
Classification and Secondary Metabolite-Biosynthetic Gene Clusters of Marine Streptomyces Strains Including a Lobophorin- and Divergolide-Producer
Abstract
:1. Introduction
2. Results
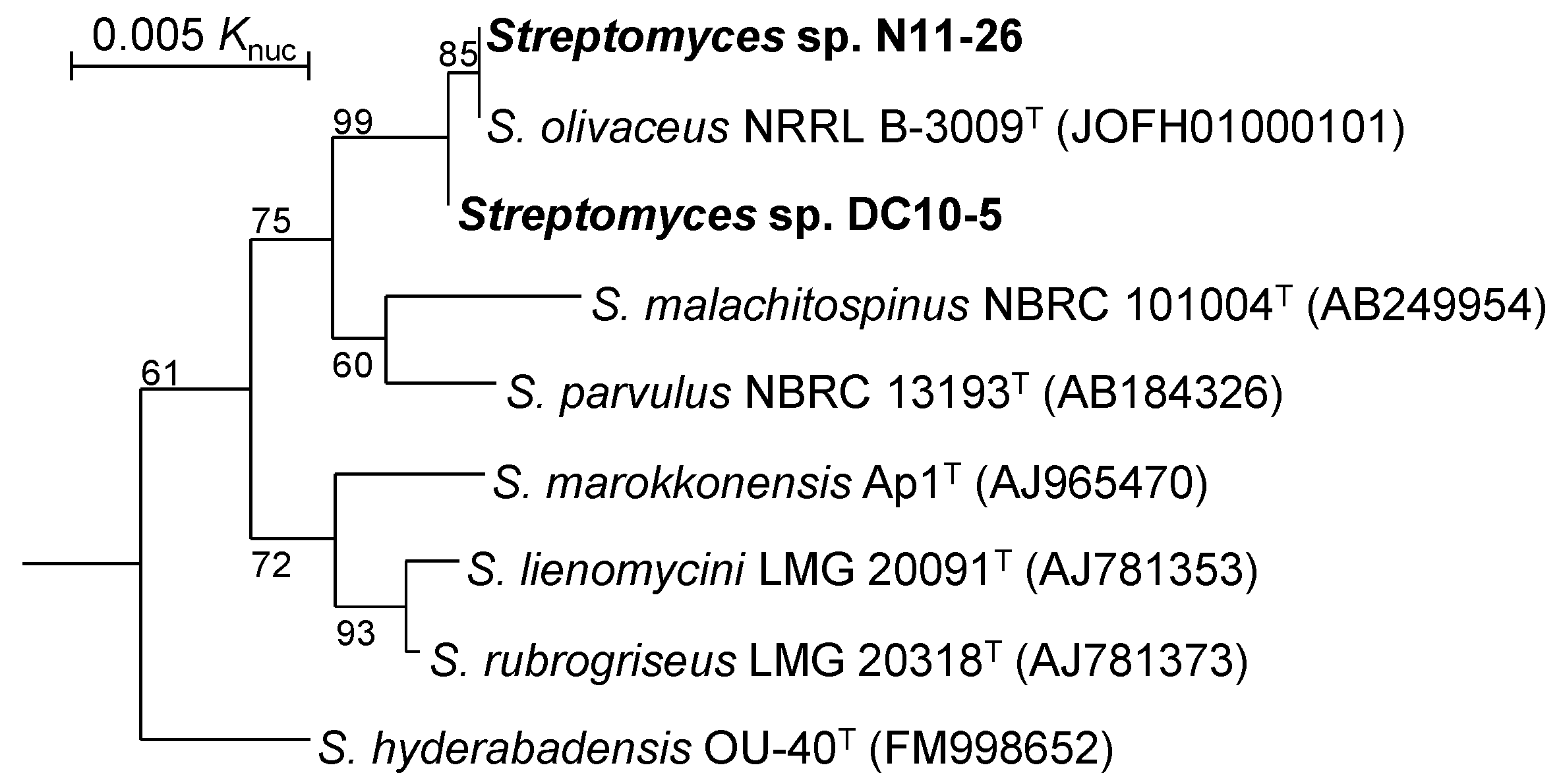
2.1. Classification of Streptomyces Strains N11-26 and DC10-5
2.2. Secondary Metabolite-Biosynthetic Gene Clusters Encoding PKSs and/or NRPSs
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2019, 36, 122–173. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Hwang, S.; Kim, J.; Cho, S.; Palsson, B.; Cho, B. Mini review: Genome mining approaches for the identification of secondary metabolite biosynthetic gene clusters in Streptomyces. Comput. Struct. Biotechnol. J. 2020, 18, 1548–1556. [Google Scholar] [CrossRef] [PubMed]
- Harunari, E.; Ogino, K.; Kanaki, S.; Kumagai, T.; Igarashi, Y. Isolation and metabolites analysis of actinomycetes from deep-sea water in Toyama Bay. Deep Ocean Water Res. 2021, 22, 49–57. [Google Scholar]
- Li, S.; Xiao, J.; Zhu, Y.; Zhang, G.; Yang, C.; Zhang, H.; Ma, L.; Zhang, C. Dissecting glycosylation steps in lobophorin biosynthesis implies an iterative glycosyltransferase. Org. Lett. 2013, 15, 1374–1377. [Google Scholar] [CrossRef]
- Li, S.; Zhao, G.; Sun, M.; He, H.; Wang, H.; Li, Y.; Lu, C.; Shen, Y. Identification and characterization of the biosynthetic gene cluster of divergolides from Streptomyces sp. W112. Gene 2014, 544, 93–99. [Google Scholar] [CrossRef]
- Song, L.; Barona-Gomez, F.; Corre, C.; Xiang, L.; Udwary, D.W.; Austin, M.B.; Noel, J.P.; Moore, B.S.; Challis, G.L. Type III polyketide synthase beta-ketoacyl-ACP starter unit and ethylmalonyl-CoA extender unit selectivity discovered by Streptomyces coelicolor genome mining. J. Am. Chem. Soc. 2006, 128, 14754–14755. [Google Scholar] [CrossRef]
- Ding, L.; Maier, A.; Fiebig, H.H.; Gorls, H.; Lin, W.; Peschel, G.; Hertweck, C. Divergolides A-D from a mangrove endophyte reveal an unparalleled plasticity in ansa-macrolide biosynthesis. Angew. Chem. Int. Ed. Engl. 2011, 50, 1630–1634. [Google Scholar] [CrossRef]
- Luo, M.; Tang, L.; Dong, Y.; Huang, H.; Deng, Z.; Sun, Y. Antibacterial natural products lobophorin L and M from the marine-derived Streptomyces sp. 4506. Nat. Prod. Res. 2021, 35, 5581–5587. [Google Scholar] [CrossRef]
- Um, S.; Lee, J.; Kim, S. Lobophorin producing endophytic Streptomyces olivaceus JB1 associated with Maesa japonica (Thunb.) Moritzi & Zoll. Front. Microbiol. 2022, 13, 881253. [Google Scholar]
- Aoki, Y.; Matsumoto, D.; Kawaide, H.; Natsume, M. Physiological role of germicidins in spore germination and hyphal elongation in Streptomyces coelicolor A3(2). J. Antibiot. 2011, 64, 607–611. [Google Scholar] [CrossRef]
- Nett, M.; Ikeda, H.; Moore, B.S. Genomic basis for natural product biosynthetic diversity in the actinomycetes. Nat. Prod. Rep. 2009, 26, 1362–1384. [Google Scholar] [CrossRef]
- Fischbach, M.A.; Walsh, C.T. Assembly-line enzymology for polyketide and nonribosomal peptide antibiotics: Logic, machinery, and mechanisms. Chem. Rev. 2006, 106, 3468–3496. [Google Scholar] [CrossRef] [PubMed]
- Doroghazi, J.R.; Albright, J.C.; Goering, A.W.; Ju, K.S.; Haines, R.R.; Tchalukov, K.A.; Labeda, D.P.; Kelleher, N.L.; Metcalf, W.W. A roadmap for natural product discovery based on large-scale genomics and metabolomics. Nat. Chem. Biol. 2014, 10, 963–968. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Baunach, M.; Ding, L.; Peng, H.; Franke, J.; Hertweck, C. Biosynthetic code for divergolide assembly in a bacterial mangrove endophyte. ChemBioChem 2014, 15, 1274–1279. [Google Scholar] [CrossRef]
- Becerril, A.; Alvarez, S.; Brana, A.F.; Rico, S.; Diaz, M.; Santamaria, R.I.; Salas, J.A.; Mendez, C. Uncovering production of specialized metabolites by Streptomyces argillaceus: Activation of cryptic biosynthesis gene clusters using nutritional and genetic approaches. PLoS ONE 2018, 13, e0198145. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; Niu, J.; Liu, N.; Lu, Y.; Liu, M.; Li, Y. Cloning and identification of the lobophorin biosynthetic gene cluster from marine Streptomyces olivaceus strain FXJ7.023. Pak. J. Pharm. Sci. 2016, 29, 287–293. [Google Scholar] [PubMed]
- Bentley, S.D.; Chater, K.F.; Cerdeno-Tarraga, A.M.; Challis, G.L.; Thomson, N.R.; James, K.D.; Harris, D.E.; Quail, M.A.; Kieser, H.; Harper, D.; et al. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 2002, 417, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Giessen, T.W.; Franke, K.B.; Knappe, T.A.; Kraas, F.I.; Bosello, M.; Xie, X.; Linne, U.; Marahiel, M.A. Isolation, structure elucidation, and biosynthesis of an unusual hydroxamic acid ester-containing siderophore from Actinosynnema mirum. J. Nat. Prod. 2012, 75, 905–914. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, M.; Zhang, Q.; Zhang, X.; Yang, P.; Liu, Z.; Deng, Y.; Zhu, Y.; Huang, X.; Han, L.; et al. Diisonitrile natural product SF2768 functions as a chalkophore that mediates copper acquisition in Streptomyces thioluteus. ACS Chem. Biol. 2017, 12, 3067–3075. [Google Scholar] [CrossRef]
- Cruz-Morales, P.; Kopp, J.F.; Martinez-Guerrero, C.; Yanez-Guerra, L.A.; Selem-Mojica, N.; Ramos-Aboites, H.; Feldmann, J.; Barona-Gomez, F. Phylogenomic analysis of natural products biosynthetic gene clusters allows discovery of arseno-organic metabolites in model streptomycetes. Genome Biol. Evol. 2016, 8, 1906–1916. [Google Scholar] [CrossRef]
- Cruz-Morales, P.; Vijgenboom, E.; Iruegas-Bocardo, F.; Girard, G.; Yanez-Guerra, L.A.; Ramos-Aboites, H.E.; Pernodet, J.L.; Anne, J.; van Wezel, G.P.; Barona-Gomez, F. The genome sequence of Streptomyces lividans 66 reveals a novel tRNA-dependent peptide biosynthetic system within a metal-related genomic island. Genome Biol. Evol. 2013, 5, 1165–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, S.; Zhang, W.; Zhang, G.; Zhu, Y.; Chen, Y.; Liu, W.; Yuan, C.; Zhang, Q.; Zhang, H.; Zhang, L.; et al. Activation and characterization of a cryptic gene cluster reveals a cyclization cascade for polycyclic tetramate macrolactams. Chem. Sci. 2017, 8, 1607–1612. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Probst, K.; Linnenbrink, A.; Arnold, M.; Paululat, T.; Zeeck, A.; Bechthold, A. Cloning and heterologous expression of three type II PKS gene clusters from Streptomyces bottropensis. ChemBioChem 2012, 13, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Sun, C.; Huang, H.; Gui, C.; Wang, L.; Li, Q.; Ju, J. Biosynthetic Baeyer-Villiger chemistry enables access to two anthracene scaffolds from a single gene cluster in deep-dea-derived Streptomyces olivaceus SCSIO T05. J. Nat. Prod. 2018, 81, 1570–1577. [Google Scholar] [CrossRef]
- Zeng, J.; Decker, R.; Zhan, J. Biochemical characterization of a type III polyketide biosynthetic gene cluster from Streptomyces toxytricini. Appl. Biochem. Biotechnol. 2012, 166, 1020–1033. [Google Scholar] [CrossRef]
- Chen, R.; Zhang, Q.; Tan, B.; Zheng, L.; Li, H.; Zhu, Y.; Zhang, C. Genome mining and activation of a silent PKS/NRPS gene cluster direct the production of totopotensamides. Org. Lett. 2017, 19, 5697–5700. [Google Scholar] [CrossRef]
- Kodani, S.; Komaki, H.; Hemmi, H.; Miyake, Y.; Kaweewan, I.; Dohra, H. Streptopeptolin, a cyanopeptolin-type peptide from Streptomyces olivochromogenes. ACS Omega 2018, 3, 8104–8110. [Google Scholar] [CrossRef]
- Banskota, A.H.; McAlpine, J.B.; Sorensen, D.; Aouidate, M.; Piraee, M.; Alarco, A.M.; Omura, S.; Shiomi, K.; Farnet, C.M.; Zazopoulos, E. Isolation and identification of three new 5-alkenyl-3,3(2H)-furanones from two Streptomyces species using a genomic screening approach. J. Antibiot. 2006, 59, 168–176. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Pokhrel, A.R.; Nguyen, C.T.; Pham, V.T.T.; Dhakal, D.; Lim, H.N.; Jung, H.J.; Kim, T.; Yamaguchi, T.; Sohng, J.K. Streptomyces sp. VN1, a producer of diverse metabolites including non-natural furan-type anticancer compound. Sci. Rep. 2020, 10, 1756. [Google Scholar] [CrossRef]
- Komaki, H.; Ichikawa, N.; Hosoyama, A.; Fujita, N.; Igarashi, Y. Draft genome sequence of Streptomyces sp. TP-A0882 reveals putative butyrolactol biosynthetic pathway. FEMS Microbiol. Lett. 2015, 362, fnv155. [Google Scholar] [CrossRef]
- McClure, R.A.; Goering, A.W.; Ju, K.S.; Baccile, J.A.; Schroeder, F.C.; Metcalf, W.W.; Thomson, R.J.; Kelleher, N.L. Elucidating the rimosamide-detoxin natural product families and their biosynthesis using metabolite/gene cluster correlations. ACS Chem. Biol. 2016, 11, 3452–3460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Wang, J.; Guo, H.; Hou, W.; Yang, N.; Ren, B.; Liu, M.; Dai, H.; Liu, X.; Song, F.; et al. Three antimycobacterial metabolites identified from a marine-derived Streptomyces sp. MS100061. Appl. Microbiol. Biotechnol. 2013, 97, 3885–3892. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.; Zhang, S.; Wang, N.; Li, Z.; Hua, H.; Hu, J.; Wang, S. New spirotetronate antibiotics, lobophorins H and I, from a South China Sea-derived Streptomyces sp. 12A35. Mar. Drugs 2013, 11, 3891–3901. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.; Li, S.; Chen, Y.; Tian, X.; Zhang, H.; Zhang, G.; Zhang, W.; Yang, X.; Zhang, S.; Ju, J.; et al. Lobophorins E and F, new spirotetronate antibiotics from a South China Sea-derived Streptomyces sp. SCSIO 01127. J. Antibiot. 2011, 64, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Cruz, P.G.; Fribley, A.M.; Miller, J.R.; Larsen, M.J.; Schultz, P.J.; Jacob, R.T.; Tamayo-Castillo, G.; Kaufman, R.J.; Sherman, D.H. Novel lobophorins inhibit oral cancer cell growth and induce Atf4- and Chop-dependent cell death in murine fibroblasts. ACS Med. Chem. Lett. 2015, 6, 877–881. [Google Scholar] [CrossRef]
- Brana, A.F.; Sarmiento-Vizcaino, A.; Osset, M.; Perez-Victoria, I.; Martin, J.; de Pedro, N.; de la Cruz, M.; Diaz, C.; Vicente, F.; Reyes, F.; et al. Lobophorin K, a new natural product with cytotoxic activity produced by Streptomyces sp. M-207 associated with the deep-sea coral Lophelia pertusa. Mar. Drugs 2017, 15, 144. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Kong, F.; Xie, Q.; Ma, Q.; Hu, Z.; Zhao, Y.; Luo, D. Divergolides T–W with apoptosis-inducing activity from the mangrove-derived actinomycete Streptomyces sp. KFD18. Mar. Drugs 2019, 17, 219. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, H.; Ju, J. On-PKS Baeyer-Villiger-type O-atom insertion catalyzed by luciferase-like monooxygenase OvmO during olimycin biosynthesis. Org. Lett. 2020, 22, 1780–1784. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Pinto-Tomas, A.A.; Cheng, K.; Huang, Y. Habitat adaptation drives speciation of a Streptomyces species with distinct habitats and disparate geographic origins. mBio 2022, 13, e0278121. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Hahnke, R.L.; Petersen, J.; Scheuner, C.; Michael, V.; Fiebig, A.; Rohde, C.; Rohde, M.; Fartmann, B.; Goodwin, L.A.; et al. Complete genome sequence of DSM 30083T, the type strain (U5/41T) of Escherichia coli, and a proposal for delineating subspecies in microbial taxonomy. Stand. Genom. Sci. 2014, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Komaki, H.; Ichikawa, N.; Oguchi, A.; Hamada, M.; Harunari, E.; Kodani, S.; Fujita, N.; Igarashi, Y. Draft genome sequence of Streptomyces sp. TP-A0867, an alchivemycin producer. Stand. Genom. Sci. 2016, 11, 85. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.; Ha, S.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Komaki, H.; Igarashi, Y.; Tamura, T. Taxonomic positions of a nyuzenamide-producer and its closely related strains. Microorganisms 2022, 10, 349. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Goker, M. TYGS and LPSN: A database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef] [PubMed]
- Komaki, H.; Ichikawa, N.; Hosoyama, A.; Takahashi-Nakaguchi, A.; Matsuzawa, T.; Suzuki, K.; Fujita, N.; Gonoi, T. Genome based analysis of type-I polyketide synthase and nonribosomal peptide synthetase gene clusters in seven strains of five representative Nocardia species. BMC Genom. 2014, 15, 323. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
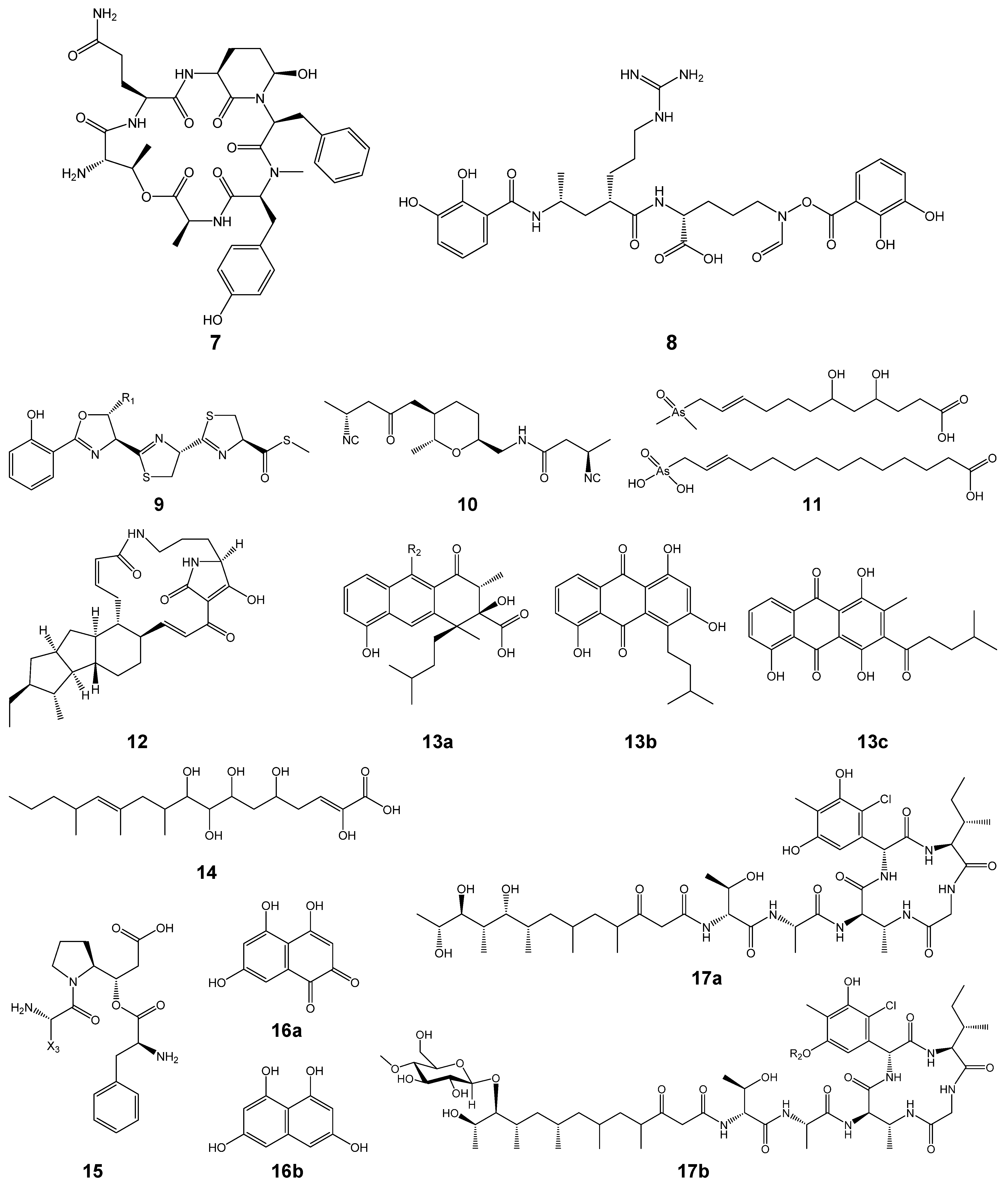{kind=link}
{kind=link}
| Strain | Contig | Genome Size | G+C Content | WGS Project | Sequenced in |
|---|---|---|---|---|---|
| Streptomyces sp. N11-26 | 6 | 8.44 Mb | 72.5% | BNEG01 | This study |
| Streptomyces sp. DC10-5 | 6 | 8.66 Mb | 72.5% | BNEF01 | This study |
| S. olivaceus NRRL B-3009T | 188 | 8.58 Mb | 72.4% | JOFH01 | Reference [14] |
| Strain | DNA–DNA Relatedness | |||
|---|---|---|---|---|
| 1 | 2 | 3 | ||
| 1. Streptomyces sp. N11-26 | - | 80.5% | 88.0% | |
| 2. Streptomyces sp. DC10-5 | 99.9% | - | 80.5% | |
| 3. S. olivaceus NRRL B-3009T | 100% | 99.9% | - | |
| 16S rRNA gene sequence similarity | ||||
| Gene Cluster | Putative Product | S. olivaceus | ||
|---|---|---|---|---|
| N11-26 | DC10-5 | NRRL B-3009T | ||
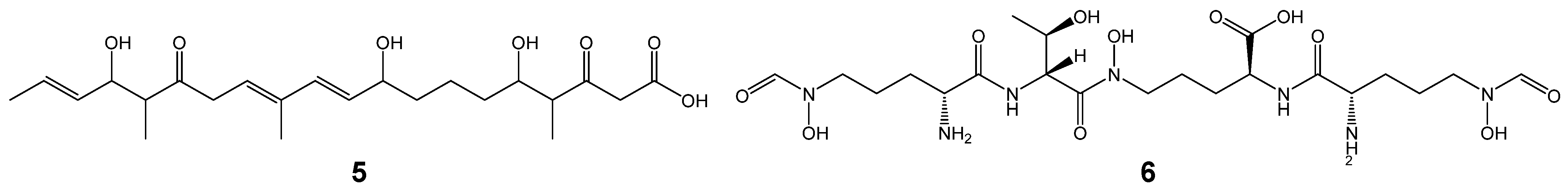
| t1pks-1 | decaketide (5) for alkenyl-franone | + | + | + |
| nrps-1 | Coelichelin (6) | + | + | + |
| nrps-2 * | Streptopeptolin-like, partial (7) | - | + | + |
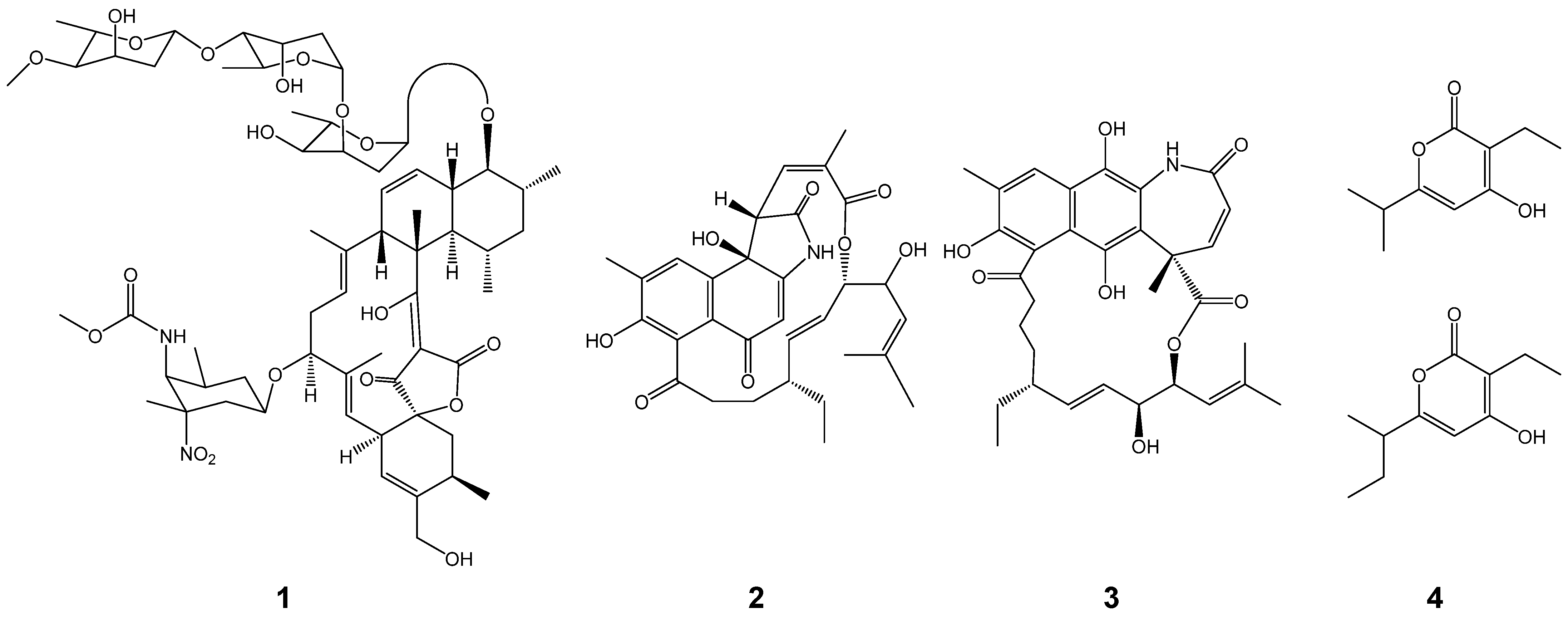
| t1pks-2 | Divergolides (such as 2 and 3 in Figure 1) | + | + | + |
| nrps-3 | Mirubactin (8) | + | + | + |
| nrps-4 | Coelibactin (9) | + | + | + |
| nrps-5 | unknown (with Val) | + | + | + |
| nrps-6 | SF2768 (10) | + | + | + |
| t1pks-3 * | arsono-polyketide (11) | - | + | - |
| pks/nrps-1 | Pactamide (12) | + | + | + |
| t2pks-1 * | Rishirilides (13a), Lupinacidin (13b), and Galvaquinone (13c) | - | + | - |
| nrps-7 | tripeptide (Orn-Orn-Orn) | + | + | + |
| pks/nrps-2 * | unknown (x, mal, Pro) | + | - | + |
| t2pks-2 | spore pigment | + | + | + |
| nrps-8 * | x-y-Gly-Phe | - | + | - |
| t1pks-4 * | Butyrolactol-like, partial (14) | + | - | + |
| pks/nrps-3 | Rimosamide-like, partial (15) | + | + | + |
| t3pks-1 | Flaviolin (16a) and 1,3,6,8-tetrahydroxynaphthalene (16b) | + | + | + |
| t3pks-2 | Germicidins (such as 4 in Figure 1) | + | + | + |
| pks/nrps-4 | Totopotensamides (17a, 17b) | + | + | + |
| t1pks-5 | Lobophorins (such as 1 in Figure 1) | + | + | + |
| Total | 17 | 19 | 18 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komaki, H.; Tamura, T.; Igarashi, Y. Classification and Secondary Metabolite-Biosynthetic Gene Clusters of Marine Streptomyces Strains Including a Lobophorin- and Divergolide-Producer. Hydrobiology 2023, 2, 151-161. https://doi.org/10.3390/hydrobiology2010010
Komaki H, Tamura T, Igarashi Y. Classification and Secondary Metabolite-Biosynthetic Gene Clusters of Marine Streptomyces Strains Including a Lobophorin- and Divergolide-Producer. Hydrobiology. 2023; 2(1):151-161. https://doi.org/10.3390/hydrobiology2010010
Chicago/Turabian StyleKomaki, Hisayuki, Tomohiko Tamura, and Yasuhiro Igarashi. 2023. "Classification and Secondary Metabolite-Biosynthetic Gene Clusters of Marine Streptomyces Strains Including a Lobophorin- and Divergolide-Producer" Hydrobiology 2, no. 1: 151-161. https://doi.org/10.3390/hydrobiology2010010