

Predicting Habitat and Distribution of an Interior Highlands Regional Endemic Winter Stonefly (Allocapnia mohri) in Arkansas Using Random Forest Models
 and
and
Abstract
:1. Introduction
2. Materials and Methods
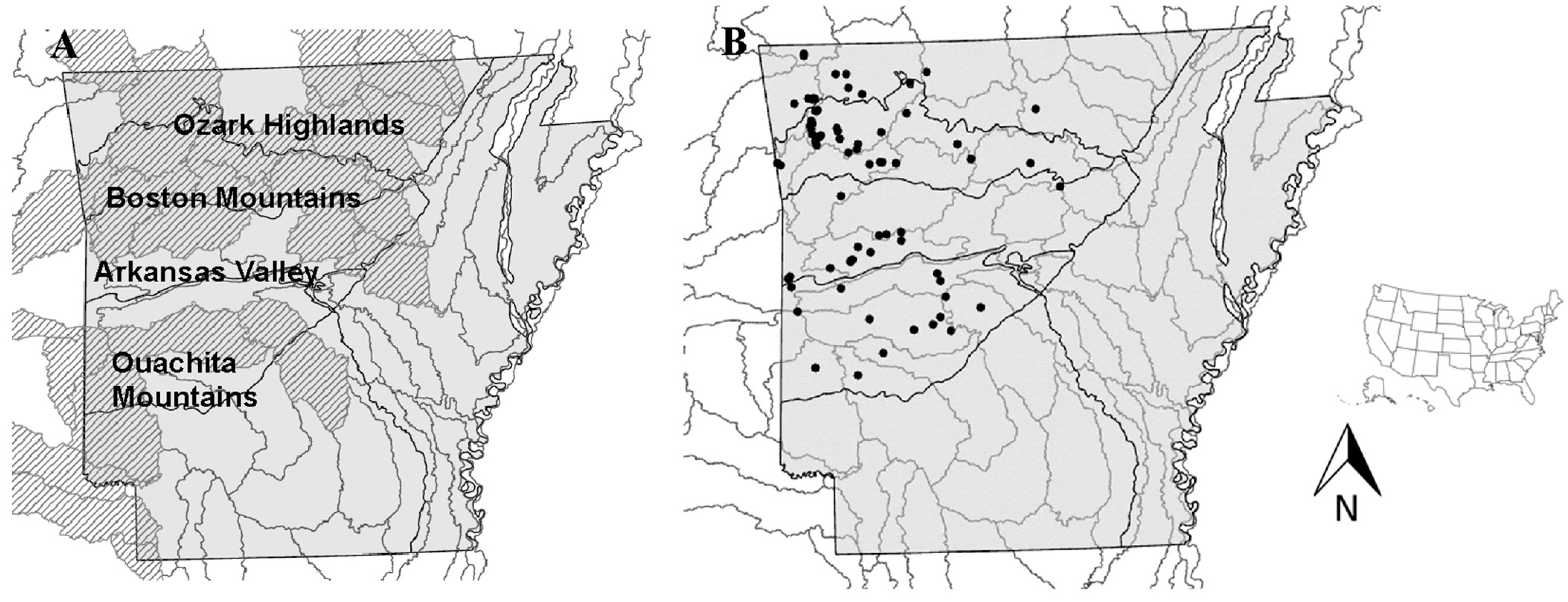
2.1. Study Design
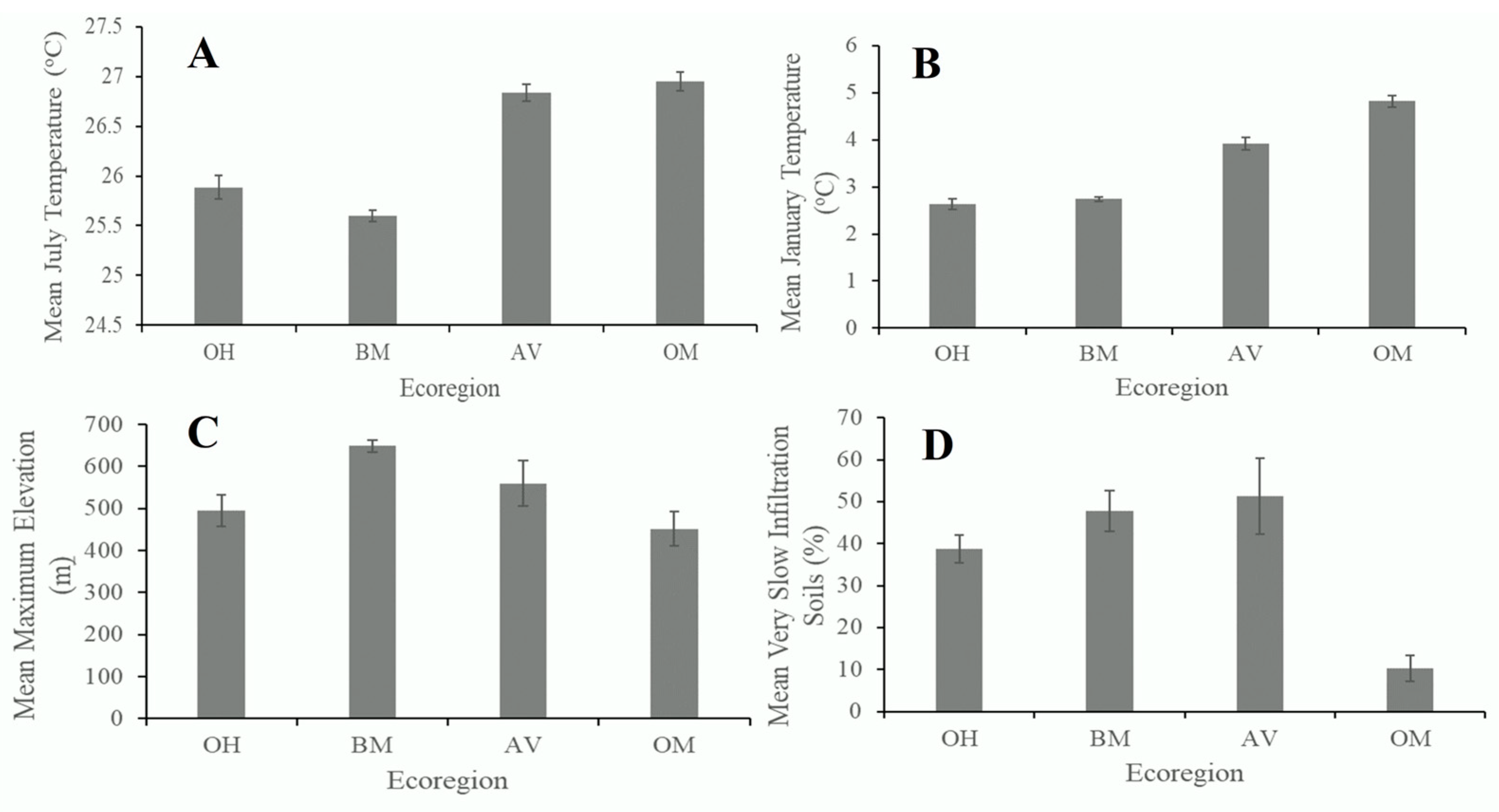
2.2. Local Site Characterization
2.3. Landscape-Level Site Characterization
2.4. A. mohri Presence
2.5. Statistical Analyses and Modeling
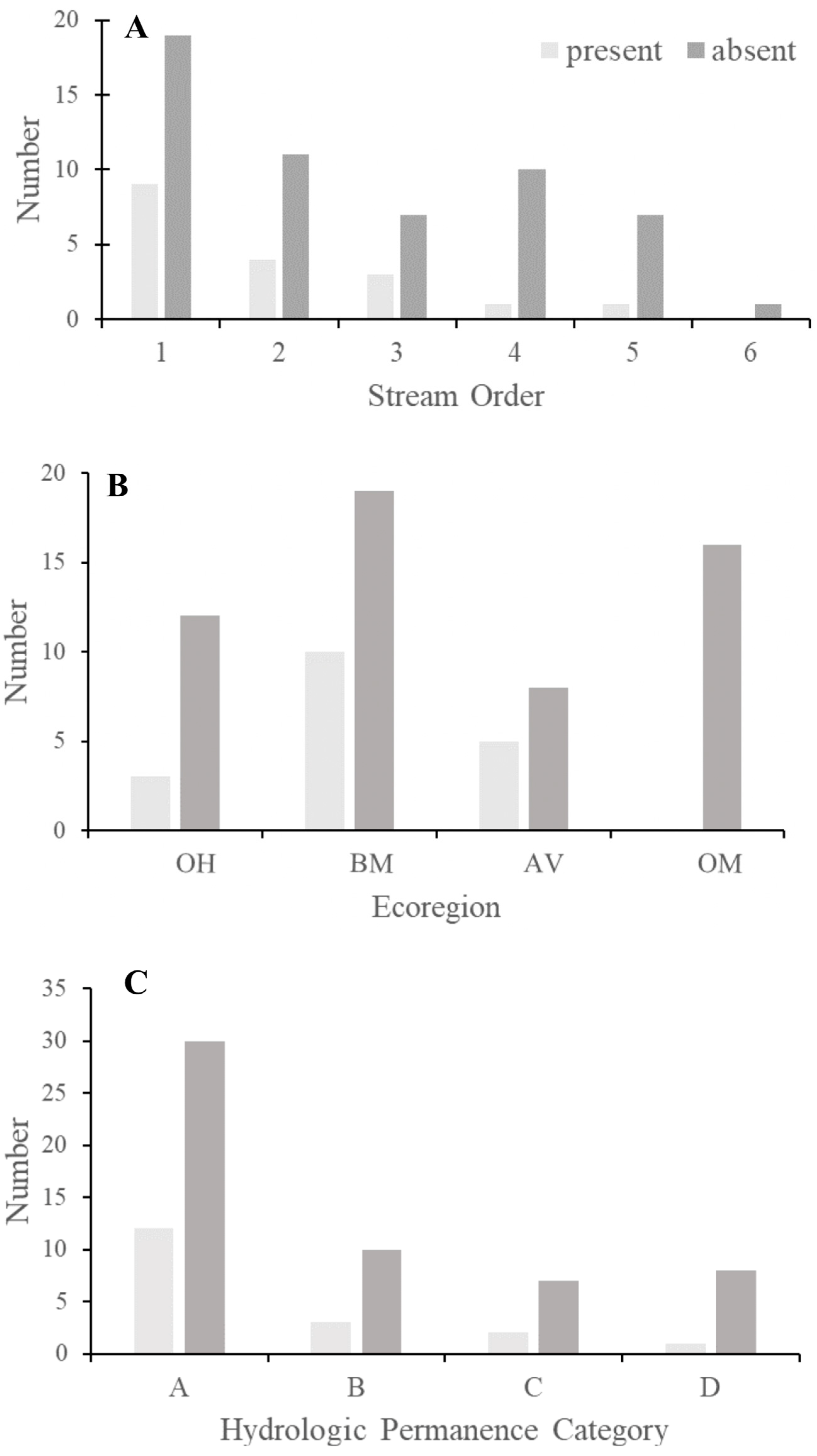
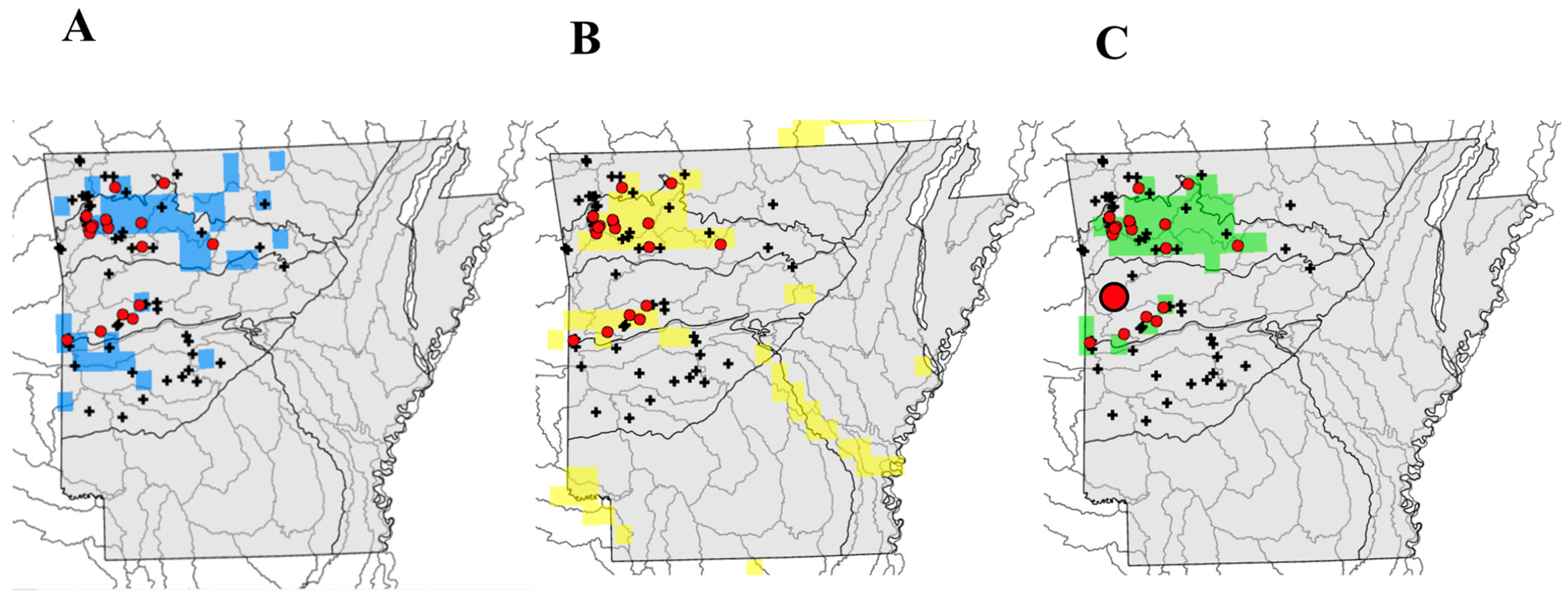
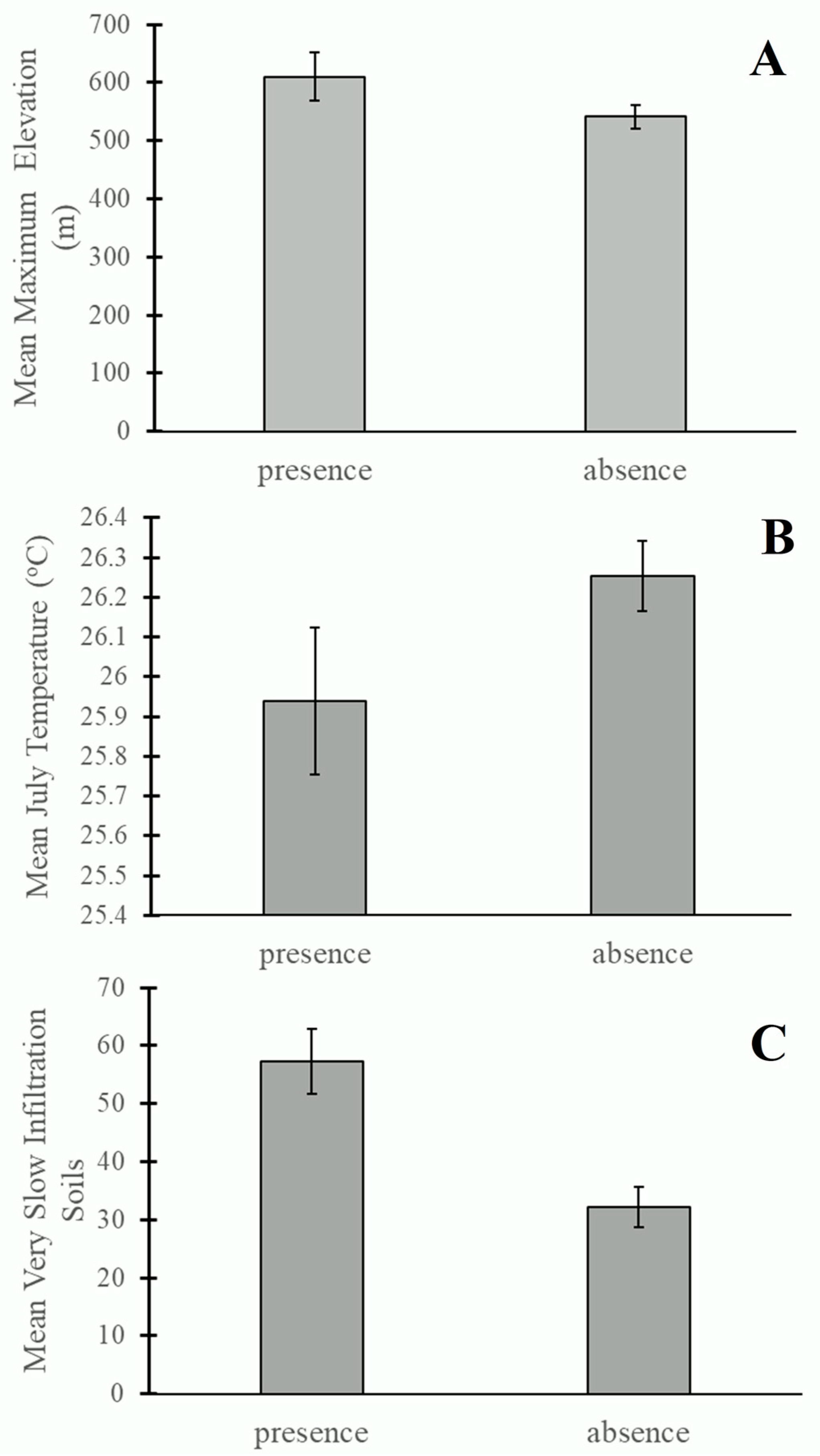
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DeWalt, R.E.; Ower, G.D. Ecosystem Services, Global Diversity, and Rate of Stonefly Species Descriptions (Insecta: Plecoptera). Insects 2019, 10, 99. [Google Scholar] [CrossRef] [PubMed]
- Fochetti, R.; De Figueroa, J.M.T. Notes on diversity and conservation of the European fauna of Plecoptera (Insecta). J. Nat. Hist. 2006, 40, 2361–2369. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Allan, J.D.; Castillo, M.M.; Capps, K.A. Stream Ecology: Structure and Function of Running Waters, 3rd ed.; Springer: Cham, Switzerland, 2021. [Google Scholar]
- Fisher, S.G.; Likens, G.E. Energy Flow in Bear Brook, New Hampshire: An Integrative Approach to Stream Ecosystem Metabolism. Ecol. Monogr. 1973, 43, 421–439. [Google Scholar] [CrossRef]
- Hornick, L.E.; Webster, J.R.; Benfield, E.F. Periphyton Production in an Appalachian Mountain Trout Stream. Am. Midl. Nat. 1981, 106, 22. [Google Scholar] [CrossRef]
- Martínez, A.; Larrañaga, A.; Pérez, J.; Basaguren, A.; Pozo, J. Leaf-litter quality effects on stream ecosystem functioning: A comparison among five species. Fundam. Appl. Limnol. 2013, 183, 239–248. [Google Scholar] [CrossRef]
- Perry, W.B.; Benfield, E.F.; Perry, S.A.; Webster, J.R. Energetics, Growth, and Production of a Leaf-Shredding Stonefly in an Appalachian Mountain Stream. J. N. Am. Benthol. Soc. 1987, 6, 12–25. [Google Scholar] [CrossRef]
- Thorp, J.H.; Rogers, D.C. Chapter 22—Stoneflies: Insect Order Plecoptera. In Field Guide to Freshwater Invertebrates of North America; Thorp, J.H., Rogers, D.C., Eds.; Academic Press: Boston, MA, USA, 2011; pp. 199–204. [Google Scholar]
- Zwick, P. Phylogenetic System and Zoogeography of the Plecoptera. Annu. Rev. Èntomol. 2000, 45, 709–746. [Google Scholar] [CrossRef] [PubMed]
- Maguire, T.J.; Mundle, S.O.C. Citizen Science Data Show Temperature-Driven Declines in Riverine Sentinel Invertebrates. Environ. Sci. Technol. Lett. 2020, 7, 303–307. [Google Scholar] [CrossRef]
- Poulton, B.C.; Stewart, K.W. The stoneflies of the Ozark and Ouachita Mountains (Plecoptera). Mem. Am. Entomol. Soc. 1991, 38, 1–116. [Google Scholar]
- IPCC, 2013: Climate Change 2013: The Physical Science Basis: Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013.
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Further evidence for a global decline of the entomofauna. Austral Èntomol. 2020, 60, 9–26. [Google Scholar] [CrossRef]
- Stanford, J.A.; Gaufin, A.R. Hyporheic Communities of Two Montana Rivers. Science 1974, 185, 700–702. [Google Scholar] [CrossRef]
- Lancaster, J.; Hildrew, A.G. Flow Refugia and the Microdistribution of Lotic Macroinvertebrates. J. N. Am. Benthol. Soc. 1993, 12, 385–393. [Google Scholar] [CrossRef]
- Pusey, B.J.; Arthington, A. Importance of the riparian zone to the conservation and management of freshwater fish: A review. Mar. Freshw. Res. 2003, 54, 1–16. [Google Scholar] [CrossRef]
- Burdon, F.J.; McIntosh, A.; Harding, J.S. Habitat loss drives threshold response of benthic invertebrate communities to deposited sediment in agricultural streams. Ecol. Appl. 2013, 23, 1036–1047. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.B.; Eggert, S.L.; Meyer, J.L.; Webster, J.R. Effects of Resource Limitation on a Detrital-Based Ecosystem. Ecol. Monogr. 1999, 69, 409–442. [Google Scholar] [CrossRef]
- Cross, W.F.; Wallace, J.B.; Rosemond, A.D.; Eggert, S.L. Whole-System Nutrient Enrichment Increases Secondary Production in a Detritus-Based Ecosystem. Ecology 2006, 87, 1556–1565. [Google Scholar] [CrossRef]
- Likens, G.E.; Bormann, F.H.; Johnson, N.M.; Fisher, D.W.; Pierce, R.S. Effects of Forest Cutting and Herbicide Treatment on Nutrient Budgets in the Hubbard Brook Watershed-Ecosystem. Ecol. Monogr. 1970, 40, 23–47. [Google Scholar] [CrossRef]
- Lee, J.-W.; Lee, S.-W.; An, K.-J.; Hwang, S.-J.; Kim, N.-Y. An Estimated Structural Equation Model to Assess the Effects of Land Use on Water Quality and Benthic Macroinvertebrates in Streams of the Nam-Han River System, South Korea. Int. J. Environ. Res. Public Health 2020, 17, 2116. [Google Scholar] [CrossRef]
- Waschbusch, R.J.; Selbig, W.; Bannerman, R.T. Sources of phosphorus in stormwater and street dirt from two urban residential basins in Madison. Wisconsin 1994, 95, 1999. [Google Scholar] [CrossRef]
- Omernik, J.M. The Influence of Land Use on Stream Nutrient Levels; US Environmental Protection Agency, Office of Research and Development, Corvallis Environmental Research Laboratory, Eutrophication Survey Branch: Corvallis, OR, USA, 1976. [Google Scholar]
- Paul, M.J.; Meyer, J.L. Streams in the Urban Landscape. In Urban Ecology: An International Perspective on the Interaction Between Humans and Nature; Marzluff, J.M., Shulenberger, E., Endlicher, W., Alberti, M., Bradley, G., Ryan, C., ZumBrunnen, C., Simon, U., Eds.; Springer US: Boston, MA, USA, 2008; pp. 207–231. [Google Scholar]
- Booth, D.B.; Jackson, C.R. Urbanization of Aquatic Systems: Degradation Thresholds, Stormwater Detection, and the Limits of Mitigation. JAWRA J. Am. Water Resour. Assoc. 1997, 33, 1077–1090. [Google Scholar] [CrossRef]
- Freeborn, J.R.; Sample, D.J.; Fox, L.J. Residential Stormwater: Methods for Decreasing Runoff and Increasing Stormwater Infiltration. J. Green Build. 2012, 7, 15–30. [Google Scholar] [CrossRef]
- King, R.S.; Scoggins, M.; Porras, A. Stream biodiversity is disproportionately lost to urbanization when flow permanence declines: Evidence from southwestern North America. Freshw. Sci. 2016, 35, 340–352. [Google Scholar] [CrossRef]
- Dodds, W.K. Trophic state, eutrophication and nutrient criteria in streams. Trends Ecol. Evol. 2007, 22, 669–676. [Google Scholar] [CrossRef]
- Stringfellow, W.; Herr, J.; Litton, G.; Brunell, M.; Borglin, S.; Hanlon, J.; Chen, C.; Graham, J.; Burks, R.; Dahlgren, R.; et al. Investigation of river eutrophication as part of a low dissolved oxygen total maximum daily load implementation. Water Sci. Technol. 2009, 59, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Suberkropp, K. Annual production of leaf-decaying fungi in a woodland stream. Freshw. Biol. 1997, 38, 169–178. [Google Scholar] [CrossRef]
- Evans-White, M.A.; Haggard, B.E.; Scott, J.T. A Review of Stream Nutrient Criteria Development in the United States. J. Environ. Qual. 2013, 42, 1002–1014. [Google Scholar] [CrossRef] [PubMed]
- Halvorson, H.M.; Scott, J.T.; Sanders, A.J.; Evans-White, M.A. A stream insect detritivore violates common assumptions of threshold elemental ratio bioenergetics models. Freshw. Sci. 2015, 34, 508–518. [Google Scholar] [CrossRef]
- Howard-Parker, B.; White, B.; Halvorson, H.M.; Evans-White, M.A. Light and dissolved nutrients mediate recalcitrant organic matter decomposition via microbial priming in experimental streams. Freshw. Biol. 2020, 65, 1189–1199. [Google Scholar] [CrossRef]
- Eckert, R.A.; Halvorson, H.M.; Kuehn, K.A.; Lamp, W.O. Macroinvertebrate community patterns in relation to leaf-associated periphyton under contrasting light and nutrient conditions in headwater streams. Freshw. Biol. 2020, 65, 1270–1287. [Google Scholar] [CrossRef]
- Rosemond, A.D.; Benstead, J.P.; Bumpers, P.M.; Gulis, V.; Kominoski, J.S.; Manning, D.W.P.; Suberkropp, K.; Wallace, J.B. Experimental nutrient additions accelerate terrestrial carbon loss from stream ecosystems. Science 2015, 347, 1142–1145. [Google Scholar] [CrossRef] [PubMed]
- Stewart, K.W.; Stark, B.P. Plecoptera (Chapter 14). In An Introduction to the Aquatic Insects of North America, 4th ed.; Merritt, R.W., Cummins, K.W., Berg, M.B., Eds.; Kendall/Hunt Publishing Company: Dubuque, IW, USA, 2007; pp. 311–384. 1158p. [Google Scholar]
- Ab Hamid, S.; Rawi, C.S. Application of Aquatic Insects (Ephemeroptera, Plecoptera and Trichoptera) in Water Quality Assessment of Malaysian Headwater. Trop. Life Sci. Res. 2017, 28, 143–162. [Google Scholar] [CrossRef]
- McCafferty, W.P.; Provonsha, A.V. Aquatic Entomology: The Fishermen’s and Ecologists’ Illustrated Guide to Insects and Their Relatives; Jones and Barlett Publishers. Inc.: Boston, MA, USA, 1983. [Google Scholar]
- Guisan, A.; Zimmermann, N.E. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Breiman, L. Classification and Regression Trees, 1st ed.; Chapman & Hall/CRC: Boca Raton, FL, USA, 1984. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Franke, I.; Herzog, S.K.; Pacheco, V.; Paniagua, L.; Quintana, H.L.; Soto, A.; Swenson, J.J.; Tovar, C.; Valqui, T.; et al. Predicting species distributions in poorly-studied landscapes. Biodivers. Conserv. 2008, 17, 1353–1366. [Google Scholar] [CrossRef]
- Grubbs, S.; Sheldon, A.L. The stoneflies (Insecta, Plecoptera) of the Talladega Mountain region, Alabama, USA: Distribution, elevation, endemism, and rarity patterns. Biodivers. Data J. 2018, 6, e22839. [Google Scholar] [CrossRef]
- Ross, H.H.; Ricker, W.E. The Classification, Evolution, and Dispersal of the Winter Stonefly Genus Allocapnia; University of Illinois Press: Urbana, IL, USA, 1971; Volume 45. [Google Scholar]
- Merritt, R.W.; Cummins, K.W.; Berg, M.B. An Introduction to the Aquatic Insects of North America, 5th ed.; Kendall/Hunt Publishing Company: Dubuque, IW, USA, 2019. [Google Scholar]
- Cao, Y.; DeWalt, R.E.; Robinson, J.L.; Tweddale, T.; Hinz, L.; Pessino, M. Using Maxent to model the historic distributions of stonefly species in Illinois streams: The effects of regularization and threshold selections. Ecol. Model. 2013, 259, 30–39. [Google Scholar] [CrossRef]
- Leasure, D.R.; Magoulick, D.D.; Longing, S.D. Natural Flow Regimes of the Ozark-Ouachita Interior Highlands Region. River Res. Appl. 2014, 32, 18–35. [Google Scholar] [CrossRef]
- Newbury, R.; Bates, D.; Alex, K.L. Restoring Habitat Hydraulics with Constructed Riffles. Earth Space Sci. 2013, 194, 353–366. [Google Scholar] [CrossRef]
- Gore, J.A. Chapter 3—Discharge Measurements and Streamflow Analysis; Academic Press: Cambridge, MA, USA, 2006. [Google Scholar]
- Sheldon, A.L.; Jr, M.L.W. Filters and templates: Stonefly (Plecoptera) richness in Ouachita Mountains streams, U.S.A. Freshw. Biol. 2009, 54, 943–956. [Google Scholar] [CrossRef]
- Stroud Water Research Center. Model My Watershed [Software]. 2020. Available online: https://wikiwatershed.org/ (accessed on 9 December 2022).
- Dewitz, J. National Land Cover Dataset (NLCD) 2016 Products; U.S. Geological Survey Data Release: Reston, VA, USA, 2019. [Google Scholar] [CrossRef]
- Soil Survey Staff, 2020. The Gridded National Soil Survey Geographic (gNATSGO) Database for Arkansas. United States Department of Agriculture, Natural Resources Conservation Service. 2020. Available online: https://nrcs.app.box.com/v/soils (accessed on 10 December 2022).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Tarter, D.C.; Chaffee, D.L.; Grubbs, S.A. Revised Checklist of The Stoneflies (Plecoptera) Of Kentucky, U.S.A. Entomol. News 2006, 117, 1–10. [Google Scholar] [CrossRef]
- McRoberts, T.; Grubbs, S. Effects of stream permanence on stonefly (Insecta, Plecoptera) community structure at Mammoth Cave National Park, Kentucky, USA. Biodivers. Data J. 2021, 9, e62242. [Google Scholar] [CrossRef] [PubMed]
- ArcGIS Pro, Version 2.7.2; ESRI: Redlands, CA, USA, 2020.
- Chicco, D.; Jurman, G. The advantages of the Matthews correlation coefficient (MCC) over F1 score and accuracy in binary classification evaluation. BMC Genom. 2020, 21, 6. [Google Scholar] [CrossRef]
- Boughorbel, S.; Jarray, F.; El Anbari, M. Optimal classifier for imbalanced data using Matthews Correlation Coefficient metric. PLoS ONE 2017, 12, e0177678. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 4 December 2022).
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef]
- Snellen, R.K.; Stewart, K.W. The Life Cycle of Perlesta placida (Plecoptera: Perlidae) in an Intermittent Stream in Northern Texas1. Ann. Èntomol. Soc. Am. 1979, 72, 659–666. [Google Scholar] [CrossRef]
- Sheldon, A.L. Possible climate-induced shift of stoneflies in a southern Appalachian catchment. Freshw. Sci. 2012, 31, 765–774. [Google Scholar] [CrossRef]
- Garcia-Raventós, A.; Viza, A.; de Figueroa, J.M.T.; Riera, J.L.; Múrria, C. Seasonality, species richness and poor dispersion mediate intraspecific trait variability in stonefly community responses along an elevational gradient. Freshw. Biol. 2017, 62, 916–928. [Google Scholar] [CrossRef]
- Sheldon, A.L.; Grubbs, S.A. Distributional ecology of a rare, endemic stonefly. Freshw. Sci. 2014, 33, 1119–1126. [Google Scholar] [CrossRef]
- Strahler, A.N. Quantitative analysis of watershed geomorphology. Eos Trans. Am. Geophys. Union 1957, 38, 913–920. [Google Scholar] [CrossRef]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The River Continuum Concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Hotaling, S.; Shah, A.A.; Dillon, M.E.; Giersch, J.J.; Tronstad, L.M.; Finn, D.S.; Woods, H.A.; Kelley, J.L. Cold Tolerance of Mountain Stoneflies (Plecoptera: Nemouridae) from the High Rocky Mountains. West. N. Am. Nat. 2021, 81, 54–62. [Google Scholar] [CrossRef]
- Mulholland, P.J.; Marzolf, E.R.; Webster, J.R.; Hart, D.; Hendricks, S.P. Evidence that hyporheic zones increase heterotrophic metabolism and phosphorus uptake in forest streams. Limnol. Oceanogr. 1997, 42, 443–451. [Google Scholar] [CrossRef]
- Burgmer, T.; Hillebrand, H.; Pfenninger, M. Effects of climate-driven temperature changes on the diversity of freshwater macroinvertebrates. Oecologia 2006, 151, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Durance, I.; Ormerod, S.J. Climate change effects on upland stream macroinvertebrates over a 25-year period. Glob. Chang. Biol. 2007, 13, 942–957. [Google Scholar] [CrossRef]
- Hering, D.; Schmidt-Kloiber, A.; Murphy, J.; Lücke, S.; Zamora-Muñoz, C.; López-Rodríguez, M.J.; Huber, T.; Graf, W. Potential impact of climate change on aquatic insects: A sensitivity analysis for European caddisflies (Trichoptera) based on distribution patterns and ecological preferences. Aquat. Sci. 2009, 71, 3–14. [Google Scholar] [CrossRef]
- Domisch, S.; Jähnig, S.C.; Haase, P. Climate-change winners and losers: Stream macroinvertebrates of a submontane region in Central Europe. Freshw. Biol. 2011, 56, 2009–2020. [Google Scholar] [CrossRef]
- Shokri, M.; Cozzoli, F.; Vignes, F.; Bertoli, M.; Pizzul, E.; Basset, A. Metabolic rate and climate change across latitudes: Evidence of mass-dependent responses in aquatic amphipods. J. Exp. Biol. 2022, 225, jeb244842. [Google Scholar] [CrossRef]
- Pélissié, M.; Johansson, F.; Hyseni, C. Pushed Northward by Climate Change: Range Shifts With a Chance of Co-occurrence Reshuffling in the Forecast for Northern European Odonates. Environ. Èntomol. 2022, 51, 910–921. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Range | Median | Mean |
|---|---|---|---|
| Stream Order | 1st–6th | 2 | 2.48 |
| Mean Width (m) | 0.66–6.52 | 2.6 | 2.99 |
| Mean Depth (cm) | 5.56–41.40 | 14.72 | 15.63 |
| Elevation (m) | 155–598 | 401 | 370.37 |
| %Forested | 8.61–98.6 | 86.25 | 76.26 |
| %Agriculture | 0.00–51.57 | 7.74 | 12.62 |
| %Urban | 0.23–57.73 | 5.03 | 10.75 |
| %High Infiltration Soils | 0.00–66.76 | 2.32 | 4.36 |
| %Moderate Infiltration Soils | 0.00–84.21 | 19.25 | 24.79 |
| %Low Infiltration Soils | 0.00–100.00 | 16.82 | 27.04 |
| %Very Slow Infiltration Soils | 0.00–96.10 | 38.15 | 38.31 |
| Total Annual Precipitation (cm) | 110.0–155.3 | 127.1 | 125.7 |
| Total February Precipitation (cm) | 5.9–10.8 | 7.9 | 7.9 |
| Total May Precipitation (cm) | 12.6–17.5 | 14.7 | 14.8 |
| Annual Mean Water Temperature (°C) | 14.2–16.9 | 14.7 | 15.2 |
| January Mean Water Temperature (°C) | 2.1–5.6 | 2.8 | 3.4 |
| July Mean Water Temperature (°C) | 25.1–27.6 | 25.9 | 26.2 |
| Model | Variables | Removed Due to Correlation and/or Collinearity |
|---|---|---|
| H1 Model | Ecoregion, Elevation, Average Elevation, Max Elevation, Min Elevation | Average Elevation, Min Elevation, Elevation |
| H2 Model | %Forest, %Agriculture, %Urban | N/A |
| H3 Model | Annual Precip Total (cm), Annual Mean Temp (°C), July Mean Temp, Jan Mean Temp, May Total Precip, Feb Total Precip | Annual Mean Temp, Jan Mean Temp, May Total Temp, Feb Total Precip |
| H4 Model | High Infiltration, Moderate Infiltration, Slow Infiltration, Very Slow Infiltration | Slow Infiltration |
| Model | Out of Bag Score | F1 Score | MSE | MCC | Accuracy | Validation Accuracy |
|---|---|---|---|---|---|---|
| Model 1: Landscape | 0.594 | 0.86 | 6.94 | 0.83 | 0.92 | 0.75 |
| Model 2: Climate | 0.375 | 0.8 | 11.008 | 0.75 | 0.92 | 0.88 |
| Model 3: Climate + Landscape | 0.406 | 0.57 | 9.631 | 0.45 | 0.91 | 0.62 |
| Model | df | Chi-Squared | p-Value | AIC Score |
|---|---|---|---|---|
| H1 Model | 3 | 0.34 | p < 0.05 | 80.37 |
| H2 Model | 2 | 0.23 | p > 0.05 | 83.36 |
| H3 Model | 2 | 0.31 | p < 0.05 | 80.35 |
| H4 Model | 2 | 0.53 | p < 0.01 | 73.49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Annaratone, B.; Larson, C.; Prater, C.; Dowling, A.; Magoulick, D.D.; Evans-White, M.A. Predicting Habitat and Distribution of an Interior Highlands Regional Endemic Winter Stonefly (Allocapnia mohri) in Arkansas Using Random Forest Models. Hydrobiology 2023, 2, 196-211. https://doi.org/10.3390/hydrobiology2010013
Annaratone B, Larson C, Prater C, Dowling A, Magoulick DD, Evans-White MA. Predicting Habitat and Distribution of an Interior Highlands Regional Endemic Winter Stonefly (Allocapnia mohri) in Arkansas Using Random Forest Models. Hydrobiology. 2023; 2(1):196-211. https://doi.org/10.3390/hydrobiology2010013
Chicago/Turabian StyleAnnaratone, Brianna, Camryn Larson, Clay Prater, Ashley Dowling, Daniel D. Magoulick, and Michelle A. Evans-White. 2023. "Predicting Habitat and Distribution of an Interior Highlands Regional Endemic Winter Stonefly (Allocapnia mohri) in Arkansas Using Random Forest Models" Hydrobiology 2, no. 1: 196-211. https://doi.org/10.3390/hydrobiology2010013