in white grapes, and 400 mg/kg in fresh apples [7,8]. The cardioprotective, antitumor, antidiabetic, antioxidant, and neuroprotective effects of RSV, as well as its effects on glucose metabolism, are well known.
In humans, the total RSV absorption rate in the intestine is as high as 70%. RSV is rapidly metabolized after oral intake and is finally eliminated by the kidneys. Therefore, its plasma bioavailability is only 20% of the oral intake. Resveratrol also rapidly metabolizes into the corresponding sulfate and glucuronic acid conjugates due to specific enzymes and intestinal microorganisms [9,10,11,12,13,14,15]. In addition, the occurrence of mild to moderate dose-related side effects, as well as the ability of RSV to interfere with the activity of drug-metabolizing enzymes, point to an optimal dosage between 100 and 1000 mg/day [10].
RSV has protective properties against DNA damage and the resulting initial mutations, consequently showing a detoxifying activity and the suppression of the carcinogenic metabolic activation. This is also due to decreased plasma levels of insulin-like growth factor-1 (IGF-1) and IGF-binding protein-3 (IGFBP-3) with a reduction in their mitogenic and antiapoptotic activity [16].
One of the most studied RSV mechanisms is the increased energy expenditure from modulating the protein targets by inducing mitochondrial biogenesis. RSV can directly activate the sirtuins, specifically sirtuin1 (SIRT1) and sirtuin5 (SIRT5), which act as caloric restriction mimetics, thus eliciting the deacetylation of peroxisome proliferator-activated receptor-gamma coactivator-1 alpha (PGC1-α), which is a key regulator of energy metabolism [17], leading to decreased glycolysis in the muscles and liver and increased lipid use [18,19]. This benefits longevity and the prevention of age-related complications. In addition, RSV inhibits ATP production by interfering with the mitochondrial function, leading to an increase in the AMP/ATP ratio, which activates the AMP-activated protein kinase (AMPK), a pivotal protein governing energy homeostasis. Furthermore, AMPK may inhibit the mammalian target of rapamycin (mTOR) signaling, which is associated with an extended lifespan, given its anti-aging effects [20].
Another important aspect of RSV is its antioxidant effects. Although the in vitro antioxidant action of RSV is far superior to that of other antioxidants, its plasma concentration in humans is generally low due to its low bioavailability. Therefore, the antioxidant effect of RSV in humans is weak unless large quantities are consumed [21]. The most important antioxidant effect is manifested on the low-density lipoprotein (LDL), which is protected from oxidation by the chelation of copper, an essential element for the oxidative process of the LDL [22,23].
Teimouri et al. [24] proposed a systematic review where they showed that RSV might exerts a powerful protective action against atherosclerosis. RSV also acts on blood platelets with an anti-inflammatory and antithrombotic mechanism. RSV has anti-inflammatory effects, especially in patients with cardiovascular disease, specifically by reducing serum levels of C-reactive proteins and TNF-alpha [24]. The anti-inflammatory action occurs by blocking cyclooxygenase, a platelet enzyme that is important in the transformation of arachidonic acid into inflammatory prostaglandins. The antithrombotic activity is due to the inhibition of the synthesis of thromboxane, a prothrombotic eicosanoid, with a consequent antiaggregating effect on the platelets [25].
Most importantly, due to its antioxidant, anti-inflammatory, immunoregulatory, and protective properties of vascular functions, and by improving neurohumoral regulation, RSV regulates the cardiomyocyte proliferation, promoting autophagy and inhibiting myocardial fibrosis and its impact on mitochondrial degradation and lipid metabolism, thus maintaining the normal function of the myocardial cells. Therefore, resveratrol can inhibit pathological cardiac remodeling, thus delaying the onset and development of cardiovascular disease [26].
RSV contributes to cardioprotection against ischemia–reperfusion injury in the myocardial tissue. In particular, RSV activates the nuclear factor erythroid-2-related factor 2 (Nrf2) signaling pathway, thus increasing the expression of the antioxidant enzymes. RSV also reduces the expression of the proapoptotic proteins, such as caspase-3, through the (phosphoinositide 3-kinases/protein kinase B (PI3K)/Akt pathway. The other effects observed after the RSV administration are a reduction in the inflammation, which is associated with a reduced expressions of the tumor necrosis factor-α (TNF-α), nuclear factor kappa-B (Nfk-B), transferrin receptor-1 (TfR1) and ferroptosis, which is associated with an increased glutathione peroxidase-4 (GPX4) activity. Furthermore, through the activation of SIRT1, resveratrol administration is associated with reduced mitochondrial reactive oxygen species (ROS) production due to a decrease in the intramitochondrial calcium concentration [27].
RSV has the potential to influence the increase in the species within the gut microbiota and alter its composition. Therefore, it may be able to influence the composition of the bacterial species in the gut in favor of human health [28]. In addition, RSV acts by attenuating the progression of the atherosclerotic plaque induced by trimethylamine N-oxide (TMAO), which is a metabolite of the intestinal flora that plays a substantial role in cardiovascular disease (CVD).
The Mediterranean diet (MD) is one of the effective model diets for delaying the effect of aging and preventing CVD, cancer, and degenerative diseases [5,6,7,8]. The beneficial effects of the MD are due to the high concentration and synergistic effect of the phytochemical components of the foods in this diet, such as polyphenols, sterols, polyunsaturated fatty acids, and vitamins [29,30]
The purpose of this brief narrative review was to analyze the available evidence to increase our understanding of the cardioprotective effects of RSV, i.e., the inhibitory action of RSV on the lipid metabolism and the progression of atherosclerotic disease.
2. Methodology
We performed a narrative review of the literature using PubMed with the following keywords: “resveratrol”, “atherosclerosis”, “cardiovascular disease”, and “Mediterranean Diet“. We focused primarily on articles written in English and original research based on its importance and relevance to this field. Our selection of references focused on the most impactful and clinically relevant reports and included published review articles. Where relevant, we focused on more recently published articles.
3. RSV Effects on Atherosclerosis Progression and Lipid Metabolism
The development and progression of atherosclerosis can be attributed to multiple risk factors, such as hypercholesterolemia, hypertension, age, sex, diabetes, impaired glucose tolerance, smoking, obesity, and genetic factors. All these are commonly known as traditional risk factors, which often coexist in the same individual and can aggravate each other, amplifying the mechanism of pathogenesis and the progression of atherosclerotic plaque.
Other risk factors, denominated as nontraditional risk factors, include poor daily lifestyle, infection, inflammation, hyperomocysteinemia, hyperuricemia, clonal hematopoiesis of indeterminate potential (CHIP), dysbiosis of the intestinal flora, depression, metabolic syndrome, hyperinsulinemia, nephrotic syndrome, organ transplantation, hyperfibrinogenemia, hypercoagulable state, obstructive sleep apnea, and extrasystolic arrhythmia, among others [31].
The transforming growth factor/extracellular regulated protein kinases (TGF/ERK) signaling pathway is also involved in the pathophysiological mechanisms that are related to the development of atherosclerosis, such as the vascular smooth muscle cell (VSMC) proliferation, inflammatory response, proliferation of fibroblasts, and accumulation of extracellular matrix [32,33]. RSV may also inhibit the TGF/ERK signaling pathway by activating SIRT1, thus potentially playing a role in slowing the development of atherosclerosis [34].
Hypertension is characterized by decreased vasodilator production and increased ROS levels in the endothelium. This leads to a decreased endothelial nitric oxide synthase (eNOS) expression and vasodilator production, including decreased nitric oxide (NO) levels. Consequently, this leads to increased vasoconstriction and blood pressure.
Several animal model studies have shown that the consumption of resveratrol, by stimulating SIRT1, Nrf2, and the AMP-activated AMPK activity, led to an increased eNOS phosphorylation and expression. This led to increases in the levels of nitric oxide and other vasodilators and a decrease in blood pressure [33].
The effect of RSV in lipid the improving profile could be due to its downregulation of the enzyme 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase, thus reducing the total cholesterol (TC) and LDL levels. RSV can also increase the expression of the LDL receptors in the liver, contributing to reductions in the LDL concentration [29,30]. Additionally, RSV has antioxidant, antiplatelet, and anti-inflammatory properties [35,36].
RSV improves glucose, lipid, and amino acid metabolism in the endothelial cells (ECs). RSV reduces glucose uptake and glycolysis by inhibiting the expression of the glucose transporter (Glut), monocarboxylate transporter (MCT), 6-phosphofructo-1-kinase (PFK), pyruvate kinase (PK), hexokinase (HK), and lactate dehydrogenase (LDH). In addition, RSV improves fatty-acid-related damage by upregulating the expression of brain and muscle arnt-like protein-1 (Bmal1). It decreases lipogenesis by suppressing the expression of fatty acid synthase (FASN) and activates fatty acid oxidation by suppressing the expression of acetyl-CoA carboxylase (ACC).
Finally, resveratrol increases the uptake and synthesis of glutamine. It upregulates the NO release by elevating the expression of eNOS, suppressing the level of serum asymmetric dimethylarginine (ADMA), and inhibiting the activity of arginase [11,37].
RSV shows a cardioprotective activity both by regulating the ROS production in cardiac cells and changing the lipid profile, and by reducing atherosclerosis progression in animal models, randomized control trials (RCTs), and prospective cohort studies.
3.1. RSV Effects on Lipid Metabolism and the Progression of Atherosclerosis: Studies on Animal Models
A case–control animal model study was conducted on guinea pigs [38], in which eight guinea pigs were provided with RSV in drinking water for 16 days (14 mg/kg body weight). Five control animals were provided with 0.2% ethanol in tap water instead of RSV. The RSV administration significantly increased the NAD(P)H:quinone oxidoreductase (DT-diaphorase) activity, expressed as Vmax (nmoles/mg protein/min) with respect to the controls (p < 0.05) without changing its affinity for the substrate, expressed as Kmax (μM) (p > 0.05). In addition, the authors measured the activities of glutathione peroxidase (Gpx, nmol/mg protein/min), glutathione reductase (GR, nmol/mg protein/min), catalase (CAT, units/mg protein), Cu/Zn superoxide dismutase (Cu/Zn-SOD, units/mg protein), and Mn superoxide dismutase (Mn-SOD, units/mg protein) in the hearts isolated from the control and the RSV-treated animals. They showed that the RSV treatment significantly increased the cardiac CAT activity (p < 0.001) but not the Gpx, GR, Cu/Zn-SOD, or Mn-SOD activity with respect to the controls. Finally, the authors measured the amount of the ROS produced by the cardiac fractions isolated from both the RSV-treated and the control guinea pigs at low (5 μM) and high (20 μM) concentrations of 2-methyl-1,4-naphthoquinone (menadione). They found that the positive inotropic effect of menadione, the activity of which is related to the amount of the ROS generated by the cardiac metabolism, substantially decreased in the left atria isolated from the RSV-treated animals compared to the controls.
A case–control study on animal models was conducted [39] to demonstrate the lipid-lowering properties of RSV. Male Apo/E-deficient mice (34 weeks old) were divided into four groups: a control group, which was fed only a standard semisynthetic diet; a second group, which was supplemented with 0.02% clofibrate (CF); a third group, which was supplemented with 0.02% RSV; and a fourth group, which was supplemented with 0.06% RSV. After 20 weeks, compared to the control group, the authors observed a significant reduction in the plasma levels (mmol/L) of the TC in the 0.02% CF, 0.02% RSV, and 0.06% RSV groups (p < 0.05). A significant reduction in the plasma levels (mmol/L) of triglycerides (TG) was also observed in the 0.02% CF, 0.02% RSV, and 0.06% RSV groups (p < 0.05). In addition, compared to the control group, a significant reduction in the plasma levels (mmol/L) of the LDL was observed in the 0.02% CF, 0.02% RSV, and 0.06% RSV groups (p < 0.05). A significant increase in the plasma levels (mmol/L) of the high-density lipoprotein (HDL) was observed after 20 weeks in all the treatment groups (p < 0.05). Consequently, the HDL/cholesterol ratio (%), which is predictive of the risk of heart disease, increased in the 0.02% CF, 0.02% RSV, and 0.06% RSV groups (p < 0.05). A decrease was observed in the atherogenic index, a strong marker predictive of the risk for coronary heart disease, which was expressed by the TC-HDL-cholesterol/HDL ratio, in the 0.02% CF, 0.02% RSV, and 0.06% RSV groups (p < 0.05). Compared to the control group, after 20 weeks, the hepatic 3-hydroxy-3-methylglutaryl-CoA reductase (HMG-CoA) activity was reduced (p < 0.05), while the acyl-CoA cholesterol acyltransferase (ACAT) activity was increased in all three treatment groups.
A case–control animal model study was conducted on three groups of 10–12-week-old female APOE*3-Leiden.CETP (E3L.CETP) transgenic mice, as a mouse model representing human CVD [40]. All the mice were fed regular chow, supplemented with RSV, atorvastatin, or RSV plus atorvastatin. In the E3L.CETP transgenic mice, the RSV supplementation reduced the progression of atherosclerotic lesions by improving the plaque stability. The authors noted a significant reduction in the total atherosclerotic lesions compared to those of the control group in the mice supplemented with RSV (p < 0.01), atorvastatin (p < 0.05), and RSV plus atorvastatin (p < 0.01). In addition, an improvement was noted in the plaque stability, expressed by the collagen/macrophage ratio in the mice supplemented with RSV (p < 0.05), atorvastatin (p < 0.01), and RSV plus atorvastatin (p < 0.001). A reduction in the plasma very-low-density lipoprotein (VLDL) levels was also observed in the mice supplemented with RSV (p = 0.08), atorvastatin (p < 0.05), and RSV plus atorvastatin (p < 0.05).
An experiment on ApoE-deficient mice [41] was conducted to assess the effects of RSV on the serum levels of the total cholesterol (TC), LDL, HDL, non-HDL, and TG and how RSV can protect against the atherosclerosis progression induced by a high-fat diet (HFD). The mice were divided into five groups: a control group (normal diet); an HFD group (1.25% cholesterol and 20% lard); an HFD plus lipopolysaccharides (LPS) group (10 mcg of LPS/mouse), with LPS as a trigger of inflammation [42,43]; an HFD plus simvastatin group (3.3 mg/kg/mouse), and a HFD plus RSV group (5 mg RSV/kg/day). Compared to the mice in the control group, after 20 weeks of the HFD plus LPS, the serum levels of the TC, TG, LDL, and non-HDL significantly increased, while the HDL level decreased. After the 20th week, in the HFD + LPS + RSV group, the serum levels of the TC, TG and non-HDL decreased, while the levels of the HDL increased. Concerning the progression of atherosclerosis plaque, after 20 weeks of the HFD plus LPS, a thickened coronary wall and a large area of atherosclerotic plaque on the aorta were observed. Both simvastatin and RSV treatment reduced the thickening of the coronary wall and the plaque area on the aorta. In addition, both simvastatin and the RSV treatments decreased the rate of the CD4+ T cells in the peripheral blood mononuclear cells (PBMCs), suggesting a reduced inflammatory status of the atherosclerotic plaque.
Similar results were obtained from a subsequent case–control animal model study [44]. Twenty-four New Zealand male rabbits were divided into three groups: a normal diet group (NDG), a fatty diet group (FDG), and a fat diet plus RSV group (RFG), in which 80 mg RSV/kg/day was added to the fat diet. The serum levels of the TG, TC, HDL, LDL, and lipoprotein-associated phospholipase A2 (Lp-PLA2), as a predictor biomarker for cardiovascular risk assessment [45], were measured. After three months, the authors observed a significant decrease in the serum levels of the TC, HDL, LDL, and Lp-PLA2 (p < 0.001), but not the TG in the RFG with respect to the FDG (p = 0.232). Furthermore, the intimal thickness and the intima/media ratio, but not the smooth muscle layer, of the RFG was significantly lower than that of the FDG (p < 0.001; p = 0.65, respectively). This was indicative of a slower progression toward atherosclerotic plaque. The authors concluded that RSV, by reducing the serum levels of the TC, LDL, and Lp-PLA2, might inhibit the thickening of the arterial intima. In addition, RSV may inhibit the production of proinflammatory cytokines, reduce the atherosclerotic effect of Lp-PLA2, and reduce the peroxidation of the LDL by macrophages by its absorption.
All the animal model studies are summarized in Table 1.
3.2. RSV Effects on Lipid Metabolism and Progression of Atherosclerosis on Humans: RCTs and Prospective Cohort Studies
A meta-analysis of the data from 10 RCTs [46] involving 600 subjects evaluated the efficacy of the RSV intake on the plasma concentration, expressed as the weighed mean difference (WMD) of the C-reactive protein (CRP), TC, LDL, HDL, TG, and glucose; systolic blood pressure (SBP); and diastolic blood pressure DBP as the selected predictors of the CVD risk. Of the 600 subjects, 344 received the RSV supplementation at a dose ranging from 8 to 1500 mg/day of RSV or trans-RSV. The duration of the RSV supplementation ranged from 60 days to 12 months. No significant effect was found for the RSV supplementation on the changes in plasma CRP (p = 0.731), TC (p = 0.859), LDL (p = 0.948), TG (p = 0.866), or glucose (p = 0.703) concentrations. Only a slight reduction in the HDL (p = 0.001) was observed. No significant change was found in either the SBP (p = 0.868) or the DBP (p = 0.674). However, the authors acknowledged the limitations of their meta-analysis, i.e., few RCTs were included, the heterogeneity concerning the patient characteristics and study design, and the low number of subjects.
In a triple-blind, randomized, placebo-controlled study [47], 75 patients with stable CAD received 350 mg/day of a placebo or grape extract containing phenolics plus 8 mg RSV or a conventional grape extract lacking RSV for 6 months with a double dose for the following 6 months. In the placebo group, after 12 months, the interleukin-10 (IL-10) and adiponectin serum levels significantly decreased by 6.6% (p = 0.03) and 12.7% (p = 0.01) vs. the baseline, respectively, whereas the plasminogen activator inhibitor type 1 (PAI-1) serum levels significantly increased by 38.5% (p < 0.001). In the RSV group, the serum adiponectin levels increased by 10% (p = 0.01) and the adiponectin serum values were 23% higher than in the placebo group (p < 0.05). An 18% (p = 0.05) decrease in the serum PAI-1 and high-sensitivity C-reactive protein (hs-CRP) levels (p = 0.17) were also observed. In the RSV-group, a significant (p < 0.05) downregulation was observed in the gene expression of six proinflammatory factors, namely, the Kruppel-like factor 2 (KLF2), NF-kB, activator protein 1 (Ap-1), proto-oncogene c-JUN (JUN), activating transcription factor 2 (ATF-2), and CREB-binding protein (CREBBP). These data suggested an inhibition of the lymphocytes/monocytes-mediated inflammatory response, with the subsequent inhibition of the inflammatory response, by the PBMCs in the RSV-containing grape extract group.
A prospective case–control study was subsequently conducted on twenty-five subjects with a BMI > 30 mg/dL and metabolic syndrome [48]. All the patients were randomly divided into a placebo group (nine subjects: physical activity program + diet + placebo), and an RSV group (thirteen subjects: physical activity program + diet + RSV, 250 mg/day).
The authors evaluated the effects of the oral RSV supplementation associated with an MD model and a physical training program on anthropometric parameters, such as the body weight, body mass index (BMI), and waist circumference (WC). The biochemical parameters were also evaluated, i.e., insulin, glycemia, hemoglobin A1c (HBA1c), leptin, uric acid, fibrinogen, urea, creatinine, albumin, TC, LDL, HDL, VLDL, TG, alanine aminotransferase (AST), and aspartate aminotransferase (ALT). After 12 weeks, the subjects treated with the physical activity program + diet + placebo showed significant reductions in the body weight (p = 0.006) and BMI (p = 0.003), WC (p = 0.001), and the levels of HBA1c (p = 0.005) and leptin (p = 0.031). The subjects treated with the physical activity program + diet + RSV 250 mg/day showed significant reductions in the body weight (p = 0.000) and BMI (p = 0.000), and the WC (p = 0.000), TC (p = 0.031), VLDL (p = 0.025), leptin (p = 0.014), urea (p = 0.046), creatinine (p = 0.021), and albumin (p = 0.000) levels. In addition, among the patients in the RSV group, a significant increase in the HDL (p = 0.026) levels was observed.
A prospective cohort study [49], involving a sample of 2318 men from the Aragon Workers Health Study (AWHS) [50], found that the subjects with the highest total polyphenol intake consumed higher amounts of fruits, vegetables, virgin olive oil, coffee, nuts, chocolate, and red wine. Notably, a higher intake of stilbenes, mainly RSV, was associated with a reduction of up to 38% in the risk of developing atherosclerotic plaques in the femoral arteries (p = 0.009). A similar result was observed regarding a high flavonoid intake (p < 0.001). A slight but not significant reduction in the risk of developing atherosclerotic plaques in the carotid arteries was observed, which was associated with higher intakes of flavonoids (p = 0.094) and stilbenes (p = 0.536). In addition, the risk of developing atherosclerotic plaques in the femoral arteries was reduced by 4% for each 100 mg/day increase in the intake of flavonoids and by 3% for each 1 mg/day increase in the intake of stilbenes. Finally, a slight but not significant reduction in the coronary calcium, evaluated by the coronary artery calcium (CACS), which is known to be a marker of coronary damage and an independent predictor of CHD [51,52,53], was observed, associated with the increased intakes of flavonoids (p = 0.346) and stilbenes (p = 0.131).
The effect of the RSV supplementation on the biomarkers of atherosclerosis was assessed in a review that pooled the biochemical parameters from 27 studies, with a mean sample size of 26 subjects [54], at baseline and postintervention. Based on the clinical relevance and a higher variation level, 12 biomarkers of atherosclerosis were selected, i.e., systolic SBP, DBP, TC, TG, LDL, HDL, hsCRP, TNF-α, IL-6, IL-10, leptin, and adiponectin. All the studies were divided into three clusters, where the patients in Clusters II and III presented comorbidities, such as type 2 diabetes mellitus (T2DM), dyslipidemia, hypertension, obesity, and stable angina. The clusters differed in the study duration (38, 74, and 175 days) and RSV dosage (1067, 454, and 264 mg). The patients in Cluster II (74.21 ± 10.36 days of study duration, 454.14 ± 129 mg of RSV) showed the best results in terms of the percentage of reductions in the SBP (p < 0.001), DBP (p = 0.026), TC (p < 0.001), LDL (p = 0.036), and TG (p < 0.001) levels. The patients in Cluster III (175.33 ± 40.96 days of study duration, 263.75 ± 88.07 mg of RSV) showed the best results in terms of the percentage of increases in the HDL (p < 0.001). No statistically significant changes were observed in the values of hsCRP, TNF-α, IL-6, IL-10, leptin, or adiponectin in Cluster I, Cluster II, or Cluster III.
More specifically considering the efficacy of the RSV supplementation on the lipid and glucose metabolism, a meta-analysis involving 1171 subjects was conducted [55] to further clarify the relationship between the RSV supplementation and lipid and glucose metabolism. All 25 articles were randomized placebo-controlled trials. The following parameters were considered: the TC, TG, LDL, HDL, body weight, BMI, waist circumference (WC), HbA1c, HOMA index, insulin, leptin, fasting glucose, fat percentage, and adiponectin level. Continuous data were pooled using the standardized mean difference (SMD) with a 95% confidence interval (CI) to calculate the effects of the RSV supplementation. The authors described significant reductions in the WC (p = 0.002), HbA1c (p < 0.001), TC (p = 0.003), LDL (p < 0.001), and HDL (p = 0.03) levels. No significant changes were observed in the body weight (p = 0.18), BMI (p = 0.83), fasting glucose (p = 0.14), insulin (p = 0.42), HOMA index (p = 0.13), fat percentage (p = 0.06), TG (p = 0.53), adiponectin (p = 0.89), or leptin (p = 0.38). Overall, significant results were observed from the intervention studies lasting longer than 17 weeks and applying a daily RSV intake of 200–500 mg/day. Concerning the lipid metabolism, significant reductions in the TC and HDL levels were observed in the intervention studies lasting less than 8 weeks. A more significant reduction in the LDL level was observed for the intervention trials ranging from 9–16 weeks. Regarding the dosage, significant reductions in the LDL and HDL levels were observed in the trials with a dose of 200 mg/day. Concerning the TC, a significant reduction occurred in the trials with a dose of 200–500 mg/day.
Finally, the EC is not only a simple internal cellular lining of the blood vessels. Instead, it plays a substantial role in the regulation of immunoproliferative–inflammatory processes and angiogenesis, thus constituting an active and fundamental component in the development process of atherosclerosis. A meta-analysis involving 1034 adult subjects from 17 RCTs evaluated the effect of the RSV supplementation on the endothelial functions, including the flow-mediated dilation (FMD), intercellular adhesion molecule-1 (ICAM-1), vascular cell adhesion molecule-1 (VCAM-1), fibrinogen, and plasminogen activator inhibitor-1 (PAI-1) serum levels, expressed as the weighted mean difference (WMD) [56]. The RSV supplementation significantly increased the FMD, but only in cross-over studies (WMD: 1.59%; 95% CI: 1.05–2.13, p < 0.001) at RSV dosages of <100 mg (WMD: 1.69%; 95% CI:1.30–2.08, p < 0.001) in the trials with acute intervention (WMD: 2.28%; 95% CI: 1.80–2.76, p < 0.001) and in the studies that included people with CVD (WMD: 2.60%; 95% CI: 1.95–3.26, p < 0.001). The RSV supplementation significantly decreased the ICAM-1 (WMD: −7.09 ng/mL, 95% CI: −7.45 to −6.73, p < 0.001) but not VCAM-1 (WMD: −15.04 ng/mL, 95% CI: −37.52 to −7.45, p = 0.19) serum levels. Finally, the RSV supplementation had no significant effect on either the fibrinogen (WMD: 0.25 g/L, 95% CI: −0.36–0.86, p = 0.42) or PAI-1 (WMD: −0.94 ng/mL, 95% CI: −3.51–1.62, p = 0.47) serum levels.
All the human studies are summarized in Table 2.
3.3. RSV Effects on Atherosclerosis Progression by Regulating the TMAO Synthesis via the Remodeling of Gut Microbiota
The gut microbiota can regulate several metabolic processes in the host, including the lipid metabolism [57,58]. The mechanisms through which dietary fatty acids affect the gut microbiota are poorly known. Most consumed fatty acids are absorbed in the small intestine. However, a minority pass through the gastrointestinal tract and modulate the colonic microbiota composition. Fatty acids have a broad spectrum of antibacterial activity, such as the lysis and solubilization of bacterial cell membranes or the inhibition of ATP production. The antibacterial action of fatty acids is affected by the carbon chain length, saturation, and double bond position. Fatty acids may also be used as metabolic substrates by the gut bacteria, thereby affecting the gut microbiota profile and the production of microbial metabolite [59,60,61,62].
TMAO is a phospholipid metabolite related to intestinal microorganisms. It is predominantly sourced from choline, which is found in foods such as red meat, fish, poultry, and eggs, or from L-carnitine, which is found in red meat and dairy products. Both choline and L-carnitine are metabolized by the gut microbiota into trimethylamine (TMA) and then into TMAO through the action of the hepatic enzyme, flavin monooxygenase-3 (FMO3). Studies have recently suggested that TMAO is a new independent risk factor for atherosclerosis. The plasma level of TMAO is reportedly positively associated with cardiovascular risk and mortality in a dose-dependent manner. The higher the concentration of TMAO in the plasma, the higher the probability of malignant cardiovascular and cerebrovascular events. A recent prospective case–control study showed that in a 10-year period, regardless of the level of baseline TMAO, the final increase in TMAO was significantly associated with an increased risk of CHD, and the TMAO–CHD relationship could be improved through dietary changes [63,64,65,66,67,68,69,70,71].
TMAO can hasten the atherosclerosis progression through the mechanisms related to the lipid metabolism and inflammation. More specifically, TMAO promotes oxidative stress and inflammation in the endothelial cells, thus aggravating the vascular dysfunction. TMAO also aggravates the process of the atherosclerotic plaque formation by promoting vascular calcification. It promotes the conversion of macrophages into foam cells and the platelet activation process. In addition, TMAO contributes to increases in the serum cholesterol levels and the progression of atherosclerosis (AS) by reversing the cholesterol transport and by inhibiting bile acid (BA) synthesis [64,66]. TMAO also induces increases in the oxidative stress and inflammation of endothelial cells, leading to an increase in the endothelial-derived NOS expression [53], thus contributing to endothelial dysfunction [72].
Resveratrol is characterized by a poor bioavailability when ingested through red wine, other foods, or food supplements. When taken orally, only 1–8% of free resveratrol is found in the serum, 25% is excreted without absorption, and over 70% is metabolized by the liver and the intestinal microbiota [62,73]. Nonetheless, resveratrol has broad antibacterial activity at concentrations that can be reached even with a dietary intake. Specifically, resveratrol seems to inhibit the bacteria involved in the saccharolytic and proteolytic activities [74].
RSV acts by lowering the TMAO levels and increasing hepatic bile acid synthesis via remodeling the gut microbiota [30]. Specifically, RSV has antibacterial properties against various bacterial species, including opportunistic intestinal pathogens, for example, Escherichia coli, Enterococcus faecalis, and Salmonella enterica [75,76,77]. The bactericidal effect of resveratrol is, however, weaker toward commensal bacterial species such as the Lactobacillus spp. [78]. RSV can, therefore, influence the composition of bacterial species in the intestine in favor of maintaining the state of health.
In summary, by modifying the composition of the intestinal flora, RSV induces reductions in the levels of microbial production of intestinal TMA, subsequently leading to a decrease in the synthesis of TMAO in the liver and, ultimately, to the inhibition of the progression of AS. Additionally, by remodeling the intestinal microbiota, RSV increases the enzymatic activity of bile salt hydrolase (BSH). This induces an increase in the production of unconjugated bile acid (BA) from the conjugated BA. This involves an increase in the fecal excretion and a contextual induction of hepatic BA neosynthesis, thus contributing to reductions in the serum cholesterol levels and the inhibition of the AS progression [79].
4. Concluding Remarks
Although the evidence provided strong results, the final effect of resveratrol in humans is not yet fully understood. This is due to the variability in the dosages used in the studies, the heterogeneity of the experimental studies conducted on small samples of subjects, the low statistical power of the same studies, and a myriad of biological and other biases [20].
Concerning the cardioprotective effects of the RSV treatment, the study results were inconsistent, mainly due to the wide variability between the clinical studies regarding the RSV dosage, treatment duration, and number and characteristics of the subjects examined. Consequently, no widespread consensus has been reached regarding whether RSV has protective effects on cardiovascular health [80,81].
The mechanisms of action of RSV are pleiotropic, varying in potential benefits for cardiovascular health, obesity, metabolic health, inflammation, and for cancer management. From the reported evidence, only one study [46] did not find a significant effect of the RSV supplementation on the plasma concentrations of CVD risk markers. This result must be attributed, as recognized by the authors themselves, to several factors; first to the small number of studies included (only 10) and second to the small number of subjects included in the studies (600 total, of which only 344 were administered RSV). This was due to the heterogeneity of the duration of the studies (60 days to 12 months) and the heterogeneity of the dosage (from 8 to 1500 mg/day) of the different compounds administered to the patients, namely RSV, trans-RSV, extract grape juice containing RSV, or Chuanhu anti-gout blend containing 149.2 μg/mL RSV.
Most of the evidence from the studies conducted both on animal models and human populations found that RSV showed a cardioprotective activity by regulating the ROS production in cardiac cells and modifying the lipid profiles, thus reducing the progression of atherosclerosis. In addition, RSV can promote vasodilation by stimulating the K+ channels activated by Ca2+ and decreasing the inactivation of nitric oxide by free radicals in the endothelium.
Overall, the evidence confirmed that RSV is one of the most important nutraceuticals belonging to the stilbenoid group. The potent antioxidant, antiplatelet, and anti-inflammatory properties of RSV explain its protective and health-promoting properties [82,83]. The studies on RSV metabolites seem to be encouraging, as these either have similar effects or can act as an RSV pool in the body, fostering the metabolic effects previously solely attributed to free RSV [84].
Currently, RSV is widely marketed primarily as a dietary supplement, available without a prescription, and as an antioxidant with generic protective effects.
In conclusion, although a wide range of in vitro and in vivo evidence shows that RSV is a promising therapeutic agent, the evidence reviewed must not lead to an erroneous conclusion about the unique role of RSV as a miraculous health protector considering the numerous other phenolic compounds found in fruit and vegetables. The only advisable indication is to adopt a healthy dietary model, for example, the MD, composed of a wide variety of unprocessed plant-based foods that are rich in RSV.
The hope is that further clinical studies will be conducted to further understand the bioavailability, metabolic pathways, and human toxicity of RSV and to confirm its potential not as a nutraceutical or prebiotic but as a real drug.
Author Contributions
All authors listed made a substantial, direct, and intellectual contribution to the study and approved it for publication. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The Fondazione Dieta Mediterranea, Ostuni, Italy endorsed this review paper—https://www.fondazionedietamediterranea.it/.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aggarwal, B.B.; Bhardwaj, A.; Aggarwal, R.S.; Seeram, N.P.; Shishodia, S.; Takada, Y. Role of resveratrol in prevention and therapy of cancer: Preclinical and clinical studies. Anticancer Res. 2004, 24, 2783–2840. [Google Scholar] [PubMed]
- Frémont, L. Biological effects of RSV. Life Sci. 2000, 66, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-X.; Li, C.-X.; Kakar, M.U.; Khan, M.S.; Wu, P.-F.; Amir, R.M.; Dai, D.-F.; Naveed, M.; Li, Q.-Y.; Saeed, M.; et al. Resveratrol (RV): A pharmacological review and call for further research. Biomed. Pharmacother. 2021, 143, 112164. [Google Scholar] [CrossRef] [PubMed]
- Ragab, A.S.; Jennifer, V.F.; Boris, J.; Joon-Hyun, P.; Bobzin, S.C. Detection and quantitation of resveratrol in tomato fruit (Lyco-persicon esculentum Mill.). J. Agric. Food Chem. 2006, 54, 7175–7179. [Google Scholar] [CrossRef]
- Hurst, W.J.; Glinski, J.A.; Miller, K.B.; Apgar, J.; Davey, M.H.; Stuart, D.A. Survey of the trans-Resveratrol and trans-Piceid Content of Cocoa-Containing and Chocolate Products. J. Agric. Food Chem. 2008, 56, 8374–8378. [Google Scholar] [CrossRef]
- Um, J.-H.; Park, S.-J.; Kang, H.; Yang, S.; Foretz, M.; McBurney, M.W.; Kim, M.K.; Viollet, B.; Chung, J.H. AMP-Activated Protein Kinase–Deficient Mice Are Resistant to the Metabolic Effects of Resveratrol. Diabetes 2010, 59, 554–563. [Google Scholar] [CrossRef]
- Arichi, H.; Kimura, Y.; Okuda, H.; Baba, K.; Kozawa, M.; Arichi, S. Effects of stilbene components of the roots of Polygonum cuspidatum Sieb. et Zucc. on lipid metabolism. Chem. Pharm. Bull. 1982, 30, 1766–1770. [Google Scholar] [CrossRef]
- Sanchez-Ballesta, M.T.; Alvarez, I.; Escribano, M.I.; Merodio, C.; Romero, I. Effect of high CO2 levels and low temperature on stilbene biosynthesis pathway gene expression and stilbenes production in white, red and black table grape cultivars during postharvest storage. Plant Physiol. Biochem. 2020, 151, 334–341. [Google Scholar] [CrossRef]
- Walle, T.; Hsieh, F.; DeLegge, M.H.; Oatis, J.E., Jr.; Walle, U.K. High absorption but very low bioavailability of oral resveratrol in humans. Drug Metab. Dispos. 2004, 32, 1377–1382. [Google Scholar] [CrossRef]
- Vesely, O.; Baldovska, S.; Kolesarova, A. Enhancing Bioavailability of Nutraceutically Used Resveratrol and Other Stilbenoids. Nutrients 2021, 13, 3095. [Google Scholar] [CrossRef]
- Raj, P.; Thandapilly, S.J.; Wigle, J.; Zieroth, S.; Netticadan, T. A Comprehensive Analysis of the Efficacy of Resveratrol in Atherosclerotic Cardiovascular Disease, Myocardial Infarction and Heart Failure. Molecules 2021, 26, 6600. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Shin, Y.G.; Chow, A.; Li, Y.; Kosmeder, J.W.; Lee, Y.S.; Hirschelman, W.H.; Pezzuto, J.M.; Mehta, R.G.; van Breemen, R.B. Human, Rat, and Mouse Metabolism of Resveratrol. Pharm. Res. 2002, 19, 1907–1914. [Google Scholar] [CrossRef] [PubMed]
- Böhmdorfer, M.; Szakmary, A.; Schiestl, R.H.; Vaquero, J.; Riha, J.; Brenner, S.; Thalhammer, T.; Szekeres, T.; Jäger, W. Involvement of UDP-Glucuronosyltransferases and Sulfotransferases in the Excretion and Tissue Distribution of Resveratrol in Mice. Nutrients 2017, 9, 1347. [Google Scholar] [CrossRef] [PubMed]
- Bode, L.M.; Bunzel, D.; Huch, M.; Cho, G.-S.; Ruhland, D.; Bunzel, M.; Bub, A.; Franz, C.M.; Kulling, S.E. In vivo and in vitro metabolism of trans-resveratrol by human gut microbiota. Am. J. Clin. Nutr. 2013, 97, 295–309. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C. Interactions of gut microbiota with dietary polyphenols and consequences to human health. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 471–476. [Google Scholar] [CrossRef]
- Patel, K.R.; Scott, E.; Brown, V.A.; Gescher, A.J.; Steward, W.P.; Brown, K. Clinical trials of resveratrol. Ann. NY Acad. Sci. USA 2011, 1215, 161–169. [Google Scholar] [CrossRef]
- Liang, H.; Ward, W.F. PGC-1α: A key regulator of energy metabolism. Adv. Physiol. Educ. 2006, 30, 145–151. [Google Scholar] [CrossRef]
- Lagouge, M.; Argmann, C.; Gerhart-Hines, Z.; Meziane, H.; Lerin, C.; Daussin, F.; Messadeq, N.; Milne, J.; Lambert, P.; Elliott, P.; et al. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1α. Cell 2006, 127, 1109–1122. [Google Scholar] [CrossRef]
- Houtkooper, R.H.; Pirinen, E.; Auwerx, J. Sirtuins as regulators of metabolism and healthspan. Nat. Rev. Mol. Cell Biol. 2012, 13, 225–238. [Google Scholar] [CrossRef]
- Springer, M.; Moco, S. Resveratrol and Its Human Metabolites—Effects on Metabolic Health and Obesity. Nutrients 2019, 11, 143. [Google Scholar] [CrossRef]
- Bitterman, J.L.; Chung, J.H. Metabolic effects of resveratrol: Addressing the controversies. Cell. Mol. Life Sci. 2014, 72, 1473–1488. [Google Scholar] [CrossRef] [PubMed]
- Belguendouz, L.; Frémont, L.; Gozzelino, M.-T. Interaction of Transresveratrol with Plasma Lipoproteins. Biochem. Pharmacol. 1998, 55, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Berrougui, H.; Grenier, G.; Loued, S.; Drouin, G.; Khalil, A. A new insight into resveratrol as an atheroprotective compound: Inhibition of lipid peroxidation and enhancement of cholesterol efflux. Atherosclerosis 2009, 207, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Teimouri, M.; Homayouni-Tabrizi, M.; Rajabian, A.; Amiri, H.; Hosseini, H. Anti-inflammatory effects of resveratrol in patients with cardiovascular disease: A systematic review and meta-analysis of randomized controlled trials. Complement. Ther. Med. 2022, 70, 102863. [Google Scholar] [CrossRef]
- Duffy, S.J.; Vita, J.A. Vita. Effects of phenolics on vascular endothelial function. Curr Opin Lipidol. 2003, 14, 21–27. [Google Scholar] [CrossRef]
- Olas, B.; Wachowicz, B. Resveratrol, a phenolic antioxidant with effects on blood platelet functions. Platelets 2005, 16, 251–260. [Google Scholar] [CrossRef]
- Fan, S.; Hu, Y.; You, Y.; Xue, W.; Chai, R.; Zhang, X.; Shou, X.; Shi, J. Role of resveratrol in inhibiting pathological cardiac remodeling. Front. Pharmacol. 2022, 13, 3452. [Google Scholar] [CrossRef]
- Rodrigo, R.; Retamal, C.; Schupper, D.; Vergara-Hernández, D.; Saha, S.; Profumo, E.; Buttari, B.; Saso, L. Antioxidant Cardioprotection against Reperfusion Injury: Potential Therapeutic Roles of Resveratrol and Quercetin. Molecules 2022, 27, 2564. [Google Scholar] [CrossRef]
- Bird, J.K.; Raederstorff, D.; Weber, P.; Steinert, R.E. Cardiovascular and Antiobesity Effects of Resveratrol Mediated through the Gut Microbiota. Adv. Nutr. Int. Rev. J. 2017, 8, 839–849. [Google Scholar] [CrossRef]
- Shao, D.; Wang, Y.; Huang, Q.; Shi, J.; Yang, H.; Pan, Z.; Jin, M.; Zhao, H.; Xu, X. Cholesterol-Lowering Effects and Mechanisms in View of Bile Acid Pathway of Resveratrol and Resveratrol Glucuronides. J. Food Sci. 2016, 81, H2841–H2848. [Google Scholar] [CrossRef]
- Yashiro, T.; Nanmoku, M.; Shimizu, M.; Inoue, J.; Sato, R. Resveratrol increases the expression and activity of the low density lipoprotein receptor in hepatocytes by the proteolytic activation of the sterol regulatory element-binding proteins. Atherosclerosis 2012, 220, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Zhou, Y.; Xie, X. Resveratrol inhibiting TGF/ERK signaling pathway can improve atherosclerosis: Backgrounds, mechanisms and effects. Biomed. Pharmacother. 2022, 155, 113775. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wang, Y.; Li, Y.; Xia, X.; Zhao, S.; Che, Y.; Sun, Y.; Lei, L. TGF-β1 promotes bovine mammary fibroblast proliferation through the ERK 1/2 signalling pathway. Cell Biol. Int. 2016, 40, 750–760. [Google Scholar] [CrossRef] [PubMed]
- Suwanabol, P.A.; Seedial, S.M.; Shi, X.; Zhang, F.; Yamanouchi, D.; Roenneburg, D.; Liu, B.; Kent, K.C. Transforming growth factor-β increases vascular smooth muscle cell proliferation through the Smad3 and extracellular signal-regulated kinase mitogen-activated protein kinases pathways. J. Vasc. Surg. 2012, 56, 446–454.e1. [Google Scholar] [CrossRef] [PubMed]
- DiNatale, J.C.; Crowe-White, K.M. Effects of resveratrol supplementation on nitric oxide-mediated vascular outcomes in hy-pertension: A systematic review. Nitric Oxide 2022, 129, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Wiciński, M.; Socha, M.; Walczak, M.; Wódkiewicz, E.; Malinowski, B.; Rewerski, S.; Górski, K.; Pawlak-Osińska, K. Beneficial Effects of Resveratrol Administration—Focus on Potential Biochemical Mechanisms in Cardiovascular Conditions. Nutrients 2018, 10, 1813. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.; Liu, C.; Zhang, Z.; Huang, K.; Wang, T.; Chen, S.; Li, Z. Progress in the Preclinical and Clinical Study of Resveratrol for Vascular Metabolic Disease. Molecules 2022, 27, 7524. [Google Scholar] [CrossRef]
- Su, M.; Zhao, W.; Xu, S.; Weng, J. Resveratrol in Treating Diabetes and Its Cardiovascular Complications: A Review of Its Mechanisms of Action. Antioxidants 2022, 11, 1085. [Google Scholar] [CrossRef]
- Floreani, M.; Napoli, E.; Quintieri, L.; Palatini, P. Oral administration of trans-resveratrol to guinea pigs increases cardiac DT-diaphorase and catalase activities, and protects isolated atria from menadione toxicity. Life Sci. 2003, 72, 2741–2750. [Google Scholar] [CrossRef]
- Do, G.-M.; Kwon, E.-Y.; Kim, H.-J.; Jeon, S.-M.; Ha, T.-Y.; Park, T.; Choi, M.-S. Long-term effects of resveratrol supplementation on suppression of atherogenic lesion formation and cholesterol synthesis in apo E-deficient mice. Biochem. Biophys. Res. Commun. 2008, 374, 55–59. [Google Scholar] [CrossRef]
- Berbée, J.F.; Wong, M.C.; Wang, Y.; van der Hoorn, J.W.; Khedoe, P.P.; van Klinken, J.B.; Mol, I.M.; Hiemstra, P.S.; Tsikas, D.; Romijn, J.A.; et al. Resveratrol protects against atherosclerosis but does not add to the antiatherogenic effect of atorvastatin, in APOE*3-Leiden.CETP mice. J. Nutr. Biochem. 2013, 24, 1423–1430. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Long, J.; Sun, Y.; Chen, W.; Qiu, R.; Yuan, D. Resveratrol ameliorates atherosclerosis induced by high-fat diet and LPS in ApoE−/− mice and inhibits the activation of CD4+ T cells. Nutr. Metab. 2020, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Horseman, M.A.; Surani, S.; Bowman, J.D. Endotoxin, Toll-like Receptor-4, and Atherosclerotic Heart Disease. Curr. Car-Diology Rev. 2017, 13, 86–93. [Google Scholar]
- Brooks, D.; Barr, L.C.; Wiscombe, S.; McAuley, D.F.; Simpson, A.J.; Rostron, A.J. Human lipopolysaccharide models provide mechanistic and therapeutic insights into systemic and pulmonary inflammation. Eur. Respir. J. 2020, 56, 1901298. [Google Scholar] [CrossRef]
- Xu, L.; Wang, R.; Liu, H.; Wang, J.; Mang, J.; Xu, Z. Resveratrol Treatment Is Associated with Lipid Regulation and Inhibition of Lipoprotein-Associated Phospholipase A2 (Lp-PLA2) in Rabbits Fed a High-Fat Diet. Evid. -Based Complement. Altern. Med. 2020, 2020, 1–8. [Google Scholar] [CrossRef]
- Carlquist, J.F.; Muhlestein, J.B.; Anderson, J.L. Lipoprotein-associated phospholipase A2: A new biomarker for cardiovascular risk assessment and potential therapeutic target. Expert Rev. Mol. Diagn. 2007, 7, 511–517. [Google Scholar] [CrossRef]
- Sahebkar, A.; Serban, C.; Ursoniu, S.; Wong, N.D.; Muntner, P.; Graham, I.M.; Mikhailidis, D.P.; Rizzo, M.; Rysz, J.; Sperling, L.S.; et al. Lack of efficacy of resveratrol on C-reactive protein and selected cardiovascular risk factors—Results from a systematic review and meta-analysis of randomized controlled trials. Int. J. Cardiol. 2015, 189, 47–55. [Google Scholar] [CrossRef]
- Tomé-Carneiro, J.; Gonzálvez, M.; Larrosa, M.; Yáñez-Gascón, M.J.; García-Almagro, F.J.; Ruiz-Ros, J.A.; Tomás-Barberán, F.A.; García-Conesa, M.T.; Espín, J.C. Grape Resveratrol Increases Serum Adiponectin and Downregulates Inflammatory Genes in Peripheral Blood Mononuclear Cells: A Triple-Blind, Placebo-Controlled, One-Year Clinical Trial in Patients with Stable Coronary Artery Disease. Cardiovasc. Drugs Ther. 2013, 27, 37–48. [Google Scholar] [CrossRef]
- Batista-Jorge, G.C.; Barcala-Jorge, A.S.; Silveira, M.F.; Lelis, D.F.; Andrade, J.M.O.; de Paula, A.M.B.; Guimarães, A.L.S.; Santos, S.H.S. Oral resveratrol supplementation improves Metabolic Syndrome features in obese patients submitted to a life-style-changing program. Life Sci. 2020, 256, 117962. [Google Scholar] [CrossRef]
- Salazar, H.M.; Mendonça, R.D.D.; Laclaustra, M.; Moreno-Franco, B.; Åkesson, A.; Guallar-Castillón, P.; Donat-Vargas, C. The intake of flavonoids, stilbenes, and tyrosols, mainly consumed through red wine and virgin olive oil, is associated with lower carotid and femoral subclinical atherosclerosis and coronary calcium. Eur. J. Nutr. 2022, 61, 2697–2709. [Google Scholar] [CrossRef]
- Laclaustra, M.; Casasnovas, J.A.; Fernandez-Ortiz, A.; Fuster, V.; Leon-Latre, M.; Jimenez-Borreguero, L.J.; Pocovi, M.; Hur-tado-Roca, Y.; Ordovas, J.M.; Jarauta, E.; et al. Femoral and carotid subclinical atherosclerosis association with risk factors and coronary calcium: The AWHS Study. J. Am. Coll. Cardiol. 2016, 67, 1263–1274. [Google Scholar] [CrossRef] [PubMed]
- Budoff, M.J.; Nasir, K.; McClelland, R.L.; Detrano, R.; Wong, N.; Blumenthal, R.S.; Kondos, G.; Kronmal, R.A. Coronary calcium predicts events better with absolute calcium scores than age-sexrace/ethnicity percentiles: MESA (Multi-Ethnic Study of Ath-erosclerosis). J. Am. Coll. Cardiol. 2009, 53, 345–352. [Google Scholar] [CrossRef]
- Lo-Kioeng-Shioe, M.S.; Rijlaarsdam-Hermsen, D.; van Domburg, R.T.; Hadamitzky, M.; Lima, J.A.; Hoeks, S.E.; Deckers, J.W. Prognostic value of coronary artery calcium score in symptomatic individuals: A meta-analysis of 34,000 subjects. Int. J. Cardiol. 2019, 299, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Tota-Maharaj, R.; Blaha, M.J.; Blankstein, R.; Silverman, M.G.; Eng, J.; Shaw, L.J.; Blumenthal, R.S.; Budoff, M.J.; Nasir, K. Association of coronary artery calcium and coronary heart disease events in young and elderly participants in the multi-ethnic study of atherosclerosis: A secondary analysis of a prospective, population-based cohort. Mayo. Clin. Proc. 2014, 89, 1350–1359. [Google Scholar] [CrossRef] [PubMed]
- Santana, T.M.; Ogawa, L.Y.; Rogero, M.M.; Barroso, L.P.; de Castro, I.A. Effect of resveratrol supplementation on biomarkers associated with atherosclerosis in humans. Complement Clin Pract. 2022, 46, 101491. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Wang, Y.; Han, X.; Fu, S.; Zhu, C.; Chen, Q. Efficacy of resveratrol supplementation on Glucose and Lipid Me-tabolism: A Meta-Analysis and Systematic Review. Front. Physiol. 2022, 13, 795980. [Google Scholar] [CrossRef]
- Mohammadipoor, N.; Shafiee, F.; Rostami, A.; Kahrizi, M.S.; Soleimanpour, H.; Ghodsi, M.; Ansari, M.J.; Bokov, D.O.; Jannat, B.; Mosharkesh, E.; et al. Resveratrol supplementation efficiently improves endothelial health: A systematic review and meta-analysis of randomized controlled trials. Phytother. Res. 2022, 36, 3529–3539. [Google Scholar] [CrossRef]
- Sonnenburg, J.L.; Bäckhed, F. Diet–microbiota interactions as moderators of human metabolism. Nature 2016, 535, 56–64. [Google Scholar] [CrossRef]
- Schoeler, M.; Caesar, R. Dietary lipids, gut microbiota and lipid metabolism. Rev. Endocr. Metab. Disord. 2019, 20, 461–472. [Google Scholar] [CrossRef]
- Jackman, J.A.; Yoon, B.K.; Li, D.; Cho, N.-J. Nanotechnology Formulations for Antibacterial Free Fatty Acids and Monoglycerides. Molecules 2016, 21, 305. [Google Scholar] [CrossRef]
- Shilling, M.; Matt, L.; Rubin, E.; Visitacion, M.P.; Haller, N.A.; Grey, S.F.; Woolverton, C.J. Antimicrobial Effects of Virgin Coconut Oil and Its Medium-Chain Fatty Acids on Clostridium difficile. J. Med. Food 2013, 16, 1079–1085. [Google Scholar] [CrossRef] [PubMed]
- Sheu, C.W.; Freese, E. Effects of fatty acids on growth and envelope proteins of Bacillus subtilis. J. Bacteriol. 1972, 111, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.J.; Yoo, J.-S.; Lee, T.-G.; Cho, H.-Y.; Kim, Y.-H.; Kim, W.-G. Fatty acid synthesis is a target for antibacterial activity of unsaturated fatty acids. FEBS Lett. 2005, 579, 5157–5162. [Google Scholar] [CrossRef] [PubMed]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of l-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Li, Q.; Jiang, H. Gut microbiota in atherosclerosis: Focus on trimethylamine N-oxide. Apmis 2020, 128, 353–366. [Google Scholar] [CrossRef]
- Jonsson, A.L.; Bäckhed, F. Role of gut microbiota in atherosclerosis. Nat. Rev. Cardiol. 2016, 14, 79–87. [Google Scholar] [CrossRef]
- Wang, B.; Qiu, J.; Lian, J.; Yang, X.; Zhou, J. Gut Metabolite Trimethylamine-N-Oxide in Atherosclerosis: From Mechanism to Therapy. Front. Cardiovasc. Med. 2021, 8, 1560. [Google Scholar] [CrossRef]
- Heianza, Y.; Ma, W.; DiDonato, J.A.; Sun, Q.; Rimm, E.B.; Hu, F.B.; Rexrode, K.M.; Manson, J.E.; Qi, L. Long-Term Changes in Gut Microbial Metabolite Trimethylamine N-Oxide and Coronary Heart Disease Risk. J. Am. Coll. Cardiol. 2020, 75, 763–772. [Google Scholar] [CrossRef]
- Gregory, J.C.; Buffa, J.A.; Org, E.; Wang, Z.; Levison, B.S.; Zhu, W.; Wagner, M.A.; Bennett, B.J.; Li, L.; DiDonato, J.A.; et al. Transmission of Atherosclerosis Susceptibility with Gut Microbial Transplantation. J. Biol. Chem. 2015, 290, 5647–5660. [Google Scholar] [CrossRef]
- Haghikia, A.; Li, X.S.; Liman, T.G.; Bledau, N.; Schmidt, D.; Zimmermann, F.; Kränkel, N.; Widera, C.; Sonnenschein, K.; Haghikia, A.; et al. Gut Microbiota–Dependent Trimethylamine N -Oxide Predicts Risk of Cardiovascular Events in Patients with Stroke and Is Related to Proinflammatory Monocytes. Arter. Thromb. Vasc. Biol. 2018, 38, 2225–2235. [Google Scholar] [CrossRef]
- Farhangi, M.A. Gut microbiota-dependent trimethylamine N-oxide and all-cause mortality: Findings from an updated systematic review and meta-analysis. Nutrition 2020, 78, 110856. [Google Scholar] [CrossRef] [PubMed]
- Schiattarella, G.G.; Sannino, A.; Toscano, E.; Giugliano, G.; Gargiulo, G.; Franzone, A.; Trimarco, B.; Esposito, G.; Perrino, C. Gut microbe-generated metabolite trimethylamine-N-oxide as cardiovascular risk biomarker: A systematic review and dose-response meta-analysis. Eur. Hear. J. 2017, 38, 2948–2956. [Google Scholar] [CrossRef] [PubMed]
- Vitaglione, P.; Sforza, S.; Galaverna, G.; Ghidini, C.; Caporaso, N.; Vescovi, P.P.; Fogliano, V.; Marchelli, R. Bioavailability of trans-resveratrol from red wine in humans. Mol. Nutr. Food Res. 2005, 49, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Walle, T. Bioavailability of resveratrol. Ann. NY Acad. Sci. USA 2011, 1215, 9–15. [Google Scholar] [CrossRef]
- Jing, L.; Zhang, H.; Xiang, Q.; Shen, L.; Guo, X.; Zhai, C.; Hu, H. Targeting Trimethylamine N-Oxide: A New Therapeutic Strategy for Alleviating Atherosclerosis. Front. Cardiovasc. Med. 2022, 9, 864600. [Google Scholar] [CrossRef]
- Chen, F.; Wen, Q.; Jiang, J.; Li, H.L.; Tan, Y.F.; Li, Y.H.; Zeng, N.K. Could the gut microbiota reconcile the oral bioavailability conundrum of traditional herbs? J. Ethnopharmacol. 2016, 179, 253–264. [Google Scholar] [CrossRef]
- Komaroff, A.L. The microbiome and risk for obesity and diabetes. JAMA 2017, 317, 355–356. [Google Scholar] [CrossRef]
- Paulo, L.; Ferreira, S.; Gallardo, E.; Queiroz, J.; Domingues, F. Antimicrobial activity and effects of resveratrol on human pathogenic bacteria. World J. Microbiol. Biotechnol. 2010, 26, 1533–1538. [Google Scholar] [CrossRef]
- Nawrocki, E.M.; Bedell, H.W.; Humphreys, T.L. Resveratrol is cidal to both classes of Haemophilus ducreyi. Int. J. Antimicrob. Agents 2013, 41, 477–479. [Google Scholar] [CrossRef]
- Chen, M.-L.; Yi, L.; Zhang, Y.; Zhou, X.; Ran, L.; Yang, J.; Zhu, J.-D.; Zhang, Q.-Y.; Mi, M.-T. Resveratrol Attenuates Trimethylamine- N -Oxide (TMAO)-Induced Atherosclerosis by Regulating TMAO Synthesis and Bile Acid Metabolism via Remodeling of the Gut Microbiota. mBio 2016, 7, e02210-15. [Google Scholar] [CrossRef]
- Salehi, B.; Mishra, A.P.; Nigam, M.; Sener, B.; Kilic, M.; Sharifi-Rad, M.; Fokou, P.V.T.; Martins, N.; Sharifi-Rad, J. Resveratrol: A Double-Edged Sword in Health Benefits. Biomedicines 2018, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- Bonnefont-Rousselot, D. Resveratrol and Cardiovascular Diseases. Nutrients 2016, 8, 250. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-F.; Chen, S.-A.; Wu, S.-N. Evidence for the stimulatory effect of resveratrol on Ca(2+)-activated K+ current in vascular endothelial cells. Cardiovasc. Res. 2000, 45, 1035–1045. [Google Scholar] [CrossRef] [PubMed]
- Orallo, F.; Álvarez, E.; Camiña, M.; Leiro, J.M.; Gómez, E.; Fernández, P. The Possible Implication of trans-Resveratrol in the Cardioprotective Effects of Long-Term Moderate Wine Consumption. Mol. Pharmacol. 2002, 61, 294–302. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chemical structure of resveratrol (drawn using ChemSketch, ACDLabs).


{kind=link}
Table 1.
RSV effects on the antioxidant activity, lipid metabolism, and progression of atherosclerosis: studies on animal models.
Table 1.
RSV effects on the antioxidant activity, lipid metabolism, and progression of atherosclerosis: studies on animal models.
| Author, Year of Publication | Study Design | Study Duration | Sample Size | RSV Supplementation, Antioxidant Activity, Lipid Profile, and Atherosclerotic Risk |
|---|---|---|---|---|
| Maura Floreani, 2003 [39] | Case control | Follow-up: 16 days | 13 Dunkin–Hartley male guinea pigs | Effect of RSV on Vmax and Km of DT-diaphorase from guinea pig cardiac tissue: RSV treatment vs. controls Vmax: 75.47 ± 3.87 vs. 57.08 ± 9.02, p < 0.05 Km: 0.23 ± 0.03 vs. 0.25 ± 0.02, p = NS Effect of RSV on the antioxidant enzyme activities of cardiac tissue: RSV treatment vs. controls Gpx: 46.49 ± 2.61 vs. 45.10 ± 1.60, p = NS GR: 18.21 ± 2.97 vs. 16.62 ± 1.93, p = NS CAT: 32.20 ± 2.39 vs. 25.14 ± 3.85, p < 0.001 Cu/Zn SOD: 32.69 ± 3.86 vs. 27.75 ± 4.15, p = NS Mn–SOD: 31.87 ± 3.55 vs. 32.70 ± 1.86, p = NS Menadione-induced ROS generation by cardiac tissue from the control and resveratrol-treated guinea pigs. RSV-treated guinea pigs: significantly lower ROS production with respect to controls (p < 0.001, data not shown) Positive inotropic effect induced by increasing the concentrations of menadione in the left atria isolated from control and RSV-treated guinea pigs. RSV-treated guinea pigs: no inotropic action of menadione at 5 M concentration; effect was 54% lower than that observed in controls (p < 0.001, data not shown) |
| Gyeong-Min Do, 2008 [40] | Case control | 20 weeks | 4-week-old male apo/E-deficient mice | Effects of resveratrol supplementation on the plasma levels (mmol/L) of the total cholesterol at 20 weeks (p < 0.05): Control diet: 9.60 ± 0.50 0.02% clofibrate-supplemented diet: 7.73 ± 0.36 0.02% resveratrol-supplemented diet: 6.33 ± 0.41 0.06% resveratrol-supplemented diet: 7.78 ± 0.51 Effects of resveratrol supplementation on the plasma levels (mmol/L) of Triglycerides at 20 weeks (p < 0.05): Control diet: 1.51 ± 0.17 0.02% clofibrate-supplemented diet: 1.10 ± 0.11 0.02% resveratrol-supplemented diet: 1.00 ± 0.11 0.06% resveratrol-supplemented diet: 1.42 ± 0.16 Effects of resveratrol supplementation on the plasma levels (mmol/L) of the LDL at 20 weeks (p < 0.05): Control diet: 8.1 ± 0.5 0.02% clofibrate-supplemented diet: 6.1 ± 0.4 0.02% resveratrol-supplemented diet: 4.8 ± 0.3 0.06% resveratrol-supplemented diet: 5.9 ± 0.5 Effects of resveratrol supplementation on the plasma levels (mmol/L) of the HDL at 20 weeks (p < 0.05): Control diet: 1.07 ± 0.10 0.02% clofibrate-supplemented diet: 1.41 ± 0.12 0.02% resveratrol-supplemented diet: 1.36 ± 0.11 0.06% resveratrol-supplemented diet: 1.87 ± 0.12 Effects of resveratrol supplementation on the plasma levels of the HDL/cholesterol (%) at 20 weeks (p < 0.05): Control diet: 11.7 ± 0.9 0.02% clofibrate-supplemented diet: 18.4 ± 1.5 0.02% resveratrol-supplemented diet: 20.9 ± 1.7 0.06% resveratrol-supplemented diet: 22.3 ± 1.5 Effects of resveratrol supplementation on the plasma levels of the atherogenic index (ratio) at 20 weeks (p < 0.05): Control diet: 7.0 ± 0.5 0.02% clofibrate-supplemented diet: 4.6 ± 0.4 0.02% resveratrol-supplemented diet: 3.9 ± 0.4 0.06% resveratrol-supplemented diet: 3.8 ± 0.3 Effects of resveratrol supplementation on the hepatic HMG-CoA reductase activity (pmol/min/mg protein) at 20 weeks (p < 0.05): Control diet: 282.15 ± 15.62 0.02% clofibrate-supplemented diet: 148.20 ± 31.38 0.02% resveratrol-supplemented diet: 113.73 ± 6.20 0.06% resveratrol-supplemented diet: 152.97 ± 11.45 Effects of resveratrol supplementation on the hepatic ACAT activity (pmol/min/mg protein) at 20 weeks: Control diet: 49.99 ± 4.17 0.02% clofibrate-supplemented diet: 52.81 ± 2.86 0.02% resveratrol-supplemented diet: 55.92 ± 2.78 0.06% resveratrol-supplemented diet: 59.77 ± 3.43 |
| Jimmy F.P. Berbée, 2013 [41] | Case control | Run-in period: 5 weeks; Drug intervention period: 14 weeks | 10–12-week-old female APOE*3- Leiden.CETP (E3L.CETP) transgenic mice | Total atherosclerotic lesion area: Resveratrol vs. control: −52% (p < 0.01) Atorvastatin vs. control: −40% (p < 0.05) Resveratrol plus atorvastatin vs. control: −47% (p < 0.01) Collagen/macrophage ratio as marker of plaque stability: Resveratrol vs. control: +108% (p < 0.05) Atorvastatin vs. control: +124% (p < 0.01) Resveratrol plus atorvastatin vs. control: +154% (p < 0.001) Reduction in the plasma VLDL levels: Resveratrol vs. control: −19% (p = 0.08) Atorvastatin vs. control: −22% (p < 0.05) Resveratrol plus atorvastatin vs. control: −21% (p < 0.05) |
| Liyu Zhou, 2020 [42] | Case control | 20 weeks | 7-week-old male apo/E-deficient mice | HFD + LPS vs. control after 20 weeks (data not shown): TC, LDL-C, non-HDL-C serum levels: increased (p < 0.001). TG serum levels: increased (p < 0.05) HDL-C serum levels decreased (p < 0.05) Atherosclerotic plaque area ratio: increased (p < 0.01) CD4+ T cells in PBMC: increased (p < 0.01) Simvastatin + LPS vs. HFD + LPS after 20 weeks (data not shown): TC, LDL-C, serum levels: decreased (p < 0.01) TG, non-HDL-C serum levels: decreased (p < 0.05) HDL-C serum levels increased (p < 0.05) Atherosclerotic plaque area ratio: decreased (p < 0.05) CD4+ T cells in PBMC: decreased (p < 0.05) Resveratrol + LPS vs. HFD + LPS after 20 weeks (data not shown): TC, LDL-C, serum levels: decreased (p < 0.01) TG, non-HDL-C serum levels: decreased (p < 0.05) HDL-C serum levels increased (p < 0.05) Atherosclerotic plaque area ratio: decreased (p < 0.05) CD4+ T cells in PBMC: decreased (p < 0.05) |
| Lei Xu, 2020 [45] | Case control | 3 months | 24 three-month-old New Zealand male white rabbits | Serum levels (mmol/L) of the total cholesterol at 3 months in RFG vs. FDG 11.84 ± 2.78 vs. 30.32 ± 5.74, p < 0.001 Serum levels (mmol/L) of the HDL at 3 months in RFG vs. FDG 1.58 ± 0.35 vs. 3.28 ± 1.61, p < 0.001 Serum levels (mmol/L) of the LDL at 3 months in RFG vs. FDG 6.23 ± 1.53 vs. 16.45 ± 3.16, p < 0.001 Serum levels (mmol/L) of Lp-PLA2 at 3 months in RFG vs. FDG 953.20 ± 96.66 vs. 1928.88 ± 385.78, p < 0.001 Serum levels (mmol/L) of the TG at 3 months in RFG vs. FDG 0.76 ± 0.28 vs. 0.90 ± 0.42, p = 0.232 Measurements (μm) of the aortic arch thickness observed under a microscope at 3 months in RFG vs. FDG Intima: 52.44 ± 14.94 vs. 124.76 ± 6.83, p < 0.001 Intima/media ratio: 0.30 ± 0.09 vs. 0.64 ± 0.04, p < 0.001 Smooth muscle layer: 173.48 ± 4.05 vs. 194.16 ± 10.2, p = 0.65 |
Table 2.
RSV effects on the antioxidant activity, lipid metabolism, and progression of atherosclerosis: data from randomized clinical trials and prospective cohort studies.
Table 2.
RSV effects on the antioxidant activity, lipid metabolism, and progression of atherosclerosis: data from randomized clinical trials and prospective cohort studies.
| Author, Year of Publication | Study Design | Duration of Study | Sample Size | RSV Intake, Atherosclerosis, and Inflammatory Biomarkers |
|---|---|---|---|---|
| Amirhossein Sahebkar, 2015 [47] | Meta-analysis from 10 randomized controlled trials | Treatment duration range: 60 days to 12 months | 600 subjects: 344 in RSV group and 256 controls | Effect of RSV supplementation on the plasma concentration of the CRP and selected predictors of CVD risk. CRP (mg/L): WMD = −0.144; 95% CI = −0.968–0.680; p = 0.731 TC (mg/dL): WMD = 1.49; 95% CI = −14.96–17.93; p = 0.859 LDL (mg/dL): WMD = −0.31; 95% CI = −9.57–8.95; p = 0.948 HDL (mg/dL): WMD = −4.18; 95% CI = −6.54–1.82; p = 0.001 TG (mg/dL): WMD = 2.67; 95% CI = −28.34–33.67; p = 0.866 Glucose (mg/dL): WMD = 1.28; 95% CI = −5.28–7.84; p = 0.703 SBP (mm Hg): WMD = 0.82; 95% CI = −8.86–10.50; p = 0.868 DBP (mm Hg): WMD = 1.72; 95% CI = −6.29–9.73; p = 0.674 |
| João Tomé-Carneiro, 2013 [48] | Triple-blind, randomized, placebo-controlled study | Follow-up: 1 year | 75 stable-CAD patients | Serum levels of the inflammatory markers after 12 months vs. baseline in placebo group: Interleukin-10 (pg/mL): 18.3 ± 12.2 vs. 19.6 ± 12.4; p = 0.03 Adiponectin (μg/mL): 9.6 ± 4.4 vs. 11.0 ± 5.8; p = 0.01 PAI–1 (ng/mL): 25.9 ± 15.0 vs. 18.7 ± 14.4; p < 0.001 hs-CRP (mg/L): 4.0 ± 1.8 vs. 3.3 ± 2.2; p = 0.66 Serum levels of the inflammatory markers after 12 months vs. baseline in RSV group: Interleukin-10 (pg/mL): 23.3 ± 16.9 vs. 23.05 ± 16.7; p = 0.45 Adiponectin (μg/mL): 13.6 ± 5.2 vs. 12.4 ± 5.6; p = 0.01 PAI–1 (ng/mL): 14.0 ± 7.0 vs. 17.2 ± 10.3; p = 0.05 hs-CRP (mg/L): 3.2 ± 2.1 vs. 3.9 ± 4.1; p = 0.17 Activation status of the inflammation-related transcription factors genes in the PBMCs in the RSV group after 12 months: KLF2: Activated Ap-1: Inhibited JUN: Inhibited ATF-2: Inhibited CREBBP: Inhibited |
| G.C. Batista-Jorge, 2020 [49] | Prospective case–control study | Follow-up: 12 weeks | 25 subjects aged 30 to 60 years old | Anthropometric and biochemical parameters of the subjects treated with the physical activity program + diet + placebo before and after the 12-weeks intervention Bodyweight (kg): 102.7 ± 12.9 vs. 95.9 ± 12.1, p = 0.006 BMI (kg/m2): 35.0 ± 3.0 vs. 32.8 ± 3.6, p = 0.003 WC (cm): 105.0 ± 9.4 vs. 96.3 ± 8.6, p = 0.001 TC (mg/dL): 198.4 ± 33.1 vs. 193.7 ± 44.1, p = 0.568 LDL (mg/dL): 123.3 ± 38.3 vs. 117.7 ± 42.0, p = 0.483 HDL (mg/dL): 47.2 ± 6.6 vs. 50.6 ± 5.8, p = 0.253 VLDL (mg/dL): 24.1 ± 6.7 vs. 27.4 ± 9.5, p = 0.209 Triglycerides (mg/dL): 120.4 ± 33.1 vs. 137.1 ± 47.4, p = 0.203 Insulin (IU): 56.7 ± 91.7 vs. 13.9 ± 6.1, p = 0.194 Glycemia (mg/dL): 87.1 ± 9.1 vs. 89.4 ± 5.7, p = 0.363 HBA1c (%): 5.5 ± 0.7 vs. 6.2 ± 0.4, p = 0.005 Leptin (mg/dL): 36.3 ± 24.2 vs. 22.7 ± 16.2, p = 0.031 Uric acid (mg/dL): 10.6 ± 15.9 vs. 4.8 ± 0.7, p = 0.345 ALT (mg/dL): 30.2 ± 14.4 vs. 30.6 ± 14.5, p = 0.948 AST (mg/dL): 33.3 ± 19.4 vs. 35.3 ± 34.1, p = 0.874 Fibrinogen (mg/dL): 97.0 ± 136.9 vs. 5.1 ± 5.7, p = 0.535 Urea (mg/dL): 30.0 ± 12.2 vs. 30.3 ± 6.5, p = 0.933 Creatinine (mg/dL): 0.9 ± 0.3 vs. 1.6 ± 2.3, p = 0.344 Albumin (mg/dL): 4.5 ± 0.4 vs. 7.8 ± 11.3, p = 0.371 Anthropometric and biochemical parameters of the subjects treated with the physical activity program + diet + RSV (250 mg/day) before and after the 12-week intervention Bodyweight (kg): 105.4 ± 13.0 vs. 98.1 ± 11.7, p = 0.000 BMI (kg/m2): 36.1 ± 4.5 vs. 33.6 ± 3.5, p = 0.000 WC (cm): 109.6 ± 8.3 vs. 102.2 ± 7.8, p = 0.000 TC (mg/dL): 221.0 ± 48.6 vs. 192.1 ± 43.9, p = 0.031 LDL (mg/dL): 127.8 ± 33.3 vs. 108.1 ± 38.1, p = 0.241 HDL (mg/dL): 42.7 ± 7.6 vs. 48.1 ± 6.2, p = 0.026 VLDL (mg/dL): 49.3 ± 25.6 vs. 39.1 ± 14.3, p = 0.025 Triglycerides (mg/dL): 294.9 ± 191.8 vs. 189.9 ± 73.2, p = 0.094 Insulin (IU): 46.6 ± 69.9 vs. 15.6 ± 4.1, p = 0.154 Glycemia (mg/dL): 138.8 ± 66.3 vs. 117.1 ± 45.9, p = 0.116 HBA1c (%): 11.3 ± 15.0 vs. 11.7 ± 15.6, p = 0.343 Leptin (mg/dL): 35.8 ± 27.9 vs. 28.0 ± 25.5, p = 0.014 Uric acid (mg/dL): 10.1 ± 14.8 vs. 5.4 ± 1.6, p = 0.302 ALT (mg/dL): 34.9 ± 12.4 vs. 29.3 ± 11.0, p = 0.099 AST (mg/dL): 46.1 ± 27.4 vs. 37.7 ± 23.5, p = 0.440 Fibrinogen (mg/dL): 82.1 ± 151.9 vs. 3.7 ± 1.2 p = 0.199 Urea (mg/dL): 35.9 ± 8.8 vs. 28.9 ± 6.3, p = 0.046 Creatinine (mg/dL): 1.0 ± 0.1 vs. 0.9 ± 0.2, p = 0.021 Albumin (mg/dL): 4.8 ± 0.6 vs. 4.0 ± 0.4, p = 0.000 |
| Henry Montero Salazar, 2022 [50] | Prospective cohort study | Mean follow-up: 10 years | 2318 subjects from cohort of AWHS | Risk of presence of at least one plaque in the femoralarteries according to a higher dietary polyphenol intake: Stilbenes: OR = 0.62; 95%CI = 0.46–0.83; p trend = 0.009 Flavonoids: OR = 0.62; 95%CI = 0.48–0.80; p trend < 0.001 Risk of presence of at least one plaque in the carotid arteries according to a higher dietary polyphenols intake: Stilbenes: OR = 0.89; 95%CI = 0.67–1.18; P trend = 0.536 Flavonoids: OR = 0.80; 95%CI = 0.62–1.02; P trend = 0.094 Risk of presence of at least one plaque in the femoral arteries according to each 1 mg/day intake of stilbenes or each 100 mg/daily intake of flavonoids: Stilbenes: OR = 0.97; 95%CI = 0.95–0.99 Flavonoids: OR = 0.94; 95%CI = 0.90–0.98 Risk of presence of a positive coronary calcium Agatston score (CACS > 0) according to a higher dietary polyphenol intake: Stilbenes: OR = 0.75; 95%CI = 0.55–1.03; P trend = 0.131 Flavonoids: OR = 0.88; 95%CI = 0.67–1.15; P trend = 0.346 |
| Tamires Miranda Santana, 2022 [55] | Review from 27 randomized clinical trials | Treatment duration range: 28–360 days | Average sample size: 26 subjects | Changes in atherosclerosis biomarkers according to Cluster II (RSV dose: 454.14 ± 129 mg): SBP (%): −6.76 ± 1.06, p < 0.001 DBP (%):−4.51 ± 1.77, p = 0.026 TC (%): −6.04 ± 1.51, p < 0.001 TG (%): −10.13 ± 5.01, p < 0.001 LDL (%): −4.81 ± 3.40, p = 0.036 HDL (%): −1.16 ± 1.42, p < 0.001 Changes in atherosclerosis biomarkers according to Cluster III (RSV dose: 263.75 ± 88.07 mg): SBP (%): −0.95 ± 1.19, p < 0.001 DBP (%): −1.39 ± 0.87, p = 0.026 TC (%): −3.28 ± 1.39, p < 0.001 TG (%): −3.51 ± 2.12, p < 0.001 LDL (%): −4.35 ± 2.38, p = 0.036 HDL (%): 6.23 ± 1.35, p < 0.001 |
| Qian Zhou, 2022 [56] | Meta-analysis and systematic review from 25 randomized controlled trials | Treatment duration range: 4–26 weeks | 1171 subjects: 578 in placebo group and 593 in RSV intervention group | Effect of RSV supplementation on the lipid, glucose, and anthropometric parameters (SMD). Best duration of intervention: more than 17 weeks; best intake of RSV: 200–500 mg/day) TC: SMD = –0.15; 95% CI = –0.01–−0.3; p = 0.003 LDL-C: SMD = –0.42; 95% CI = –0.27–−0.57; p < 0.001 HDL-C: SMD = −0.16 95% CI = –0.02–−0.31; p = 0.03 HbA1c: SMD = –0.48, 95% CI = –0.2–−0.69; p < 0.001 WC: SMD = –0.36; 95% CI = –0.14–−0.59; p = 0.002 TG: SMD = 0.06; 95%CI = –0.12–0.23; p = 0.53 Body weight: SMD = 0.12; 95%CI = –0.05–0.28; p = 0.18 BMI: SMD = 0.02; 95%CI = –0.13–0.17; p = 0.83 HOMA index: SMD = 0.14; 95%CI = 0.04–0.31; p = 0.13 Insulin: SMD = 0.1; 95%CI = –0.06–0.25; p = 0.42 Leptin: SMD = 0.11; 95%CI = –0.13–0.36; p = 0.38 Fasting glucose: SMD = –0.10; 95%CI = –0.24–0.03; p = 0.14 Fat percentage: SMD = –0.26; 95%CI = –0.54–0.01; p = 0.06 Adiponectin: SMD = 0.02; 95%CI = –0.22–0.25; p = 0.89 |
| Nazanin Mohammadipoor, 2022 [57] | Meta-analysis and systematic review from 17 randomized controlled trials | Treatment duration: 90 min–24 weeks for evaluating FMD; 4–54 weeks for evaluating fibrinogen, PAI-1, ICAM-1, VCAM-1 | Effect of RSV supplementation on FMD (%), ICAM-1, VCAM-1, fibrinogen, PAI-1 (WMD). FMD in cross-over studies: WMD = 1.59%; 95% CI: 1.05–2.13, p < 0.001 FMD at dosages <100 mg: WMD = 1.69%; 95% CI:1.30–2.08, p < 0.001 FMD on acute intervention: WMD = 2.28%; 95% CI: 1.80–2.76, p < 0.001 FMD on patients with CVD: WMD = 2.60%; 95% CI: 1.95–3.26, p < 0.001 ICAM-1: WMD = −7.09 ng/mL, 95% CI: −7.45–−6.73, p < 0.001 VCAM-1: WMD = −15.04 ng/mL, 95% CI: −37.52–−7.45, p = 0.19 Fibrinogen: WMD = 0.25 g/L, 95% CI: −0.36–0.86, p = 0.42 PAI-1: WMD = −0.94 ng/mL, 95% CI: −3.51–1.62, p = 0.47 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Capurso, C.; Bellanti, F.; Lo Buglio, A.; Vendemiale, G. Cardioprotective Effects of Resveratrol in the Mediterranean Diet: A Short Narrative Review. Dietetics 2023, 2, 174-190. https://doi.org/10.3390/dietetics2020014
AMA Style
Capurso C, Bellanti F, Lo Buglio A, Vendemiale G. Cardioprotective Effects of Resveratrol in the Mediterranean Diet: A Short Narrative Review. Dietetics. 2023; 2(2):174-190. https://doi.org/10.3390/dietetics2020014
Chicago/Turabian StyleCapurso, Cristiano, Francesco Bellanti, Aurelio Lo Buglio, and Gianluigi Vendemiale. 2023. "Cardioprotective Effects of Resveratrol in the Mediterranean Diet: A Short Narrative Review" Dietetics 2, no. 2: 174-190. https://doi.org/10.3390/dietetics2020014