
Antidepressant Effect of Alpha Lipoic Acid in Rats Exposed to Chronic Unpredictable Mild Stress: Putative Role of Neurotransmitters and 5HT3 Receptor
 , ,
, ,
Abstract
:
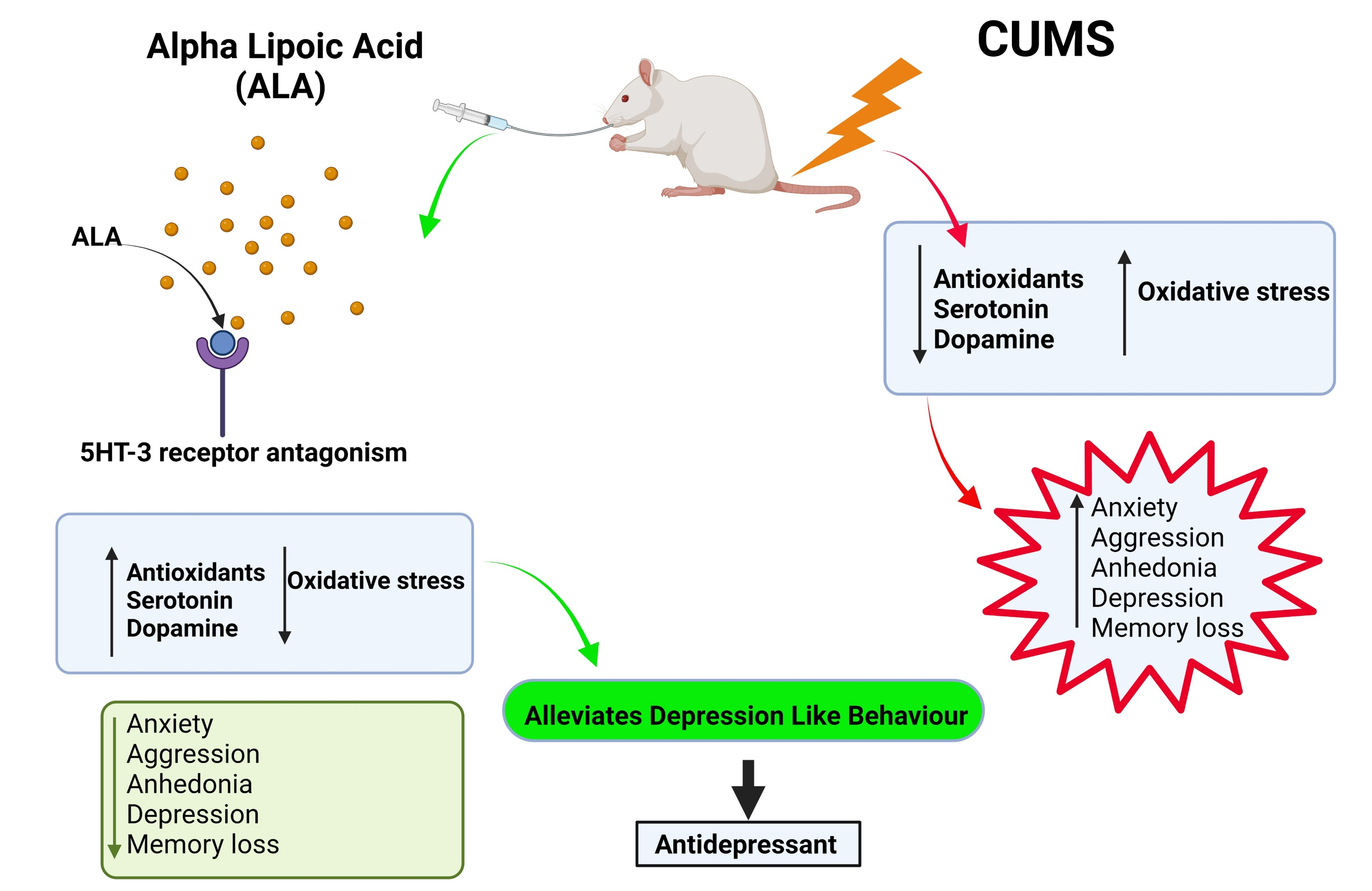
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Drugs and Chemicals
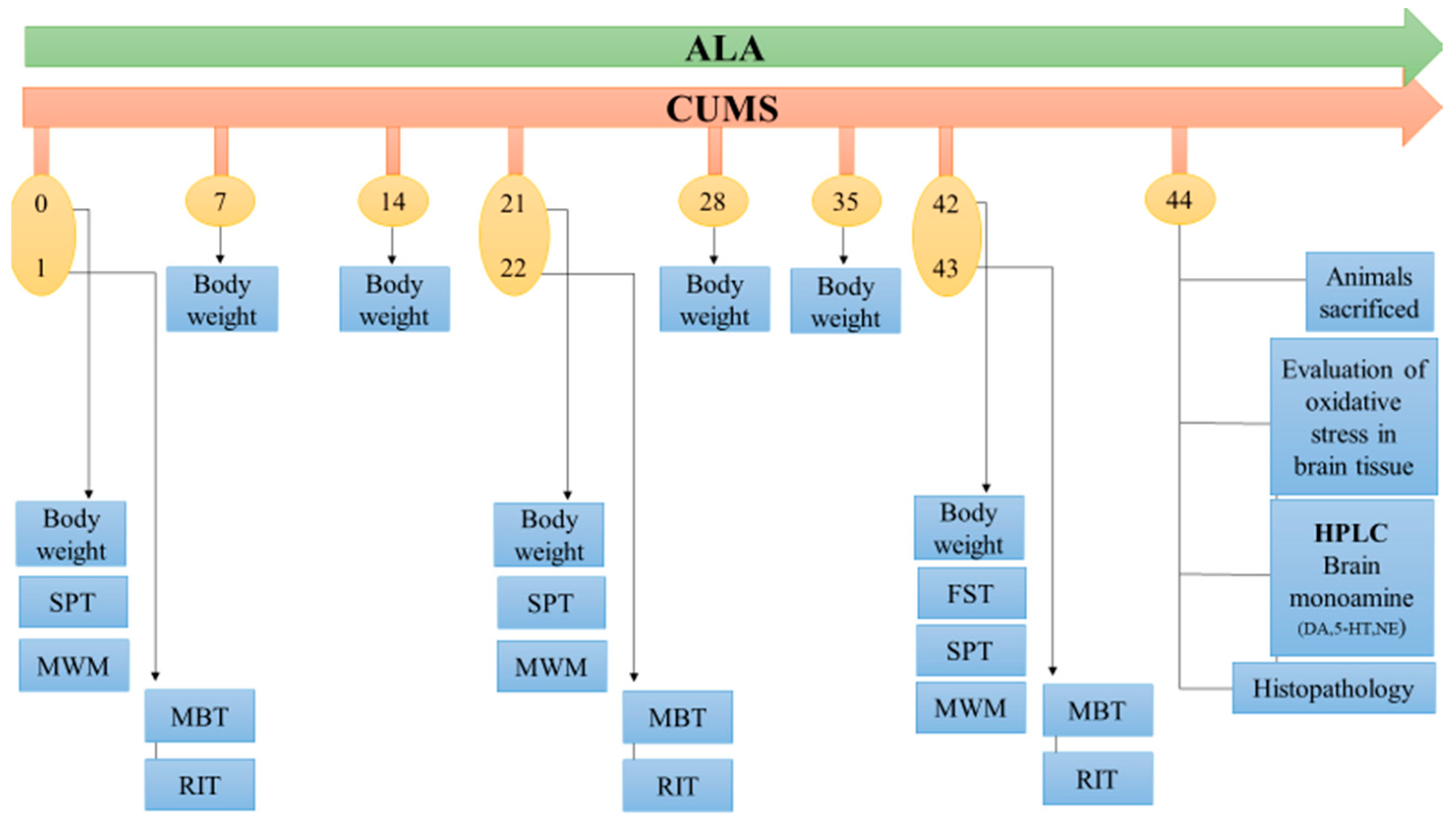
2.3. Experimental design
2.4. CUM-Induced Depression-Like Behaviour in Rats
2.5. Body Weight
2.6. Forced Swim Test (FST)
2.7. Sucrose Preference Test
2.8. Morris Water Maze
2.9. Marble Burying Test (MBT)
2.10. Resident Intruder Test (RIT)
2.11. Brain Tissue Homogenate Preparation for Antioxidant Parameters
2.12. Evaluation of Antioxidant Parameters
2.12.1. Assay of GSH
2.12.2. TBARS Assay
2.12.3. Assay of SOD
2.12.4. Assay of NO
2.12.5. Assay of Catalase
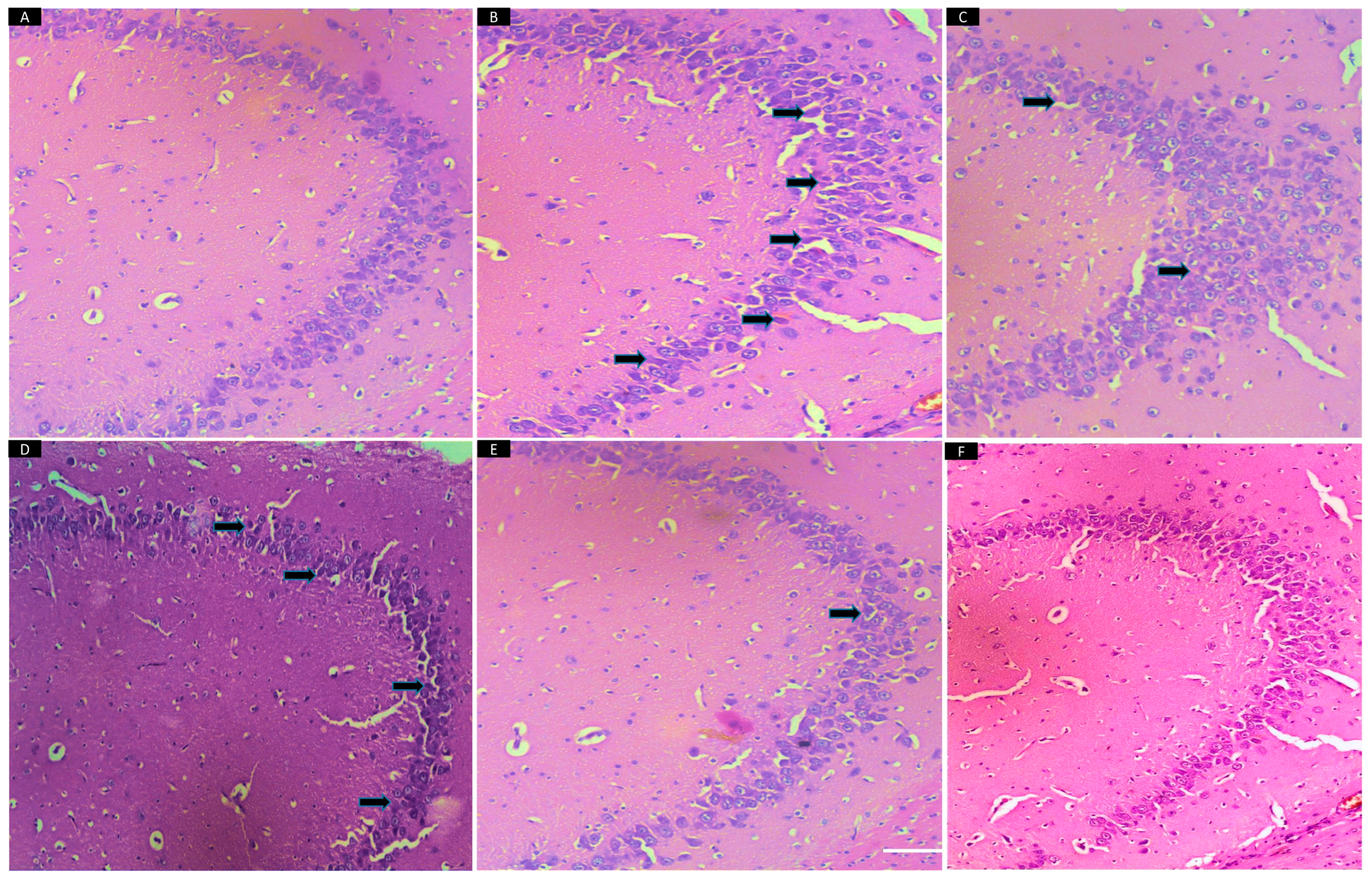
2.13. Histopathological Study of Brain Tissues
2.14. Monoamine’s Estimations
2.14.1. Tissue Preparation
2.14.2. High-Performance Liquid Chromatography (HPLC) Conditions
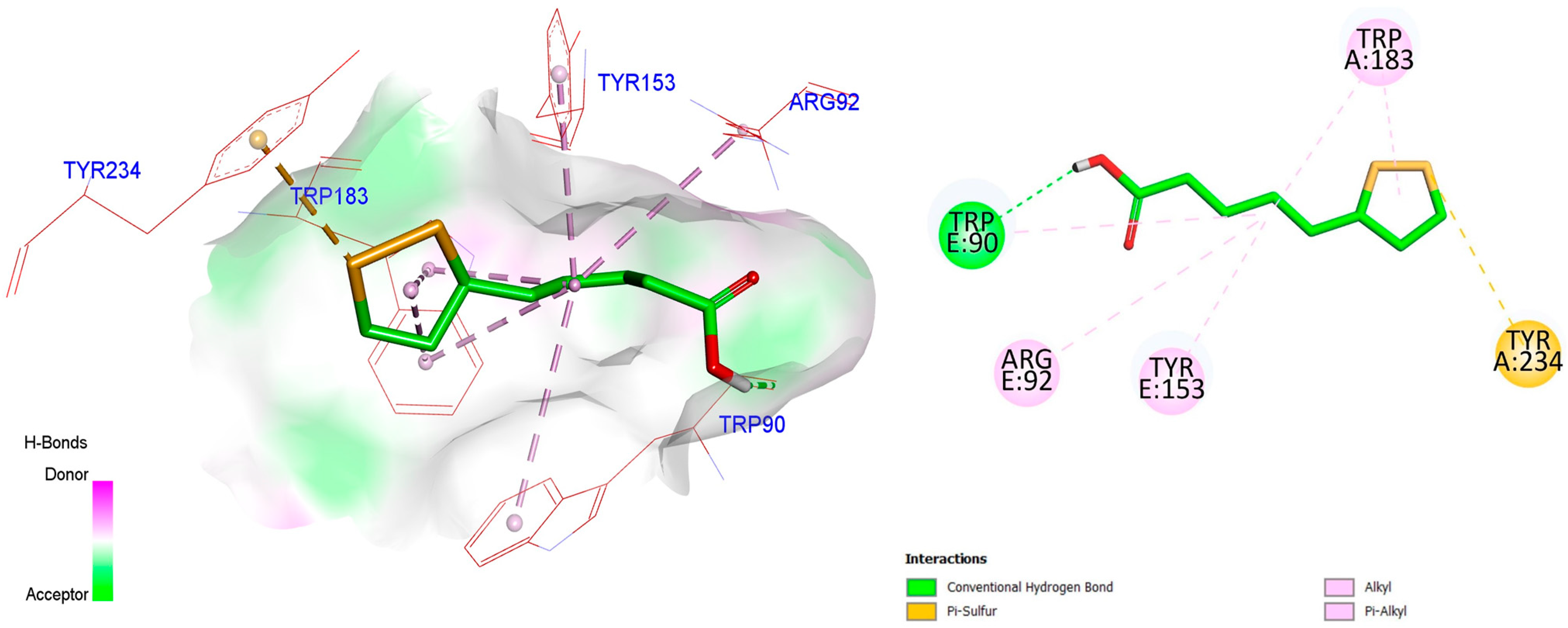
2.15. Molecular Docking Study
2.16. Statistical Analysis
3. Results
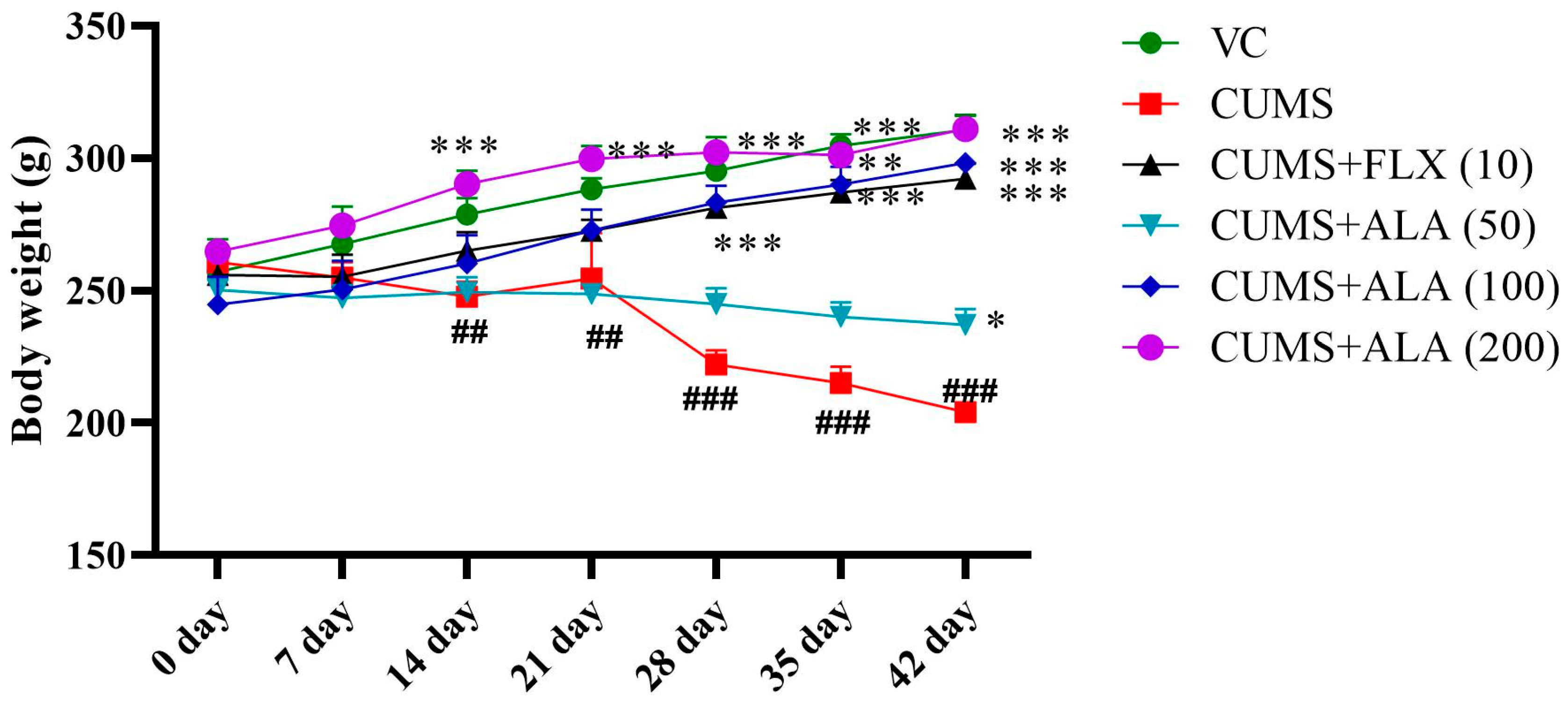
3.1. Effect of ALA (50, 100, 200) and FLX (10 mg/kg) on Body Weight
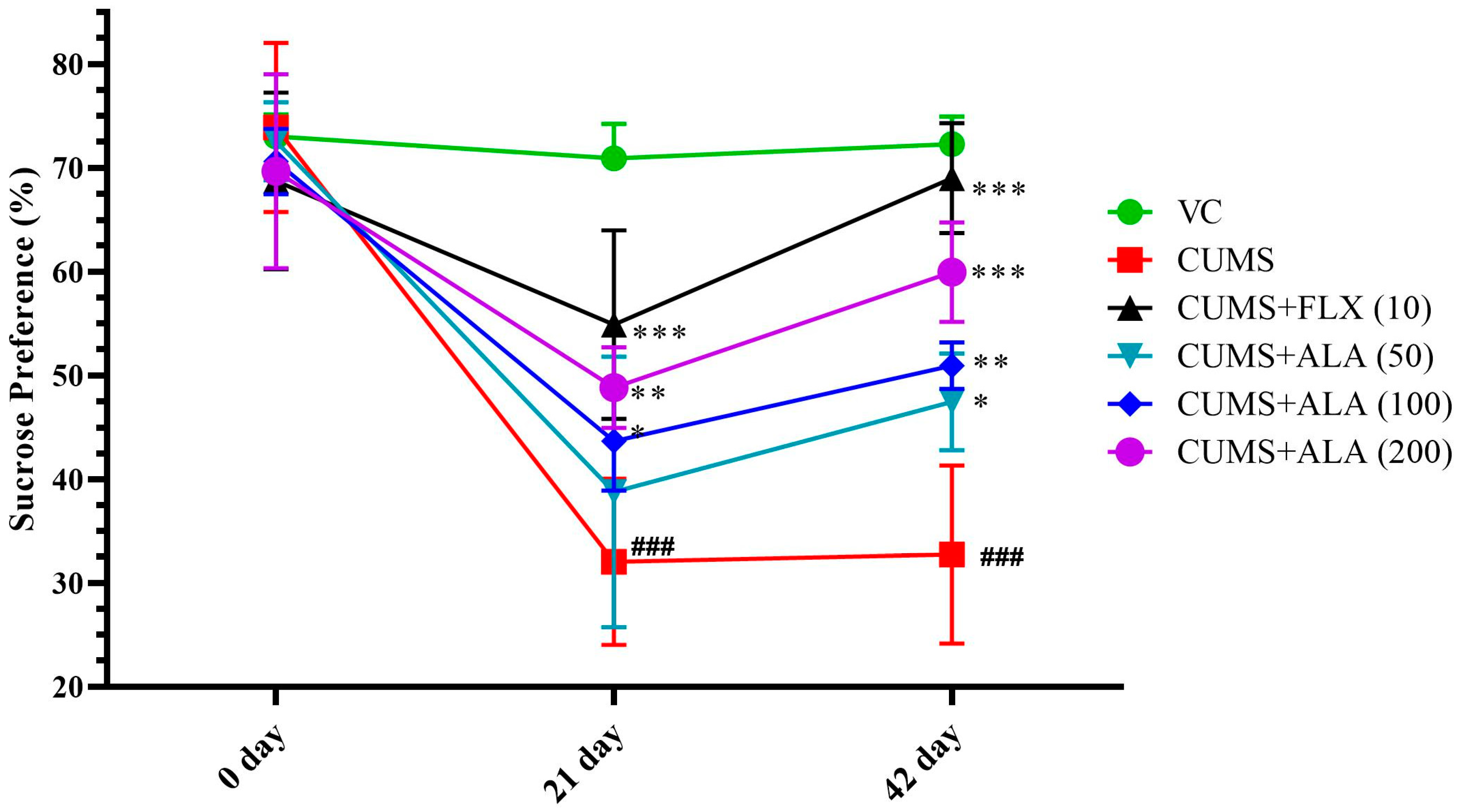
3.2. Effect of ALA (50, 100, 200 mg/kg) and FLX (10 mg/kg) on Sucrose Preference
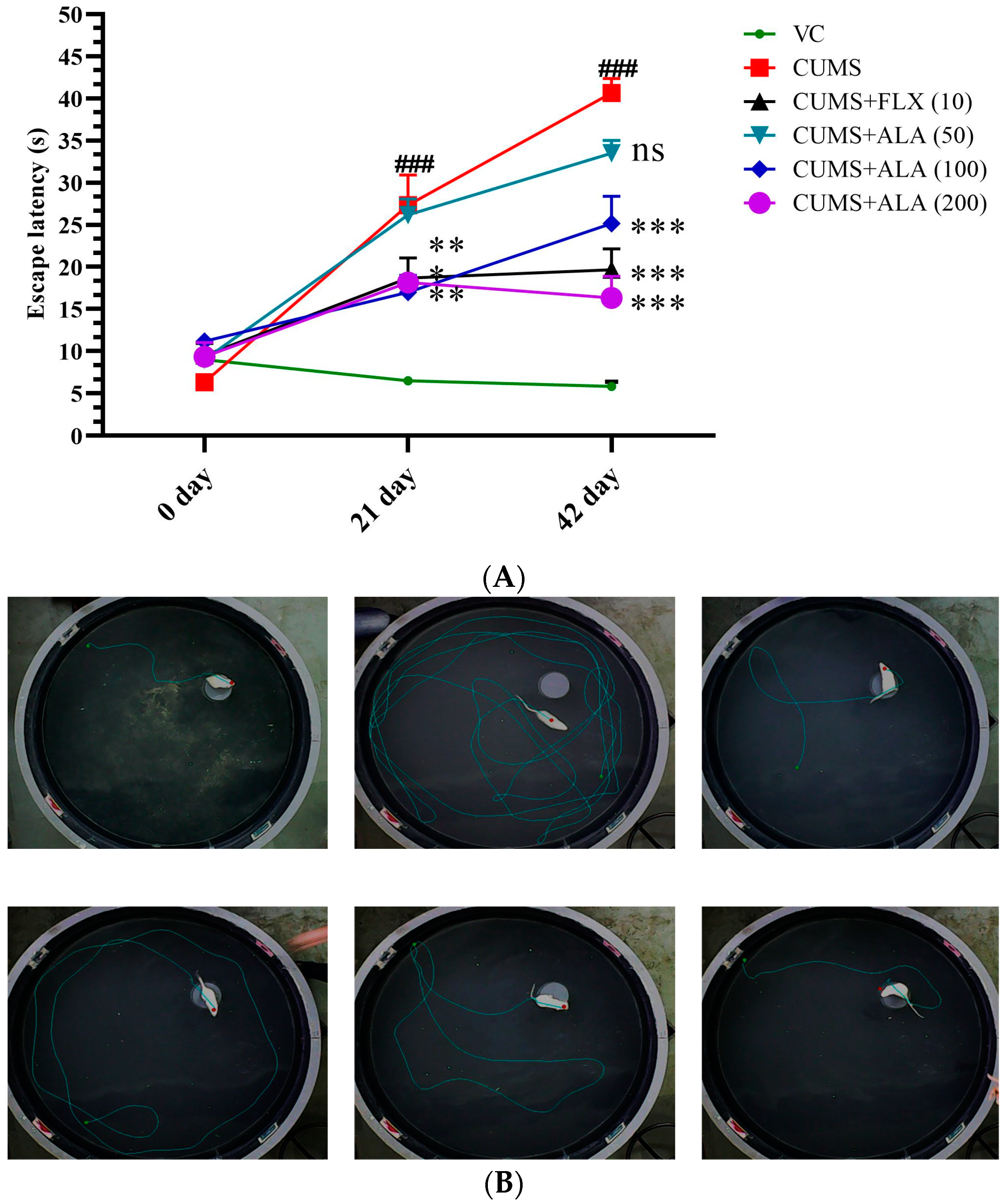
3.3. Effect of ALA (50,100,200 mg/kg) and FLX (10 mg/kg) on Escape Latency, Path Length, and Time Spent away from the Target Zone Using MWM
3.4. Effect of ALA (50,100,200 mg/kg) and FLX (10 mg/kg) on the Number of Marbles Buried Using MBT
3.5. Effect of ALA (50, 100, 200 mg/kg) and FLX (10 mg/kg) on Attack Latency Using RIT
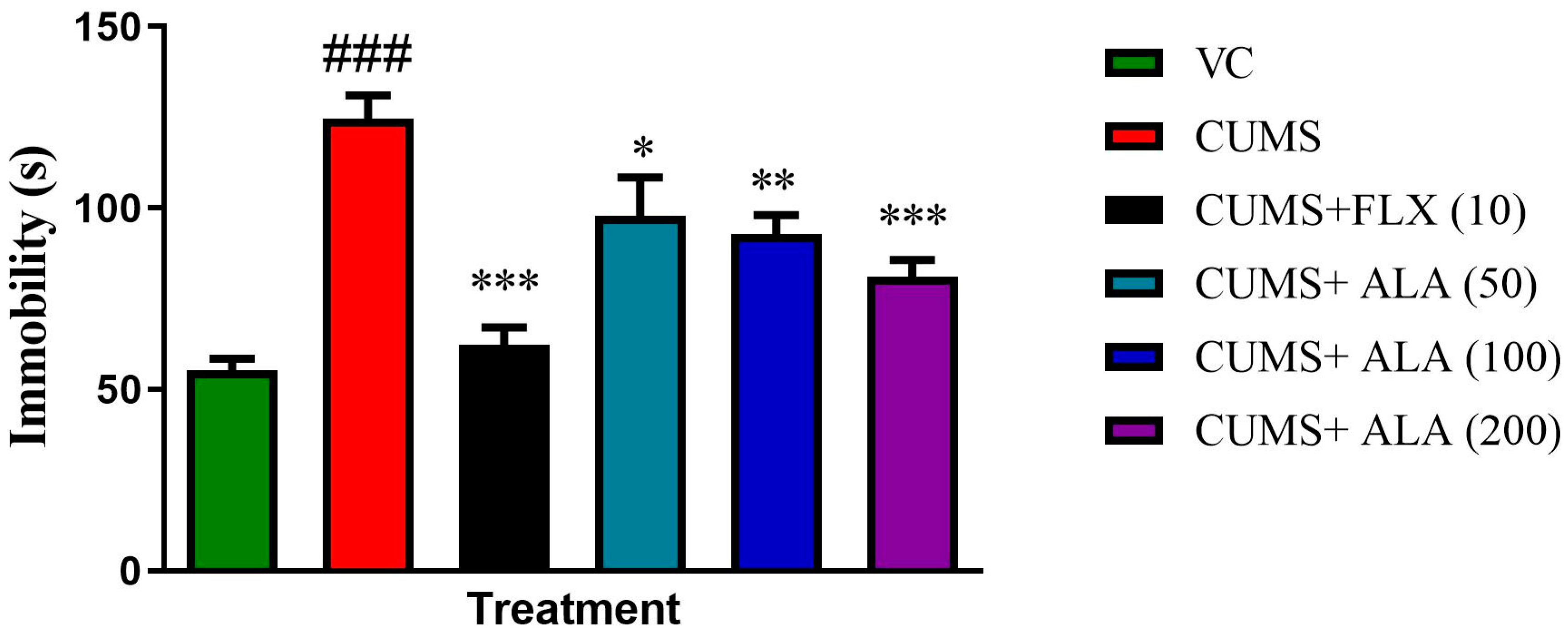
3.6. Effect of ALA (50,100,200 mg/kg) and FLX (10 mg/kg) on Immobility Using FST
3.7. Effect of ALA (50, 100, 200 mg/kg) and FLX (10 mg/kg) on Antioxidant Parameters
3.8. Effect of ALA (50,100,200 mg/kg) and FLX (10 mg/kg) on the Brain Monoamines (DA, NE, 5HT)
3.9. Histopathology
3.10. Molecular Docking Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Perez-Caballero, L.; Torres-Sanchez, S.; Romero-Lopez-Alberca, C.; Gonzalez-Saiz, F.; Mico, J.A.; Berrocoso, E. Monoaminergic system and depression. Cell Tissue Res. 2019, 377, 107–113. [Google Scholar] [CrossRef]
- Xinxing, W.; Wei, L.; Lei, W.; Rui, Z.; Baoying, J.; Lingjia, Q. A neuroendocrine mechanism of co-morbidity of depression-like behavior and myocardial injury in rats. PLoS ONE 2014, 9, e88427. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Guo, J.; Guo, R. Effect of XingPiJieYu decoction on spatial learning and memory and cAMP-PKA-CREB-BDNF pathway in rat model of depression through chronic unpredictable stress. BMC Complement. Altern. Med. 2017, 17, 73. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J.; Barrot, M.; DiLeone, R.J.; Eisch, A.J.; Gold, S.J.; Monteggia, L.M. Neurobiology of depression. Neuron 2002, 34, 13–25. [Google Scholar] [CrossRef]
- Chang, C.H.; Grace, A.A. Amygdala-ventral pallidum pathway decreases dopamine activity after chronic mild stress in rats. Biol. Psychiatry 2014, 76, 223–230. [Google Scholar] [CrossRef]
- Abe, H.; Hidaka, N.; Kawagoe, C.; Odagiri, K.; Watanabe, Y.; Ikeda, T.; Ishizuka, Y.; Hashiguchi, H.; Takeda, R.; Nishimori, T.; et al. Prenatal psychological stress causes higher emotionality, depression-like behavior, and elevated activity in the hypothalamo-pituitary-adrenal axis. Neurosci. Res. 2007, 59, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Patki, G.; Atrooz, F.; Alkadhi, I.; Solanki, N.; Salim, S. High aggression in rats is associated with elevated stress, anxiety-like behavior, and altered catecholamine content in the brain. J. Neurosci. Lett. 2015, 584, 308–313. [Google Scholar] [CrossRef]
- Ou, C.H.; Hall, W.A. Anger in the context of postnatal depression: An integrative review. Birth 2018, 45, 336–346. [Google Scholar] [CrossRef]
- Ma, M.; Chang, X.; Wu, H. Animal models of stress and stress-related neurocircuits: A comprehensive review. Stress Brain 2021, 10, 108–127. [Google Scholar] [CrossRef]
- Ma, Z.X.; Zhang, R.Y.; Rui, W.J.; Wang, Z.Q.; Feng, X. Quercetin alleviates chronic unpredictable mild stress-induced depressive-like behaviors by promoting adult hippocampal neurogenesis via FoxG1/CREB/ BDNF signaling pathway. Behav. Brain Res. 2021, 406, 113245. [Google Scholar] [CrossRef]
- Maes, M.; Fisar, Z.; Medina, M.; Scapagnini, G.; Nowak, G.; Berk, M. New drug targets in depression: Inflammatory, cell-mediated immune, oxidative and nitrosative stress, mitochondrial, antioxidant, and neuroprogressive pathways. And new drug candidates--Nrf2 activators and GSK-3 inhibitors. Inflammopharmacology 2012, 20, 127–150. [Google Scholar] [CrossRef]
- Nandam, L.S.; Brazel, M.; Zhou, M.; Jhaveri, D.J. Cortisol and Major depressive disorder-translating findings from humans to animal models and back. Front. Psychiatry 2020, 10, 974. [Google Scholar] [CrossRef]
- Mikulska, J.; Juszczyk, G.; Gawronska-Grzywacz, M.; Herbet, M. HPA Axis in the Pathomechanism of Depression and Schizophrenia: New Therapeutic Strategies Based on Its Participation. Brain Sci. 2021, 11, 1298. [Google Scholar] [CrossRef]
- Gareri, P.; Falconi, U.; De Fazio, P.; De Sarro, G. Conventional and new antidepressant drugs in the elderly. Prog. Neurobiol. 2000, 61, 353–396. [Google Scholar] [CrossRef]
- Bradberry, S.M.; Thanacoody, H.K.; Watt, B.E.; Thomas, S.H.; Vale, J.A. Management of the cardiovascular complications of tricyclic antidepressant poisoning: Role of sodium bicarbonate. Toxicol. Rev. 2005, 24, 195–204. [Google Scholar] [CrossRef]
- Solmi, M.; Fornaro, M.; Ostinelli, E.G.; Zangani, C.; Croatto, G.; Monaco, F.; Krinitski, D.; Fusar-Poli, P.; Correll, C.U. Safety of 80 antidepressants, antipsychotics, anti-attention-deficit/hyperactivity medications and mood stabilizers in children and adolescents with psychiatric disorders: A large scale systematic meta-review of 78 adverse effects. World Psychiatry 2020, 19, 214–232. [Google Scholar] [CrossRef] [PubMed]
- Rush, A.J. Limitations in efficacy of antidepressant monotherapy. J. Clin. Psychiatry 2007, 68 (Suppl. 10), 8–10. [Google Scholar] [PubMed]
- Fornaro, M.; Anastasia, A.; Valchera, A.; Carano, A.; Orsolini, L.; Vellante, F.; Rapini, G.; Olivieri, L.; Di Natale, S.; Perna, G. The FDA “black box” warning on antidepressant suicide risk in young adults: More harm than benefits? Front. Psychiatry 2019, 10, 294. [Google Scholar] [CrossRef]
- Cuijpers, P.; Quero, S.; Dowrick, C.; Arroll, B. Psychological Treatment of Depression in Primary Care: Recent Developments. Curr. Psychiatry Rep. 2019, 21, 129. [Google Scholar] [CrossRef]
- Shay, K.P.; Moreau, R.F.; Smith, E.J.; Smith, A.R.; Hagen, T.M. Alpha-lipoic acid as a dietary supplement: Molecular mechanisms and therapeutic potential. Biochimica Biophysica Acta (BBA)-Gen. Subj. 2009, 1790, 1149–1160. [Google Scholar] [CrossRef] [PubMed]
- Carrier, B.; Rideout, T.C. Anti-obesity and lipid-lowering properties of alpha-lipoic acid. Food Sci. Nutr. 2013, 1, 1002. [Google Scholar]
- Pal, S.N.; Dandiya, P. Glutathione as a cerebral substrate in depressive behavior. Pharmacol. Biochem. Behav. 1994, 48, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Karunakaran, U.; Jeoung, N.H.; Jeon, J.H.; Lee, I.K. Physiological effect and therapeutic application of alpha lipoic acid. Curr. Med. Chem. 2014, 21, 3636–3645. [Google Scholar] [CrossRef]
- Saha, M.; Ahammad, H.; Bhoumik, N.C.; Shakil, M.S.; Shawan, M.; Morshed, M.; Hossan, T.; Sarker, S.R.; Rahman, M.N.; Rahman, S.B. Extraction and estimation of alpha lipoic acid content in different food samples by reverse phase HPLC: Effect of heat treatment. Int. J. Biosci. 2018, 13, 473–482. [Google Scholar]
- Badran, M.; Abuyassin, B.; Golbidi, S.; Ayas, N.; Laher, I. Alpha Lipoic Acid Improves Endothelial Function and Oxidative Stress in Mice Exposed to Chronic Intermittent Hypoxia. Oxid. Med. Cell. Longev. 2019, 2019, 4093018. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, C.N.; Meneses, L.N.; Vasconcelos, G.S.; Silva, M.C.; da Silva, J.C.; Macedo, D.; de Lucena, D.F.; Vasconcelos, S.M. Reversal of corticosterone-induced BDNF alterations by the natural antioxidant alpha-lipoic acid alone and combined with desvenlafaxine: Emphasis on the neurotrophic hypothesis of depression. Psychiatry Res. 2015, 230, 211–219. [Google Scholar] [CrossRef]
- Wei, W.; Wang, H.; Wu, Y.; Ding, K.; Li, T.; Cong, Z.; Xu, J.; Zhou, M.; Huang, L.; Ding, H.; et al. Alpha lipoic acid inhibits neural apoptosis via a mitochondrial pathway in rats following traumatic brain injury. Neurochem. Int. 2015, 87, 85–91. [Google Scholar] [CrossRef]
- Poon, H.F.; Farr, S.A.; Thongboonkerd, V.; Lynn, B.C.; Banks, W.A.; Morley, J.E.; Klein, J.B.; Butterfield, D.A. Proteomic analysis of specific brain proteins in aged SAMP8 mice treated with alpha-lipoic acid: Implications for aging and age-related neurodegenerative disorders. Neurochem. Int. 2005, 46, 159–168. [Google Scholar] [CrossRef]
- Macedo, D.S.; Medeiros, C.D.; Cordeiro, R.C.; Sousa, F.C.; Santos, J.V.; Morais, T.A.; Hyphantis, T.N.; McIntyre, R.S.; Quevedo, J.; Carvalho, A.F. Effects of alpha-lipoic acid in an animal model of mania induced by D-amphetamine. Bipolar Disord. 2012, 14, 707–718. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Gupta, Y. Effect of alpha lipoic acid on intracerebroventricular streptozotocin model of cognitive impairment in rats. Eur. Neuropsychopharmacol. 2003, 13, 241–247. [Google Scholar] [CrossRef]
- Li, S.; Wang, C.; Wang, W.; Dong, H.; Hou, P.; Tang, Y. Chronic mild stress impairs cognition in mice: From brain homeostasis to behavior. Life Sci. 2008, 82, 934–942. [Google Scholar] [CrossRef]
- Harro, J.; Haidkind, R.; Harro, M.; Modiri, A.R.; Gillberg, P.G.; Pahkla, R.; Matto, V.; Oreland, L. Chronic mild unpredictable stress after noradrenergic denervation: Attenuation of behavioural and biochemical effects of DSP-4 treatment. Eur. Neuropsychopharmacol. 1999, 10, 5–16. [Google Scholar] [CrossRef]
- Castagne, V.; Porsolt, R.D.; Moser, P. Use of latency to immobility improves detection of antidepressant-like activity in the behavioral despair test in the mouse. Eur. J. Pharmacol. 2009, 616, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Aswar, U.; Chepurwar, S.; Shintre, S.; Aswar, M. Telmisartan attenuates diabetes induced depression in rats. Pharmacol. Rep. 2017, 69, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Willner, P.; Scheel-Kruger, J.; Belzung, C. The neurobiology of depression and antidepressant action. Neurosci. Biobehav. Rev. 2013, 37, 2331–2371. [Google Scholar] [CrossRef]
- D’Hooge, R.; De Deyn, P.P. Applications of the Morris water maze in the study of learning and memory. Brain Res. 2001, 36, 60–90. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Sripanidkulchai, K.; Wyss, J.M.; Sripanidkulchai, B. Curcuma comosa improves learning and memory function on ovariectomized rats in a long-term Morris water maze test. J. Ethnopharmacol. 2010, 130, 70–75. [Google Scholar] [CrossRef]
- Handley, S.L. Evaluation of marble-burying behavior as a model of anxiety. Pharmacol. Biochem. Behav. 1991, 38, 63–67. [Google Scholar]
- Wei, S.; Zhang, H.; Gao, J.; Xue, L.; Sun, P.; Chao, Y.; Xue, G.; Qiao, M. Impact of social isolation and resident intruder stress on aggressive behavior in the male rat. Neural Regen Res. 2010, 5, 1175–1179. [Google Scholar]
- Moron, M.; Depierre, J.; Mannervik, B. Biochim Biophys Acta. Gen. Subj. 1979, 582, 67–78. [Google Scholar] [CrossRef]
- Garcia, L.S.; Comim, C.M.; Valvassori, S.S.; Reus, G.Z.; Barbosa, L.M.; Andreazza, A.C.; Stertz, L.; Fries, G.R.; Gavioli, E.C.; Kapczinski, F.; et al. Acute administration of ketamine induces antidepressant-like effects in the forced swimming test and increases BDNF levels in the rat hippocampus. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 140–144. [Google Scholar] [CrossRef]
- Sun, Y.J. A simple method for clinical assay of superoxide dismutase. Clin. Chem. 1988, 34, 497–500. [Google Scholar] [CrossRef]
- Giustarini, D.; Rossi, R.; Milzani, A.; Dalle-Donne, I. Nitrite and nitrate measurement by Griess reagent in human plasma: Evaluation of interferences and standardization. Methods Enzymol. 2008, 440, 361–380. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.K. Colorimetric assay of catalase. Anal. Biochem. 1972, 47, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Ayuob, N.N.; Balgoon, M.J. Histological and molecular techniques utilized to investigate animal models of depression. An updated review. Microsc. Res. Tech. 2018, 81, 1143–1153. [Google Scholar] [CrossRef]
- Thorré, K.; Pravda, M.; Sarre, S.; Ebinger, G.; Michotte, Y. New antioxidant mixture for long term stability of serotonin, dopamine and their metabolites in automated microbore liquid chromatography with dual electrochemical detection. J. Chromatogr. B Biomed. Sci. Appl. 1997, 694, 297–303. [Google Scholar] [CrossRef] [PubMed]
- De Benedetto, G.E.; Fico, D.; Pennetta, A.; Malitesta, C.; Nicolardi, G.; Lofrumento, D.D.; De Nuccio, F.; La Pesa, V. A rapid and simple method for the determination of 3,4-dihydroxyphenylacetic acid, norepinephrine, dopamine, and serotonin in mouse brain homogenate by HPLC with fluorimetric detection. J. Pharm. Biomed. Anal. 2014, 98, 266–270. [Google Scholar] [CrossRef]
- Bétry, C.; Etiévant, A.; Oosterhof, C.; Ebert, B.; Sanchez, C.; Haddjeri, N. Role of 5-HT3 receptors in the antidepressant response. Pharmaceuticals 2011, 4, 603–629. [Google Scholar] [CrossRef]
- Jiang, N.; Zhang, B.Y.; Dong, L.M.; Lv, J.W.; Lu, C.; Wang, Q.; Fan, L.X.; Zhang, H.X.; Pan, R.L.; Liu, X.M. Antidepressant effects of dammarane sapogenins in chronic unpredictable mild stress-induced depressive mice. Phytother. Res. 2018, 32, 1023–1029. [Google Scholar] [CrossRef]
- Abdul Shukkoor, M.S.; Baharuldin, M.T.; Mat Jais, A.M.; Mohamad Moklas, M.A.; Fakurazi, S. Antidepressant-Like Effect of Lipid Extract of Channa striatus in Chronic Unpredictable Mild Stress Model of Depression in Rats. Evid. Based Complement. Alternat. Med. 2016, 2016, 2986090. [Google Scholar] [CrossRef]
- Kim, M.-S.; Park, J.-Y.; Namkoong, C.; Jang, P.-G.; Ryu, J.-W.; Song, H.-S.; Yun, J.-Y.; Namgoong, I.-S.; Ha, J.; Park, I.-S. Anti-obesity effects of α-lipoic acid mediated by suppression of hypothalamic AMP-activated protein kinase. Nat. Med. 2004, 10, 727–733. [Google Scholar] [CrossRef]
- Pratt, W.E.; Lin, P.; Pierce-Messick, Z.; Ilesanmi, A.O.; Clissold, K.A. Contrasting effects of 5-HT3 receptor stimulation of the nucleus accumbens or ventral tegmentum on food intake in the rat. Behav. Brain Res. 2017, 323, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Stanton, C.H.; Holmes, A.J.; Chang, S.W.; Joormann, J. From stress to anhedonia: Molecular processes through functional circuits. Trends Neurosci. 2019, 42, 23–42. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Zhang, X.; Xing, N.; Qu, H.; Zhang, K. Uncovering pharmacological mechanisms of Zhi-Zi-Hou-Po decoction in chronic unpredictable mild stress induced rats through pharmacokinetics, monoamine neurotransmitter and neurogenesis. J. Ethnopharmacol. 2019, 243, 112079. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.; He, B.; Wan, S.; Xu, M.; Yang, H.; Xiao, F.; Bi, K.; Jia, Y. Antidepressant-like effects and cognitive enhancement of Schisandra chinensis in chronic unpredictable mild stress mice and its related mechanism. Sci. Rep. 2017, 7, 6903. [Google Scholar] [CrossRef]
- Huang, H.J.; Zhu, X.C.; Han, Q.Q.; Wang, Y.L.; Yue, N.; Wang, J.; Yu, R.; Li, B.; Wu, G.C.; Liu, Q.; et al. Ghrelin alleviates anxiety- and depression-like behaviors induced by chronic unpredictable mild stress in rodents. Behav. Brain Res. 2017, 326, 33–43. [Google Scholar] [CrossRef]
- Aswar, U.; Shende, H.; Aswar, M. Buspirone, a 5-HT1A agonist attenuates social isolation-induced behavior deficits in rats: A comparative study with fluoxetine. Behav. Pharmacol. 2022, 33, 309–321. [Google Scholar] [CrossRef]
- Yang, C.R.; Bai, Y.Y.; Ruan, C.S.; Zhou, H.F.; Liu, D.; Wang, X.F.; Shen, L.J.; Zheng, H.Y.; Zhou, X.F. Enhanced aggressive behaviour in a mouse model of depression. Neurotox. Res. 2015, 27, 129–142. [Google Scholar] [CrossRef]
- Salazar, M.R. Alpha lipoic acid: A novel treatment for depression. Med. Hypotheses 2000, 55, 510–512. [Google Scholar] [CrossRef]
- de Sousa, C.N.S.; Meneses, L.N.; Vasconcelos, G.S.; da Silva Medeiros, I.; Silva, M.C.C.; Mouaffak, F.; Kebir, O.; da Silva Leite, C.M.G.; Patrocinio, M.C.A.; Macedo, D.; et al. Neuroprotective evidence of alpha-lipoic acid and desvenlafaxine on memory deficit in a neuroendocrine model of depression. Naunyn Schmiedebergs Arch. Pharmacol. 2018, 391, 803–817. [Google Scholar] [CrossRef]
- Lackovic, M.; Rovcanin, B.; Pantovic, M.; LvkovIc, M.; Petronijevic, N.; Damjanovic, A. Association of oxidative stress with the pathophysiology of depresion and bipolar disorder. Arch. Biol. Sci. 2013, 65, 369–373. [Google Scholar]
- Correia, A.S.; Cardoso, A.; Vale, N. Oxidative Stress in Depression: The Link with the Stress Response, Neuroinflammation, Serotonin, Neurogenesis and Synaptic Plasticity. Antioxidants 2023, 12, 470. [Google Scholar] [CrossRef]
- Li, X.; Wu, T.; Yu, Z.; Li, T.; Zhang, J.; Zhang, Z.; Cai, M.; Zhang, W.; Xiang, J.; Cai, D. Apocynum venetum leaf extract reverses depressive-like behaviors in chronically stressed rats by inhibiting oxidative stress and apoptosis. Biomed. Pharmacother. 2018, 100, 394–406. [Google Scholar] [CrossRef]
- Yusha’u, Y.; Muhammad, U.; Mustapha, S.; Umar, A.; Imam, M.; Umar, B.; Alhassan, A.; Saleh, M.; Ya’u, J. Alpha-lipoic acid attenuates depressive symptoms in mice exposed to chronic unpredictable mild stress. J. Afr. Assoc. Physiol. Sci. 2021, 9, 58–68. [Google Scholar]
- Yusha’u, Y.; Adam, U.M.; Wahab, A.A.; Saleh, M.I.A.; Ya’u, J. Alpha-lipoic acid enhances short-term spatial memory of mice in open-space forced swim-induced depression mouse model. Neurosci. Res. Notes 2021, 4, 36–50. [Google Scholar] [CrossRef]
- Bergstrom, A.; Jayatissa, M.N.; Mork, A.; Wiborg, O. Stress sensitivity and resilience in the chronic mild stress rat model of depression; an in situ hybridization study. Brain Res. 2008, 1196, 41–52. [Google Scholar] [CrossRef]
- Lesch, K.-P.; Araragi, N.; Waider, J.; van den Hove, D.; Gutknecht, L. Targeting brain serotonin synthesis: Insights into neurodevelopmental disorders with long-term outcomes related to negative emotionality, aggression and antisocial behaviour. Philos. Biol. Sci. 2012, 367, 2426–2443. [Google Scholar] [CrossRef]
- Hong, L.Z.; Huang, K.F.; Hung, S.W.; Kuo, L.T. Chronic fluoxetine treatment enhances sympathetic activities associated with abnormality of baroreflex function in conscious normal rats. Eur. J. Pharmacol. 2017, 811, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.; Devadoss, T.; Manjula, S.N.; Rajangam, J. 5-HT3 receptor antagonism: A potential therapeutic approach for the treatment of depression and other disorders. Curr. Neuropharmacol. 2021, 19, 1545. [Google Scholar] [CrossRef]
- Bravo, G.; Maswood, S. Acute treatment with 5-HT3 receptor antagonist, tropisetron, reduces immobility in intact female rats exposed to the forced swim test. Pharmacol. Biochem. Behav. 2006, 85, 362–368. [Google Scholar] [CrossRef]
- Gupta, D.; Radhakrishnan, M.; Kurhe, Y.; Thangaraj, D.; Prabhakar, V.; Kanade, P. Antidepressant-like effects of a novel 5-HT3 receptor antagonist 6z in acute and chronic murine models of depression. Acta Pharmacol. Sin. 2014, 35, 1493–1503. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Parameters | Days | VC | CUMS | CUMS + FLX (10) | CUMS + ALA (50) | CUMS + ALA (100) | CUMS + ALA (200) |
|---|---|---|---|---|---|---|---|
| MWM—Path length | |||||||
| Distance traveled (cm) | day 0 | 150 ± 13 | 137 ± 13 | 142 ± 11 | 182 ± 13 | 174 ± 29 | 145 ± 44 |
| day 21 | 130 ± 15 | 410 ± 72 ## | 360 ± 36 ** | 380 ± 15 | 350 ± 77 *** | 250 ± 76 ** | |
| day 42 | 105 ± 49 | 630 ± 71 ### | 275 ± 23 *** | 510 ± 13* | 234 ± 72 *** | 180 ± 118 *** | |
| MWM—Time spent away from target zone | |||||||
| Time (s) | day 0 | 4.0 ± 0.1 | 3 ± 0.3 | 5 ± 1 | 5 ± 0.5 | 4 ± 0.3 | 4 ± 0.9 |
| day 21 | 1.8 ± 0.1 | 24 ± 6 ### | 4 ± 0.6 *** | 19 ± 4 | 9 ± 1 *** | 12 ± 2 ** | |
| day 42 | 6.1 ± 0.3 | 33 ± 4 ### | 11 ± 1 *** | 21 ± 3 ** | 20 ± 11 *** | 11 ± 2 *** | |
| MBT Number of marbles buried | day 0 | 2 ± 0.5 | 3 ± 0.8 | 3 ± 0.4 | 2 ± 0.3 | 2 ± 0.7 | 2 ± 0.3 |
| day 21 | 2 ± 0.5 | 8 ± 0.8 ### | 3 ± 0.8 *** | 6 ± 0.4 | 6 ± 0.8 * | 6 ± 1 * | |
| day 42 | 1 ± 0.3 | 10 ± 0.7 ### | 3 ± 0.4 *** | 6 ± 0.6 ** | 6 ± 1 ** | 5 ± 0.5 *** | |
| RIT Attack latency (s) | day 0 | 272 ± 17 | 235 ± 16 | 231 ± 21 | 235 ± 0 | 231 ± 0 | 249 ± 03 |
| day 21 | 260 ± 30 | 138 ± 17 ## | 193 ± 22 ** | 138 ± 14 | 193 ± 10 * | 228 ± 17 * | |
| day 42 | 267 ± 26 | 82 ± 5 ### | 239 ± 14 ** | 82 ± 15 | 239 ± 19 ** | 260 ± 12 *** | |
| Antioxidant parameters | |||||||
| GSH (ng/g tissue weight) | 201 ± 14.6 | 121 ± 12.1 ### | 182.3 ± 8.4 ** | 136.4 ± 9.50 | 182.1 ±7.96 ** | 232.8 ± 9.27*** | |
| CAT (μg/g of tissue weight) | 0.25 ± 0.03 | 0.05 ± 0.01 ### | 0.14 ± 0.01 ** | 0.06 ± 0.01 | 0.13 ± 0.01 ** | 0.17 ± 0.01 *** | |
| SOD (U/g tissue weight) | 61.58 ± 0.92 | 20.35 ± 2.92 ### | 39.85 ± 5.46 *** | 16.59 ± 1.47 | 23.18 ± 2.69 | 33.26 ± 2.79 * | |
| NO (μg/g of tissue weight) | 356.8 ± 26.88 | 1291 ± 27.33 ### | 420.5 ± 24.62 *** | 1125 ± 56.58 | 1038 ± 56.10 * | 709.4 ± 20.75 *** | |
| MDA (nM/g tissue weight) | 207.3 ± 4.77 | 276.3 ± 12.34 ## | 205.3 ± 4.92 ** | 256.9 ± 13.72 | 193.1 ± 19.03 *** | 209.3 ± 8.53 ** | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akotkar, L.; Aswar, U.; Patil, R.; Kumar, D.; Aswar, M.; Pandey, J.; Gurav, S. Antidepressant Effect of Alpha Lipoic Acid in Rats Exposed to Chronic Unpredictable Mild Stress: Putative Role of Neurotransmitters and 5HT3 Receptor. Future Pharmacol. 2023, 3, 407-425. https://doi.org/10.3390/futurepharmacol3020025
Akotkar L, Aswar U, Patil R, Kumar D, Aswar M, Pandey J, Gurav S. Antidepressant Effect of Alpha Lipoic Acid in Rats Exposed to Chronic Unpredictable Mild Stress: Putative Role of Neurotransmitters and 5HT3 Receptor. Future Pharmacology. 2023; 3(2):407-425. https://doi.org/10.3390/futurepharmacol3020025
Chicago/Turabian StyleAkotkar, Likhit, Urmila Aswar, Rashmi Patil, Dileep Kumar, Manoj Aswar, Jyoti Pandey, and Shailendra Gurav. 2023. "Antidepressant Effect of Alpha Lipoic Acid in Rats Exposed to Chronic Unpredictable Mild Stress: Putative Role of Neurotransmitters and 5HT3 Receptor" Future Pharmacology 3, no. 2: 407-425. https://doi.org/10.3390/futurepharmacol3020025