
Flow Cytometry of Oxygen and Oxygen-Related Cellular Stress

Abstract
:1. Introduction: Oxygen, ROS and Oxidative Stress
2. Flow Cytometry as a Tool for Functional Cell Research
2.1. Specific Features and Limitations of Functional FCM
2.1.1. Multiparametric Data Acquisition
2.1.2. Multivariate Data Analysis
2.1.3. Fast Analysis of Large Number of Live Cells
2.1.4. Real-Time Flow Cytometry
2.1.5. Individual Cell Sorting
2.1.6. Limitations of Functional Flow Cytometry
3. General Strategies in Flow Cytometric Analysis of Oxygen and Oxidative Stress
- Performing cell-based studies in hypoxic conditions:
- Monitoring intracellular oxygen in hypoxic conditions:
- Direct detection of ROS, the initiators of the oxidative stress process:
- Detection of more stable oxidized end products:
- Assessment of antioxidant defenses, mostly GSH and SH-containing proteins:
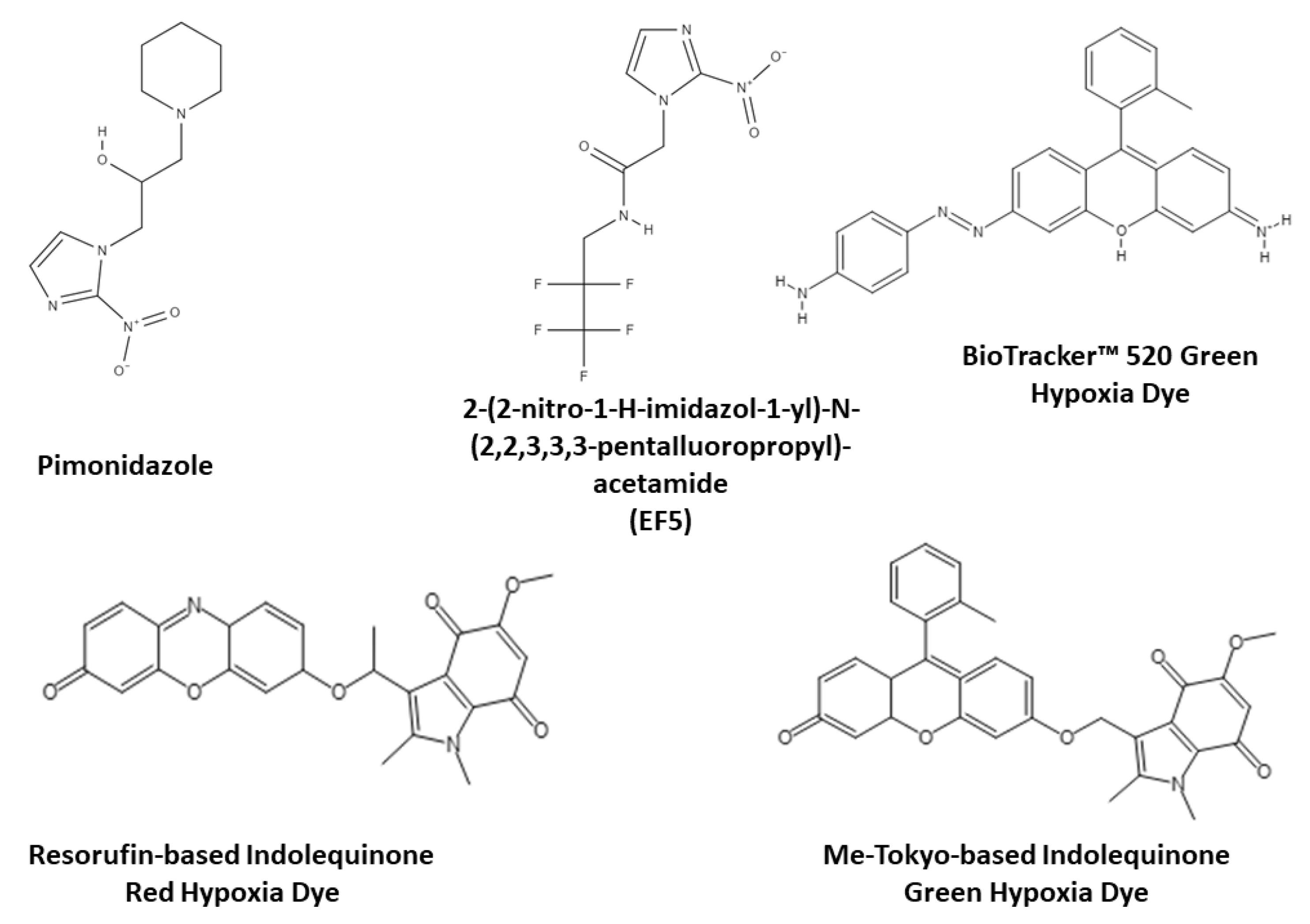
3.1. Monitoring Intracellular Oxygen in Hypoxic Conditions
3.2. Direct Detection of ROS with Fluorogenic Substrates
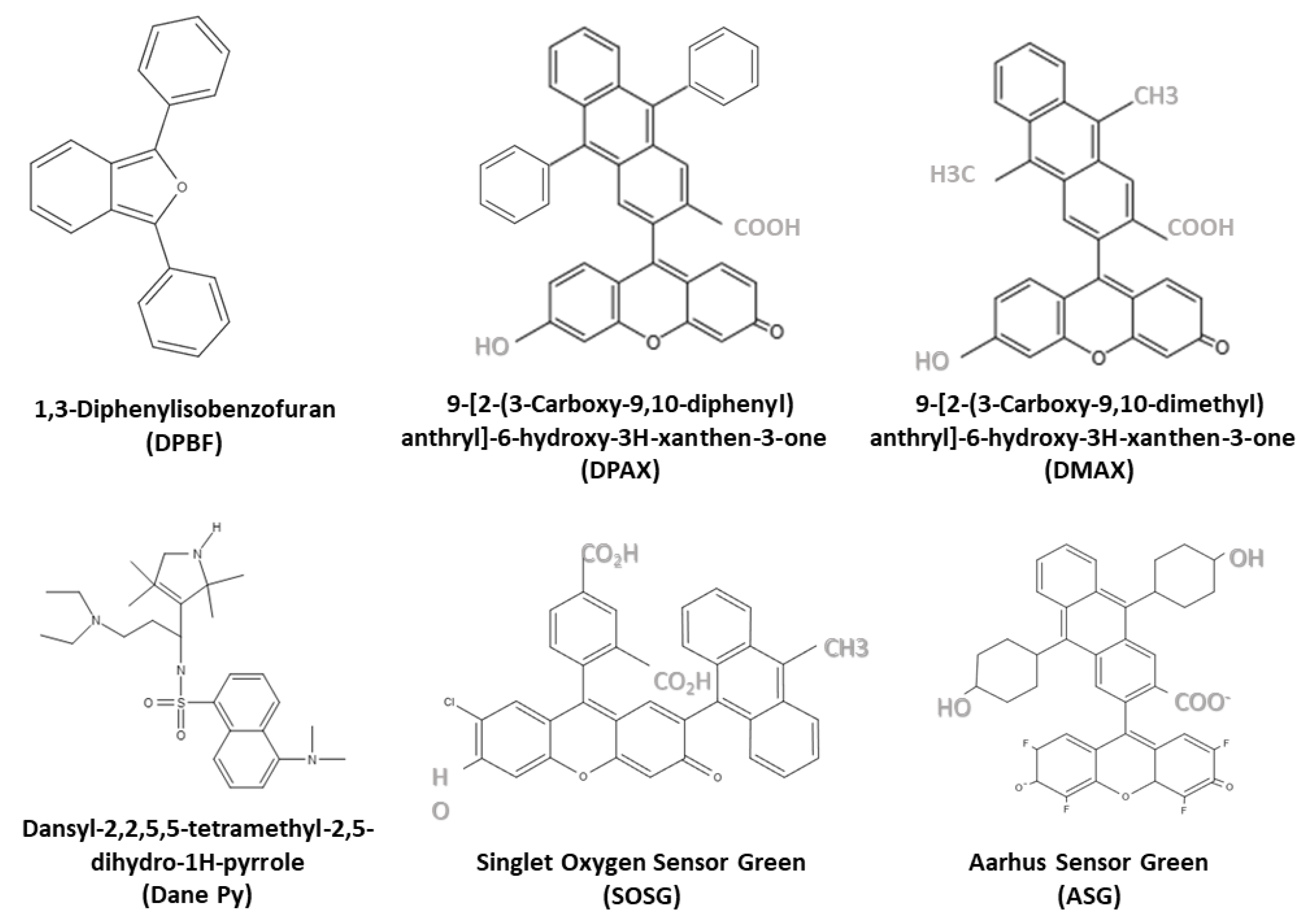
3.2.1. 1O2 Probes
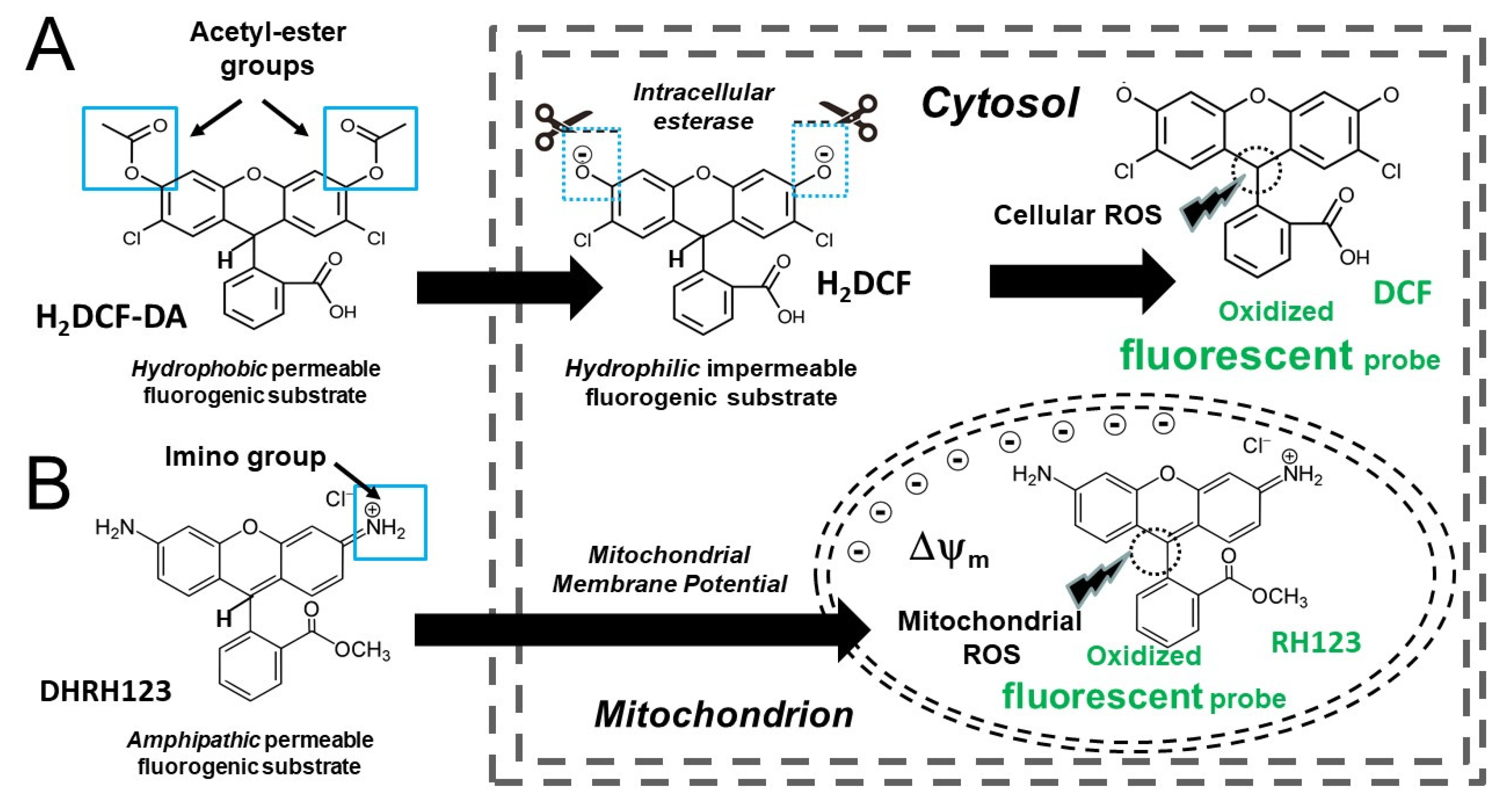
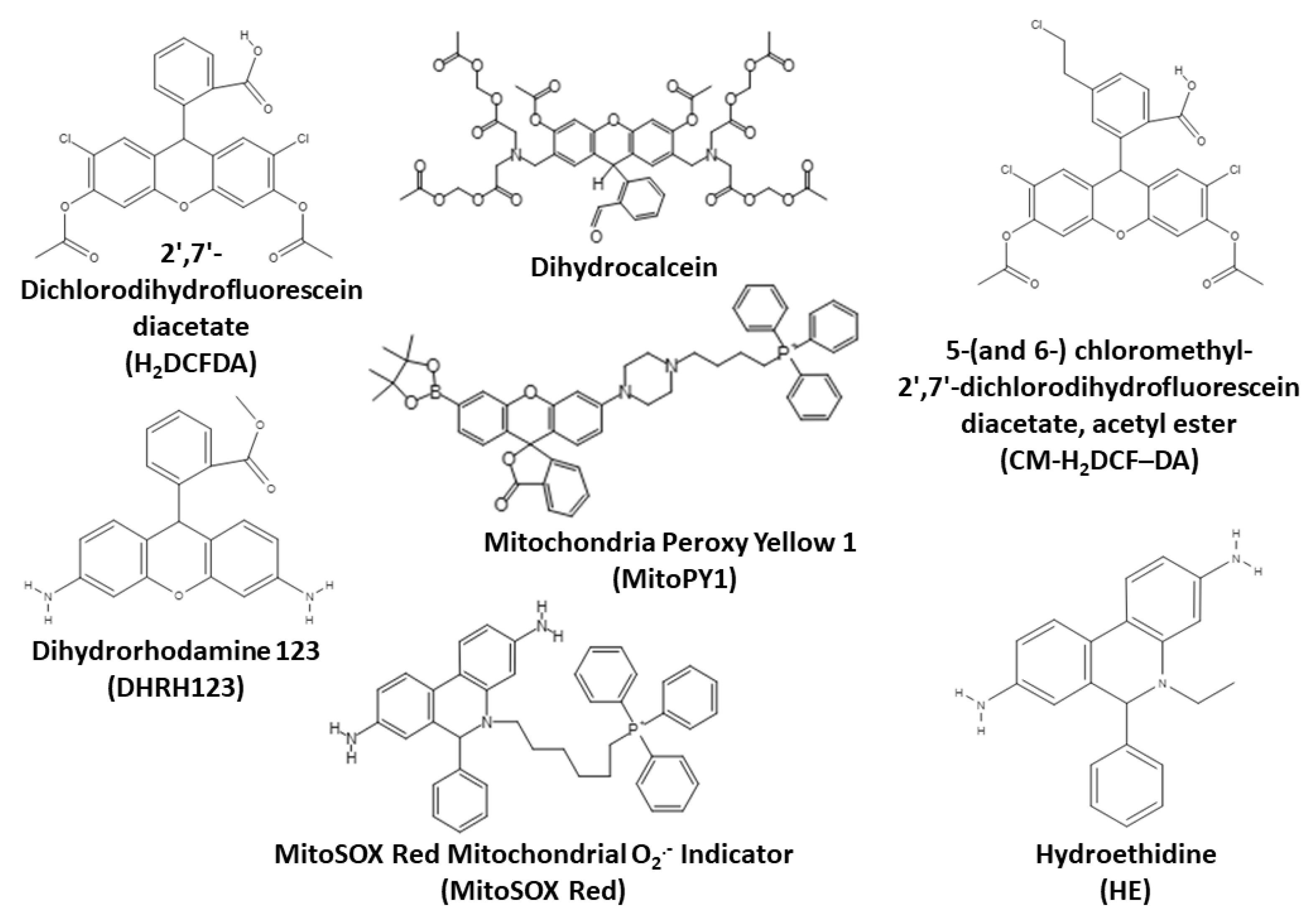
3.2.2. 2′,7′-Dichlorodihydrofluorescein Diacetate and Related Probes
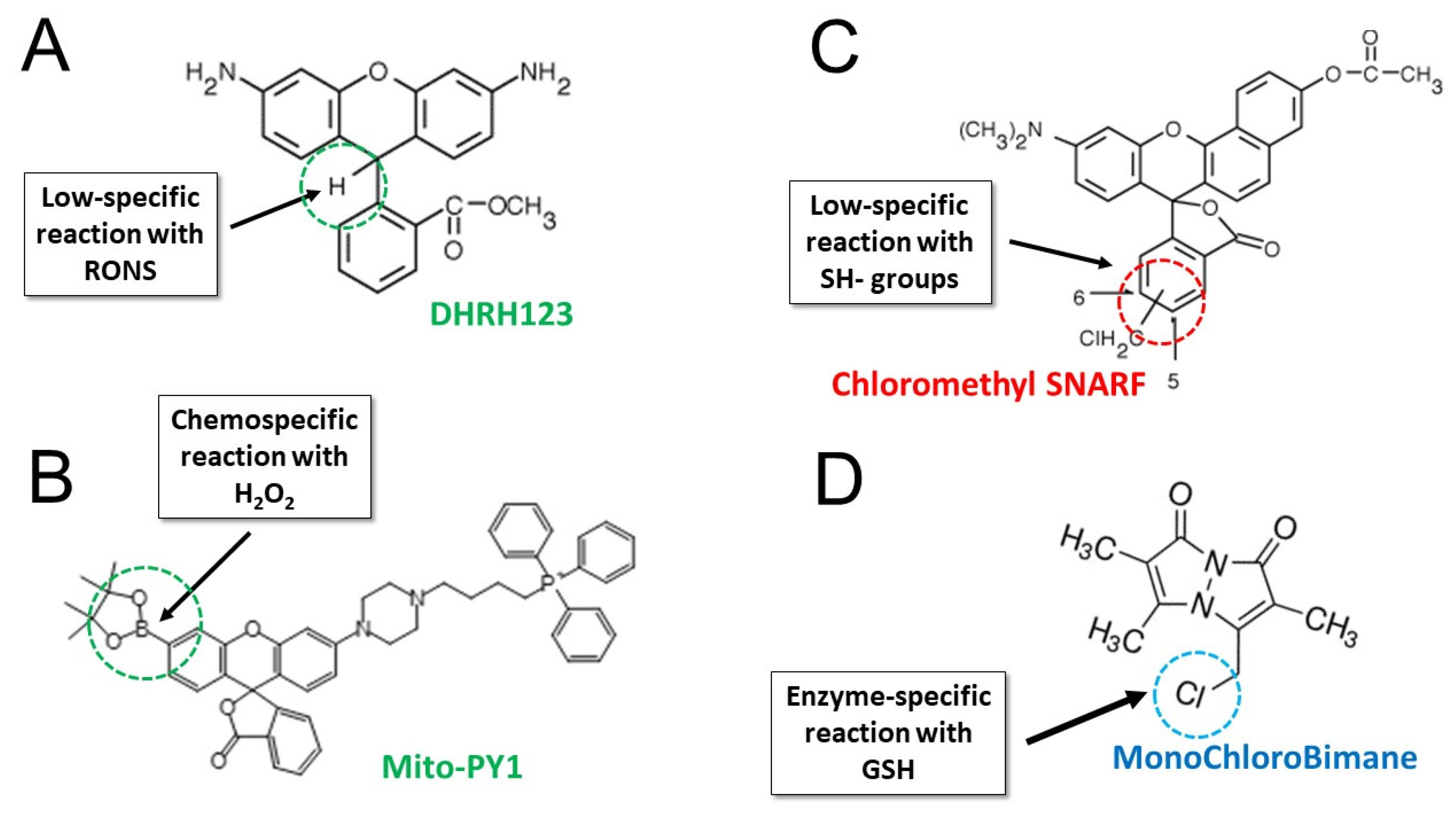
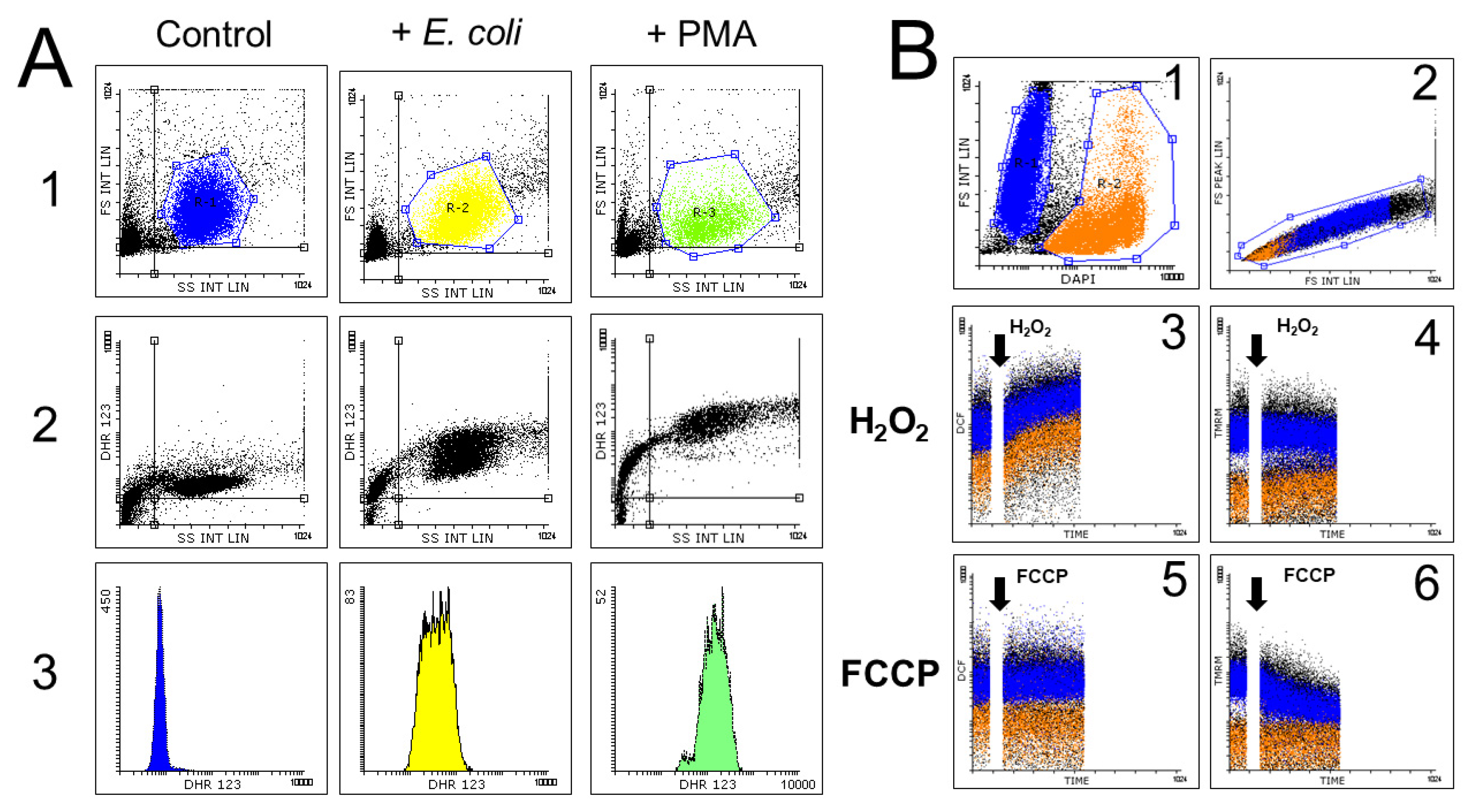
3.2.3. Dihydrorhodamine 123
3.2.4. Mitochondria Peroxy Yellow 1 and Related Arylboronate Fluorescent Probes
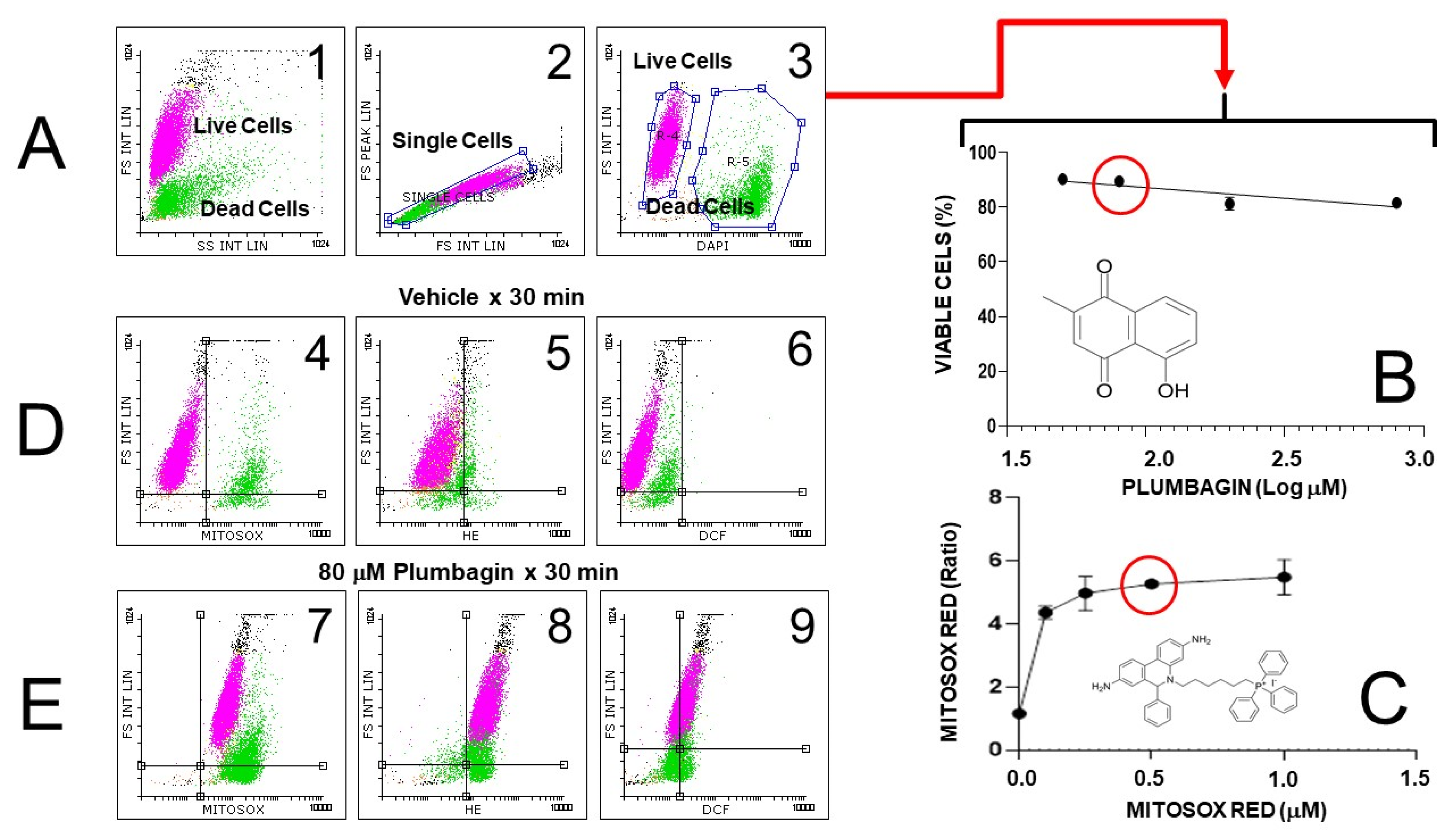
3.2.5. Hydroethidine and MitoSOX Mitochondrial O2•− Indicators
3.2.6. CellROXTM Reagents
3.2.7. ROS-IDTM Reagents
3.3. Detection of More Stable Products of ROS Reaction
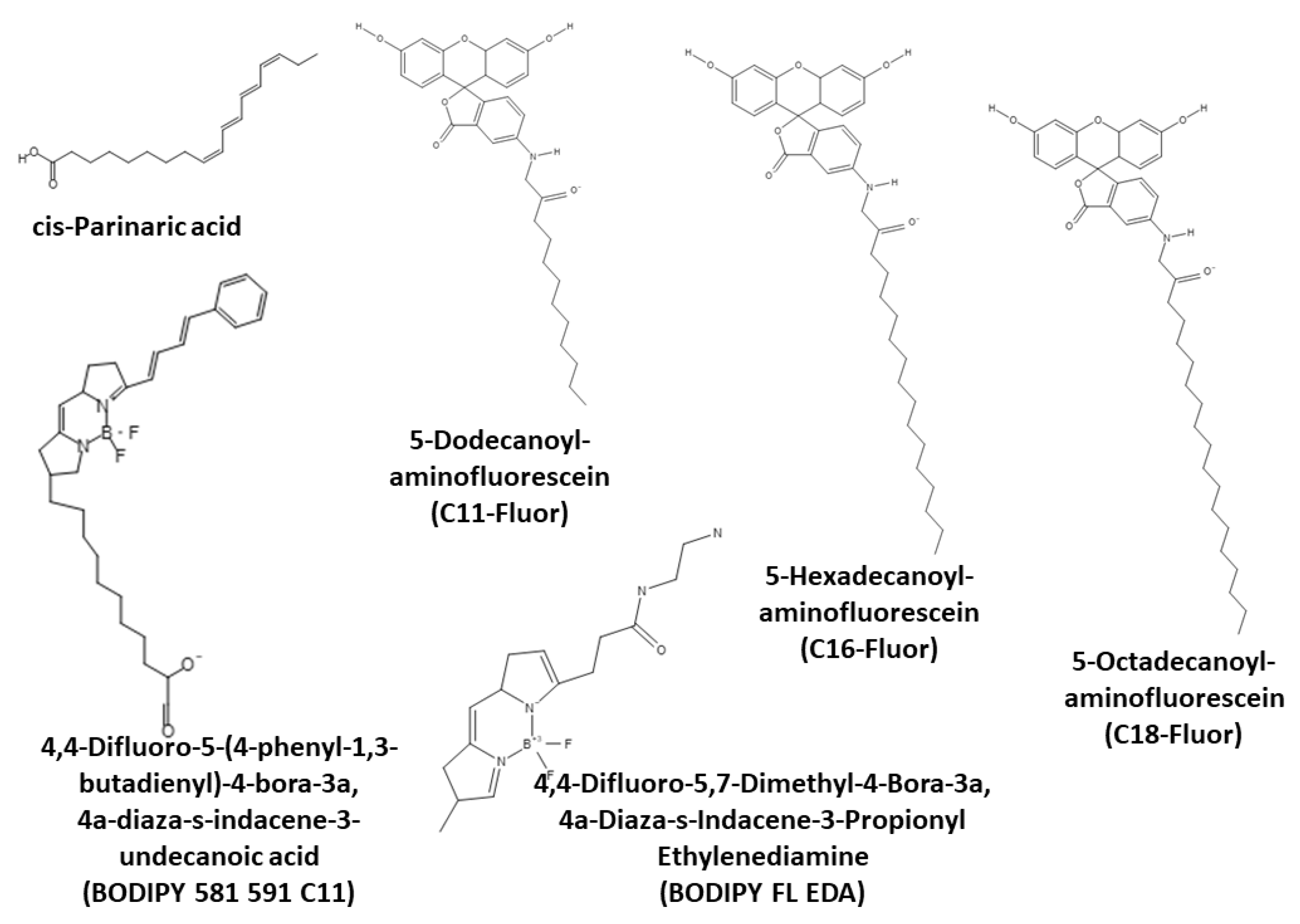
3.3.1. Detection of Lipid Peroxidation
cis-Parinaric Acid
BODIPY 581/591 C11 and Related Probes
Lipophilic Fluorescein Derivatives
3.3.2. Detection of Metabolic Derivatives of Peroxidized Lipids
Detection of 4-Hydroxy-2-nonenal by Immunofluorescence
Immunofluorescent Detection of Oxidized Bases in DNA
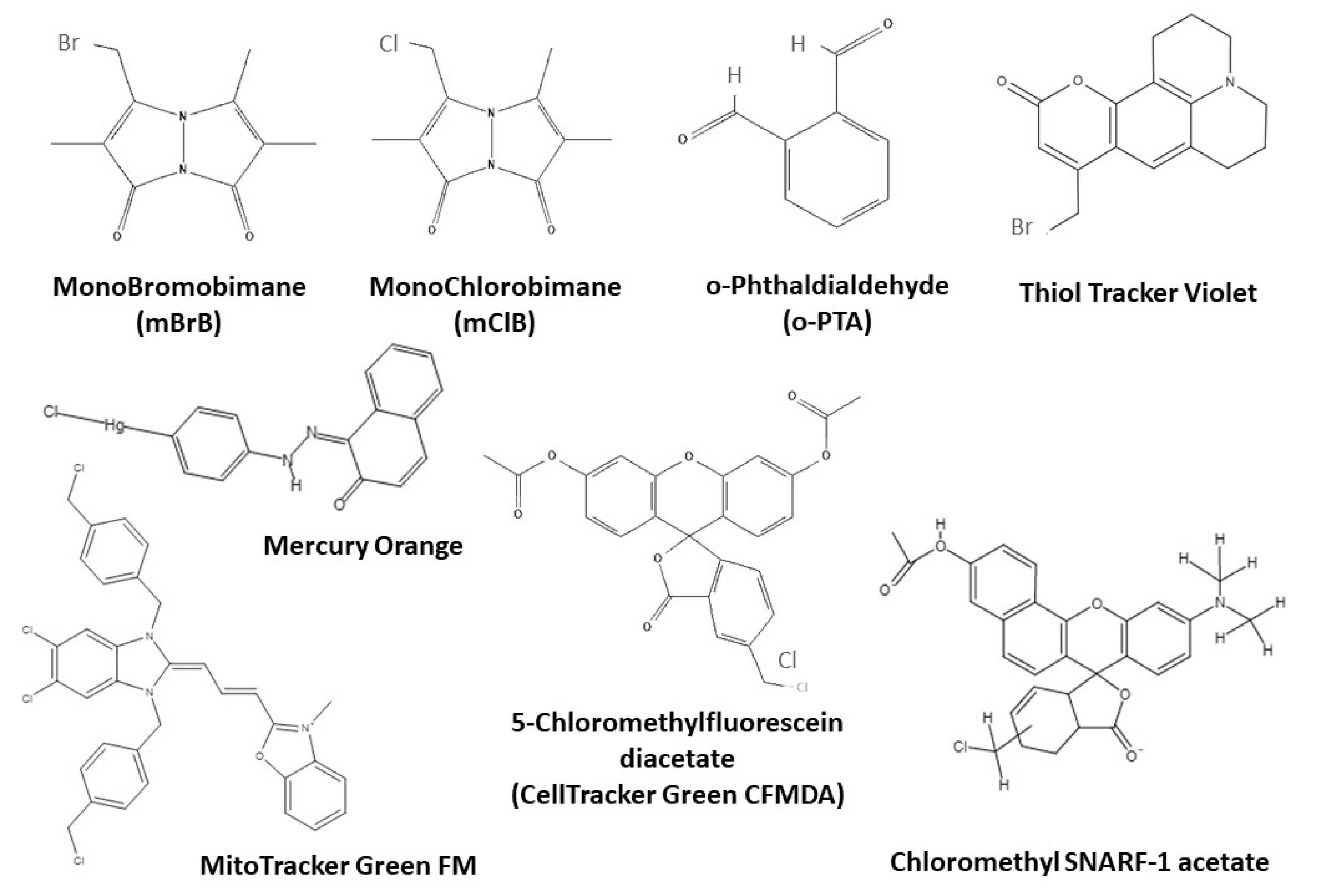
3.4. Assessment of Antioxidant Defenses: Glutathione and Thiols
4. FCM in Oxidative Stress Research
4.1. Variability in Half-Life and Intracellular Sources of ROS
4.2. Interactions among and between ROS and RNS
4.3. Influence of the Probes on the Experimental System
4.4. Cell Integrity and Intracellular Retention of Probes
4.5. Experimental Artifacts
4.6. Intrinsic Limitations of Fluorogenic Substrates and Probes
4.6.1. Probes Used for Detection of H2O2 and Organic Peroxides
4.6.2. Probes Used for Detection of O2•−
4.6.3. Probes Used for Detection of Lipid Peroxides
4.6.4. Probes Used for the Determination of GSH
5. Recommendations for Performing FCM Analysis of ROS, RNS and Oxidative Stress
5.1. Inclusion of Experimental Controls
5.1.1. Positive Controls
5.1.2. Negative Controls
5.1.3. Genetically-Modified Organisms as Controls
5.2. Choice of Fluorescent Probes
5.3. Titration of Fluorescent Probes
5.4. Range-Finding Experiments and Exclusion of Dead/Injured Cells
5.5. Data Generation, Presentation, Data Analysis and Publication
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Taverne, Y.J.; Merkus, D.; Bogers, A.J.; Halliwell, B.; Duncker, D.J.; Lyons, T.W. Reactive Oxygen Species: Radical Factors in the Evolution of Animal Life. Bioessays 2018, 40, 1700158. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, G.; Fasciolo, G.; Venditti, P. The Ambiguous Aspects of Oxygen. Oxygen 2022, 2, 382–409. [Google Scholar] [CrossRef]
- Forman, H.J.; Augusto, O.; Brigelius-Flohe, R.; Dennery, P.A.; Kalyanaraman, B.; Ischiropoulos, H.; Mann, G.E.; Radi, R.; Roberts, L.J.; Vina, J.; et al. Even free radicals should follow some rules: A Guide to free radical research terminology and methodology. Free Radic. Biol. Med. 2015, 78, 233–235. [Google Scholar] [CrossRef]
- Prasad, A.; Sedlářová, M.; Pospíšil, P. Singlet oxygen imaging using fluorescent probe Singlet Oxygen Sensor Green in photosynthetic organisms. Sci. Rep. 2018, 8, 13685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxid. Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef] [PubMed]
- Speckmann, B.; Steinbrenner, H.; Grune, T.; Klotz, L.-O. Peroxynitrite: From interception to signaling. Arch. Biochem. Biophys. 2016, 595, 153–160. [Google Scholar] [CrossRef]
- Aranda-Rivera, A.K.; Cruz-Gregorio, A.; Arancibia-Hernández, Y.L.; Hernández-Cruz, E.Y.; Pedraza-Chaverri, J. RONS and Oxidative Stress: An Overview of Basic Concepts. Oxygen 2022, 2, 437–478. [Google Scholar] [CrossRef]
- Clancy, D.; Birdsall, J. Flies, worms and the Free Radical Theory of ageing. Ageing Res. Rev. 2013, 12, 404–412. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, G.; Cortopassi, G. Oxidative stress in inherited mitochondrial diseases. Free Radic. Biol. Med. 2015, 88, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Moulin, M.; Ferreiro, A. Muscle redox disturbances and oxidative stress as pathomechanisms and therapeutic targets in early-onset myopathies. Semin. Cell Dev. Biol. 2017, 64, 213–223. [Google Scholar] [CrossRef]
- Reula, A.; Pellicer, D.; Castillo, S.; Magallón, M.; Armengot, M.; Herrera, G.; O’connor, J.-E.; Bañuls, L.; Navarro-García, M.M.; Escribano, A.; et al. New Laboratory Protocol to Determine the Oxidative Stress Profile of Human Nasal Epithelial Cells Using Flow Cytometry. J. Clin. Med. 2021, 10, 1172. [Google Scholar] [CrossRef]
- Beltrán, B.; Nos, P.; Dasí, F.; Iborra, M.; Bastida, G.; Martínez, M.; O’connor, J.-E.; Sáez, G.; Moret, I.; Ponce, J. Mitochondrial dysfunction, persistent oxidative damage, and catalase inhibition in immune cells of naïve and treated Crohn’s disease. Inflamm. Bowel Dis. 2010, 16, 76–86. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative Stress: An Essential Factor in the Pathogenesis of Gastrointestinal Mucosal Diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Tan, H.-Y.; Wang, N.; Zhang, Z.-J.; Lao, L.; Wong, C.-W.; Feng, Y. The Role of Oxidative Stress and Antioxidants in Liver Diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef] [Green Version]
- Siti, H.N.; Kamisah, Y.; Kamsiah, J. The role of oxidative stress, antioxidants and vascular inflammation in cardiovascular disease (a review). Vasc. Pharmacol. 2015, 71, 40–56. [Google Scholar] [CrossRef]
- Santilli, F.; D’Ardes, D.; Davì, G. Oxidative stress in chronic vascular disease: From prediction to prevention. Vasc. Pharmacol. 2015, 74, 23–37. [Google Scholar] [CrossRef]
- Li, H.; Horke, S.; Förstermann, U. Vascular oxidative stress, nitric oxide and atherosclerosis. Atherosclerosis 2014, 237, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Montezano, A.C.; Dulak-Lis, M.; Tsiropoulou, S.; Harvey, A.; Briones, A.M.; Touyz, R.M. Oxidative Stress and Human Hypertension: Vascular Mechanisms, Biomarkers, and Novel Therapies. Can. J. Cardiol. 2015, 31, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, E.; Palomo, I. Role of oxidative stress on platelet hyperreactivity during aging. Life Sci. 2016, 148, 17–23. [Google Scholar] [CrossRef]
- Collado, R.; Ivars, D.; Oliver, I.; Tormos, C.; Egea, M.; Miguel, A.; Sáez, G.T.; Carbonell, F. Increased Oxidative Damage Associated with Unfavorable Cytogenetic Subgroups in Chronic Lymphocytic Leukemia. BioMed Res. Int. 2014, 2014, 686392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, B.; Figtree, G.; Costa, D.; Eade, T.; Hruby, G.; Lim, S.; Elfiky, A.; Martine, N.; Rosenthal, D.; Clarke, S.; et al. Oxidative stress in prostate cancer patients: A systematic review of case control studies. Prostate Int. 2016, 4, 71–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Wen, J.; Huang, Z.; Nice, E.C.; Huang, C.; Zhang, H.; Li, Q. Redox proteomics screening cellular factors associated with oxidative stress in hepatocarcinogenesis. Proteom. Clin. Appl. 2017, 11, 1600089. [Google Scholar] [CrossRef] [PubMed]
- Marengo, B.; Nitti, M.; Furfaro, A.L.; Colla, R.; De Ciucis, C.; Marinari, U.M.; Pronzato, M.A.; Traverso, N.; Domenicotti, C. Redox Homeostasis and Cellular Antioxidant Systems: Crucial Players in Cancer Growth and Therapy. Oxid. Med. Cell. Longev. 2016, 2016, 6235641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, D.; Zhelev, Z.; Aoki, I.; Bakalova, R.; Higashi, T. Overproduction of reactive oxygen species—obligatory or not for induction of apoptosis by anticancer drugs. Chin. J. Cancer Res. 2016, 28, 383–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, A.V.; Valuev-Elliston, V.T.; Ivanova, O.N.; Kochetkov, S.N.; Starodubova, E.S.; Bartosch, B.; Isaguliants, M.G. Oxidative Stress during HIV Infection: Mechanisms and Consequences. Oxid. Med. Cell Longev. 2016, 2016, 8910396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santus, P.; Danzo, F.; Zuffi, A.; Pini, S.; Saad, M.; Visconti, A.; Radovanovic, D. Oxidative stress and viral Infections: Rationale, experiences, and perspectives on N-acetylcysteine. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 8582–8590. [Google Scholar] [CrossRef] [PubMed]
- Theofanous, T.; Kourti, M. Abrogating Oxidative Stress as a Therapeutic Strategy against Parkinson’s Disease: A Mini Review of the Recent Advances on Natural Therapeutic Antioxidant and Neuroprotective Agents. Med. Chem. 2022, 18, 772–783. [Google Scholar] [CrossRef]
- Carvalho, A.N.; Branco, V.; van Horssen, J.; Saso, L. Editorial: Redox-signaling in neurodegenerative diseases: Biomarkers, targets, and therapies. Front. Cell. Neurosci. 2023, 17, 1198669. [Google Scholar] [CrossRef]
- Kamat, P.K.; Kalani, A.; Rai, S.; Swarnkar, S.; Tota, S.; Nath, C.; Tyagi, N. Mechanism of Oxidative Stress and Synapse Dysfunction in the Pathogenesis of Alzheimer’s Disease: Understanding the Therapeutics Strategies. Mol. Neurobiol. 2016, 53, 648–661. [Google Scholar] [CrossRef] [Green Version]
- Rani, V.; Deep, G.; Singh, R.K.; Palle, K.; Yadav, U.C.S. Oxidative stress and metabolic disorders: Pathogenesis and therapeutic strategies. Life Sci. 2016, 148, 183–193. [Google Scholar] [CrossRef]
- Forman, H.J. Redox signaling: An evolution from free radicals to aging. Free Radic. Biol. Med. 2016, 97, 398–407. [Google Scholar] [CrossRef] [Green Version]
- Huwiler, A.; Beck, K.-F.; Pfeilschifter, J. Cross-Regulation of the Cellular Redox System, Oxygen, and Sphingolipid Signalling. Metabolites 2023, 13, 426. [Google Scholar] [CrossRef]
- Münzel, T.; Daiber, A. Vascular Redox Signaling, Endothelial Nitric Oxide Synthase Uncoupling, and Endothelial Dysfunction in the Setting of Transportation Noise Exposure or Chronic Treatment with Organic Nitrates. Antioxid. Redox Signal. 2023, 38, 1001–1021. [Google Scholar] [CrossRef]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef] [PubMed]
- El-Benna, J.; Hurtado-Nedelec, M.; Marzaioli, V.; Marie, J.-C.; Gougerot-Pocidalo, M.-A.; Dang, P.M.-C. Priming of the neutrophil respiratory burst: Role in host defense and inflammation. Immunol. Rev. 2016, 273, 180–193. [Google Scholar] [CrossRef]
- Qin, S.; Li, B.; Ming, H.; Nice, E.C.; Zou, B.; Huang, C. Harnessing redox signaling to overcome therapeutic-resistant cancer dormancy. Biochim. Biophys. Acta (BBA) Rev. Cancer 2022, 1877, 188749. [Google Scholar] [CrossRef] [PubMed]
- Lionaki, E.; Markaki, M.; Tavernarakis, N. Autophagy and ageing: Insights from invertebrate model organisms. Ageing Res. Rev. 2013, 12, 413–428. [Google Scholar] [CrossRef] [Green Version]
- Gibellini, L.; De Biasi, S.; Pinti, M.; Nasi, M.; Riccio, M.; Carnevale, G.; Cavallini, G.M.; de Oyanguren, F.J.S.; O’connor, J.E.; Mussini, C.; et al. The protease inhibitor atazanavir triggers autophagy and mitophagy in human preadipocytes. Aids 2012, 26, 2017–2026. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, B.C.; Chang, C.J. Chemistry and biology of reactive oxygen species in signaling or stress responses. Nat. Chem. Biol. 2011, 7, 504–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative Stress, Prooxidants, and Antioxidants: The Interplay. BioMed Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frijhoff, J.; Winyard, P.G.; Zarkovic, N.; Davies, S.S.; Stocker, R.; Cheng, D.; Knight, A.R.; Taylor, E.L.; Oettrich, J.; Ruskovska, T.; et al. Clinical Relevance of Biomarkers of Oxidative Stress. Antioxid. Redox Signal. 2015, 23, 1144–1170. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Ren, X.; Zhang, L.; Liu, Z.; Hai, Z. Mitochondria-Targeted Fluorescent and Photoacoustic Imaging of Hydrogen Peroxide in Inflammation. Anal. Chem. 2020, 92, 14244–14250. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Ouyang, W.; Qiu, H.; Zhang, Z.; Wang, Z.; Xing, B. Detection of Reactive Oxygen and Nitrogen Species by Upconversion Nanoparticle-Based Near-Infrared Nanoprobes: Recent Progress and Perspectives. Chem. A Eur. J. 2022, 28, e202201966. [Google Scholar] [CrossRef]
- Geng, Y.; Wang, Z.; Zhou, J.; Zhu, M.; Liu, J.; James, T.-D. Recent progress in the development of fluorescent probes for imaging pathological oxidative stress. Chem. Soc. Rev. 2023. ahead of print. [Google Scholar] [CrossRef]
- Hong, S.; Pawel, G.T.; Pei, R.; Lu, Y. Recent progress in developing fluorescent probes for imaging cell metabolites. Biomed. Mater. 2021, 16, 044108. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Zhu, T.; Xu, J.; Zhao, Y.; Kuang, Y.; Sun, M.; Chen, Y.; He, W.; Wang, Z.; Jiang, T.; et al. Organic Fluorescent Probes for Monitoring Micro-Environments in Living Cells and Tissues. Molecules 2023, 28, 3455. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Haghighatbin, M.A.; Hogan, C.F.; New, E.J. A FRET-based ratiometric redox probe for detecting oxidative stress by confocal microscopy, FLIM and flow cytometry. Chem. Commun. 2015, 51, 10510–10513. [Google Scholar] [CrossRef]
- Fallico, V.; Rea, M.; Stanton, C.; Ilestam, N.; McKinney, J. Next-generation multiparameter flow cytometry assay improves the assessment of oxidative stress in probiotics. Food Microbiol. 2020, 91, 103501. [Google Scholar] [CrossRef]
- Kraaij, T.; Tengström, F.C.; Kamerling, S.W.; Pusey, C.D.; Scherer, H.U.; Toes, R.E.; Rabelink, T.J.; van Kooten, C.; Teng, Y.O. A novel method for high-throughput detection and quantification of neutrophil extracellular traps reveals ROS-independent NET release with immune complexes. Autoimmun. Rev. 2016, 15, 577–584. [Google Scholar] [CrossRef] [Green Version]
- Bulkeley, E.; Santistevan, A.C.; Varner, D.; Meyers, S. Imaging flow cytometry to characterize the relationship between abnormal sperm morphologies and reactive oxygen species in stallion sperm. Reprod. Domest. Anim. 2023, 58, 10–19. [Google Scholar] [CrossRef]
- Kalyanaraman, B.; Darley-Usmar, V.; Davies, K.J.A.; Dennery, P.A.; Forman, H.J.; Grisham, M.B.; Mann, G.E.; Moore, K.; Roberts, L.J., II; Ischiropoulos, H. Measuring reactive oxygen and nitrogen species with fluorescent probes: Challenges and limitations. Free Radic. Biol. Med. 2012, 52, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Debowska, K.; Debski, D.; Hardy, M.; Jakubowska, M.; Kalyanaraman, B.; Marcinek, A.; Michalski, R.; Michalowski, B.; Ouari, O.; Sikora, A.; et al. Toward selective detection of reactive oxygen and nitrogen species with the use of fluorogenic probes—Limitations, progress, and perspectives. Pharmacol. Rep. 2015, 67, 756–764. [Google Scholar] [CrossRef] [Green Version]
- O’connor, J.-E.; Herrera, G.; Sala-De-Oyanguren, F.; Jávega, B.; Martínez-Romero, A. Cytomics of Oxidative Stress: Probes and Problems. In Single Cell Analysis: Contemporary Research and Clinical Applications; Robinson, J., Cossarizza, A., Eds.; Springer: Singapore, 2017; pp. 83–118. [Google Scholar] [CrossRef]
- Goetz, C.; Hammerbeck, C.; Bonnevier, J. Flow Cytometry Basics for the Non-Expert; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Ortolani, C. Flow Cytometry Today: Everything You Need to Know about Flow Cytometry; Springer: Cham, Switzerland, 2022. [Google Scholar]
- Robinson, J.P. Spectral flow cytometry—Quo vadimus? Cytom. Part A 2019, 95, 823–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, I. Flow Cytometry: Recent Perspectives; Intechopen Limited: London, UK, 2012. [Google Scholar] [CrossRef]
- Cossarizza, A.; Chang, H.-D.; Radbruch, A.; Acs, A.; Adam, D.; Adam-Klages, S.; Agace, W.W.; Aghaeepour, N.; Akdis, M.; Allez, M.; et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies (second edition). Eur. J. Immunol. 2019, 49, 1457–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortolani, C. Flow Cytometry of Hematological Malignancies, 2nd ed.; Wiley Blackwell: Hoboken, NJ, USA, 2021. [Google Scholar]
- Robinson, J.P. Flow cytometry: Past and future. Biotechniques 2022, 72, 159–169. [Google Scholar] [CrossRef]
- Spidlen, J.; Moore, W.E.; Parks, D.R.; Goldberg, M.L.; Bray, C.L.; Bierre, P.; Gorombey, P.; Hyun, B.; Hubbard, M.; Lange, S.; et al. Data File Standard for Flow Cytometry, version FCS 3.1. Cytom. Part A 2010, 77, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Felipo-Benavent, M.; Martínez-Romero, A.; Rubio-Guerri, C.; Álvaro-Álvarez, T.; Gil, D.; García-Párraga, D.; O’Connor, J. Flow cytometric kinetic assay of calcium mobilization in whole blood platelets of bottlenose dolphins (Tursiops truncatus). Cytom. Part A 2023, 103, 347–352. [Google Scholar] [CrossRef]
- Sun, G.; Qu, L.; Azi, F.; Liu, Y.; Li, J.; Lv, X.; Du, G.; Chen, J.; Chen, C.-H.; Liu, L. Recent progress in high-throughput droplet screening and sorting for bioanalysis. Biosens. Bioelectron. 2023, 225, 115107. [Google Scholar] [CrossRef]
- Sikora, J.P.; Sobczak, J.; Zawadzki, D.; Przewratil, P.; Wysocka, A.; Burzyńska, M. Respiratory Burst and TNF-α Receptor Expression of Neutrophils after Sepsis and Severe Injury-Induced Inflammation in Children. Int. J. Environ. Res. Public Health 2021, 18, 2187. [Google Scholar] [CrossRef]
- De Gaetano, A.; Gibellini, L.; Zanini, G.; Nasi, M.; Cossarizza, A.; Pinti, M. Mitophagy and Oxidative Stress: The Role of Aging. Antioxidants 2021, 10, 794. [Google Scholar] [CrossRef]
- Gibellini, L.; Pinti, M.; Nasi, M.; De Biasi, S.; Roat, E.; Bertoncelli, L.; Cossarizza, A. Interfering with ROS Metabolism in Cancer Cells: The Potential Role of Quercetin. Cancers 2010, 2, 1288–1311. [Google Scholar] [CrossRef] [Green Version]
- Petit, P.X.; Ardilla-Osorio, H.; Penalvia, L.; Rainey, N.E. Tafazzin Mutation Affecting Cardiolipin Leads to Increased Mitochondrial Superoxide Anions and Mitophagy Inhibition in Barth Syndrome. Cells 2020, 9, 2333. [Google Scholar] [CrossRef] [PubMed]
- Ivars, D.; Orero, M.T.; Javier, K.; Díaz-Vico, L.; García-Giménez, J.L.; Mena, S.; Tormos, C.; Egea, M.; Pérez, P.L.; Arrizabalaga, B.; et al. Oxidative imbalance in low/intermediate-1-risk myelodysplastic syndrome patients: The influence of iron overload. Clin. Biochem. 2017, 50, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Cocchi, V.; Jávega, B.; Gasperini, S.; O’connor, J.-E.; Lenzi, M.; Hrelia, P. 6-(Methylsulfonyl) Hexyl Isothiocyanate: A Chemopreventive Agent Inducing Autophagy in Leukemia Cell Lines. Biomolecules 2022, 12, 1485. [Google Scholar] [CrossRef] [PubMed]
- Urios, A.; López-Gresa, M.; González, M.; Primo, J.; Martínez, A.; Herrera, G.; Escudero, J.C.; O’Connor, J.-E.; Blanco, M. Nitric oxide promotes strong cytotoxicity of phenolic compounds against Escherichia coli: The influence of antioxidant defenses. Free Radic. Biol. Med. 2003, 35, 1373–1381. [Google Scholar] [CrossRef]
- Hirota, K. Basic Biology of Hypoxic Responses Mediated by the Transcription Factor HIFs and Its Implication for Medicine. Biomedicines 2020, 8, 32. [Google Scholar] [CrossRef] [Green Version]
- Godet, I.; Doctorman, S.; Wu, F.; Gilkes, D.M. Detection of Hypoxia in Cancer Models: Significance, Challenges, and Advances. Cells 2022, 11, 686. [Google Scholar] [CrossRef]
- Dey, D.; Shrivastava, V.; Joshi, D.; Singal, C.M.S.; Tyagi, S.; Bhat, M.A.; Jaiswal, P.; Sharma, J.B.; Palanichamy, J.K.; Sinha, S.; et al. Hypoxia Induces Early Neurogenesis in Human Fetal Neural Stem Cells by Activating the WNT Pathway. Mol. Neurobiol. 2023, 60, 2910–2921. [Google Scholar] [CrossRef]
- Pachón-Peña, G.; Serena, C.; Ejarque, M.; Petriz, J.; Duran, X.; Oliva-Olivera, W.; Simó, R.; Tinahones, F.J.; Fernández-Veledo, S.; Vendrell, J. Obesity Determines the Immunophenotypic Profile and Functional Characteristics of Human Mesenchymal Stem Cells From Adipose Tissue. Stem Cells Transl. Med. 2016, 5, 464–475. [Google Scholar] [CrossRef]
- Nunez-Espinosa, C.; Ferreira, I.; Rios-Kristjansson, J.; Rizo-Roca, D.; Godoy, M.G.; Rico, L.; Rubi-Sans, G.; Torrella, J.; Pages, T.; Petriz, J.; et al. Effects of Intermittent Hypoxia and Light Aerobic Exercise on Circulating Stem Cells and Side Population, after Strenuous Eccentric Exercise in Trained Rats. Curr. Stem Cell Res. Ther. 2015, 10, 132–139. [Google Scholar] [CrossRef]
- Synowiec, A.; Brodaczewska, K.; Wcisło, G.; Majewska, A.; Borkowska, A.; Filipiak-Duliban, A.; Gawrylak, A.; Wilkus, K.; Piwocka, K.; Kominek, A.; et al. Hypoxia, but Not Normoxia, Reduces Effects of Resveratrol on Cisplatin Treatment in A2780 Ovarian Cancer Cells: A Challenge for Resveratrol Use in Anticancer Adjuvant Cisplatin Therapy. Int. J. Mol. Sci. 2023, 24, 5715. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wang, J.; Jin, Y.; Lin, J.; Gong, L.; Xu, Y. Hypoxia upregulates the expression of lncRNA H19 in non-small cell lung cancer cells and induces drug resistance. Transl. Cancer Res. 2022, 11, 2876–2886. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gao, J.; Liu, C.; Bu, N.; Zhan, S.; Wu, H.; Zhang, R.; Sun, H.; Fan, H. USP22 knockdown protects against cerebral ische-mia/reperfusion injury via destabilizing PTEN protein and activating the mTOR/TFEB pathway. Naunyn Schmiedebergs Arch. Pharmacol. 2023; ahead of print. [Google Scholar] [CrossRef]
- Chen, G.; Yang, Y.; Xu, C.; Gao, S. A Flow Cytometry-based Assay for Measuring Mitochondrial Membrane Potential in Cardiac Myocytes After Hypoxia/Reoxygenation. J. Vis. Exp. 2018, 137, e57725. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Liu, Y.; Hao, Y.; Hou, D.; Yang, R. Lycium barbarum polysaccharide protects cardiomyocytes from hypoxia/reoxygenation injury via activation of SIRT3/CypD signaling. Ann. Transl. Med. 2023, 11, 72. [Google Scholar] [CrossRef]
- Feng, J.; He, W.; Xia, J.; Huang, Q.; Yang, J.; Gu, W.; Zhang, N.; Liu, Y. Human umbilical cord mesenchymal stem cells-derived exosomal circDLGAP4 promotes angiogenesis after cerebral ischemia–reperfusion injury by regulating miR-320/KLF5 axis. FASEB J. 2023, 37, e22733. [Google Scholar] [CrossRef]
- Qiao, J.; Wang, M.; Cui, M.; Fang, Y.; Li, H.; Zheng, C.; Li, Z.; Xu, Y.; Hua, H.; Li, D. Small-molecule probes for fluorescent detection of cellular hypoxia-related nitroreductase. J. Pharm. Biomed. Anal. 2021, 203, 114199. [Google Scholar] [CrossRef] [PubMed]
- Wallabregue, A.L.D.; Bolland, H.; Faulkner, S.; Hammond, E.M.; Conway, S.J. Two Color Imaging of Different Hypoxia Levels in Cancer Cells. J. Am. Chem. Soc. 2023, 145, 2572–2583. [Google Scholar] [CrossRef]
- Labiano, S.; Serrano-Mendioroz, I.; Rodriguez-Ruiz, M.E. Assessment of hypoxia by pimonidazole staining following radiotherapy. Methods Cell Biol. 2022, 172, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Foehrenbacher, A.; Su, J.; Patel, R.; Hay, M.P.; Hicks, K.O.; Wilson, W.R. The 2-Nitroimidazole EF5 Is a Biomarker for Oxidoreductases That Activate the Bioreductive Prodrug CEN-209 under Hypoxia. Clin. Cancer Res. 2012, 18, 1684–1695. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, K.; Homma, M.; Suzuki, M.; Yokouchi, Y.; Matsuda, T.; Takakura, H.; Hirata, K.; Kuge, Y.; Ogawa, M. Reduction of tumor hypoxia by anti-PD-1 therapy assessed using pimonidazole and [18F]FMISO. Nucl. Med. Biol. 2022, 108, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Gravelle, P.; Jean, C.; Familiades, J.; Decaup, E.; Blanc, A.; Bezombes-Cagnac, C.; Laurent, C.; Savina, A.; Fournié, J.-J.; Laurent, G. Cell Growth in Aggregates Determines Gene Expression, Proliferation, Survival, Chemoresistance, and Sensitivity to Immune Effectors in Follicular Lymphoma. Am. J. Pathol. 2014, 184, 282–295. [Google Scholar] [CrossRef] [PubMed]
- Chang, Q.; Ornatsky, O.I.; Koch, C.J.; Chaudary, N.; Marie-Egyptienne, D.T.; Hill, R.P.; Tanner, S.D.; Hedley, D.W. Single-cell measurement of the uptake, intratumoral distribution and cell cycle effects of cisplatin using mass cytometry. Int. J. Cancer 2015, 136, 1202–1209. [Google Scholar] [CrossRef]
- Document Connect (thermofisher.com). Available online: https://www.thermofisher.com/document-connect/document-connect.html?url=https://assets.thermofisher.com/TFS-Assets%2FLSG%2Fmanuals%2FMAN0017632_Hypoxia_Green_for_Flow_Cytometry_UG.pdf (accessed on 27 April 2023).
- BioTracker 520 Green Hypoxia Dye Live Cell Imaging Millipore (sigmaaldrich.com). Available online: https://www.sigmaaldrich.com/ES/es/product/mm/sct033 (accessed on 27 April 2023).
- Document Connect (thermofisher.com). Available online: https://www.thermofisher.com/document-connect/document-connect.html?url=https://assets.thermofisher.com/TFS-Assets%2FLSG%2Fmanuals%2FMAN0013497_Image_iT_Hypoxia_Reagents_UG.pdf (accessed on 27 April 2023).
- Zhang, S.; Hosaka, M.; Yoshihara, T.; Negishi, K.; Iida, Y.; Tobita, S.; Takeuchi, T. Phosphorescent Light–Emitting Iridium Complexes Serve as a Hypoxia-Sensing Probe for Tumor Imaging in Living Animals. Cancer Res. 2010, 70, 4490–4498. [Google Scholar] [CrossRef] [Green Version]
- Bogdanov, A.; Mishin, A.; Yampolsky, I.; Belousov, V.V.; Chudakov, D.; Subach, F.V.; Verkhusha, V.; Lukyanov, S.; Lukyanov, K. Green fluorescent proteins are light-induced electron donors. Nat. Chem. Biol. 2009, 5, 459–461. [Google Scholar] [CrossRef]
- Lin, X.; Gately, D.P.; Hom, D.; Mishima, M.; Los, G.; Howell, S.B. Quantification of tumor cell injury in vitro and in vivo using expression of green fluorescent protein under the control of the GADD153 promoter. Int. J. Cancer 2001, 91, 555–562. [Google Scholar] [CrossRef]
- Chia, H.E.; Koebke, K.J.; Rangarajan, A.A.; Koropatkin, N.M.; Marsh, E.N.G.; Biteen, J.S. New Orange Ligand-Dependent Fluorescent Reporter for Anaerobic Imaging. ACS Chem. Biol. 2021, 16, 2109–2115. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Wang, A.; Huang, L.; Zou, Y.; Gu, Y.; Chen, X.; Zhao, Y.; Yang, Y. Monitoring cellular redox state under hypoxia using a fluorescent sensor based on eel fluorescent protein. Free Radic. Biol. Med. 2018, 120, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Cho, S.; Han, Y.; You, Y.; Nam, W. Ratiometric Fluorescent Probes for Detection of Intracellular Singlet Oxygen. Org. Lett. 2013, 15, 3582–3585. [Google Scholar] [CrossRef]
- Ragàs, X.; Jiménez-Banzo, A.; Sánchez-García, D.; Batllori, X.; Nonell, S. Singlet oxygen photosensitisation by the fluorescent probe Singlet Oxygen Sensor Green®. Chem. Commun. 2009, 20, 2920–2922. [Google Scholar] [CrossRef] [PubMed]
- Murotomi, K.; Umeno, A.; Sugino, S.; Yoshida, Y. Quantitative kinetics of intracellular singlet oxygen generation using a fluorescence probe. Sci. Rep. 2020, 10, 10616. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, N.; Tanaka, K.; Urano, Y.; Kikuchi, K.; Higuchi, T.; Nagano, T. Novel Fluorescent Probes for Singlet Oxygen. Angew. Chem. Int. Ed. Engl. 1999, 38, 2899–2901. [Google Scholar] [CrossRef]
- Hideg, É.; Kálai, T.; Kós, P.B.; Asada, K.; Hideg, K. Singlet Oxygen in Plants—Its Significance and Possible Detection with Double (Fluorescent and Spin) Indicator Reagents. Photochem. Photobiol. 2006, 82, 1211–1218. [Google Scholar] [CrossRef]
- Gollmer, A.; Arnbjerg, J.; Blaikie, F.H.; Pedersen, B.W.; Breitenbach, T.; Daasbjerg, K.; Glasius, M.; Ogilby, P.R. Singlet Oxygen Sensor Green®: Photochemical Behavior in Solution and in a Mammalian Cell. Photochem. Photobiol. 2011, 87, 671–679. [Google Scholar] [CrossRef]
- Kim, S.; Fujitsuka, M.; Majima, T. Photochemistry of Singlet Oxygen Sensor Green. J. Phys. Chem. B 2013, 117, 13985–13992. [Google Scholar] [CrossRef]
- Flors, C.; Fryer, M.J.; Waring, J.; Reeder, B.; Bechtold, U.; Mullineaux, P.M.; Nonell, S.; Wilson, M.T.; Baker, N.R. Imaging the production of singlet oxygen in vivo using a new fluorescent sensor, Singlet Oxygen Sensor Green(R). J. Exp. Bot. 2006, 57, 1725–1734. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Carter, P.J.H.; Laan, A.C.; Eelkema, R.; Denkova, A.G. Singlet Oxygen Sensor Green is not a Suitable Probe for 1O2 in the Presence of Ionizing Radiation. Sci. Rep. 2019, 9, 8393. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, S.K.; Holmehave, J.; Blaikie, F.H.; Gollmer, A.; Breitenbach, T.; Jensen, H.H.; Ogilby, P.R. Aarhus Sensor Green: A Fluorescent Probe for Singlet Oxygen. J. Org. Chem. 2014, 79, 3079–3087. [Google Scholar] [CrossRef]
- Yazdani, M. Concerns in the application of fluorescent probes DCDHF-DA, DHR 123 and DHE to measure reactive oxygen species in vitro. Toxicol. Vitr. 2015, 30, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C. The challenges of using fluorescent probes to detect and quantify specific reactive oxygen species in living cells. Biochim. Biophys. Acta (BBA) Gen. Subj. 2014, 1840, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Yu, S.; Jiang, Z.; Liang, C.; Yu, W.; Li, J.; Du, X.; Wang, H.; Gao, X.; Wang, X. N-Acetyl-Serotonin Protects HepG2 Cells from Oxidative Stress Injury Induced by Hydrogen Peroxide. Oxidative Med. Cell Longev. 2014, 2014, 310504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prokhorova, I.V.; Gorbach, O.I.; Yakshibaeva, Y.R.; Shliakhtova, N.; Solyanik, G.I. ROS production by circulating phagocytes and Guerin carcinoma resistance to cisplatin. Exp. Oncol. 2021, 43, 26–30. [Google Scholar] [CrossRef]
- Qiao, Y.; Zhou, Y.; Zhang, X.; Faulkner, S.; Liu, H.; Wang, L. Toxic effects of triphenyltin on the development of zebrafish (Danio rerio) embryos. Sci. Total Environ. 2023, 885, 163783. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.-R.; Lee, G.-A.; Kim, G.-S.; Hwang, K.-A.; Choi, K.-C. Phytochemical-induced reactive oxygen species and endoplasmic reticulum stress-mediated apoptosis and differentiation in malignant melanoma cells. Phytomedicine 2018, 39, 100–110. [Google Scholar] [CrossRef]
- Przyborowski, K.; Kurpinska, A.; Wojkowska, D.; Kaczara, P.; Suraj-Prazmowska, J.; Karolczak, K.; Malinowska, A.; Pelesz, A.; Kij, A.; Kalvins, I.; et al. Protein disulfide isomerase-A1 regulates intraplatelet reactive oxygen species–thromboxane A2-dependent pathway in human platelets. J. Thromb. Haemost. 2022, 20, 157–169. [Google Scholar] [CrossRef]
- Kose, T.; Sharp, P.A.; Latunde-Dada, G.O. Upregulation of Nrf2 Signalling and the Inhibition of Erastin-Induced Ferroptosis by Ferulic Acid in MIN6 Cells. Int. J. Mol. Sci. 2022, 23, 15886. [Google Scholar] [CrossRef]
- Rachakhom, W.; Banjerdpongchai, R. Effect of Calomelanone, a Dihydrochalcone Analogue, on Human Cancer Apoptosis/Regulated Cell Death in an In Vitro Model. BioMed Res. Int. 2020, 2020, 4926821. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.W.; Choi, S.; Song, H.; Lee, M.J.; Kwon, J.E.; Lee, H.A.R.; Kim, K. Effect of Hyperbaric Oxygen Therapy on Acute Liver Injury and Survival in a Rat Cecal Slurry Peritonitis Model. Life 2020, 10, 283. [Google Scholar] [CrossRef]
- Probes for Reactive Oxygen Species, Including Nitric Oxide—Chapter 18|Thermo Fisher Scientific—ES. Available online: https://www.thermofisher.com/es/es/home/references/molecular-probes-the-handbook/probes-for-reactive-oxygen-species-including-nitric-oxide.html (accessed on 27 April 2023).
- Razavi, M.; Ren, G.; Wang, J.; Kimura, R.; Thakor, A.S. Imaging Tumor Oxidative Stress with Surface Enhanced Raman Scattering Gold Nanoparticles. J. Biomed. Nanotechnol. 2019, 15, 2130–2141. [Google Scholar] [CrossRef]
- Prolo, C.; Rios, N.; Piacenza, L.; Álvarez, M.N.; Radi, R. Fluorescence and chemiluminescence approaches for peroxynitrite detection. Free Radic. Biol. Med. 2018, 128, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Sadowska-Bartosz, I.; Gajewska, A.; Skolimowski, J.; Szewczyk, R.; Bartosz, G. Nitroxides protect against peroxynitrite-induced nitration and oxidation. Free Radic. Biol. Med. 2015, 89, 1165–1175. [Google Scholar] [CrossRef]
- Miller, E.W.; Albers, A.E.; Pralle, A.; Isacoff, E.Y.; Chang, C.J. Boronate-Based Fluorescent Probes for Imaging Cellular Hydrogen Peroxide. J. Am. Chem. Soc. 2005, 127, 16652–16659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickinson, B.C.; Huynh, C.; Chang, C.J. A Palette of Fluorescent Probes with Varying Emission Colors for Imaging Hydrogen Peroxide Signaling in Living Cells. J. Am. Chem. Soc. 2010, 132, 5906–5915. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Aleyasin, H.; Dickinson, B.C.; Haskew-Layton, R.E.; Ratan, R.R. Recent advances in hydrogen peroxide imaging for biological applications. Cell Biosci. 2014, 4, 64. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Aleyasin, H.; Howard, S.S.; Dickinson, B.C.; Lin, V.S.; Haskew-Layton, R.E.; Xu, C.; Chen, Y.; Ratan, R.R. Two-photon fluorescence imaging of intracellular hydrogen peroxide with chemoselective fluorescent probes. J. Biomed. Opt. 2013, 18, 106002. [Google Scholar] [CrossRef] [Green Version]
- Albers, A.E.; Okreglak, V.S.; Chang, C.J. A FRET-Based Approach to Ratiometric Fluorescence Detection of Hydrogen Peroxide. J. Am. Chem. Soc. 2006, 128, 9640–9641. [Google Scholar] [CrossRef] [PubMed]
- Jávega, B.; Herrera, G.; O’connor, J.-E. Flow Cytometric Analysis of Oxidative Stress in Escherichia coli B Strains Deficient in Genes of the Antioxidant Defence. Int. J. Mol. Sci. 2022, 10, 6537. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.-F.; Gong, Y.-X.; Li, H.-F.; Sun, F.-L.; Li, W.-L.; Chen, D.-Q.; Xie, D.-P.; Ren, C.-X.; Guo, X.-Y.; Wang, Z.-Y.; et al. Curcumin Activates ROS Signaling to Promote Pyroptosis in Hepatocellular Carcinoma HepG2 Cells. In Vivo 2021, 35, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.-Y.; Duchen, M.R. A Plate Reader-Based Measurement of the Cellular ROS Production Using Dihydroethidium and MitoSOX. Methods Mol. Biol. 2022, 2497, 333–337. [Google Scholar] [CrossRef]
- Walrand, S.; Valeix, S.; Rodriguez, C.; Ligot, P.; Chassagne, J.; Vasson, M.-P. Flow cytometry study of polymorphonuclear neutrophil oxidative burst: A comparison of three fluorescent probes. Clin. Chim. Acta 2003, 331, 103–110. [Google Scholar] [CrossRef]
- Talib, J.; Maghzal, G.J.; Cheng, D.; Stocker, R. Detailed protocol to assess in vivo and ex vivo myeloperoxidase activity in mouse models of vascular inflammation and disease using hydroethidine. Free Radic. Biol. Med. 2016, 97, 124–135. [Google Scholar] [CrossRef]
- Barbacanne, M.-A.; Souchard, J.-P.; Darblade, B.; Iliou, J.-P.; Nepveu, F.; Pipy, B.; Bayard, F.; Arnal, J.-F. Detection of superoxide anion released extracellularly by endothelial cells using cytochrome c reduction, ESR, fluorescence and lucigenin-enhanced chemiluminescence techniques. Free Radic. Biol. Med. 2000, 29, 388–396. [Google Scholar] [CrossRef]
- Münzel, T.; Afanas’ev, I.B.; Kleschyov, A.L.; Harrison, D.G. Detection of Superoxide in Vascular Tissue. Arter. Thromb. Vasc. Biol. 2002, 22, 1761–1768. [Google Scholar] [CrossRef] [Green Version]
- Tarpey, M.M.; Wink, D.A.; Grisham, M.B. Methods for detection of reactive metabolites of oxygen and nitrogen: In vitro and in vivo considerations. Am. J. Physiol. Integr. Comp. Physiol. 2004, 286, R431–R444. [Google Scholar] [CrossRef] [Green Version]
- Guo, T.L.; Miller, M.A.; Shapiro, I.M.; Shenker, B.J. Mercuric Chloride Induces Apoptosis in Human T Lymphocytes: Evidence of Mitochondrial Dysfunction. Toxicol. Appl. Pharmacol. 1998, 153, 250–257. [Google Scholar] [CrossRef]
- Le, S.B.; Hailer, M.K.; Buhrow, S.; Wang, Q.; Flatten, K.; Pediaditakis, P.; Bible, K.C.; Lewis, L.D.; Sausville, E.A.; Pang, Y.-P.; et al. Inhibition of Mitochondrial Respiration as a Source of Adaphostin-induced Reactive Oxygen Species and Cytotoxicity. J. Biol. Chem. 2007, 282, 8860–8872. [Google Scholar] [CrossRef] [Green Version]
- De Biasi, S.; Gibellini, L.; Bianchini, E.; Nasi, M.; Pinti, M.; Salvioli, S.; Cossarizza, A. Quantification of mitochondrial reactive oxygen species in living cells by using multi-laser polychromatic flow cytometry. Cytom. Part A 2016, 89, 1106–1110. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.M.; Janes, M.S.; Beckman, J.S. The selective detection of mitochondrial superoxide by live cell imaging. Nat. Protoc. 2008, 3, 941–947. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, J.; Kalyanaraman, B. Hydroethidine- and MitoSOX-derived red fluorescence is not a reliable indicator of intracellular superoxide formation: Another inconvenient truth. Free Radic. Biol. Med. 2010, 48, 983–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Zhang, G.; Yang, T.; Gan, J.; Xu, L.; Yang, H. A flow-cytometry-based protocol for detection of mitochondrial ROS production under hypoxia. STAR Protoc. 2021, 2, 100466. [Google Scholar] [CrossRef]
- Ahn, H.-Y.; Fairfull-Smith, K.E.; Morrow, B.J.; Lussini, V.; Kim, B.; Bondar, M.V.; Bottle, S.E.; Belfield, K.D. Two-Photon Fluorescence Microscopy Imaging of Cellular Oxidative Stress Using Profluorescent Nitroxides. J. Am. Chem. Soc. 2012, 134, 4721–4730. [Google Scholar] [CrossRef] [Green Version]
- Davison, C.A.; Durbin, S.M.; Thau, M.R.; Zellmer, V.R.; Chapman, S.E.; Diener, J.; Wathen, C.; Leevy, W.M.; Schafer, Z.T. Antioxidant Enzymes Mediate Survival of Breast Cancer Cells Deprived of Extracellular Matrix. Cancer Res. 2013, 73, 3704–3715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deloughery, Z.; Luczak, M.W.; Zhitkovich, A. Monitoring Cr Intermediates and Reactive Oxygen Species with Fluorescent Probes during Chromate Reduction. Chem. Res. Toxicol. 2014, 27, 843–851. [Google Scholar] [CrossRef]
- Davila, M.P.; Muñoz, P.M.; Tapia, J.A.; Ortega-Ferrusola, C.; Da Silva, C.M.B.; Peña, F.J. Inhibition of Mitochondrial Complex I Leads to Decreased Motility and Membrane Integrity Related to Increased Hydrogen Peroxide and Reduced ATP Production, while the Inhibition of Glycolysis Has Less Impact on Sperm Motility. PLoS ONE 2015, 10, e0138777. [Google Scholar] [CrossRef] [Green Version]
- mp10422.pdf (fishersci.com). Available online: https://assets.fishersci.com/TFS-Assets/LSG/manuals/mp10422.pdf?_ga=2.231040351.1726096505.1681672212-1827320652.1681672212 (accessed on 27 April 2023).
- ROS-ID® Total ROS/Superoxide detection kit—ENZ-51010—Enzo Life Sciences. Available online: https://www.enzolifesciences.com/ENZ-51010/ros-id-total-ros-superoxide-detection-kit/ (accessed on 27 April 2023).
- Egawa, N.; Tanaka, T.; Matsufuji, S.; Yamada, K.; Ito, K.; Kitagawa, H.; Okuyama, K.; Kitajima, Y.; Noshiro, H. Antitumor effects of low-dose tipifarnib on the mTOR signaling pathway and reactive oxygen species production in HIF-1α-expressing gastric cancer cells. FEBS Open Bio 2021, 11, 1465–1475. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Liu, S.; Wang, X.; Deng, X.; Fan, Y.; Sun, C.; Wang, Y.; Mehta, J.L. Hemodynamic Shear Stress via ROS Modulates PCSK9 Expression in Human Vascular Endothelial and Smooth Muscle Cells and Along the Mouse Aorta. Antioxid. Redox Signal. 2015, 22, 760–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedley, D.; Chow, S. Flow cytometric measurement of lipid peroxidation in vital cells using parinaric acid. Cytometry 1992, 13, 686–692. [Google Scholar] [CrossRef]
- Zhan, B.; Arumugam, S.; Kennedy, M.W.; Tricoche, N.; Lian, L.-Y.; Asojo, O.A.; Bennuru, S.; Bottazzi, M.E.; Hotez, P.J.; Lustigman, S.; et al. Ligand binding properties of two Brugia malayi fatty acid and retinol (FAR) binding proteins and their vaccine efficacies against challenge infection in gerbils. PLoS Negl. Trop. Dis. 2018, 12, e0006772. [Google Scholar] [CrossRef]
- Oliveira, A.C.; Filipe, H.A.L.; Loura, L.M.S. Fluorescent Probes cis- and trans-Parinaric Acids in Fluid and Gel Lipid Bilayers: A Molecular Dynamics Study. Molecules 2023, 28, 2241. [Google Scholar] [CrossRef]
- Yoshida, Y.; Shimakawa, S.; Itoh, N.; Niki, E. Action of DCFH and BODIPY as a probe for radical oxidation in hydrophilic and lipophilic domain. Free Radic. Res. 2003, 37, 861–872. [Google Scholar] [CrossRef]
- Brouwers, J.F.; Gadella, B.M. In situ detection and localization of lipid peroxidation in individual bovine sperm cells. Free Radic. Biol. Med. 2003, 35, 1382–1391. [Google Scholar] [CrossRef] [PubMed]
- Cheloni, G.; Slaveykova, V.I. Optimization of the C11-BODIPY581/591dye for the determination of lipid oxidation in Chlamydomonas reinhardtii by flow cytometry. Cytom. Part A 2013, 83, 952–961. [Google Scholar] [CrossRef] [PubMed]
- Peluso, I.; Adorno, G.; Raguzzini, A.; Urban, L.; Ghiselli, A.; Serafini, M. A new flow cytometry method to measure oxidative status: The Peroxidation of Leukocytes Index Ratio (PLIR). J. Immunol. Methods 2013, 390, 113–120. [Google Scholar] [CrossRef]
- Donato, M.T.; Martínez-Romero, A.; Jiménez, N.; Negro, A.; Herrera, G.; Castell, J.V.; O’connor, J.-E.; Gómez-Lechón, M.J. Cytometric analysis for drug-induced steatosis in HepG2 cells. Chem. Interact. 2009, 181, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Makrigiorgos, G.; Kassis, A.I.; Mahmood, A.; Bump, E.A.; Savvides, P. Novel Fluorescein-Based Flow-Cytometric Method For Detection of Lipid Peroxidation. Free Radic. Biol. Med. 1997, 22, 93–100. [Google Scholar] [CrossRef]
- du Plessis, L.; Laubscher, P.J.; Jooste, J.; du Plessis, J.; Franken, A.; Van Aarde, M.N.; Eloff, F.C. Flow Cytometric Analysis of the Oxidative Status in Human Peripheral Blood Mononuclear Cells of Workers Exposed to Welding Fumes. J. Occup. Environ. Hyg. 2010, 7, 367–374. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, T.; Li, J.; Xia, M.; Li, Y.; Wang, X.; Liu, C.; Zheng, T.; Chen, R.; Kan, D.; et al. Oxidative Stress and 4-hydroxy-2-nonenal (4-HNE): Implications in the Pathogenesis and Treatment of Aging-related Diseases. J. Immunol. Res. 2022, 2022, 2233906. [Google Scholar] [CrossRef]
- Sharma, S.; Sharma, P.; Bailey, T.; Bhattarai, S.; Subedi, U.; Miller, C.; Ara, H.; Kidambi, S.; Sun, H.; Panchatcharam, M.; et al. Electrophilic Aldehyde 4-Hydroxy-2-Nonenal Mediated Signaling and Mitochondrial Dysfunction. Biomolecules 2022, 12, 1555. [Google Scholar] [CrossRef]
- Spickett, C.M. The lipid peroxidation product 4-hydroxy-2-nonenal: Advances in chemistry and analysis. Redox Biol. 2013, 1, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, P.M.; Ferrusola, C.O.; Vizuete, G.; Dávila, M.P.; Martinez, H.R.; Peña, F.J. Depletion of Intracellular Thiols and Increased Production of 4-Hydroxynonenal that Occur During Cryopreservation of Stallion Spermatozoa Lead to Caspase Activation, Loss of Motility, and Cell Death1. Biol. Reprod. 2015, 93, 143. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.M.; Correa, R.; Rosenberg, S.M.; Hastings, P.J. Persistent damaged bases in DNA allow mutagenic break repair in Escherichia coli. PLoS Genet. 2017, 13, e1006733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Møller, P.; Roursgaard, M. Biomarkers of DNA Oxidation Products: Links to Exposure and Disease in Public Health Studies. Chem. Res. Toxicol. 2021, 34, 2235–2250. [Google Scholar] [CrossRef] [PubMed]
- Oxy dna assay kit|Sigma-Aldrich (sigmaaldrich.com). Available online: https://www.sigmaaldrich.com/ES/en/search/oxy-dna-assay-kit?focus=products&page=1&perpage=30&sort=relevance&term=oxy%20dna%20assay%20kit&type=product (accessed on 27 April 2023).
- Nagy, S.; Kakasi, B.; Bercsényi, M.; Information, R. Flow cytometric detection of oxidative DNA damage in fish spermatozoa exposed to cadmium—Short communication. Acta Veter-Hung. 2016, 64, 120–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esperanza, M.; Cid, Á.; Herrero, C.; Rioboo, C. Acute effects of a prooxidant herbicide on the microalga Chlamydomonas reinhardtii: Screening cytotoxicity and genotoxicity endpoints. Aquat. Toxicol. 2015, 165, 210–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cambi, M.; Tamburrino, L.; Marchiani, S.; Olivito, B.; Azzari, C.; Forti, G.; Baldi, E.; Muratori, M. Development of a specific method to evaluate 8-hydroxy,2-deoxyguanosine in sperm nuclei: Relationship with semen quality in a cohort of 94 subjects. Reproduction 2013, 145, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, C.; Meseguer, M.; García-Herrero, S.; Gil-Salom, M.; O’Connor, J.E.; Garrido, N. Relevance of testicular sperm DNA oxidation for the outcome of ovum donation cycles. Fertil. Steril. 2010, 94, 979–988. [Google Scholar] [CrossRef]
- Meseguer, M.; Martínez-Conejero, J.; O’Connor, J.E.; Pellicer, A.; Remohí, J.; Garrido, N. The significance of sperm DNA oxidation in embryo development and reproductive outcome in an oocyte donation program: A new model to study a male infertility prognostic factor. Fertil. Steril. 2008, 89, 1191–1199. [Google Scholar] [CrossRef]
- da Silva, C.B.; Ortega-Ferrusola, C.; Morrell, J.; Martínez, H.R.; Peña, F. Flow Cytometric Chromosomal Sex Sorting of Stallion Spermatozoa Induces Oxidative Stress on Mitochondria and Genomic DNA. Reprod. Domest. Anim. 2016, 51, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Durand, R.E.; Olive, P.L. Flow Cytometry Techniques for Studying Cellular Thiols. Radiat. Res. 1983, 95, 456. [Google Scholar] [CrossRef]
- Treumer, J.; Valet, G. Flow-cytometric determination of glutathione alterations in vital cells by o-phthaldialdehyde (OPT) staining. Exp. Cell Res. 1986, 163, 518–524. [Google Scholar] [CrossRef]
- O’Connor, J.E.; Kimler, B.F.; Morgan, M.C.; Tempas, K.J. A flow cytometric assay for intracellular nonprotein thiols using mercury orange. Cytometry 1988, 9, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Cell Viability Assays|Thermo Fisher Scientific—ES. Available online: https://www.thermofisher.com/es/es/home/life-science/cell-analysis/cell-viability-and-regulation/cell-viability.html (accessed on 27 April 2023).
- Nair, S.; Singh, S.V.; Krishan, A. Flow cytometric monitoring of glutathione content and anthracycline retention in tumor cells. Cytometry 1991, 12, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Hedley, D.W.; Chow, S. Evaluation of methods for measuring cellular glutathione content using flow cytometry. Cytometry 1994, 15, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Skindersoe, M.E.; Kjaerulff, S. Comparison of three thiol probes for determination of apoptosis-related changes in cellular redox status. Cytom. Part A 2014, 85, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Jemaà, M.; Fezai, M.; Bissinger, R.; Lang, F. Methods Employed in Cytofluorometric Assessment of Eryptosis, the Suicidal Erythrocyte Death. Cell Physiol. Biochem. 2017, 43, 431–444. [Google Scholar] [CrossRef] [Green Version]
- Keller, A.; Mohamed, A.; Dröse, S.; Brandt, U.; Fleming, I.; Brandes, R.P. Analysis of Dichlorodihydrofluorescein and Dihydrocalcein as Probes for the Detection of Intracellular Reactive Oxygen Species. Free Radic. Res. 2004, 38, 1257–1267. [Google Scholar] [CrossRef]
- Swift, L.M.; Sarvazyan, N. Localization of dichlorofluorescin in cardiac myocytes: Implications for assessment of oxidative stress. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, H982–H990. [Google Scholar] [CrossRef]
- Balaguer, S.; Diaz, L.; Gomes, A.; Herrera, G.; O’Connor, J.-E.; Urios, A.; Felipo, V.; Montoliu, C. Real-time cytometric assay of nitric oxide and superoxide interaction in peripheral blood monocytes: A no-wash, no-lyse kinetic method. Cytom. Part B Clin. Cytom. 2017, 92, 211–217. [Google Scholar] [CrossRef]
- Saengkhae, C.; Loetchutinat, C.; Garnier-Suillerot, A. Kinetic analysis of fluorescein and dihydrofluorescein effluxes in tumour cells expressing the multidrug resistance protein, MRP1. Biochem. Pharmacol. 2003, 65, 969–977. [Google Scholar] [CrossRef]
- Grzelak, A.; Rychlik, B.; Bartosz, G. Reactive oxygen species are formed in cell culture media. Acta Biochim. Pol. 2000, 47, 1197–1198. [Google Scholar]
- Petasne, R.G.; Zika, R.G. Fate of superoxide in coastal sea water. Nature 1987, 325, 516–518. [Google Scholar] [CrossRef]
- Hopwood, M.J.; Rapp, I.; Schlosser, C.; Achterberg, E.P. Hydrogen peroxide in deep waters from the Mediterranean Sea, South Atlantic and South Pacific Oceans. Sci. Rep. 2017, 7, 43436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramaniam, R.; Fan, X.-J.; Scivittaro, V.; Yang, J.; Ha, C.-E.; Petersen, C.E.; Surewicz, W.K.; Bhagavan, N.V.; Weiss, M.F.; Monnier, V.M. Cellular Oxidant Stress and Advanced Glycation Endproducts of Albumin: Caveats of the Dichlorofluorescein Assay*. Arch. Biochem. Biophys. 2002, 400, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Chignell, C.F.; Sik, R.H. A photochemical study of cells loaded with 2′,7′-dichlorofluorescin: Implications for the detection of reactive oxygen species generated during UVA irradiation. Free Radic. Biol. Med. 2003, 34, 1029–1034. [Google Scholar] [CrossRef]
- Sikora, A.; Zielonka, J.; Lopez, M.; Joseph, J.; Kalyanaraman, B. Direct oxidation of boronates by peroxynitrite: Mechanism and implications in fluorescence imaging of peroxynitrite. Free Radic. Biol. Med. 2009, 47, 1401–1407. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Kalivendi, S.; Zhang, H.; Joseph, J.; Nithipatikom, K.; Vásquez-Vivar, J.; Kalyanaraman, B. Superoxide reacts with hydroethidine but forms a fluorescent product that is distinctly different from ethidium: Potential implications in intracellular fluorescence detection of superoxide. Free Radic. Biol. Med. 2003, 34, 1359–1368. [Google Scholar] [CrossRef]
- Zhao, H.; Joseph, J.; Fales, H.M.; Sokoloski, E.A.; Levine, R.L.; Vasquez-Vivar, J.; Kalyanaraman, B. Detection and characterization of the product of hydroethidine and intracellular superoxide by HPLC and limitations of fluorescence. Proc. Natl. Acad. Sci. USA 2005, 102, 5727–5732. [Google Scholar] [CrossRef] [Green Version]
- Papapostolou, I.; Patsoukis, N.; Georgiou, C.D. The fluorescence detection of superoxide radical using hydroethidine could be complicated by the presence of heme proteins. Anal. Biochem. 2004, 332, 290–298. [Google Scholar] [CrossRef]
- Roychowdhury, S.; Luthe, A.; Keilhoff, G.; Wolf, G.; Horn, T.F. Oxidative stress in glial cultures: Detection by DAF-2 fluorescence used as a tool to measure peroxynitrite rather than nitric oxide. Glia 2002, 38, 103–114. [Google Scholar] [CrossRef]
- VanderVen, A.; Mier, P.; Peters, W.; Dolstra, H.; van Erp, P.; Koopmans, P.; VanderMeer, J. Monochlorobimane Does Not Selectively Label Glutathione in Peripheral Blood Mononuclear Cells. Anal. Biochem. 1994, 217, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Woolley, J.; Stanicka, J.; Cotter, T. Recent advances in reactive oxygen species measurement in biological systems. Trends Biochem. Sci. 2013, 38, 556–565. [Google Scholar] [CrossRef]
- Jávega, B. Aplicación de la Citometría De Flujo Al Estudio En Tiempo Real De Las Interacciones Entre Especies Reactivas De Oxígeno Y Ni-Trógeno En El Estrés Oxidativo Inducido Por Xenobióticos. Ph.D. Thesis, Universitat de València, Valencia, Spain, 2022. [Google Scholar]
- Nimse, S.B.; Pal, D. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef] [Green Version]
- Zinovkin, R.A.; Lyamzaev, K.G.; Chernyak, B.V. Current perspectives of mitochondria-targeted antioxidants in cancer prevention and treatment. Front. Cell Dev. Biol. 2023, 11, 1048177. [Google Scholar] [CrossRef]
- Sakuma, S.; Abe, M.; Kohda, T.; Fujimoto, Y. Hydrogen peroxide generated by xanthine/xanthine oxidase system represses the proliferation of colorectal cancer cell line Caco-2. J. Clin. Biochem. Nutr. 2015, 56, 15–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicoletto, R.E.; Ofner, C.M. Cytotoxic mechanisms of doxorubicin at clinically relevant concentrations in breast cancer cells. Cancer Chemother. Pharmacol. 2022, 89, 285–311. [Google Scholar] [CrossRef]
- Hall, S.; McDermott, C.; Anoopkumar-Dukie, S.; McFarland, A.J.; Forbes, A.; Perkins, A.V.; Davey, A.K.; Chess-Williams, R.; Kiefel, M.J.; Arora, D.; et al. Cellular Effects of Pyocyanin, a Secreted Virulence Factor of Pseudomonas aeruginosa. Toxins 2016, 8, 236. [Google Scholar] [CrossRef] [Green Version]
- Lew, S.Y.; Hisam, N.S.M.; Phang, M.W.L.; Rahman, S.N.S.A.; Poh, R.Y.Y.; Lim, S.H.; Kamaruzzaman, M.A.; Chau, S.C.; Tsui, K.C.; Lim, L.W.; et al. Adenosine Improves Mitochondrial Function and Biogenesis in Friedreich’s Ataxia Fibroblasts Following L-Buthionine Sulfoximine-Induced Oxidative Stress. Biology 2023, 12, 559. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, Ł.; Kepinska, M.; Milnerowicz, H. Inhibition of copper-zinc superoxide dismutase activity by selected environmental xenobiotics. Environ. Toxicol. Pharmacol. 2018, 58, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Mahaseth, T.; Kuzminov, A. Potentiation of hydrogen peroxide toxicity: From catalase inhibition to stable DNA-iron complexes. Mutat. Res. Rev. Mutat. Res. 2017, 773, 274–281. [Google Scholar] [CrossRef]
- Yousef, A.; Saleh, I.; Abd-Allah, A.R.; Elnagar, M.R.; Akool, E.-S. PEG-SOD attenuates the mitogenic ERK1/2 signaling cascade induced by cyclosporin A in the liver and kidney of albino mice. Chem. Interact. 2020, 330, 109245. [Google Scholar] [CrossRef]
- Herrera, G.; Martinez, A.; Blanco, M. Assessment ofEscherichia coli B with enhanced permeability to fluorochromes for flow cytometric assays of bacterial cell function. Cytometry 2002, 49, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Herrera, G.; Martínez, A.; O’Cornor, J.; Blanco, M. Functional Assays of Oxidative Stress Using Genetically Engineered Escherichia coli Strains. Curr. Protoc. Cytom. 2003, 24, 11–16. [Google Scholar] [CrossRef]
- Yoon, S.H.; Jeong, H.; Kwon, S.-K.; Kim, J.F. Genomics, Biological Features, and Biotechnological Applications of Escherichia coli B: “Is B for better?!”. In Systems Biology and Biotechnology of Escherichia coli; Lee, S.Y., Ed.; Springer: Dordrecht, The Netherlands, 2009; pp. 1–17. [Google Scholar] [CrossRef]
- Martínez, A.; Urios, A.; Blanco, M. Mutagenicity of 80 chemicals in Escherichia coli tester strains IC203, deficient in OxyR, and its oxyR+ parent WP2 uvrA/pKM101: Detection of 31 oxidative mutagens. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2000, 467, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Felim, A.; Urios, A.; Neudörffer, A.; Herrera, G.; Blanco, M.; Largeron, M. Bacterial Plate Assays and Electrochemical Methods: An Efficient Tandem for Evaluating the Ability of Catechol−Thioether Metabolites of MDMA (“Ecstasy”) to Induce Toxic Effects through Redox-Cycling. Chem. Res. Toxicol. 2007, 20, 685–693. [Google Scholar] [CrossRef]
- Tugcu, F.T.; Turhan, K.; Karadayi, M.; Gulluce, M. Genotoxic evaluation of newly synthesized iminothiazolidinones. Toxicol. Ind. Health 2017, 33, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Han, M.-J.; Jeong, H.; Lee, C.H.; Xia, X.-X.; Lee, D.-H.; Shim, J.H.; Lee, S.Y.; Oh, T.K.; Kim, J.F. Comparative multi-omics systems analysis of Escherichia coli strains B and K-12. Genome Biol. 2012, 13, 37. [Google Scholar] [CrossRef] [Green Version]
- Sen, A.; Imlay, J.A. How Microbes Defend Themselves From Incoming Hydrogen Peroxide. Front. Immunol. 2021, 12, 667343. [Google Scholar] [CrossRef]
- Wardman, P. Methods to Measure the Reactivity of Peroxynitrite-Derived Oxidants Toward Reduced Fluoresceins and Rhodamines. Methods Enzymol. 2008, 441, 261–282. [Google Scholar] [CrossRef]
- Fukui, M.; Choi, H.J.; Zhu, B.T. Rapid generation of mitochondrial superoxide induces mitochondrion-dependent but caspase-independent cell death in hippocampal neuronal cells that morphologically resembles necroptosis. Toxicol. Appl. Pharmacol. 2012, 262, 156–166. [Google Scholar] [CrossRef] [Green Version]
- Clothier, R.; Gómez-Lechón, M.; Kinsner-Ovaskainen, A.; Kopp-Schneider, A.; O’connor, J.; Prieto, P.; Stanzel, S. Comparative analysis of eight cytotoxicity assays evaluated within the ACuteTox Project. Toxicol. Vitr. 2013, 27, 1347–1356. [Google Scholar] [CrossRef]
- Porzani, S.J.; Lima, S.T.; Metcalf, J.S.; Nowruzi, B. In Vivo and In Vitro Toxicity Testing of Cyanobacterial Toxins: A Mini-Review. Rev. Environ. Contam. Toxicol. 2021, 258, 109–150. [Google Scholar] [CrossRef] [PubMed]
- Rooprai, H.K.; Lawrence, P.; Keshavarz, S.; Yashod, P.; Gullan, R.W.; Selway, R.P.; Davies, D. DRAQ7 as an Alternative to MTT Assay for Measuring Viability of Glioma Cells Treated with Polyphenols. Anticancer Res. 2020, 40, 5427–5436. [Google Scholar] [CrossRef] [PubMed]
- McBee, M.E.; Chionh, Y.H.; Sharaf, M.L.; Ho, P.; Cai, M.W.L.; Dedon, P.C. Production of Superoxide in Bacteria Is Stress- and Cell State-Dependent: A Gating-Optimized Flow Cytometry Method that Minimizes ROS Measurement Artifacts with Fluorescent Dyes. Front. Microbiol. 2017, 8, 459. [Google Scholar] [CrossRef] [Green Version]
- Spidlen, J.; Brinkman, R.R. Use FlowRepository to share your clinical data upon study publication. Cytom. Part B Clin. Cytom. 2018, 94, 196–198. [Google Scholar] [CrossRef]
- Spidlen, J.; Breuer, K.; Brinkman, R. Preparing a Minimum Information about a Flow Cytometry Experiment (MIFlowCyt) Compliant Manuscript Using the International Society for Advancement of Cytometry (ISAC) FCS File Repository (FlowRepository.org). Curr. Protoc. Cytom. 2012, 61, 10–18. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Critical Points and Limitations |
|---|
| Identification of blood cells in whole-blood samples without lysis of erythrocytes |
| Preparation of single-cell suspensions from adherent cell models |
| Maintenance of viability and functional competence of the cells along sample preparation and experiment performance |
| Identification of the optimal incubation time and concentration for staining |
| Access of fluorogenic substrates to intracellular sites or intracellular processes |
| Retention of fluorogenic substrates and oxidized fluorescent probes |
| Preparation of single-cell suspensions from adherent cell models |
| Lack of absolute specificity of fluorogenic substrates for specific RONS |
| Interference of the probes with ROS biology or ROS-relevant cell functions |
| Selection of the time window for kinetic assays |
| Assay calibration for data expression in biochemical units |
| Adapted from [53] |
| Biological Process of Interest and Experimental In Vitro Setting | ||||||
|---|---|---|---|---|---|---|
| Peroxidative Activity | Redox Cycling | Antioxidant Defense | ||||
| Probe | Mito-PY1 | H2DCF-DA | DHRH123 | MitoSOX Red | HE | Monochloro-bimane |
| Viability Stain | DAPI or PI | DAPI or PI | DAPI or PI | DAPI | DAPI | PI |
| Positive control | H2O2 | t-BOOH | CHP | Menadione | Plumbagin | N-Acetyl cysteine GSH-ester |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jávega, B.; Herrera, G.; Martínez-Romero, A.; O’Connor, J.-E. Flow Cytometry of Oxygen and Oxygen-Related Cellular Stress. Oxygen 2023, 3, 222-255. https://doi.org/10.3390/oxygen3020016
Jávega B, Herrera G, Martínez-Romero A, O’Connor J-E. Flow Cytometry of Oxygen and Oxygen-Related Cellular Stress. Oxygen. 2023; 3(2):222-255. https://doi.org/10.3390/oxygen3020016
Chicago/Turabian StyleJávega, Beatriz, Guadalupe Herrera, Alicia Martínez-Romero, and José-Enrique O’Connor. 2023. "Flow Cytometry of Oxygen and Oxygen-Related Cellular Stress" Oxygen 3, no. 2: 222-255. https://doi.org/10.3390/oxygen3020016