
Population Status of the Tropical Freshwater Shrimp Xiphocaris elongata in Urban and Forest Streams in Puerto Rico

Abstract
:1. Introduction
2. Materials and Methods
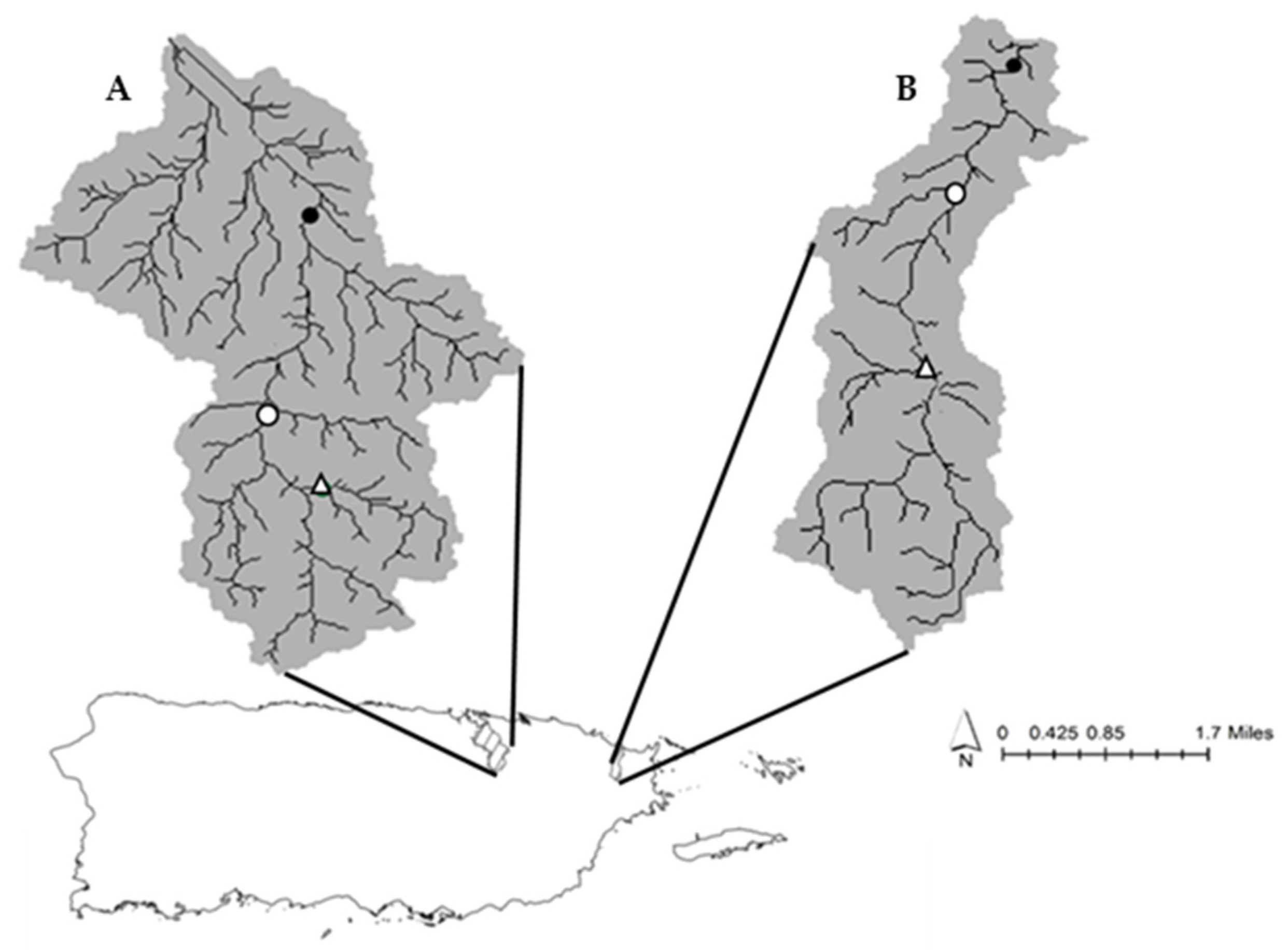
2.1. Study Sites
2.2. Physicochemical Variables
2.3. Freshwater Shrimp Sampling
2.4. Statistical Analyses
3. Results
3.1. Physicochemical Variables
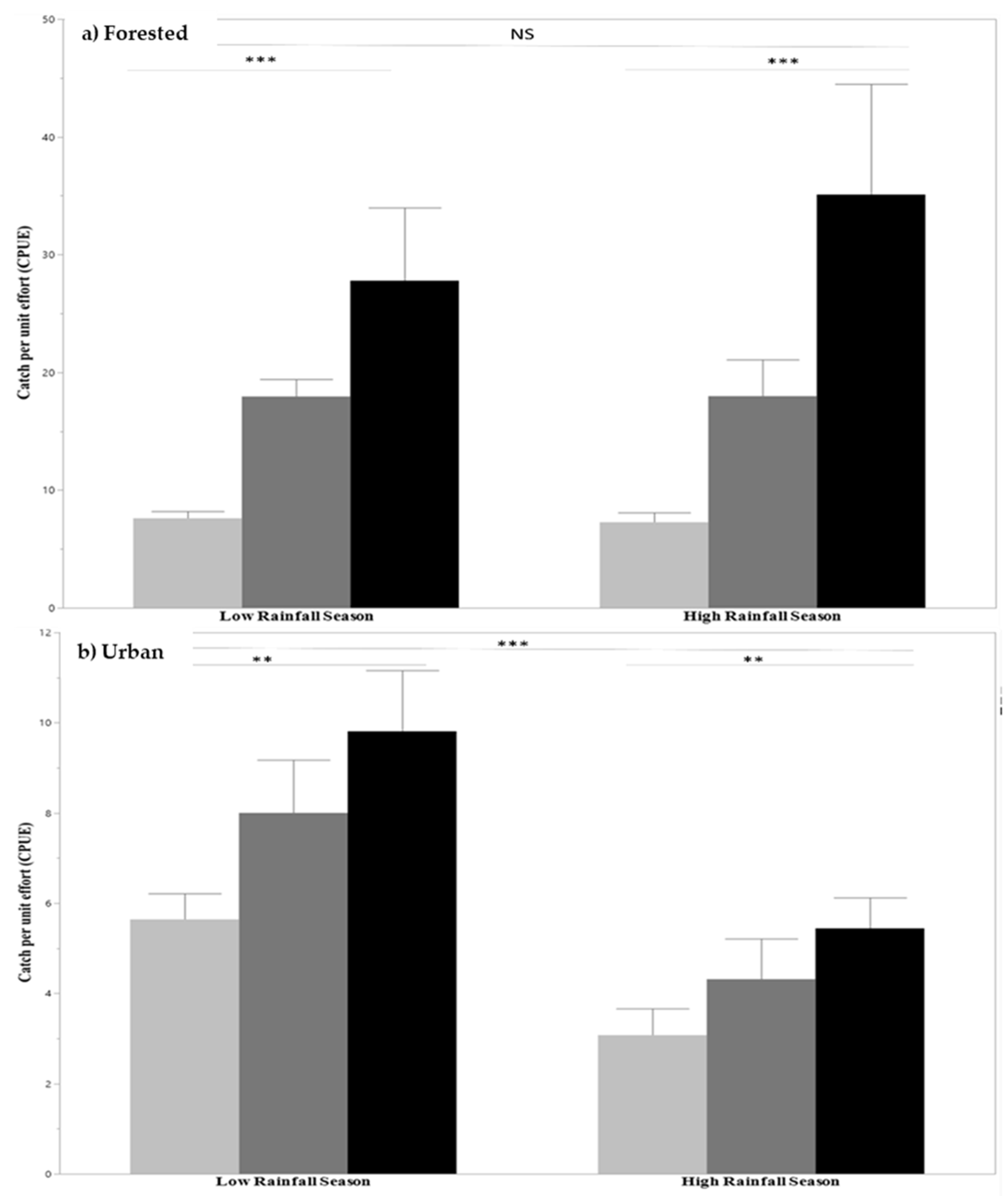
3.2. Freshwater Shrimp Sampling
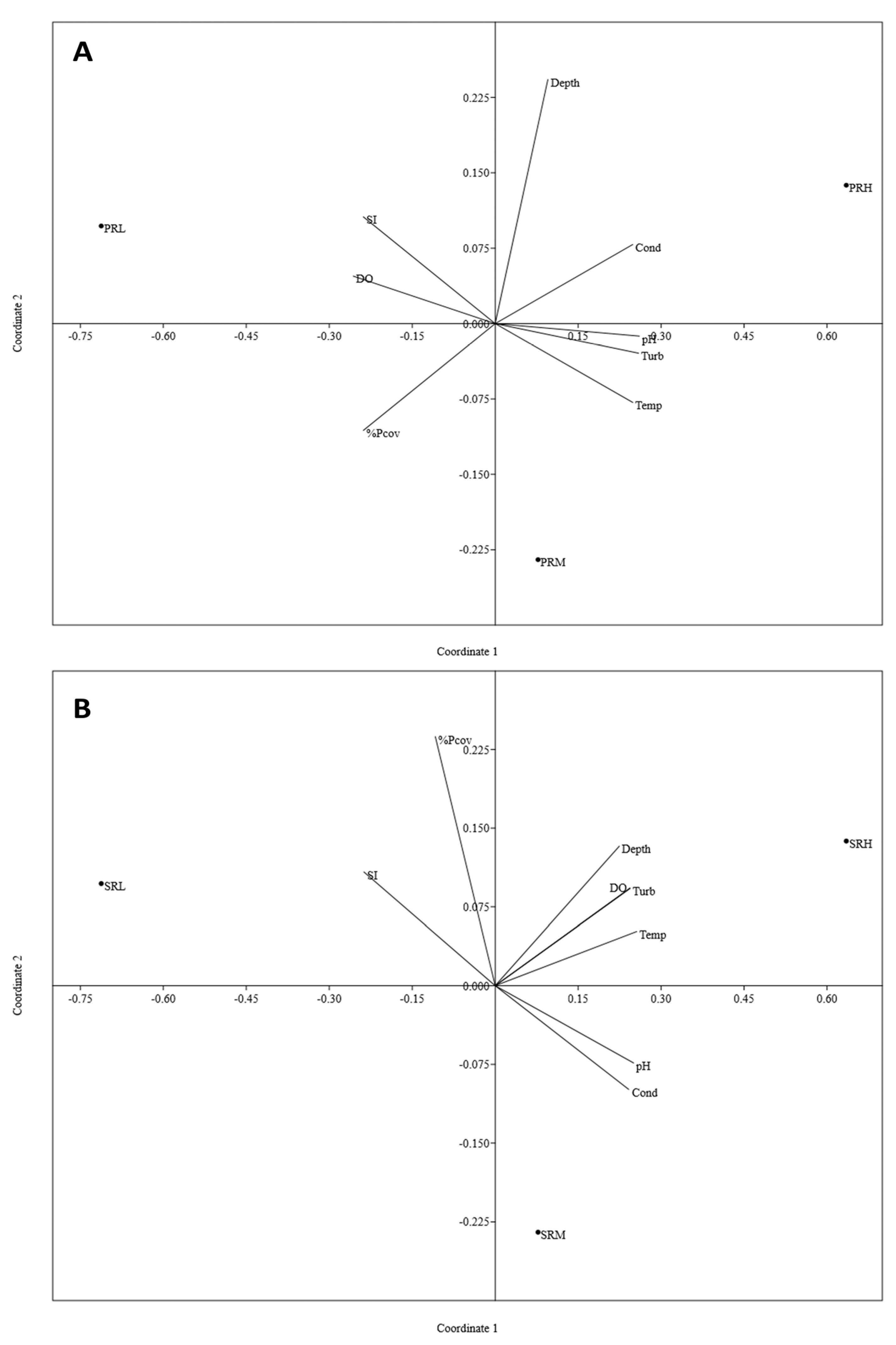
3.3. Multivariable Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Watch Institute. State of the World: Our Urban; W. W. Norton & Company: Washington, DC, USA, 2007. [Google Scholar]
- Walsh, C.J.; Roy, A.H.; Feminella, J.W.; Cottingham, P.D.; Groffman, P.M.; Morgan, R.P., II. The Urban Stream Syndrome: Current Knowledge and the Search for a Cure. J. North Am. Benthol. Soc. 2005, 24, 706–723. [Google Scholar] [CrossRef]
- Arnold, C.L.; Gibbons, J.C. Impervious surface coverage: The emergence of a key environmental indicator. J. Am. Plan. Assoc. 1996, 62, 243–258. [Google Scholar] [CrossRef]
- Paul, M.J.; Meyer, E. Streams in Urban Landscape. Annu. Rev. Ecol. Syst. 2001, 32, 333–365. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Stanley, E.H.; Vander Zanden, M.J. State of the world’s freshwater ecosystems: Physical, chemical, and biological changes. Annu. Rev. Environ. Resour. 2011, 36, 75–99. [Google Scholar] [CrossRef] [Green Version]
- Carpenter, S.R.; Caraco, N.F.; Correll, D.L.; Howarth, R.W.; Sharpley, A.N.; Smith, V.H. Nonpoint pollution of surface waters with phosphorus and nitrogen. Ecol. Appl. 1998, 8, 559–568. [Google Scholar] [CrossRef]
- Pérez-Reyes, O.; Crowl, T.; Covich, A. Comparison of decapod communities across an urban-forest land use gradient in Puerto Rican streams. Urban Ecosyst. 2016, 19, 181–203. [Google Scholar] [CrossRef]
- Covich, A.P.; Crowl, T.A.; Scatena, F.N. Effects of extreme low flows on freshwater shrimps in a perennial tropical stream. Freshw. Biol. 2003, 48, 1199–1206. [Google Scholar] [CrossRef]
- Covich, A.P.; Crowl, T.A.; Heartsill-Scalley, T. Effect of drought and hurricane disturbances on headwater distributions of palemonid river shrimp (Macrobrachium spp.) in the Luquillo Mountains, Puerto Rico. J. North Am. Benthol. Soc. 2006, 25, 99–107. [Google Scholar] [CrossRef]
- Covich, A.P.; Crowl, T.A.; Hein, C.L.; Townsend, M.J.; McDowell, W.H. Predator– prey interactions in river networks: Comparing shrimp spatial refugia in two drainage basins. Freshw. Biol. 2009, 54, 450–465. [Google Scholar] [CrossRef]
- Pérez-Reyes, O.; Crowl, T.A.; Hernández-García, P.; Ledesma-Fuste, R.; Villar-Fornes, F.A.; Covich, A.P. Freshwater decapods of Puerto Rico: A checklist and reports of new localities. Zootaxa 2013, 3717, 329–334. [Google Scholar] [CrossRef] [Green Version]
- Kikkert, D.A.; Crowl, T.A.; Covich, A.P. Upstream migration of amphidromous shrimps in the Luquillo Experimental Forest, Puerto Rico: Temporal patterns and environmental cues. J. North Am. Benthol. Soc. 2009, 28, 233–246. [Google Scholar] [CrossRef]
- Bauer, R.T. Amphidromy migrations of freshwater shrimps, I.I. Delivery of hatching larvae to the sea, return juvenile upstream migration, and human impacts. In New Frontiers in Crustacean Biology; Brill: Leiden, The Netherlands, 2011; pp. 157–168. [Google Scholar] [CrossRef] [Green Version]
- Bauer, R.T. Amphidromy in shrimps: A life cycle between rivers and the sea. Lat. Am. J. Aquat. Res. 2013, 4, 633–650. [Google Scholar] [CrossRef]
- Guérin-Méneville, F.E. Crustáceos. In Historia Física, Político y Natural de la isla de Cuba; De la Sagra, R., Ed.; Arthus Bertrand: Paris, French, 1856. [Google Scholar]
- Covich, A.P.; McDowell, W.H. The Stream Community: The Food Web of a Tropical Rain Forest; University of Chicago Press: Chicago, IL, USA, 1996. [Google Scholar]
- Gutiérrez-Fonseca, P.E.; Ramírez, A.; Pringle, C.M.; Torres, P.J.; McDowell, W.H.; Covich, A.; Crowl, T.; Pérez-Reyes, O. When the rainforest dries: Drought effects on a montane tropical stream ecosystem in Puerto Rico. Freshw. Sci. 2020, 39, 197–212. [Google Scholar] [CrossRef]
- Rosas, K.G.; Colón-Gaud, C.; Ramírez, A. Trophic basis of production in tropical headwater streams, Puerto Rico: An assessment of the importance of allochthonous resources in fueling food webs. Hydrobiologia 2020, 847, 1961–1975. [Google Scholar] [CrossRef]
- US Census Bureau. 2010 Census of Population and Housing, Summary Population and Housing Characteristics, CPH-1-53; Puerto Rico U.S. Government Printing Office: Washington, DC, USA, 2012.
- Lugo, A.E.; Ramos, O.M.; Rodríguez, C.R. The Río Piedras Watershed and Its Surrounding Environment; International Institute of Tropical Forestry: San Juan, PR, USA, 2011.
- Ramírez, A.; Rosas, K.G.; Lugo, A.E.; Ramos-González, O.M. Spatio-temporal variation in stream water chemistry in a tropical urban watershed. Ecol. Soc. 2014, 19, 45. [Google Scholar] [CrossRef] [Green Version]
- De Jesús-Crespo, R.; Ramírez, A. Effects of Urbanization on Stream Physicochemistry and Macroinvertebrate Assemblages in a Tropical Urban Watershed in Puerto Rico. J. North Am. Benthol. Soc. 2011, 30, 739–750. [Google Scholar] [CrossRef]
- Ramírez, A.; De Jesús-Crespo, R.; Martinó-Cardona, D.M.; Martínez-Rivera, N.; Burgos-Caraballo, S. Urban Streams in Puerto Rico: What can we learn from the tropics? J. N. Am. Benthol. Soc. 2009, 28, 1070–1079. [Google Scholar] [CrossRef]
- Ramos-González, O.M. The green areas of San Juan, Puerto Rico. Ecol. Soc. 2014, 19, 21. [Google Scholar] [CrossRef] [Green Version]
- Heartsill-Scalley, T.; López-Marrero, T. Land-cover composition, water resources and land management in the watersheds of the Luquillo Mountains, northeastern Puerto Rico. Caribb. Geogr. 2014, 19, 43–68. [Google Scholar]
- Thomlinson, J.R.; Rivera, L.Y. Suburban growth in Luquillo, Puerto Rico: Some consequences of development on natural and semi-natural systems. Landsc. Urban Plan. 2000, 49, 15–23. [Google Scholar] [CrossRef]
- Pottier, G.; Bargier, N.; Bennevault, Y.; Vigouroux, R.; Azam, D.; Marchand, F.; Nevoux, M.; Roussel, J.M. Optimising electrofishing settings for shrimp and fish in shallow tropical streams. Fish. Res. 2022, 256, 106457. [Google Scholar] [CrossRef]
- Ocasio-Torres, M.E.; Giray, T.; Crowl, T.A.; Sabat, A.M. Antipredator defence mechanism in the amphidromous shrimp Xiphocaris elongata (Decapoda: Xiphocarididae): Rostrum length. J. Nat. Hist. 2014, 49, 1493–1506. [Google Scholar] [CrossRef]
- Urban, M.C.; Skelly, D.K.; Burchsted, D.; Price, W.; Lowry, S. Stream communities across a rural-urban landscape gradient. Divers. Distrib. 2006, 12, 337–350. [Google Scholar] [CrossRef]
- Brasher, A.M.D. Impacts of Human Disturbances on Biotic Communities in Hawaiian Streams. BioScience 2003, 53, 1052–1059. [Google Scholar] [CrossRef] [Green Version]
- Colin, P.L. L’influence des Différents Cortèges Floristiques de la Ripisylve sur la Composition des Communautés Aquatiques (Crustacés et Poissons). Master’s Thesis, Université Des Antilles, Fouillole, French, 2020. [Google Scholar]
- Dugdale, S.J.; Malcolm, I.A.; Kantola, K.; Hannah, D.M. Stream temperature under contrasting riparian forest cover: Understanding thermal dynamics and heat exchange processes. Sci. Total Environ. 2018, 610–611, 1375–1389. [Google Scholar] [CrossRef]
- Kaushal, S.S.; Belt, K.T. The urban watershed continuum: Evolving spatial and temporal dimensions. Urban Ecosyst. 2012, 15, 409–435. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Watershed | Total Urban Population along Reach Areas (Inhabitants) | ||
|---|---|---|---|
| LU | MU | HU | |
| Río Sabana | 877 | 1821 | 5327 |
| Río Piedras | 985 | 2515 | 30,118 |
| Watershed | Physicochemical Variables | LU | MU | HU | ANOVA |
|---|---|---|---|---|---|
| Urban | Temperature (°C) | 22.4 ± 0.04 | 23.8 ± 0.03 | 23.9 ± 0.03 | *** |
| pH | 7.9 ± 0.03 | 7.9 ± 0.02 | 8.0 ± 0.03 | *** | |
| Turbidity (NTU) | 0.2 ± 0.001 | 0.2 ± 0.001 | 0.2 ± 0.001 | *** | |
| Conductivity (μS·cm−1) | 330 ± 1 | 337 ± 2 | 355 ± 1 | *** | |
| Dissolved oxygen (mg·L−1) | 7.9 ± 0.1 | 7.8 ± 0.1 | 7.7 ± 0.1 | *** | |
| Forested | Temperature (°C) | 21.3 ± 0.03 | 21.7 ± 0.06 | 22.3 ± 0.03 | *** |
| pH | 7.4 ± 0.03 | 7.6 ± 0.03 | 7.6 ± 0.03 | *** | |
| Turbidity (NTU) | 0.1 ± 0.0004 | 0.1 ± 0.0004 | 0.1 ± 0.001 | *** | |
| Conductivity (μS·cm−1) | 124 ± 1 | 154 ± 1 | 155 ± 1 | *** | |
| Dissolved oxygen (mg·L−1) | 8.9 ± 0.1 | 9.0 ± 0.1 | 0.1 ± 0.0004 | NS |
| LU | MU | HU | ANOVA | |
|---|---|---|---|---|
| Urb For | Urb For | Urb For | ||
| Temperature (°C) | 22.4 ± 0.04 21.3 ± 0.03 | 23.8 ± 0.03 21.7 ± 0.06 | 23.9 ± 0.03 22.3 ± 0.03 | *** |
| pH | 7.8 ± 0.03 7.4 ± 0.03 | 7.9 ± 0.02 7.6 ± 0.03 | 7.9 ± 0.03 7.6 ± 0.03 | *** |
| Turbidity (NTU) | 0.2 ± 0.001 0.1 ± 0.0004 | 0.2 ± 0.001 0.1 ± 0.0004 | 0.2 ± 0.001 0.1 ± 0.001 | *** |
| Conductivity (μS·cm−1) | 330 ± 1 124 ± 1 | 337 ± 2 154 ± 1 | 355 ± 1 155 ± 1 | *** |
| Dissolved oxygen (mg·L−1) | 7.9 ± 0.1 8.9 ± 0.1 | 7.8 ± 0.1 9.0 ± 0.1 | 7.7 ± 0.1 9.1 ± 0.1 | *** |
| Physicochemical Variables | LU | MU | HU | ANOVA |
|---|---|---|---|---|
| Urban Forested | Urban Forested | Urban Forested | ||
| Depth (m) | 0.42 ± 0.004 0.36 ± 0.006 | 0.30 ± 0.003 0.37 ± 0.004 | 0.53 ± 0.05 0.40 ± 0.003 | *** |
| Substrate index (SI) | 1.52 ± 0.1 3.29 ± 0.13 | 1.20 ± 0.03 1.79 ± 0.069 | 1.19 ± 0.02 1.79 ± 0.05 | *** |
| Pool canopy cover (%) | 29.53 ± 0.99 39.69 ± 0.86 | 27.33 ± 0.99 34.01 ± 1.07 | 19.21 ± 0.67 22.41 ± 1.18 | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Perez, W.X.; Perez-Reyes, O. Population Status of the Tropical Freshwater Shrimp Xiphocaris elongata in Urban and Forest Streams in Puerto Rico. Hydrobiology 2023, 2, 277-288. https://doi.org/10.3390/hydrobiology2010018
Torres-Perez WX, Perez-Reyes O. Population Status of the Tropical Freshwater Shrimp Xiphocaris elongata in Urban and Forest Streams in Puerto Rico. Hydrobiology. 2023; 2(1):277-288. https://doi.org/10.3390/hydrobiology2010018
Chicago/Turabian StyleTorres-Perez, Wesley X., and Omar Perez-Reyes. 2023. "Population Status of the Tropical Freshwater Shrimp Xiphocaris elongata in Urban and Forest Streams in Puerto Rico" Hydrobiology 2, no. 1: 277-288. https://doi.org/10.3390/hydrobiology2010018