
Odonata as Indicators? Dragonflies and Damselflies Respond to Riparian Conditions along Ozark Spring Streams


Department of Biology, Missouri State University, Springfield, MO 65897, USA
*
Author to whom correspondence should be addressed.
Hydrobiology 2023, 2(1), 260-276; https://doi.org/10.3390/hydrobiology2010017
Submission received: 7 December 2022
/
Revised: 5 February 2023
/
Accepted: 27 February 2023
/
Published: 5 March 2023
(This article belongs to the Special Issue Ecology of Aquatic Invertebrates in Springs and Headwater Streams: Imperiled Habitats in a Changing World)
Abstract
:The Odonata are not typically regarded as informative taxa for stream bioassessment in North America compared to other insects, particularly when monitoring the ecological impacts of organic pollution. However, we hypothesized that stream-dwelling odonates are useful bioindicators of riparian conditions because vegetation associated with streams is used for oviposition and establishing breeding territories and is likely a cue for a suitable nymphal habitat. We sampled odonates from multiple microhabitats and all macroinvertebrates from riffle habitat in 12 Ozark Highlands (USA) spring streams along a gradient of riparian conditions. We also measured a standard suite of physical and chemical variables in each stream. We compared various aspects of the odonate and riffle macroinvertebrate assemblages among sites to evaluate sensitivity to variables associated with riparian structure compared to the other physical and chemical variables measured. The odonates were strongly associated with riparian-specific variables, while riffle macroinvertebrates were associated with riparian variables to a lesser degree. The additional environmental variables explained minimal variation in either assemblage. Overall, our results suggest that Odonata alone could be useful for biomonitoring associated with riparian structure around Ozark spring streams. The sensitivity of odonates to riparian conditions in other Nearctic regions should be further studied to identify regional and species-specific differences.
1. Introduction
Riparian zones provide important habitat for many riverine organisms and serve as the primary ecotone between streams and terrestrial ecosystems [1,2,3]. Degradation to riparian zones can have a strong impact on instream communities. Decreased shading over streams following loss of canopy species can increase water temperatures and increase algal growth [4]. The removal of riparian vegetation can increase fine sediment inputs to streams, which may reduce heterogeneity of substrata habitat for macroinvertebrate organisms or alter channel morphology [5]. Riparian degradation also typically reduces allochthonous inputs to aquatic ecosystems, affecting the base of stream food webs and altering animal community composition [6,7]. The importance of riparian zones around streams merits action to preserve them to benefit aquatic resources, so there is a need for rapid assessment methods that indicate the biological suitability of riparian habitats.
Macroinvertebrates are standard bioindicators for the rapid assessment of freshwater ecosystems [8]. Biotic indices used for streams often characterize benthic macroinvertebrates according to sensitivities to water quality, particularly chemical parameters associated with organic pollution. Tolerance values (TVs) are numeric values that reflect the sensitivity of specific benthic taxa to changes in environmental variables of interest and their weighted average produces a single biotic index score for an individual sample site. Most of the commonly applied biotic indices in streams follow the general approach of Chutter [9] and Hilsenhoff [10], applying TVs associated with responses to organic pollution, rather than other instream variables such as sediments [11]. Chutter [9], Hilsenhoff [10] and others (e.g., [12]) stressed the importance of refining identifications to the lowest taxonomic level (species) and using species-specific TVs to achieve the most accurate biological index calculations, because closely related taxa can sometimes have substantially different responses. However, highly resolved taxonomic identification can be difficult to accomplish for many to most macroinvertebrate taxa, thanks to the degree of taxonomic expertise required. The order Odonata is an exception, however, as odonates achieve a relatively large size, and multiple keys exist for species level identifications in North America [13,14,15,16].
Despite their appeal as a highly recognizable taxon, odonates are not often used as focal taxa for stream bioassessment, particularly in temperate regions, due in large part to their general tolerance to organic pollution and their presence in rarely sampled microhabitats in streams. Stream benthic samples are often taken predominantly from one habitat (e.g., riffles) [17] where taxa most sensitive to a focal stressor such as organic pollution tend to be abundant. As such, only the few odonate taxa adapted to that microhabitat are collected while a greater diversity of odonates may occupy other instream habitats (e.g., root-wads, fine sediments, pools, backwaters).
Although mostly pollution-tolerant, Nearctic odonates can be especially sensitive to physical habitat structure [18,19], and the physical structure of the riparian zone is likely to influence community dynamics. Odonates are believed to navigate the world principally by sight and adults have large compound eyes with a wide field of vision that they use to identify predators, prey, mates, rivals and suitable habitat [20,21]. The adults of many odonates, particularly anisopterous species, are strong fliers that can travel long distances to find suitable breeding sites that they select primarily according to local environmental conditions. Key conditions for many species include shading around water bodies, specific vegetation structure for breeding and oviposition [22], or nymphal microhabitat availability (e.g., Aeshna viridis (Eversman) nymphal commensalism with Stratiotes aloides (L.), Rantala et al. [23]). Changes in habitat structure driven by invasive trees [24], agricultural development [25,26,27], urbanization [28] and deforestation within riparian zones [29,30] can affect odonate species residency and abundance in streams and ponds, usually absent of any change in water chemistry. In recent years, several Brazilian studies have shown a broad impact on odonate community structure in response to riparian and substrate configuration (e.g., [27,31,32,33,34,35]). Accordingly, resident odonate assemblages in streams, although largely pollution-tolerant, have strong potential to reflect physical habitat structure, including that of the riparian zone.
Here, we examined and compared sensitivities of odonates versus total macroinvertebrates to the riparian zone condition in spring-fed streams in the Ozark Highlands (USA). We collected odonates at all life stages from multiple instream and riparian microhabitats (henceforth, the “odonate assemblage”), and we collected riffle-dwelling macroinvertebrates by standard methods (henceforth, the “riffle assemblage”). Our overarching hypothesis was that the odonate assemblage would respond more strongly to riparian conditions than the riffle assemblage. We additionally predicted that the riffle assemblage would respond more strongly to environmental variables associated with water chemistry and benthic substrate characteristics. Finally, we discuss a possibility to develop new, odonate-specific TVs that could be applicable for rapid assessment of the riparian condition associated with spring-fed streams.
2. Materials and Methods
2.1. Study Sites
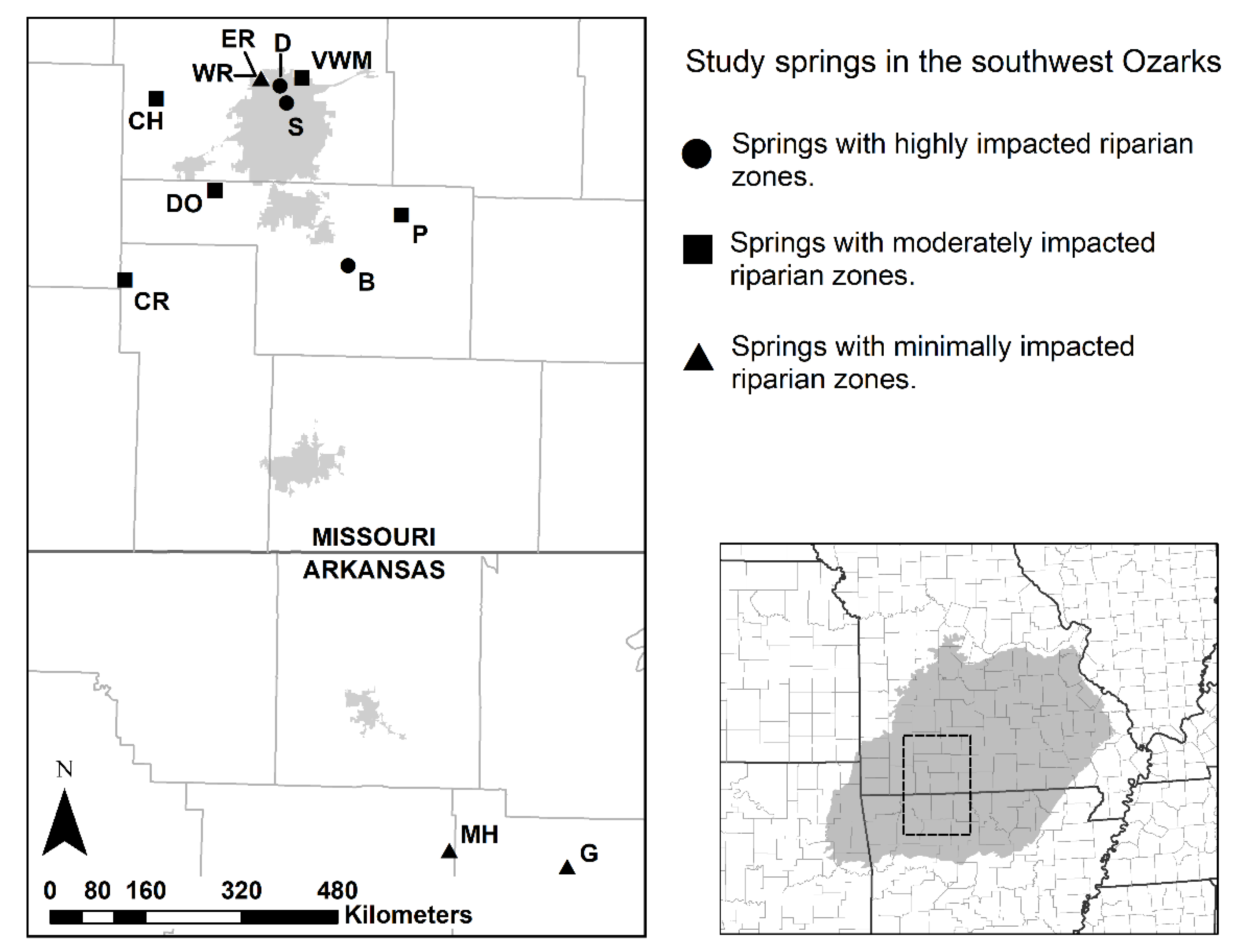
The Ozark Highlands is a karst-dominated ecoregion with a plethora of groundwater-fed springs that provide unique habitat for a diverse array of freshwater macroinvertebrates. The region is home to 258 odonate species [36], of which several are known to use Ozark springs and spring-fed systems [37,38]. We selected 12 springs, each draining either the Springfield or Salem Plateau subregions and occupying the Sac and White River watersheds (Table 1). Site selection criteria included consistent year-round flow magnitude (spring size), and conditions immediately adjacent to each spring stream fell along a gradient of riparian structure from intact riparian zones with a complex three-dimensional structure to heavily impacted riparian zones devoid of woody vegetation. Because stream size influences habitat and macroinvertebrate communities [8,39,40], and springs on the Ozark Plateau vary greatly in magnitude, we primarily selected third magnitude (28–280 L/s) springs (N = 10) and larger fourth magnitude (6.3–28 L/s) springs (N = 2) for this study [41].
At each of the 12 spring streams, we defined the length of the sampling reach according to United States Geological Survey (USGS) protocols as twenty times the wetted width of the sampling point [42]. The sampling point was the focal point for riffle assemblage sample collection. In cases where a stream segment’s total length was less than twenty times its wetted width (Silver and Chiles springs) due to a nearby confluence, we sampled its entire length. At most streams, the sampling reaches we selected were near the spring source (within 50 m). East Ritter, Doling, Brown, and Patterson springs had sampling reaches more distant from the source (Table 1) either due to greater ease of access to the stream or the presence of favorable habitat (i.e., shallow riffles or runs) to collect riffle assemblage samples. We defined the riparian zone adjacent to each sample reach as four times the wetted width of the stream (following Barbour et al. [8]) on both sides of the stream. Due to stable flow in these spring-fed systems, the active channels did not contain surface-dry portions with terrestrial vegetation that would alter riparian zone characterization. The 12 spring streams were categorized into 3 classes of riparian impact based on land-use immediately surrounding the streams: minimally impacted (N = 4), moderately impacted (N = 5) and highly impacted (N = 3) (Figure 1, Table 1). Minimally impacted streams had riparian zones that were undisturbed by human use and were located in mostly forested settings. Moderately impacted streams were in rural or suburban areas with varying degrees of agricultural land use but had at least partially intact riparian zones (i.e., including natural forest vegetation). Sometimes these sites were modified for public recreational use or partly deforested by landowners (e.g., cleared to expand hay fields). Highly impacted streams had riparian zones that were reduced to manicured lawn or inorganic rubble, lacked natural forest settings, and included varying degrees of rockwork or concrete lining their margins downstream of their source.
2.2. Data Collection
2.2.1. Environmental Variables
We measured instream physicochemical variables during daylight hours in June and July 2018 at the focal riffle unit. Instream variables including pH, specific conductivity (µS/cm), total dissolved solids (mg/L), oxidation-reduction potential (mV), dissolved oxygen (% saturation) and temperature (°C) were measured with a Hanna Instruments HI98194 Multiparameter Meter (Hanna Instruments, Woonsocket, Rhode Island). We used Hach Model PO-19 and Model NI-12 test kits (Hach Instruments, Loveland, Colorado) to measure orthophosphate (mg/L), nitrate (mg/L) and nitrite (mg/L) concentrations, respectively. Nitrite concentrations were below detection level at all streams and were omitted from later analyses. We measured volumetric flow at a single cross-section at the focal riffle using a Hach FH950 portable velocity meter (Hach Instruments, Loveland, Colorado) fitted to a top-setting wading rod. We conducted pebble counts in the focal riffles by assigning size classes for 20 particles following Bowles et al. [44], then calculating median particle size (D50). We categorized embeddedness and canopy cover at each site into quinaries of roughly 0, 25, 50, 75 and 100 percent following Barbour et al. [8].
We quantified vegetation at each stream using 3 methods, including a visual estimate of percent cover of instream emergent vegetation and an evaluation of riparian condition at both local and landscape scales. To estimate local riparian condition, we visually approximated the proportion of the riparian zone that was undisturbed and contained mostly natural woody vegetation. The assessment of the local riparian condition was based on a modification of Barbour’s [8] visual assessment protocols. For the landscape riparian condition, we used ArcMap [43] to approximate the percent of undisturbed riparian conditions within a 500 m radius around each focal riffle. We used land-use classification data from the 2011 USGS National Land Cover Database (NLCD). We visually estimated the proportion of each landscape riparian condition buffer area as ‘impacted’ or ‘natural’ habitat based on the NLCD land-use classes. Classes we considered natural habitat included deciduous forest, evergreen forest, mixed forest, woody wetlands, emerging herbaceous wetlands and shrub/scrub.
2.2.2. Biological Samples
We sampled all riffle assemblages during the same time (June and July 2018) that we collected environmental data. Three replicate samples were collected at each site using a Slack Surber sampler with a 500 µm mesh net and a 0.25 m2 sampling frame. Collection of replicate samples (henceforth ‘Surbers’) involved disturbing the substrate within the sampling frame with a long-handled garden claw cultivator for 2 min [40]. Each Surber was collected from riffles or shallow runs with predominantly very coarse gravel substrates (32–64 mm in diameter). We preserved Surbers in 95% ethanol for transport to the laboratory. We identified most taxa to genus, with the exception of Chironomidae, which were only identified to family. Amphipods in the genera Gammarus (Fabricius) and Crangonyx (Bate) were abundant in several Surbers, but many individuals were tiny and difficult to confidently distinguish so these 2 genera were grouped together for statistical analysis. Large Surbers with >2000 specimens were sub-sampled at 25% according to methods described by Bowles et al. [44], using a round 500-µm USGS sieve marked into quadrants.
To characterize the odonate assemblage at each stream, we sampled once during each of 3 seasons, including summer (June/July 2018), fall (October 2018) and spring (March 2019) using a spatially and temporally thorough, semi-quantitative approach that combined species collected with a variety of methods. The rationale for sampling sites over 3 seasons was to aid the identification of any species present as early instar nymphs in one season and account for differences in developmental phenologies among resident species. Here, ‘resident’ refers to odonate species using the streams to complete their life cycles. An ‘occupant’, alternatively, is a species temporarily present at a stream for foraging or exploration for potential breeding habitat [45]. Nymphs and exuviae were the focus of collection because their presence indicates habitat suitability for completing the life cycle. We recorded the presence of every species of adult odonate but only used breeding and ovipositing individuals to indicate species residency. We collected nymphs using the qualitative multihabitat methods for sampling streams described by Moulton et al. [46] by sampling all microhabitats found in each sampling reach with a 500 µm mesh D-frame kick net. Streams were sampled for 15 min during summer, and sample timing was increased to 60 min in fall and spring to ensure thorough investigation of all microhabitats in each sampling reach for detection of rare or cryptic species. Exuviae were collected anywhere encountered while collecting nymphs. Whenever possible, we collected at least 1 adult for each odonate species observed at a stream, including tandem pairs, to verify habitat use regardless of the presence or absence of nymphs of the same species. All odonates collected during Surber sampling for the riffle assemblage were also included in the semi-quantitative odonate counts. Counts were summed across the 3 sampling seasons.
2.3. Data Analysis
We used R version 3.5.3 [47] to conduct all statistical analyses. We performed MANOVA (α = 0.05) to test for differences in the full suite of environmental variables not directly associated with riparian vegetation among the 3 site classes, and we investigated relationships between each vegetation-specific environmental variable (local and landscape riparian conditions, canopy cover, and emergent vegetation cover) and non-vegetation specific variables using Pearson’s correlation analysis. We adjusted α for multiple comparisons using the Holm–Bonferroni method.
We used the ‘vegan’ package [48] to run multivariate analyses of community structure of the odonate assemblage, the riffle assemblage, as well as the insect-only subset of the riffle assemblage. We included an analysis of the insect-only subset because insects tend to be less pollution-tolerant than the non-insects among the stream macroinvertebrates. To examine the similarity of each of these 3 assemblages across study streams, we plotted cluster dendrograms from pairwise Bray–Curtis dissimilarity matrices using the vegdist function. We also used the metaMDS function in the vegan package to perform nonmetric multidimensional scaling (NMS) analyses to further assess variation in the community structure for the 3 assemblages across the 3 riparian-associated site classes. Input data for NMS were the square-root transformed total abundance of each taxon standardized by Wisconsin double standardization with Bray–Curtis dissimilarities. The best configuration for each NMS ordination was determined by selecting the analysis with the lowest stress value after 20 random starts. Here, ’stress‘ is a statistical value that varies from 0 to 1 and represents how well relationships are summarized in an ordination with values closer to 0 indicating better fit. We performed multiple response permutation procedure (MRPP) analyses using Bray–Curtis distance with 9999 permutations to identify whether there were consistent differences in community structure among the 3 site classes for each type of assemblage (riffle assemblage, insect-only component of the riffle assemblage, and odonate assemblage). Then we plotted 95% confidence ellipses in each of the 3 NMS bi-plots around each class of sites in ordination space.
We combined all riparian and instream environmental data into a secondary matrix and then fitted these data as vectors to assemblage ordinations based on correlation coefficients between these data and axis values in each ordination. Before plotting the vectors, they were scaled by their correlation so that the length of each arrow represented the predictive strength of its corresponding variable (longer arrows have higher correlation coefficients). The significance of the vectors was assessed following 9999 random permutations of the data [49]. We plotted vectors on the NMS bi-plots for any variable with significance at p < 0.1.
To determine whether any odonate species was strongly associated with streams in any of the 3 riparian classes, we used the ‘indicspecies’ package [50] for indicator species analysis (ISA). We used untransformed abundance data for odonate species that had multiple detections among sites and across sampling events (n = 13) in ISA. Using the multipatt function, we calculated point biserial correlation coefficients (rpb) for each species in the ISA. These are Pearson correlations that compare the abundance of a species within a site class or grouping of site classes to its abundance within every other site class [50].
3. Results
We identified 85 unique taxa in the riffle assemblage (Table S2) across the 12 spring streams. Dominant orders were Diptera (24 taxa), Coleoptera (14), Ephemeroptera (13) and Trichoptera (13). Other insect orders with just 2 or 3 taxa present in the riffle assemblage were Plecoptera, Megaloptera, Odonata and Hemiptera. There were nine non-insect taxa encountered infrequently in Surbers, but pericaridan crustaceans (amphipods and isopods) were overwhelmingly dominant in some of the streams.
From the 3 seasons of semi-quantitative sampling across all instream microhabitats, we identified 22 resident odonate taxa represented by 11 zygopterous and 11 anisopterous species (Table 2). Ten resident odonate taxa occupied the minimally impacted sites, 14 occupied moderately impacted sites, and 6 occupied highly impacted sites, with some taxa occupying multiple classes. We collected a single Gomphurus (Needham) sp. nymph from the Mitch Hill Spring riffle assemblage which was the only odonate species uniquely found in Surber samples. Argia funebris (Hagen) was the only other odonate species we observed in any riffle assemblage.
Many instream physicochemical habitat variables spanned a wide range across streams, with the exception of pH and ORP (Table 3). The temperature was similar (16.8 ± 3.3, mean ± standard deviation) across all but one site, Doling spring, where the sampling reach was located 180 m downstream of the source allowing this shallow stream to warm considerably (26.6 °C) as it flowed through a concrete channel. Dissolved oxygen ranged from 52.9% saturation at East Ritter spring to 107.1% saturation at Valley Water Mill spring. Conductivity ranged from 316 to 616 µS/cm and TDS from 158 to 308 mg/L, where Patterson and Gilbert springs had the lowest measurements and East Ritter spring had the highest. The substrate size across springs was similar (Table 3), and most streams had high to moderate embeddedness (70.83 ± 23.43%, where 100% equates to fully embedded substrate), except for Patterson spring which had the least embedded substrate (25%). Nitrate and orthophosphate concentrations varied considerably across sites (12.83 ± 5.12, 0.03 ± 0.04, mg/L respectively). However, despite the substantial among-site variability in many physicochemical variables they were not significantly different among the three riparian-associated site classes (F = 0.0836, P = 0.999).
Vegetation-specific habitat variables including local and landscape scale riparian conditions, canopy cover and emergent vegetation cover were correlated with one another. Higher estimates of local riparian condition tended to be associated with higher estimates of landscape riparian condition (r = 0.62, P = 0.031) and greater canopy cover (r = 0.80, P = 0.0019). Emergent vegetation cover generally decreased with increasing canopy cover (r = −0.59, P = 0.042) and landscape riparian condition (r = −0.63, P = 0.029). Excluding local riparian condition, from which the three site classes were directly defined, only canopy cover (F = 4.765, df = 2,9, P = 0.038) was significantly different among site classes. The four minimally impacted sites had primarily intact local riparian zones and likewise, mostly intact riparian vegetation at the broader spatial scale (landscape riparian condition) and 50% or greater canopy cover over their sampling reaches (Table 4). Their channels had ≤50% emergent vegetation. Moderately impacted sites had estimates of local riparian condition at 40–65% and estimates of landscape riparian condition higher than their corresponding estimates of local riparian condition, aside from Double spring (LSRC = 10%). Moderately impacted sites also had ≤50% canopy cover and a wide range of emergent vegetation cover (Table 4). Silver, Doling and Brown springs were heavily impacted with local riparian condition ranging from 0 to 20%. Silver and Doling springs also had a landscape riparian condition of 0 and 10%, respectively, whereas Brown spring had a landscape riparian condition of 80%. Doling spring had a more developed canopy (50%) than Silver and Brown springs (0%). Emergent vegetation peaked at Silver spring (90%), while Doling and Brown springs had emergent vegetation occupying approximately one third of their channels.
Cluster analyses produced different results for each of the three different assemblages tested. Results from the cluster analysis of the riffle assemblages were best explained by relative abundances of peracaridan crustaceans and geographic proximity between sites rather than the riparian condition (Figure S1). The insect-only component of the riffle assemblages showed no clear patterns from the cluster analysis with no distinct natural groupings that explain the most closely related sites (Figure S2). The cluster analysis of the 12 odonate assemblages showed three broad clusters (Figure 2). All minimally impacted sites fell within the largest cluster, along with two moderately impacted sites. All sites within this cluster were represented by a high abundance of Calopteryx maculata (Palisot de Beauvois). The smallest cluster included Brown (highly impacted) and Chiles (moderately impacted) springs, which together had the highest densities of Ischnura posita (Hagen). The final cluster contained the remaining two highly impacted streams and two faunistically similar moderately impacted sites. The highly impacted Silver and Doling springs had a high abundance of Argia funebris (Hagen), while Double and Crane Creek springs had a moderate abundance of this species. All four of these streams with moderate to high abundances of Argia funebris had dense patches of Nasturtium officinale (W.T. Aiton).
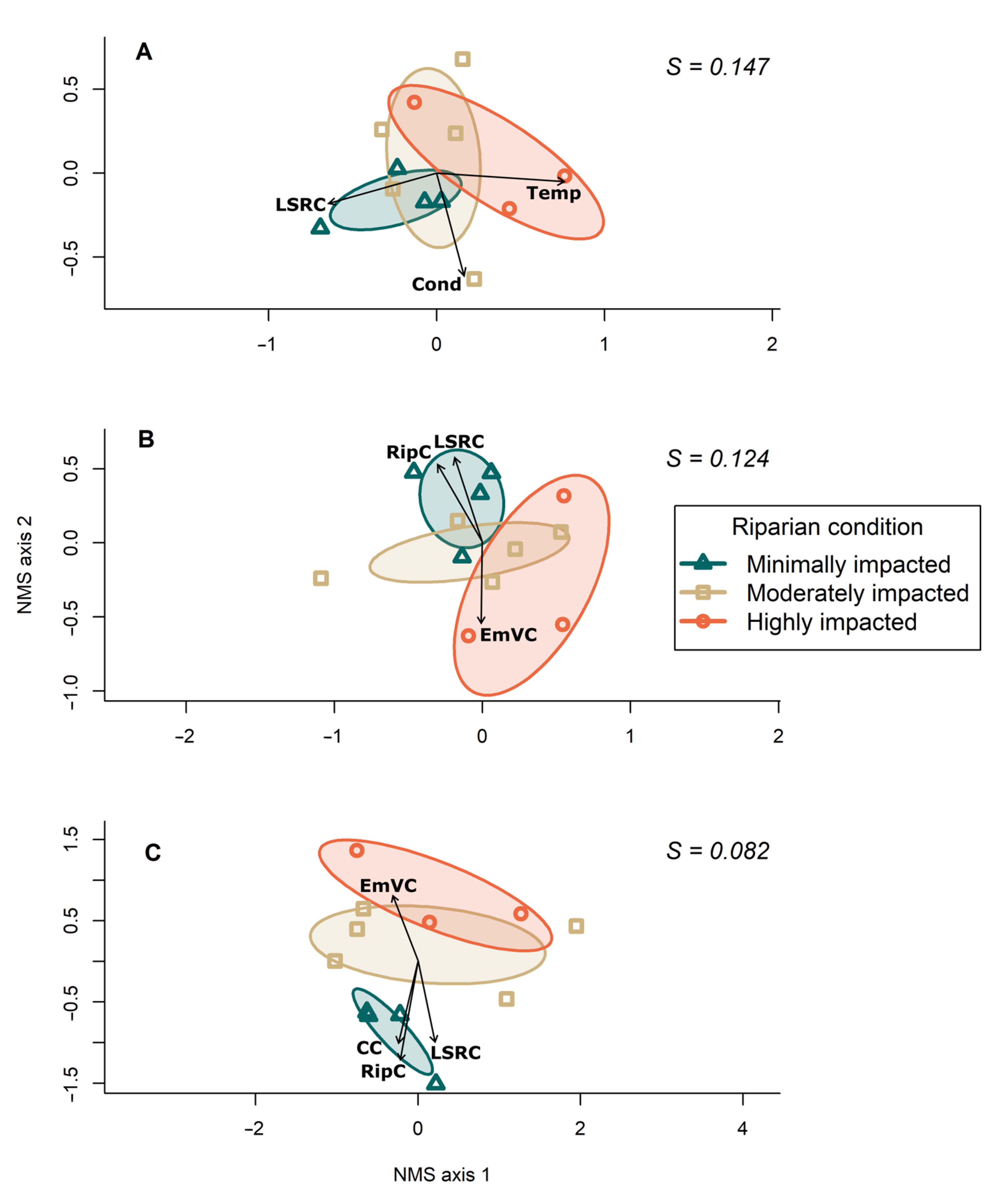
The NMS analysis showed that clusters of sites according to the three site classes of riparian impact became increasingly differentiated in order from riffle assemblage, to the insect-only component of the riffle assemblage, to the odonate assemblage (Figure 3). Statistical stress also decreased with each of these ordinations from the riffle assemblage (S = 0.147), the insect-only component of the riffle assemblage (S = 0.124), and the odonate assemblage (S = 0.082). Multi-response permutation procedure revealed a significant difference among site classes only in the odonate assemblage ordination (MRPP, A = 0.1172, P = 0.04). In the odonate assemblage ordination, the minimally impacted sites were significantly different from moderately impacted (MRPP, A = 0.1111, P = 0.04) and highly impacted sites (MRPP, A = 0.2137, P = 0.01). MRPP showed no significant differences among site classes in the riffle assemblage and the insect-only component of the riffle assemblage (MRPP, A = 0.03671, P = 0.187 and A = 0.01609, P = 0.307, respectively).
Environmental variables associated with the multivariate community structure varied according to assemblage type. Variation among the riffle assemblages was best described by temperature (r = 0.60, P = 0.0044) and to a lesser extent by landscape riparian conditions (r = 0.46, P = 0.0616), and conductivity (r = 0.41, P = 0.0898). For the insect-only component, local (r = 0.54, P = 0.0292) and landscape (r = 0.53, P = 0.0322) riparian conditions were more strongly correlated. The odonate assemblages were strongly correlated with local riparian conditions (r = 0.82, P = 0.0003), followed by canopy cover (r = 0.58, P = 0.0203), landscape riparian condition (r = 0.56, P = 0.0277) and emergent vegetation (r = 0.41, P = 0.0965). Most vegetation-related fitted vectors correlated more strongly with the odonate assemblages than with assemblages collected from the riffles (Table 5).
Among the 22 odonate species collected, only two were significant indicators of riparian class in the indicator species analysis (Table 6). We found the dragonfly Cordulegaster obliqua (Say) at each minimally impacted site on at least two sampling occasions and it was a significant indicator species (rpb = 0.84, P = 0.0071) for streams with minimally impacted local riparian conditions. The damselfly Argia funebris was also a significant indicator species (rpb = 0.61, P = 0.048), but for streams with highly impacted local riparian conditions, despite its apparent absence from the highly impacted Brown spring.
4. Discussion
The odonates sampled across multiple habitat types at each spring stream were more strongly associated with riparian conditions than the riffle assemblage sampled using a standard stream bioassessment method. Odonates were also more sensitive to riparian conditions than the insect-only component of the riffle assemblage, supporting our hypothesis that riparian structure around streams is a crucial predictor of odonate assemblage structure, relative to other physical and chemical instream variables. While odonates typically make up just a small component of stream invertebrate communities, occupying specific types of instream habitats as is common among aquatic macroinvertebrates [51], they appear to be reliable indicators of riparian condition along spring-fed streams. Furthermore, because their large size allows easier species identification compared to other benthic macroinvertebrates, we argue that odonates may be highly useful for the assessment of riparian conditions.
Interestingly, although environmental conditions varied widely among streams, all assemblage types responded to some aspect of riparian structure, and the riffle assemblage only weakly responded to a few measured non-riparian variables, providing little support for our second hypothesis. The odonate assemblage showed no response to measured physical and chemical variables aside from those directly associated with riparian and emergent vegetation. The riparian condition likely impacts the odonate assemblage both directly (as visual cues for adults) but also indirectly, in that the heterogeneity of instream habitat important for the nymph stage is also strongly influenced by riparian condition (especially local riparian conditions) [52]. For example, streams with minimally impacted riparian zones often had more trees along their streambanks and as a result, more root-wad and large wood habitat. This is likely a key factor dictating species-specific sensitivities to local riparian conditions because riparian vegetation structure can greatly influence instream conditions that are optimal for nymphal development and survival [2,31,32].
Despite high variability among abiotic instream conditions across sites, we did not find evidence that the riffle assemblage responded to water chemistry and substrate size aside from a response to conductivity. Instead, the riffle assemblage responded most strongly to temperature, which was most likely driven by the warmest site (Doling spring) where peracaridan crustacean counts were much lower than other sites. Increasing temperature and salinity is known to increase mortality and respiratory ventilation in some gammarid amphipods [53,54], which likely explains the low abundance of these peracaridan crustaceans at Doling spring. NMS analysis of the insect component of the riffle assemblage revealed a significant relationship with local and landscape riparian conditions but these assemblages were not significantly different among our three pre-defined riparian-associated classes. Only the riffle assemblage NMS analysis showed any response to instream environmental variables, and these accounted for less variability among sites than temperature or landscape riparian conditions, providing weak support for our hypothesis that the riffle assemblage would be best described by non-riparian instream variables. However, it is possible that multi-season sampling of the riffle assemblage across sites, as was done with odonates, would yield a few more taxa which might reveal more highly resolved patterns in response to other environmental variables.
While we did not detect strong effects on the riffle assemblage from other environmental variables, they did vary strongly and independently to riparian variables. There were a few extreme chemistry readings at a few sites which included parameters related to dissolved ions or DO, but these parameters were not associated with the riparian condition. Valley Water Mill spring had a much higher DO content than any other stream, perhaps due to abundant aquatic vegetation growing around its source, but with no other outstanding environmental parameters, it is uncertain why this stream lacks many non-odonate insects. It is likely that unmeasured variables like calcium carbonate may favor some taxa (e.g., amphipods). The Ritter and Valley Water Mill springs had high conductivity and their insect assemblages were dominated by elmid and chironomid larvae, which have both been shown to tolerate moderately elevated conductivity in experiments using representatives from montane streams [55]. Nutrient data were measured at coarse resolution with Hach test kits, and it is difficult to attribute much of our nutrient data to local landscape characteristics. Some spring streams in the Ozark ecoregion have high nitrate concentrations due to agricultural practices including extensive use of animal waste as fertilizer, but the ranges reported from previously studied northwest Arkansas springs were typically below most of our readings [56,57]. Further research is needed to evaluate the repeatability and potential drivers of extremely high nitrate concentrations at springs such as Brown, Double and Crane Creek. Furthermore, it would be useful to measure additional chemical characteristics such as various forms of dissolved organic carbon to better evaluate effects of water chemistry on the riffle assemblage.
Although we sampled the riffle assemblage at varying distances downstream of spring sources, it is unlikely this had a strong impact on assemblage characterization. Carroll and Thorp [58] studied three Ozark spring streams where they observed ecotonal shifts from community dominance by peracaridan crustaceans near stream sources to dominance by insects further downstream from the source (up to 145 m), even though physicochemical variability throughout each stream’s channel was negligible. In the current study, Brown spring had the highest abundance of peracaridan crustaceans but was sampled further from its source than all but East Ritter and Doling springs. East Ritter spring was sampled furthest from its source (240 m) and had the fourth highest abundance of amphipods, more so than streams sampled closer to their source like Mitch Hill and Chiles springs, therefore it is difficult to attribute assemblage differences we observed to distance from spring sources. Additionally, water chemistry did not appear to vary with distance sampled from a stream’s source, as Carroll and Thorp [58] also observed. Based on this observation and similar findings from other spring stream studies, Carroll and Thorp [58] concluded that physicochemical variables had little effect on macroinvertebrate community composition in the spring streams they studied, further supporting our own results that show little response to physicochemical variables by the riffle assemblage.
Odonate assemblage patterns were likely driven by two key factors: (1) microhabitat availability at differently impacted sites and (2) dominant taxa. We attribute the higher odonate species richness at moderately impacted sites to a wider array of suitable instream microhabitat diversity. Local riparian conditions at moderately impacted sites may be satisfying the minimum preferences for adult odonates that usually prefer conditions nearer one or the other end of the riparian gradient. For example, some species that trend towards the minimally impacted end of the riparian gradient might find just enough cover or root-wad habitat, while species that trend towards the more heavily impacted end of the gradient find their own suitable habitat characteristics such as Nasturtium patches. Additionally, multivariate dissimilarities of the odonate assemblage were best explained by the two most abundant species, A. funebris and C. maculata. These species were often either rare or absent in streams or the dominant taxon. All anisopterous species, with exception of E. simplicicolis were restricted to the streams falling into the largest cluster containing all minimally impacted sites (Figure 2), which had more shading than streams in the other clusters, save Doling spring, with 50% canopy cover.
Odonate assemblage patterns we observed here conflict with findings from similar studies in Brazil where anisopterous species were more common at streams with greater exposure to sunlight resulting from environmental degradation, and zygopterous species tended to prefer more shaded streams with natural vegetation [27,30,31,33,34,35]. In our study, there were several zygopterous species such as A. funebris that were most abundant in less shaded streams, while Miguel et al. [30] and Calvão et al. [27] observed most Argia spp. seeming to prefer or at least tolerate heavily shaded streams. Calopteryx maculata was the only zygopterous species in our study that mostly preferred minimally impacted riparian conditions with ample cover. The Brazilian studies explain the riparian preferences between suborders in part by the thermoregulatory differences between adults, but biogeographical differences between temperate and tropical zones may explain why the suborders had opposite environmental responses between regions. Preferences may be species-specific, hence species-level identifications will likely be important for bioassessment. Additionally, all taxa collected by Miguel et al. [30] and Calvão et al. [27] were adults, which do not necessarily use the streams they forage around as breeding habitat, highlighting the importance of determining occupant versus resident taxa.
Imperfect detections will inevitably occur when sampling but sometimes a species inexplicably appeared absent in the nymphal stage despite the presence of breeding adults or seemingly favorable instream habitat. For example, Argia funebris was typical at sites containing little or no canopy cover and shallow depths, with dense patches of emergent vegetation like N. officinale (which may serve as a preferred oviposition substrate for A. funebris [59]). However, we also observed breeding adults at the minimally impacted Mitch Hill and moderately impacted Patterson springs, sites that also had some sunny, shallow, and well-vegetated sections in their sampling reaches. Nonetheless, nymphs were never detected at these streams, even with extensive sampling effort. We also observed A. funebris ovipositing into moss at the minimally impacted West Ritter spring during summer, but never detected nymphs. The apparent absence of A. funebris nymphs at these springs might be explained by biotic interactions such as predation from abundant Gammarus amphipods, which have been noted as dominant predators in some stream microhabitats [60] and as facultative predators preferring Baetis (Leach) mayflies to leaf litter under laboratory conditions [61]. Additionally, some there are notable cases of invertebrates that are adept at colonizing harsh or suboptimal habitats but are poor competitors in more optimal locations [62], resulting in distributions restricted to harsh extremes of environmental gradients (e.g., [63,64]). In the case of Argia funebris, nymphs may fail to thrive in spring streams with apparently prime nymphal due to abundant gammarid amphipods or to the presence of competitors for similar resources. Amphipods, for example, were much less abundant in streams where A. funebris was abundant. ISA revealed A. funebris to be an indicator species for highly impacted riparian zones, but it was not resident at the highly impacted Brown spring which mostly lacked shallow reaches and had the highest recorded amphipod abundance.
Cordulegaster obliqua was an indicator for minimally impacted spring streams and the nymphs in this genus are burrowers in fine sediments [16]. All streams in this study had either sand, silt or clay substrates dispersed throughout their sampling reaches, and C. obliqua nymphs were discovered in all three sediment types. Regardless of quantity or type (clay, silt, sand) of nymphal habitat, this species was a resident at all minimally impacted sites, and we failed to detect any in highly to moderately impacted sites, even as non-breeding adults, suggesting that favorable riparian conditions are more important drivers of C. obliqua distribution than nymphal habitat factors like specific sediment characteristics.
While Odonata have not been considered useful for standard bioassessment associated with organic pollution, the results we present here suggest that odonates may be quite useful for bioassessment of riparian conditions. An odonate-specific biotic index assessing riparian integrity could follow the same formula as Hilsenhoff’s biotic index but apply tolerance values (TVs) based strictly on well-understood riparian associations. Tolerance values for an odonate-specific biotic index of riparian integrity could be derived in a few different ways (see Yuan [65]) but should maximize temporal and spatial collection effort. Preliminary examples using a weighted averaging approach [65] based on local riparian conditions applied to observations in this study would produce TVs of roughly 8.5 for A. funebris and 0.65 for C. obliqua. Another potential approach could mirror a biotic index proposed by Huggins and Moffett [66] that incorporates multiple independent TVs derived from specific stream pollutants/stressors (i.e., not necessarily ‘organic pollution’). Riparian-associated TVs will likely be region-specific, so their effectiveness in other locations will need to be tested before application. Just as Lenat’s [67] TVs for southeastern United States taxa sometimes contrasted with Hilsenhoff’s TVs, even at the species level, riparian-specific TVs developed for odonates would be expected to vary among ecoregions. Future sampling of other Ozark streams can help fine tune TVs created for accurate bioassessment throughout the region just as Hilsenhoff [68] improved tolerance values for Wisconsin taxa after collecting data from many additional streams to bolster his original set of TVs [10].
Odonates are shown here to be the most useful group in Ozark spring streams to reflect riparian conditions. We only detected two odonate species in Surber samples taken from riffle units across all 12 streams but identified 21 species in samples taken from the various microhabitats throughout each sampling reach. Although we needed three seasons of sampling to fully characterize the odonate assemblage, the ease of identifying individuals of this order in the lab vastly outweighs the drawback of additional sample dates. Because many odonate taxa are infrequently collected in standard benthic collections for bioassessment, our results have strong implications for future management and bioassessment of riparian ecosystems. Multi-season sampling should be considered in accordance with regional differences in climate and species phenologies. Uncovering species-specific sensitivities should allow effective management and monitoring of riparian zones without the time-consuming processing and identification required by most benthic biomonitoring protocols used with taxa that are less sensitive to riparian conditions. Furthermore, there is great potential in developing a straightforward odonate-specific biotic index based on tolerance values of species to riparian condition.
Supplementary Materials
The following supporting information can be downloaded at: doi.org/10.6084/m9.figshare.21688727.v2 (accessed on 7 December 2022), Figure S1: Supplementary Figure S1; doi.org/10.6084/m9.figshare.21688778.v2 (accessed on 7 December 2022), Figure S2: Supplementary Figure S2; doi.org/10.6084/m9.figshare.22011050.v1 (accessed on 7 December 2022), Table S1: Supplementary Table S1; doi.org/10.6084/m9.figshare.21657797.v2 (accessed on 7 December 2022), Table S2: Supplementary Table S2.
Author Contributions
Conceptualization, C.R.C.; methodology, C.R.C. and D.S.F.; software, C.R.C.; validation, C.R.C. and D.S.F.; formal analysis, C.R.C.; investigation, C.R.C.; resources, D.S.F.; data curation, C.R.C.; writing—original draft preparation, C.R.C.; writing—review and editing, D.S.F.; visualization, C.R.C.; supervision, D.S.F.; project administration, D.S.F.; funding acquisition, C.R.C. and D.S.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Biology Department and Graduate College of Missouri State University.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data used in this study are largely provided in tables and supplementary materials but are also maintained by the corresponding author and are freely available upon request.
Acknowledgments
We thank Nathan Dorff and David Bowles for assistance with fieldwork. We also thank David Bowles for advice provided throughout the data collection and taxonomic ID components of the work, and later for serving as editor of this Hydrobiology special issue. Jeff Williams helped design the study map. Jaxson Priest, Robert Brewer and Skyler Walrath helped process riffle samples. Sean Maher provided help scripting code in R. We thank the National Park Service, Marceline Matlock and the O’Reilly family, Bob Lovett, Mike and Dan Chiles, and City of Springfield, Missouri officials for permissions to access many of the spring streams used in this study. Three anonymous reviewers provided valuable feedback on an earlier version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Thomas, J.; Maser, C.; Rodiek, J. Wildlife Habitats in Managed Rangelands—The Great Basin of Southern Oregon: Riparian Zones; Pacific Northwest Forest and Range Experiment Station, US Department of Agriculture, Forest Service: Portland, OR, USA, 1979; p. 18. [Google Scholar]
- Gregory, S.V.; Swanson, F.J.; Mckee, W.A.; Cummins, K.W. An ecosystem perspective of riparian zones: Focus on links between land and water. Bioscience 1991, 41, 540–551. [Google Scholar] [CrossRef]
- Naiman, R.J.; Décamps, H. The ecology of interfaces: Riparian zones. Annu. Rev. Ecol. Syst. 1997, 28, 621–658. [Google Scholar] [CrossRef] [Green Version]
- Richardson, J.S.; Danehy, R.J. A synthesis of the ecology of headwater streams and their riparian zones in temperate forests. For. Sci. 2007, 53, 131–147. [Google Scholar]
- Henley, W.F.; Patterson, M.A.; Neves, R.J.; Lemly, A.D. Effects of Sedimentation and Turbidity on Lotic Food Webs: A Concise Review for Natural Resource Managers. Rev. Fish. Sci. 2000, 8, 125–139. [Google Scholar] [CrossRef] [Green Version]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Cummins, K.W.; Wilzbach, M.A.; Gates, D.M.; Perry, J.B.; Taliaferro, W.B. Shredders and Riparian Vegetation. Bioscience 1989, 39, 24–30. [Google Scholar] [CrossRef]
- Barbour, M.T.; Gerritsen, J.; Snyder, B.D.; Stribling, J.B. Rapid Bioassessment Protocols For Use in Streams And Wadeable Rivers: Periphyton, Benthic Macroinvertebrates, and Fish, 2nd ed.; U.S. Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999. [Google Scholar]
- Chutter, F. An empirical biotic index of the quality of water in south African streams and rivers. Water Res. 1972, 6, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Hilsenhoff, W.L. Use of Arthropods to Evaluate Water Quality of Streams, 100th ed.; Technical Bulletin-Wisconsin Department of Natural Resources, Division of Conservation: Madison, WI, USA, 1977; pp. 1–15. [Google Scholar]
- Zweig, L.D.; Rabeni, C.F. Biomonitoring for deposited sediment using benthic invertebrates: A test on 4 Missouri streams. J. North Am. Benthol. Soc. 2001, 20, 643–657. [Google Scholar] [CrossRef]
- Lenat, D.R.; Resh, V.H. Taxonomy and stream ecology—The benefits of genus- and species-level identifications. J. North Am. Benthol. Soc. 2001, 20, 287–298. [Google Scholar] [CrossRef]
- Daigle, J.J. Florida Damselflies (Zygoptera): A Species Key to the Aquatic Larval Stages; Technical Series; Florida Department of Environmental Regulation: Tallahassee, FL, USA, 1991; Volume 11, pp. 1–15. [Google Scholar]
- Westfall, M.J.J.; May, M.L. Damselflies of North America; Scientific Publishers: Gainsville, FL, USA, 1996. [Google Scholar]
- Landwer, B.H.P.; Sites, R.W. The Larval Odonata of Ponds in the Prairie Region of Missouri. Trans. Am. Èntomol. Soc. 2010, 136, 1–105. [Google Scholar] [CrossRef]
- Tennessen, K.J. Dragonfly Nymphs of North America: An Identification Guide; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Bowles, D.E.; Williams, M.H.; Dodd, H.R.; Morrison, L.W.; Hinsey, J.A.; Ciak, C.E.; Rowell, G.A.; DeBacker, M.D.; Haack, J. Protocol for Monitoring Aquatic Invertebrates of Small Streams in the Heartland Inventory & Monitoring Network, Version 2.1. 2021. Available online: https://irma.nps.gov/DataStore/Reference/Profile/2284622 (accessed on 15 January 2022). [CrossRef]
- Smith, J.; Samways, M.J.; Taylor, S. Assessing Riparian Quality Using Two Complementary Sets of Bioindicators. Biodivers. Conserv. 2006, 16, 2695–2713. [Google Scholar] [CrossRef]
- Villalobos-Jimenez, G.; Dunn, A.M.; Hassall, C. Dragonflies and damselflies (Odonata) in urban ecosystems: A review. Eur. J. Èntomol. 2016, 113, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Corbet, P.S. Dragonflies: Behaviour and Ecology of Odonata; Harley Books: Colchester, UK, 1999. [Google Scholar]
- Bybee, S.; Córdoba-Aguilar, A.; Duryea, M.C.; Futahashi, R.; Hansson, B.; Lorenzo-Carballa, M.O.; Schilder, R.; Stoks, R.; Suvorov, A.; Svensson, E.I.; et al. Odonata (dragonflies and damselflies) as a bridge between ecology and evolutionary genomics. Front. Zool. 2016, 13, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Corbet, P.S. A Biology of Dragonflies; Witherby: London, UK, 1962. [Google Scholar]
- Rantala, M.J.; Ilmonen, J.; Koskimäki, J.; Suhonen, J.; Tynkkynen, K. The macrophyte, Stratiotes aloides, protects larvae of dragonfly Aeshna viridis against fish predation. Aquat. Ecol. 2004, 38, 77–82. [Google Scholar] [CrossRef]
- Remsburg, A.J.; Olson, A.C.; Samways, M.J. Shade Alone Reduces Adult Dragonfly (Odonata: Libellulidae) Abundance. J. Insect Behav. 2008, 21, 460–468. [Google Scholar] [CrossRef]
- Hornung, J.P.; Rice, C.L. Odonata and wetland quality in southern Alberta, Canada: A preliminary study. Odonatologica 2003, 32, 119–129. [Google Scholar]
- García-García, P.L.; Vázquez, G.; Novelo-Gutiérrez, R.; Favila, M.E. Effects of land use on larval Odonata assemblages in cloud forest streams in central Veracruz, Mexico. Hydrobiologia 2016, 785, 19–33. [Google Scholar] [CrossRef]
- Calvão, L.B.; Juen, L.; Junior, J.M.B.D.O.; Batista, J.D.; Júnior, P.D.M. Land use modifies Odonata diversity in streams of the Brazilian Cerrado. J. Insect Conserv. 2018, 22, 675–685. [Google Scholar] [CrossRef]
- Córdoba-Aguilar, A.; Rocha-Ortega, M. Damselfly (Odonata: Calopterygidae) Population Decline in an Urbanizing Watershed. J. Insect Sci. 2019, 19, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, K.A.; Ali, S.; Ramachandra, T.V. Odonata as indicators of riparian ecosystem health a case study from south western Karnataka, India. Fraseria 2008, 7, 83–95. [Google Scholar]
- Miguel, T.B.; Oliveira-Junior, J.M.B.; Ligeiro, R.; Juen, L. Odonata (Insecta) as a tool for the biomonitoring of environmental quality. Ecol. Indic. 2017, 81, 555–566. [Google Scholar] [CrossRef]
- Pires, M.M.; Müller, N.F.D.; Stenert, C.; Maltchik, L. Influence of different riparian vegetation widths and substrate types on the communities of larval Odonata (Insecta) in southern Brazilian streams. Acta Limnol. Bras. 2020, 32, 1–16. [Google Scholar] [CrossRef]
- Silva, L.F.R.; Castro, D.M.P.; Juen, L.; Callisto, M.; Hughes, R.M.; Hermes, M.G. A matter of suborder: Are Zygoptera and Anisoptera larvae influenced by riparian vegetation in Neotropical Savanna streams? Hydrobiologia 2021, 848, 4433–4443. [Google Scholar] [CrossRef]
- de Oliveira-Junior, J.M.B.; Junior, P.D.M.; Dias-Silva, K.; Leitão, R.P.; Leal, C.; Pompeu, P.S.; Gardner, T.A.; Hughes, R.M.; Juen, L. Effects of human disturbance and riparian conditions on Odonata (Insecta) assemblages in eastern Amazon basin streams. Limnogical 2017, 66, 31–39. [Google Scholar] [CrossRef]
- Oliveira-Junior, J.M.B.; Juen, L. Structuring of dragonfly communities (Insecta: Odonata) in eastern Amazon: Effects of environmental and spatial factors in preserved and altered streams. Insects 2019, 10, 322. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira-Junior, J.M.B.; Shimano, Y.; Gardner, T.A.; Hughes, R.M.; de Marco Júnior, P.; Juen, L. Neotropical dragonflies (Insecta: Odonata) as indicators of ecological condition of small streams in the eastern Amazon. Austral Ecol. 2015, 40, 733–744. [Google Scholar] [CrossRef]
- Sims, G.G. A Distribution of Ozark Odonates; Central Wyoming College: Riverton, WY, USA, 2015; (Personal Communication; Unpublished Data). [Google Scholar]
- Trial, L. Atlas of Missouri Odonata-Revised 2005; Missouri Department of Conservation Report: Columbia, MO, USA, 2005. [Google Scholar]
- Zeller, M.M. Comparative Analysis of Aquatic Insect, Amphipod, and Isopod Community Composition Along Environmental Gradients in Rheocrene Spring Systems of Missouri; University of Missouri: Columbia, MO, USA, 2010. [Google Scholar]
- Brönmark, C.; Herrmann, J.; Malmqvist, B.; Otto, C.; Sjöström, P. Animal community structure as a function of stream size. Hydrobiologia 1984, 112, 73–79. [Google Scholar] [CrossRef]
- Minshall, G.W.; Robinson, C.T. Macroinvertebrate community structure in relation to measures of lotic habitat heterogeneity. Arch. Für Hydrobiol. 1998, 141, 129–151. [Google Scholar] [CrossRef]
- Meinzer, O.E. Large Springs in the United States; Geological Survey Water-Supply Paper 557, Government Printing Office: Washington, DC, USA, 1927; pp. 1–94. [Google Scholar]
- Fitzpatrick, F.A.; Waite, I.R.; D’Arconte, P.J.; Meador, M.R.; Maupin, M.A.; Gurtz, M.E. Revised Methods for Characterizing Stream Habitat in the National Water-Quality Assessment Program; US Geological Survey: Reston, VA, USA, 1998; Volume 98. [Google Scholar]
- ESRI. ArcGIS Version 10. 2012. Available online: http://www.esri.com (accessed on 24 October 2019).
- Bowles, D.E.; Dodd, H.R.; Williams, M.H.; Morrison, L.W.; James, K.; DeBacker, M.D.; Ciak, C.E.; Hinsey, J.A.; Rowell, G.A.; Haack, J.L. Protocol for Monitoring Spring Communities at Ozark National Scenic Riverways, Missouri; National Park Service: Fort Collins, CO, USA, 2008. [Google Scholar]
- Chovanec, A.; Waringer, J. Ecological integrity of river-floodplain systems? assessment by dragonfly surveys (Insecta: Odonata). Regul. Rivers: Res. Manag. 2001, 17, 493–507. [Google Scholar] [CrossRef]
- Moulton II, S.R.; Kennen, J.G.; Goldstein, R.M.; Hambrook, J.A. Revised Protocols for Sampling Algal, Invertebrate, and Fish Communities as Part of the National Water-Quality Assessment Program; Geological Survey (US): Reston, VA, USA, 2002. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, L.G.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-5. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 8 March 2019).
- Oksanen, J. Vegan: An Introduction to Ordination. 2015. Available online: http://cran.r-project.org/web/packages/vegan/vignettes/introvegan.pdf (accessed on 8 March 2019).
- De Cáceres, M.; Legendre, P. Associations between species and groups of sites: Indices and statistical inference. Ecology 2009, 90, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Downing, J.A. The effect of habitat structure on the spatial distribution of freshwater invertebrate populations. Habitat Struct. Phys. Arrange. Objects Space 1991, 8, 87–106. [Google Scholar] [CrossRef]
- Brito, J.S.; Michelan, T.S.; Juen, L. Aquatic macrophytes are important substrates for Libellulidae (Odonata) larvae and adults. Limnology 2020, 22, 139–149. [Google Scholar] [CrossRef]
- Wijnhoven, S.; Van Riel, M.; Van Der Velde, G. Exotic and indigenous freshwater gammarid species: Physiological tolerance to water temperature in relation to ionic content of the water. Aquat. Ecol. 2003, 37, 151–158. [Google Scholar] [CrossRef]
- Sornom, P.; Felten, V.; Médoc, V.; Sroda, S.; Rousselle, P.; Beisel, J.-N. Effect of gender on physiological and behavioural responses of Gammarus roeseli (Crustacea Amphipoda) to salinity and temperature. Environ. Pollut. 2010, 158, 1288–1295. [Google Scholar] [CrossRef]
- Clements, W.H.; Kotalik, C. Effects of major ions on natural benthic communities: An experimental assessment of the US Environmental Protection Agency aquatic life benchmark for conductivity. Freshw. Sci. 2016, 35, 126–138. [Google Scholar] [CrossRef]
- Davis, R.K.; Brahana, J.V.; Johnston, J.S. Ground Water in Northwest Arkansas: Minimizing Nutrient Contamination from Non-Point Sources in Karst Terrane; Arkansas Water Resources Center: Fayetteville, AR, USA, 2000; pp. 1–69. [Google Scholar]
- Peterson, E.W.; Davis, R.K.; Brahana, J.; Orndorff, H.A. Movement of nitrate through regolith covered karst terrane, northwest Arkansas. J. Hydrol. 2002, 256, 35–47. [Google Scholar] [CrossRef]
- Carroll, T.M.; Thorp, J.H. Ecotonal shifts in diversity and functional traits in zoobenthic communities of karst springs. Hydrobiologia 2014, 738, 1–20. [Google Scholar] [CrossRef]
- Bick, G.H.; Bick, J.C. Substrate utilization during reproduction by Argia plana Calvert and Argia moesta (Hagen) (Odonata: Coenagrionidae). Odonatologica 1972, 1, 3–9. [Google Scholar]
- Alam, K.; Negishi, J.N.; Rahman, M.A.T.; Tolod, J.R. Stable isotope ratios of emergent adult aquatic insects can be used as indicators of water pollution in the hyporheic food web. Ecol. Indic. 2020, 118, 106738. [Google Scholar] [CrossRef]
- Kelly, D.W.; Dick, J.T.A.; Montgomery, W.I. The functional role of Gammarus (Crustacea, Amphipoda): Shredders, predators, or both? Hydrobiologia 2002, 485, 199–203. [Google Scholar] [CrossRef]
- Connell, J.H. The Influence of Interspecific Competition and Other Factors on the Distribution of the Barnacle Chthamalus Stellatus. Ecology 1961, 42, 710–723. [Google Scholar] [CrossRef] [Green Version]
- Flory, E.A.; Milner, A.M. Macroinvertebrate community succession in Wolf Point Creek, Glacier Bay National Park, Alaska. Freshw. Biol. 2000, 44, 465–480. [Google Scholar] [CrossRef]
- Madsen, P.B.; Morabowen, A.; Andino, P.; Espinosa, R.; Cauvy-Fraunié, S.; Dangles, O.; Jacobsen, D. Altitudinal distribution limits of aquatic macroinvertebrates: An experimental test in a tropical alpine stream. Ecol. Èntomol. 2015, 40, 629–638. [Google Scholar] [CrossRef]
- Yuan, L.L. Estimation and Application of Macroinvertebrate Tolerance Values. US EPA, ORD; National Center for Environmental Assessment: Washington, DC, USA, 2006. [Google Scholar]
- Huggins, D.G.; Moffett, M. Proposed Biotic and Habitat Indices for Use in Kansas Streams: Report No. 35 of the Kansas Biological Survey; Kansas Biological Survey: Lawrence, KS, USA, 1988. [Google Scholar]
- Lenat, D.R. A Biotic Index for the Southeastern United States: Derivation and List of Tolerance Values, with Criteria for Assigning Water-Quality Ratings. J. North Am. Benthol. Soc. 1993, 12, 279–290. [Google Scholar] [CrossRef]
- Hilsenhoff, W.L. An improved biotic index of organic stream pollution. Gt. Lakes Entomol. 1987, 20, 7. [Google Scholar]
Figure 1.
Map of the stream locations in the Ozark Plateau region of Arkansas and Missouri. This map was created with ArcMap 10.5.1 GIS software [43]. Site abbreviations are defined in Table 1 and site-specific coordinates can be found in Table S1.

Figure 2.
Bray–Curtis dissimilarity dendrogram of the resident odonate assemblage at 12 investigated streams. Shading and shapes indicate the three classes of riparian impact: Orange and circles = highly impacted, Tan and squares = moderately impacted, Green and triangles = minimally impacted. Site abbreviations are defined in Table 1. The ‘height’ scale on the vertical axis of cluster dendrograms gives Bray–Curtis distance measure values between clusters of assemblages.
Figure 2.
Bray–Curtis dissimilarity dendrogram of the resident odonate assemblage at 12 investigated streams. Shading and shapes indicate the three classes of riparian impact: Orange and circles = highly impacted, Tan and squares = moderately impacted, Green and triangles = minimally impacted. Site abbreviations are defined in Table 1. The ‘height’ scale on the vertical axis of cluster dendrograms gives Bray–Curtis distance measure values between clusters of assemblages.

Figure 3.
NMS ordination bi-plots plotting sites according to their: (A) riffle assemblage, (B) insect-only riffle assemblage and (C) odonate assemblage. Ninety-five percent confidence ellipses are plotted for minimally, moderately and highly impacted sites. Vectors represent environmental variables that significantly (p < 0.1) correlated with each ordination after running permutation tests (n = 9999 permutations). Vector length is proportional to the correlation between the environmental variable and the ordination. Abbreviated variables are local riparian conditions (RipC), landscape riparian conditions (LSRC), canopy cover (CC), emergent vegetation cover (EmVC), temperature (Temp) and conductivity (Cond). Stress (S) is reported for each model.
Figure 3.
NMS ordination bi-plots plotting sites according to their: (A) riffle assemblage, (B) insect-only riffle assemblage and (C) odonate assemblage. Ninety-five percent confidence ellipses are plotted for minimally, moderately and highly impacted sites. Vectors represent environmental variables that significantly (p < 0.1) correlated with each ordination after running permutation tests (n = 9999 permutations). Vector length is proportional to the correlation between the environmental variable and the ordination. Abbreviated variables are local riparian conditions (RipC), landscape riparian conditions (LSRC), canopy cover (CC), emergent vegetation cover (EmVC), temperature (Temp) and conductivity (Cond). Stress (S) is reported for each model.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Location and general characteristics of study streams. Includes numeric values in meters (m) for the distance between the source of each stream to the beginning of the sampling reach (Source to Reach), the total length of each stream’s sampling reach (Reach Length) and the width of the local-scale riparian zone from the left and right streambanks (RipC Width).
Table 1.
Location and general characteristics of study streams. Includes numeric values in meters (m) for the distance between the source of each stream to the beginning of the sampling reach (Source to Reach), the total length of each stream’s sampling reach (Reach Length) and the width of the local-scale riparian zone from the left and right streambanks (RipC Width).
| Stream | Stream Code | Spring Magnitude | Source to Reach (m) | Reach Length (m) | RipC Width (m) | Riparian Impactedness |
|---|---|---|---|---|---|---|
| West Ritter | WR | 3rd | 16.5 | 68 | 13.6 | Minimal |
| Valley Water Mill | VWM | 3rd | 43.0 | 124 | 24.8 | Moderate |
| Silver | S | 3rd | 0.0 | 41 | 17.2 | High |
| Patterson | P | 3rd | 72.0 | 68 | 13.6 | Moderate |
| Chiles | CH | 3rd | 0.0 | 33 | 10.6 | Moderate |
| East Ritter | ER | 3rd | 260.0 | 62 | 12.4 | Minimal |
| Doling | D | 4th | 180.0 | 34 | 6.8 | High |
| Mitch Hill | MH | 3rd | 0.0 | 82 | 16.4 | Minimal |
| Gilbert | G | 3rd | 11.0 | 74 | 14.8 | Minimal |
| Double | DO | 3rd | 0.0 | 56 | 11.2 | Moderate |
| Brown | B | 3rd | 140.0 | 54 | 10.8 | High |
| Crane Creek | CR | 4th | 0.0 | 24 | 4.8 | Moderate |
Table 2.
Odonate assemblage data from resident larval detections made from multi-habitat and Slack Surber collections and breeding adult sightings at each stream. Counts represent larval abundance for one-hour-effort kick-net sampling from all different microhabitats within a sampling reach combined with any breeding/ovipositing adult detections and Surber data across three sampling seasons. See text (Methods) for more detail and Table 1 for stream code designations. Superscripts indicate a count that included adults (A) or exuviae (E).
Table 2.
Odonate assemblage data from resident larval detections made from multi-habitat and Slack Surber collections and breeding adult sightings at each stream. Counts represent larval abundance for one-hour-effort kick-net sampling from all different microhabitats within a sampling reach combined with any breeding/ovipositing adult detections and Surber data across three sampling seasons. See text (Methods) for more detail and Table 1 for stream code designations. Superscripts indicate a count that included adults (A) or exuviae (E).
| Species | WR | VWM | S | P | CH | ER | D | MH | G | DO | B | CR | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Anisoptera | |||||||||||||
| Aeshna umbrosa Walker, 1908 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 16 A | 0 | 0 | 0 | 0 | 16 |
| Anax junius Drury, 1773 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Basiaeschna janata Say, 1840 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 7 |
| Boyeria vinosa Say, 1840 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 4 |
| Cordulegaster obliqua Say, 1840 | 14 | 0 | 0 | 0 | 0 | 2 A | 0 | 9 | 8 | 0 | 0 | 0 | 33 |
| Epitheca cyanosura Say, 1840 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| Erythemis simplicicollis Say, 1840 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 1 A | 0 | 4 |
| Gomphurus sp. Needham, 1901 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| Libellula incesta Hagen, 1861 | 0 | 1 E | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Somatochlora tenebrosa Say, 1840 | 0 | 1 | 0 | 0 | 0 | 11 A | 0 | 2 | 0 | 0 | 0 | 0 | 14 |
| Sympetrum corruptum Hagen, 1861 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Zygoptera | |||||||||||||
| Argia funebris Hagen, 1861 | 1 A | 0 | 244 A | 2 A | 0 | 0 | 85 A | 2 A | 0 | 51 A | 0 | 48 A | 433 |
| Argia translata Hagen in Selys, 1865 | 0 | 0 | 0 | 0 | 0 | 0 | 6 | 0 | 0 | 0 | 0 | 0 | 6 |
| Calopteryx maculata Palisot de Beauvois, 1807 | 19 A | 33 A | 0 | 24 A | 0 | 42 A | 18 A | 18 | 28 A | 10 A | 2 A | 2 A | 196 |
| Enallagma basidens Calvert, 1902 | 0 | 25 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 25 |
| Enallagma divagans Selys, 1876 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 1 |
| Enallagma geminatum Kellicott, 1895 | 0 | 6 | 0 | 0 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 14 |
| Enallagma signatum Hagen, 1861 | 0 | 3 | 0 | 0 | 16 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 19 |
| Enallagma vesperum Calvert, 1919 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Ischnura hastata Say, 1840 | 0 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 4 |
| Ischnura posita Hagen, 1861 | 0 | 13 | 0 | 0 | 22 | 0 | 5 | 0 | 0 | 0 | 43 | 0 | 83 |
| Ischnura verticalis Say, 1840 | 0 | 2 | 0 | 0 | 18 A | 0 | 0 | 0 | 0 | 0 | 2 A | 0 | 22 |
| Total | 34 | 87 | 244 | 26 | 71 | 55 | 114 | 61 | 36 | 61 | 48 | 50 |
Table 3.
Physicochemical variables used in ordination analyses. Variables listed here are discharge (Q), pH, oxidation-reduction potential (ORP), dissolved oxygen percent saturation (DO), specific conductivity (Cond), total dissolved solids (TDS), temperature (Temp), nitrates (NO3−), orthophosphates (PO43−), embeddedness (Embed), and median substrate size (D50). All measurements are mid-day readings during summer sampling.
Table 3.
Physicochemical variables used in ordination analyses. Variables listed here are discharge (Q), pH, oxidation-reduction potential (ORP), dissolved oxygen percent saturation (DO), specific conductivity (Cond), total dissolved solids (TDS), temperature (Temp), nitrates (NO3−), orthophosphates (PO43−), embeddedness (Embed), and median substrate size (D50). All measurements are mid-day readings during summer sampling.
| Stream | Q (m3/s) | pH | ORP (mV) | DO (%) | Cond (µS/cm) | TDS (mg/L) | Temp (°C) | NO3− (mg/L) | PO43− (mg/L) | Embed (%) | D50 (mm) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| West Ritter | 0.107 | 6.93 | 261.7 | 73.2 | 586 | 293 | 14.99 | 13.2 | 0.08 | 75 | 38.5 |
| Valley Water Mill | 0.082 | 6.88 | 277.7 | 107.1 | 596 | 298 | 16.50 | 8.8 | 0.00 | 50 | 46.5 |
| Silver | 0.030 | 6.93 | 308.3 | 70.1 | 499 | 250 | 17.37 | 8.8 | 0.04 | 100 | 27.3 |
| Patterson | 0.170 | 7.16 | 246.2 | 73.2 | 316 | 158 | 14.07 | 4.4 | 0.00 | 25 | 54.5 |
| Chiles | 0.029 | 7.03 | 313.9 | 72.9 | 434 | 217 | 14.54 | 17.6 | 0.14 | 75 | 54.5 |
| East Ritter | 0.039 | 7.05 | 204.4 | 52.9 | 616 | 308 | 16.24 | 13.2 | 0.04 | 75 | 38.5 |
| Doling | 0.006 | 7.59 | 231.7 | 56.8 | 594 | 297 | 26.63 | 8.8 | 0.00 | 100 | 77.0 |
| Mitch Hill | 0.100 | 6.77 | 328.7 | 62.1 | 457 | 229 | 15.73 | 13.2 | 0.00 | 75 | 54.5 |
| Gilbert | 0.033 | 7.47 | 264.5 | 84.3 | 317 | 159 | 18.27 | 8.8 | 0.06 | 100 | 38.5 |
| Double | 0.060 | 7.16 | 275.2 | 70.6 | 357 | 178 | 17.29 | 17.6 | 0.04 | 50 | 54.5 |
| Brown | 0.053 | 7.17 | 265.8 | 79.0 | 448 | 224 | 16.00 | 22.0 | 0.02 | 50 | 46.5 |
| Crane Creek | 0.009 | 6.86 | 321.1 | 58.9 | 405 | 203 | 14.96 | 17.6 | 0.04 | 75 | 38.5 |
Table 4.
Vegetation-associated variables used in ordination analyses. Variables listed here are local riparian conditions (RipC), landscape riparian conditions (LSRC), canopy cover (CC) and emergent vegetation cover (EmVC). All measurements are estimations made during summer sampling.
Table 4.
Vegetation-associated variables used in ordination analyses. Variables listed here are local riparian conditions (RipC), landscape riparian conditions (LSRC), canopy cover (CC) and emergent vegetation cover (EmVC). All measurements are estimations made during summer sampling.
| Stream | RipC (% Undisturbed) | LSRC (% Undisturbed) | CC (% Cover) | EmVC (% Cover) |
|---|---|---|---|---|
| West Ritter | 95 | 78 | 75 | 20 |
| Valley Water Mill | 65 | 65 | 50 | 70 |
| Silver | 0 | 0 | 0 | 90 |
| Patterson | 45 | 60 | 50 | 50 |
| Chiles | 40 | 50 | 25 | 20 |
| East Ritter | 95 | 80 | 50 | 50 |
| Doling | 20 | 15 | 50 | 35 |
| Mitch Hill | 85 | 100 | 50 | 10 |
| Gilbert | 100 | 75 | 100 | 0 |
| Double | 50 | 10 | 25 | 90 |
| Brown | 10 | 80 | 0 | 35 |
| Crane Creek | 40 | 75 | 0 | 60 |
Table 5.
Correlation coefficients (r) and p-values for fitted vectors in NMS bi-plots produced from permutation procedure (n = 9999 permutations). See Table 3 and Table 4 for explanation of variable abbreviations. See Figure 3 for explanation of each NMS bi-plot (i.e., A, B and C). Significance codes: ‘***’ p<0.001; ‘**’ p<0.01; ‘*’ p<0.05; ‘·’ p<0.1.
Table 5.
Correlation coefficients (r) and p-values for fitted vectors in NMS bi-plots produced from permutation procedure (n = 9999 permutations). See Table 3 and Table 4 for explanation of variable abbreviations. See Figure 3 for explanation of each NMS bi-plot (i.e., A, B and C). Significance codes: ‘***’ p<0.001; ‘**’ p<0.01; ‘*’ p<0.05; ‘·’ p<0.1.
| Fitted Vector | NMS A r | NMS A p Value | NMS B r | NMS B p Value | NMS C r | NMS C p Value |
|---|---|---|---|---|---|---|
| RipC | 0.39 | 0.105 | 0.54 | 0.0292 * | 0.82 | 0.0003 *** |
| LSRC | 0.46 | 0.0616 · | 0.53 | 0.0322 * | 0.56 | 0.0277 * |
| CC | 0.26 | 0.250 | 0.24 | 0.280 | 0.58 | 0.0203 * |
| EmVC | 0.24 | 0.284 | 0.43 | 0.0772 · | 0.41 | 0.0965 · |
| Temp | 0.60 | 0.0044 ** | 0.32 | 0.175 | 0.036 | 0.896 |
| Cond | 0.41 | 0.0898 · | 0.27 | 0.241 | 0.091 | 0.641 |
Table 6.
Results from the Indicator Species Analysis for resident odonate taxa with multiple detections across sampling seasons. Point-biserial correlation coefficients are designated as (rpb) and p-values were calculated using 9999 permutations. Impactedness refers to the site class each odonate species is an indicator for. Significance codes: ‘***’ p<0.001; ‘**’ p<0.01; ‘*’ p<0.05; ‘·’ p<0.1.
Table 6.
Results from the Indicator Species Analysis for resident odonate taxa with multiple detections across sampling seasons. Point-biserial correlation coefficients are designated as (rpb) and p-values were calculated using 9999 permutations. Impactedness refers to the site class each odonate species is an indicator for. Significance codes: ‘***’ p<0.001; ‘**’ p<0.01; ‘*’ p<0.05; ‘·’ p<0.1.
| Odonate Taxa | rpb | p Value | Impactedness |
|---|---|---|---|
| Cordulegaster obliqua | 0.84 | 0.0071** | Minimally impacted |
| Argia funebris | 0.61 | 0.048* | Highly impacted |
| Calopteryx maculata | 0.58 | 0.14 | Minimally impacted |
| Enallagma geminatum | 0.54 | 0.28 | Moderately impacted |
| Argia translata | 0.50 | 0.24 | Highly impacted |
| Somatochlora tenebrosa | 0.49 | 0.15 | Minimally impacted |
| Enallagma signatum | 0.44 | 0.48 | Moderately impacted |
| Ischnura posita | 0.42 | 0.32 | Highly impacted |
| Aeshna umbrosa | 0.42 | 0.57 | Minimally impacted |
| Boyeria vinosa | 0.42 | 0.57 | Minimally impacted |
| Ischnura verticalis | 0.38 | 0.73 | Moderately impacted |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cheri, C.R.; Finn, D.S. Odonata as Indicators? Dragonflies and Damselflies Respond to Riparian Conditions along Ozark Spring Streams. Hydrobiology 2023, 2, 260-276. https://doi.org/10.3390/hydrobiology2010017
AMA Style
Cheri CR, Finn DS. Odonata as Indicators? Dragonflies and Damselflies Respond to Riparian Conditions along Ozark Spring Streams. Hydrobiology. 2023; 2(1):260-276. https://doi.org/10.3390/hydrobiology2010017
Chicago/Turabian StyleCheri, Cameron R., and Debra S. Finn. 2023. "Odonata as Indicators? Dragonflies and Damselflies Respond to Riparian Conditions along Ozark Spring Streams" Hydrobiology 2, no. 1: 260-276. https://doi.org/10.3390/hydrobiology2010017