

Assessment of Embryonic and Larval Development of Nile Tilapia under the Traditional and Re-Circulatory Thermostatic System in Relation to Climatic and Water Quality Variations

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Study Area
2.3. Collection and Domestication of O. niloticus for Gonadal Maturation
2.4. Collection, Rearing of Eggs and Larvae of O. niloticus
2.5. Observation of Embryonic Developmental Stages of O. niloticus
2.6. Determination of Hatching and Survival Rates of O. niloticus
2.7. Observation of Larval Development and Growth Parameters of O. niloticus
2.8. Determination of Climatic Variables and Water Quality Parameters in Traditional Hatchery and Re-Circulatory Thermostatic Systems
2.9. Relationship of Climatic Variables, Water Quality Parameters and Morphometric Change of O. niloticus Larvae under Traditional Hatchery System
2.10. Statistical Analysis
3. Results
3.1. Observation of Embryonic Development Stages of O. niloticus
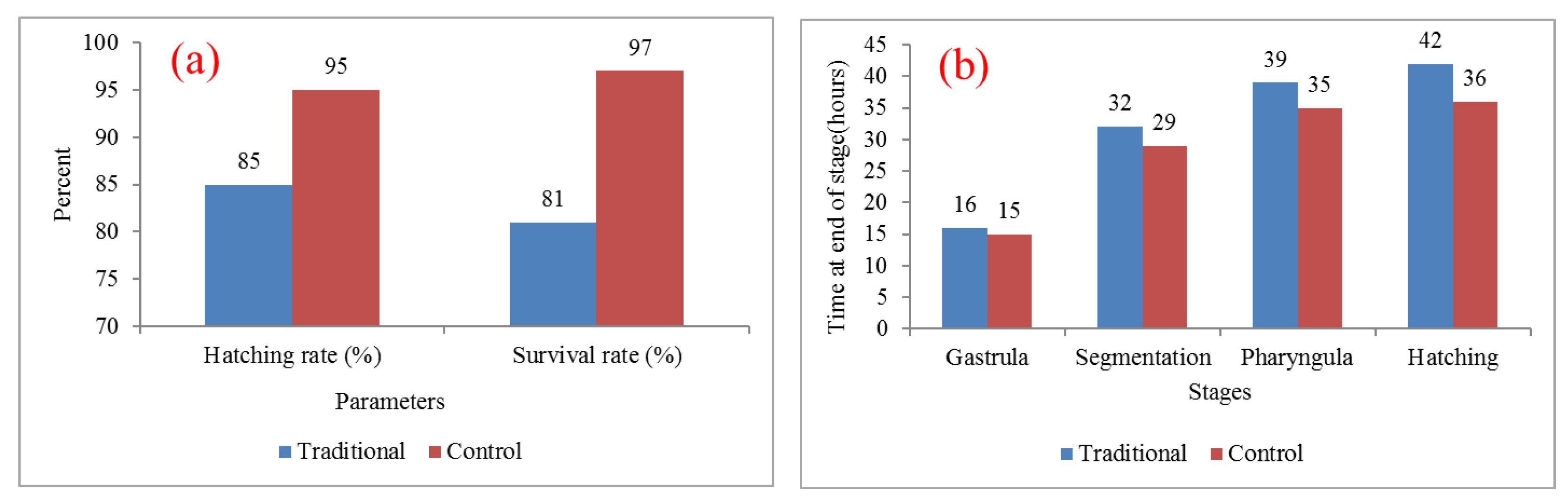
3.2. Hatching and Survival Rate of O. niloticus
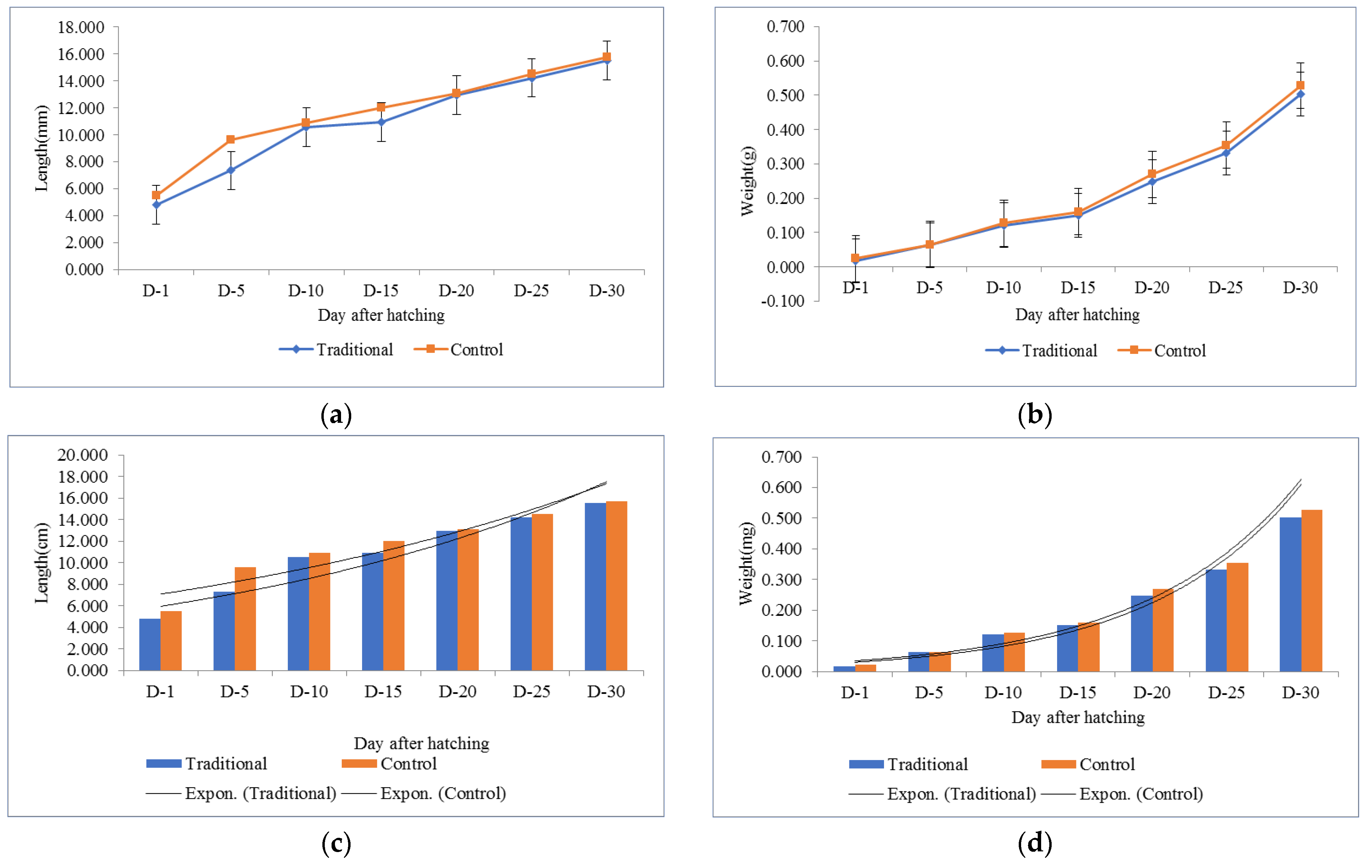
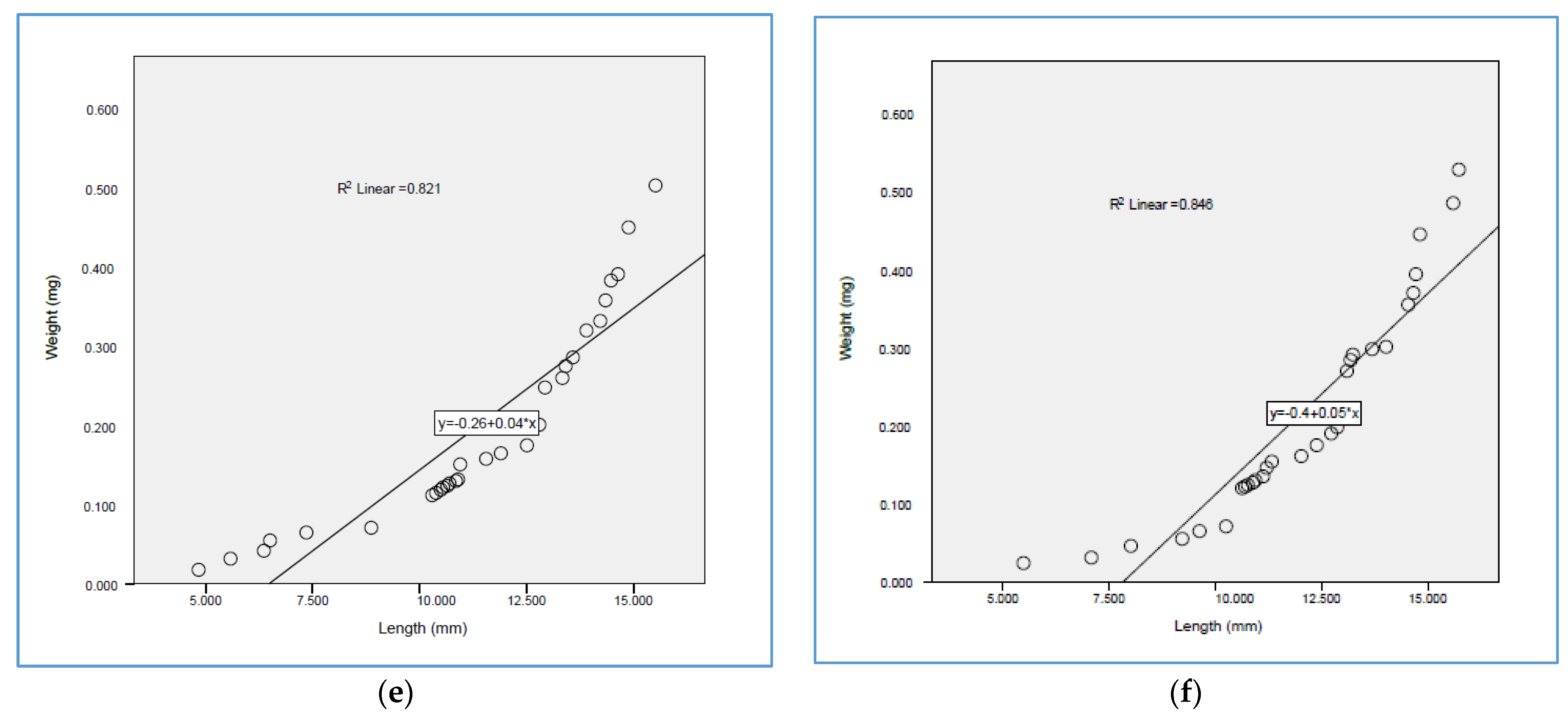
3.3. Larval Development of O. niloticus during Study Period
3.4. Determination of Climatic Variables and Water Quality Parameters in the Traditional Hatchery System and the Re-Circulatory Thermostatic System
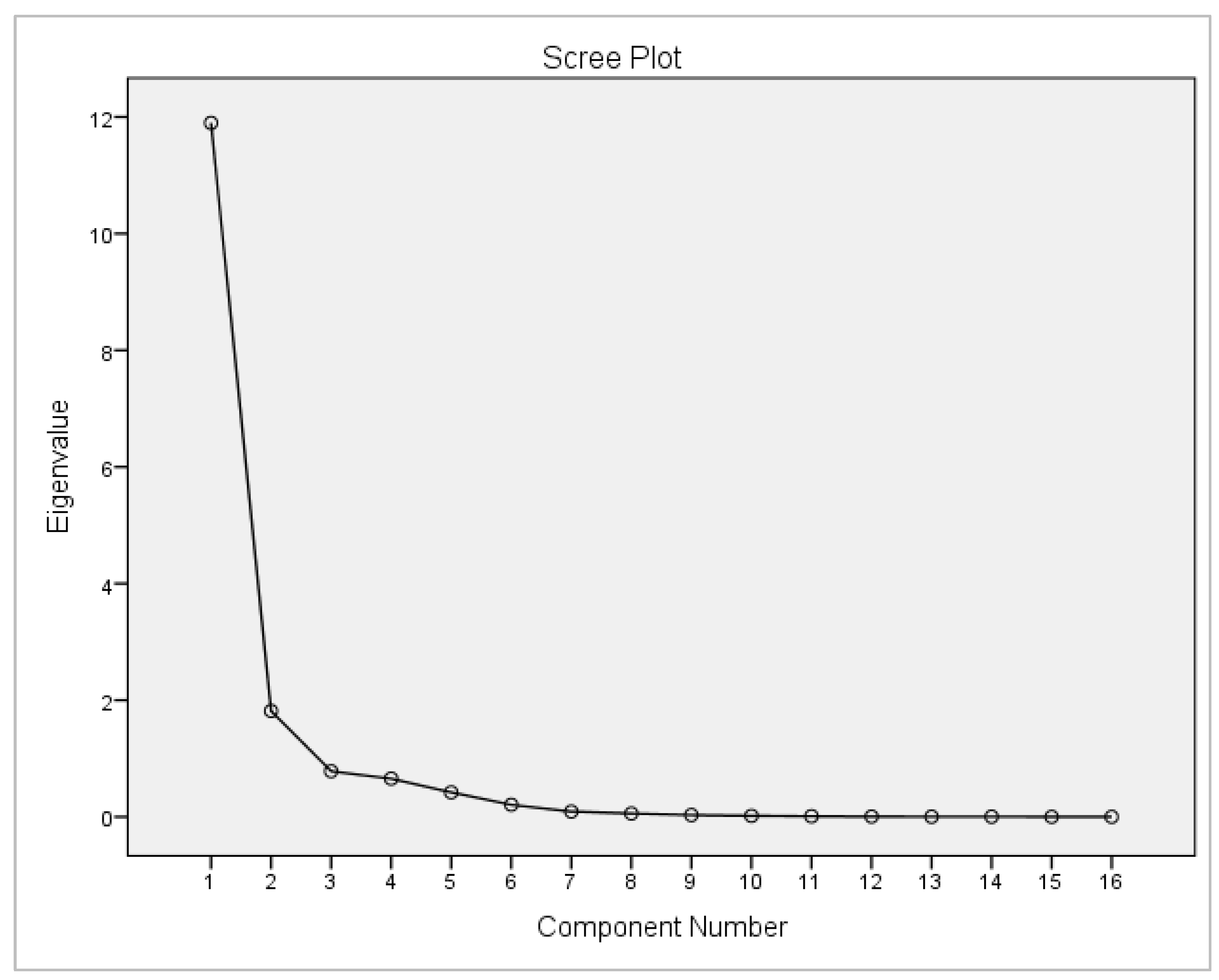
3.5. Analysis of the Relationship of Climatic Variables, Water Quality Parameters, and Morphometric Change of O. niloticus Larvae under the Traditional Hatchery System
4. Discussion
4.1. Observation of Embryonic Development Stages of O. niloticus
4.2. Hatching and Survival Rates of Tilapia, O. niloticus
4.3. Larval Development of O. niloticus during the Study Period
4.4. Determination of Climatic Variables and Water Quality Parameters in the Traditional Hatchery System and the Re-Circulatory Thermostatic System
4.5. The Relationship of Climatic Variables, Water Quality Parameters, and Morphometric Changes of O. niloticus Larvae under the Traditional Hatchery System
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DoF. Yearbook of Fisheries Statistics 2019-20; Fisheries Resources Survey System (FRSS), Department of Fisheries. Bangladesh: Ministry of Fisheries and Livestock: Dhaka, Bangladesh, 2021; Volume 37, p. 141.
- Haque, M.M.; Hasan, N.A.; Eltholth, M.M.; Saha, P.; Mely, S.S.; Rahman, T.; Murray, F.J. Assessing the impacts of in-feed probiotic on the growth performance and health condition of pangasius (Pangasianodon hypophthalmus) in a farm trial. Aquac. Rep. 2021, 20, 100699. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.M.A.; Sarkar, S.I.; Miah, M.A.; Rashid, H. Management strategies for Nile tilapia (Oreochromis niloticus) hatchery in the face of climate change induced rising temperature. Aquac. Stud. 2021, 21, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Hossan, S.M.; Ulka, S.B.; Motin, A.B.; Tarafder, A.K.; Sukhan, Z.P.; Rashid, H. Egg and fry production performance of female tilapia related to fluctuating temperature and size variation. In Proceedings of the 4th the International Conference on Environmental Aspects of Bangladesh, Fukuoka, Japan, 24–26 August 2013; Available online: https://www.researchgate.net/publication/276355629 (accessed on 12 January 2023).
- Siddique, M.A.B.; Ahammad, A.K.S.; Mahalder, B.; Alam, M.M.; Hasan, N.A.; Bashar, A.; Biswas, J.C.; Haque, M.M. Perceptions of the impact of climate change on performance of fish hatcheries in Bangladesh: An empirical study. Fishes 2022, 7, 270. [Google Scholar] [CrossRef]
- Fujimura, K.; Okada, N. Development of the embryo, larva and early juvenile of Nile tilapia Oreochromisniloticus (Pisces: Cichlidae). Developmental staging system. Dev. Growth Differ. 2007, 49, 301–324. [Google Scholar] [CrossRef] [PubMed]
- Haque, M.M. Decentralised Fish Seed Networks in Northwest Bangladesh: Impacts on Rural Livelihoods. Ph.D. Thesis, University of Stirling, Stirling, UK, 2007; p. 493. [Google Scholar]
- Rahman, M.L.; Shahjahan, M.; Ahmed, N. Tilapia farming in Bangladesh: Adaptation to climate change. Sustainability 2021, 13, 7657. [Google Scholar] [CrossRef]
- Gupta, N.; Haque, M.M.; Khan, M. Growth performance of tilapia fingerling in cage in ponds managed by Adivasi households: An assessment through length-weight relationship. J. Bangladesh Agric. Univ. 2012, 10, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Kristiana, V.; Mukti, A.T.; Agustono. Increasing growth performances of Nile tilapia (Oreochromis niloticus) by supplementation of noni morinda citrifolia fruit extract via diet. AACL Bioflux 2020, 13, 159–166. [Google Scholar]
- Faruk, M.A.R.; Mausumi, M.I.; Anka, I.Z.; Hasan, M.M. Effects of temperature on the egg production and growth of monosex Nile tilapia Oreochromis niloticus fry. Bangladesh Res. Pub. J. 2012, 7, 367–377. [Google Scholar]
- Nivelle, R.; Gennotte, V.; Kalala, E.J.K.; Ngoc, N.B.; Muller, M.; Mélard, C.; Rougeot, C. Temperature preference of Nile tilapia (Oreochromis niloticus) juveniles induces spontaneous sex reversal. PLoS ONE 2019, 14, e0212504. [Google Scholar] [CrossRef]
- Valeta, J.S.; Likongwe, J.S.; Kassam, D.; Maluwa, A.O. Temperature-dependent egg development rates, hatchability and fry survival rate of Lake Malawi Tilapia (Chambo). IJFA 2013, 5, 55–59. [Google Scholar]
- Usman, I.; Auta, J.; Abdullahi, S.A. Effect of monthly variation in water temperature on artificial breeding of common carp (Cyprinuscarpio L.) in Zaria, Nigeria. Int. J. Fish. Aquat. Stud. 2015, 3, 353–356. Available online: www.fisheriesjournal.com (accessed on 12 January 2023).
- Sapkale, P.H.; Singh, R.K.; Desai, A.S. Optimal water temperature and ph for development of eggs and growth of spawn of common carp (Cyprinuscarpio). J. Appl. Anim. Res. 2011, 39, 339–345. [Google Scholar] [CrossRef]
- Ahmad, K.; Yuliana, A.R.; Syazili, A.; Surahman. Increasing growth and survival rate of tilapia larvae (Oreochromisniloticus) by adding polychaeta Nereissp. dry meal into feed formulation. Earth Environ. Sci. 2021, 890, 012027. [Google Scholar] [CrossRef]
- Karmakar, S.; Purkait, S.; Das, A.; Samanta, R.; Kumar, K. Climate change and inland fisheries: Impact and mitigation strategies. J. Exp. Zool. 2018, 21, 329–335. [Google Scholar]
- Ahammad, A.K.S.; Asaduzzaman; Ahmed, B.U.; Akter, S.; Islam, S.; Haque, M.M.; Ceylan, H.; Wong, L.L. Muscle cellularity, growth performance and growth-related gene expression of juvenile climbing perch Anabas testudineus in response to different eggs incubation temperature. J. Therm. Biol. 2021, 96, 102830. [Google Scholar] [CrossRef]
- Pankhurst, N.W.; Munday, P.L. Effects of climate change on fish reproduction and early life history stages. Mar. Freshw. Res. 2011, 62, 1015–1026. [Google Scholar] [CrossRef] [Green Version]
- Farid, S.M.; Miah, M.I.; Akter, M.; Saha, D.; Rahman, M.M. Embryonic and larval development of tarabaim (Macrognathusaculeatus). J. Agrofor. Environ. 2008, 2, 123–129. [Google Scholar]
- Aral, F.; Sahnoz, E.; Dogu, Z. Embryonic and larval development of freshwater fish. In Recent Advances in Fish Farms; InTech: Rijeka, Croatia, 2011. [Google Scholar] [CrossRef]
- Politis, S.N.; Mazurais, D.; Servili, A.; Zambonino-Infante, J.L.; Miest, J.J.; Sørensen, S.R.; Tomkiewicz, J.; Butts, I.A.E. Temperature effects on gene expression and morphological development of European eel, Anguilla anguilla larvae. PLoS ONE 2017, 12, e0182726. [Google Scholar] [CrossRef]
- Cuevas-Rodríguez, B.L.; García-Ulloa, M.; Hernández-Lamas, A.; Racotta, I.; Valdez-González, F.J.; Polanco-Torres, A.; Rodríguez-González, H. Evaluating quality of Nile tilapia (Oreochromisniloticus) eggs and juveniles from different commercial hatcheries. Lat. Am. J. Aquat. Res. 2017, 45, 213–217. [Google Scholar] [CrossRef]
- Hasananuzzaman, A.F.M.; Sayeed, M.A.B.; Rahman, S.M.B.; Huq, K.A. Implication of climate change for fisheries and aquaculture in the Sundarban region of Bangladesh. Khulna Univ. Stud. 2010, 73–82. [Google Scholar] [CrossRef]
- Islam, M.M.; Islam, N.; Habib, A.; Mozumder, M.M.H. Climate change impacts on a tropical fishery ecosystem: Implications and societal responses. Sustainability 2020, 12, 7970. [Google Scholar] [CrossRef]
- Mallick, A.; Panigrahi, A.K. Effect of temperature variation on disease proliferation of common fishes in perspective of climate change. Int. J. Exp. Res. Rev. 2018, 16, 40–49. [Google Scholar] [CrossRef]
- Islam, A.; Islam, S.; Wahab, A. Impacts of climate change on shrimph framing in the south-west coastal regions of Bangladesh. Res. Agric. Livest. Fish. 2016, 3, 227–239. [Google Scholar] [CrossRef]
- Dadras, H.; Dzyuba, B.; Cosson, J.; Golpour, A.; Siddique, M.A.M.; Linhart, O. Effect of water temperature on the physiology of fish spermatozoon function: A brief review. Aquac. Res. 2017, 48, 729–740. [Google Scholar] [CrossRef]
- Khater, E.S.G.; Ali, S.A.; Mohamed, W.E. Effect of water temperature on masculinization and growth of Nile tilapia fish. J. Aquac. Res. Dev. 2017, 8, 8–12. [Google Scholar] [CrossRef]
- Shen, Z.G.; Eissa, N.; Yao, H.; Xie, Z.G.; Wang, H.P. Effects of temperature on the expression of two ovarian differentiation-related genes foxl2and cyp19a1a. Front. Physiol. 2018, 9, 1208. [Google Scholar] [CrossRef]
- Islam, M.N.; Amstel, V.A. Bangladesh II: Climate Change Impacts, Mitigation and Adaptation in Developing Countries, 1st ed.; Springer Climate: Berlin/Heidelberg, Germany, 2021. [Google Scholar] [CrossRef]
- Wang, H.L.; Tsai, L.C. Effects of temperature on the deformity and sex differentiation of tilapia, Oreochromis mossambicus. J. Exp. Zool. 2000, 286, 534–537. [Google Scholar] [CrossRef]
- Santo, A.H.E.; de Alba, G.; da Silva Reis, Y.; Costa, L.S.; Sánchez-Vázquez, F.J.; Luz, R.K.; Ribeiro, P.A.P.; López-Olmeda, J.F. Effects of temperature regime on growth and daily rhythms of digestive factors in Nile tilapia (Oreochromis niloticus) larvae. J. Aquac. 2020, 528, 735545. [Google Scholar] [CrossRef]
- Hartati, S.; Harjoko, A.; Supardi, T.W. The digital microscope and its image processing utility. Telkomnika 2011, 9, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Wolf, D.E.; Samarasekera, C.; Swedlow, J.R. Quantitative analysis of digital microscope images. Methods Cell Biol. 2007, 81, 365–396. [Google Scholar] [CrossRef]
- Mojekwu, T.O.; Anumudu, C.I. Advanced techniques for morphometric analysis in fish. J. Aquac. Res. Dev. 2015, 6, 354. [Google Scholar] [CrossRef]
- Statheropoulos, M.; Vassiliadis, N.; Pappa, A. Principal component and canonical correlation analysis for examining air pollution and meteorological data. Atmos. Environ. 1998, 32, 1087–1095. [Google Scholar] [CrossRef]
- Mabe, F.N.; Donkoh, S.A.; Al-Hassan, S. Technology adoption typology and rice yield differentials in Ghana: Principal component analysis approach. Afr. J. Sci. Technol. Innov. Dev. 2019, 11, 555–567. [Google Scholar] [CrossRef]
- Puvaneswari, S.; Marimuthu, K.; Karuppasamy, R.; Haniffa, M.A. Early embryonic and larval development of Indian catfish, Heteropneustesfossilis. Eur. Asian J. BioSci. 2009, 96, 84–96. [Google Scholar] [CrossRef]
- Marimuthu, K.; Haniffa, M.A. Embryonic and larval development of the striped snakehead Channa striatus. Taiwania 2007, 52, 84–92. [Google Scholar]
- Sarma, D.; Das, J.; Dutta, A.; Goswami, U.C. Early embryonic and larval development of Ompokpabo with notes on its nursery rearing. Eur. J. Exp. Biol. 2012, 2, 253–260. [Google Scholar]
- Myint, S.S.; Soe, A.K. Morphological embryonic development stages of Barbonymus gonionotus (Bleeker, 1850). IOP Conf. Ser. Earth Environ. Sci. 2020, 416, 012003. [Google Scholar] [CrossRef]
- Rijnsdorp, A.D.; Peck, M.A.; Engelhard, G.H.; Möllmann, C.; Pinnegar, J.K. Resolving the effect of climate change on fish populations. ICES J. Mar. Sci. 2009, 66, 1570–1583. [Google Scholar] [CrossRef]
- Parvez, S.; Rahman, M.A.; Hasan, J.; Rasel, S.E. Role of hatchary on fish seed production in Patuakhali district of Bangladesh. Int. J. Chem. Environ. Biol. Sci. 2018, 6, 1–7. [Google Scholar]
- Watanabe, W.O.; Lee, C.S.; Ellis, S.C.; Ellis, E.P. Hatchery study of the effects of temperature on eggs and yolksac larvae of the Nassau grouper. Epinephelusstriatus Aquac. 1995, 136, 141–147. [Google Scholar] [CrossRef]
- Okunsebor, S.; Ofojekwu, P.; Kakwi, D.; Audu, B. Effect of temperature on fertilization, hatching and survival rates of Heterobranchusbidorsalis eggs and hatchlings. Br. J. Appl. Sci. Technol. 2015, 7, 372–376. [Google Scholar] [CrossRef]
- McCarthy, I.; Moksness, E.; Pavlov, D.A. The effects of temperature on growth rate and growth efficiency of juvenile common wolffish. Aquac. Int. 1998, 6, 207–218. [Google Scholar] [CrossRef]
- Levit, S.M. A Literature Review of Effects of Ammonia on Fish; The Nature Conservancy: Arlington, VA, USA, 2010; pp. 1–11. [Google Scholar]
- Azaza, M.S.; Dhraief, M.N. Modeling the effects of water temperature on growth rates, gastric evacuation and the return of appetite in juvenile Nile tilapia, Oreochromis niloticus. J. Agric. Sci. 2020, 12, 191. [Google Scholar] [CrossRef]
- Ismarica, I.; Fazriansyah Putra, D.; Khairina, U. Ontogeny and morphological development of Tilapia (Oreochromis niloticus) larvae in Aceh Province, Indonesia. E3S Web Conf. 2022, 339, 01007. [Google Scholar] [CrossRef]
- Sallam, G.A.H.; Elsayed, E.A. Estimating the impact of air temperature and relative humidity change on the water quality of Lake Manzala, Egypt. J. Nat. Resour. Dev. 2015, 5, 76–87. [Google Scholar] [CrossRef] [Green Version]
- Pandit, N.P.; Nakamura, M. Effect of high temperature on survival, growth and feed conversion ratio of Nile tilapia, Oreochromis niloticus. Our Nat. 2010, 8, 219–224. [Google Scholar] [CrossRef] [Green Version]
- Gullian-Klanian, M.; Arámburu-Adame, C. Performance of Nile tilapia Oreochromis niloticus fingerlings in a hyper-intensive recirculating aquaculture system with low water exchange. Lat. Am. J. Aquat. Res. 2013, 41, 150–162. [Google Scholar] [CrossRef]
- Mazumder, S.K.; De, M.; Mazlan, A.G.; Zaidi, C.C.; Rahim, S.M.; Simon, K.D. Impact of global climate change on fish growth, digestion and physiological status: Developing a hypothesis for cause and effect relationships. J. Water Clim. Chang. 2015, 6, 200–226. [Google Scholar] [CrossRef]
- Mahalder, B.; Haque, M.M.; Siddique, M.A.B.; Hasan, N.A.; Alam, M.M.; Talukdar, M.M.N.; Shohan, M.H.; Ahasan, N.; Hasan, M.M.; Ahammad, A.K.S. Embryonic and Larval Development of Stinging Catfish, Heteropneustes fossilis, in Relation to Climatic and Water Quality Parameters. Life 2023, 13, 583. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Traditional Hatchery System | Re-Circulatory Thermostatic System | ||||
|---|---|---|---|---|---|---|
| Highest | Lowest | Average | Highest | Lowest | Average | |
| Air temperature (°C) | 33.87 | 29.63 | 31.52 ± 0.21 | - | - | - |
| Humidity (%) | 69.94 | 45.62 | 58.72 ± 1.47 | - | - | - |
| Water temperature (°C) | 35.50 | 30.50 | 32.71 ± 0.22 | 28.50 | 28.50 | 28.50 |
| pH | 8.60 | 7.50 | 7.99 ± 0.06 | 8.40 | 7.20 | 7.96 ± 0.04 |
| DO (mg/L) | 9.75 | 7.20 | 8.01 ± 0.11 | 9.00 | 7.10 | 8.06 ± 0.09 |
| Ammonia (mg/L) | 0 | 0 | 0 | 0 | 0 | 0 |
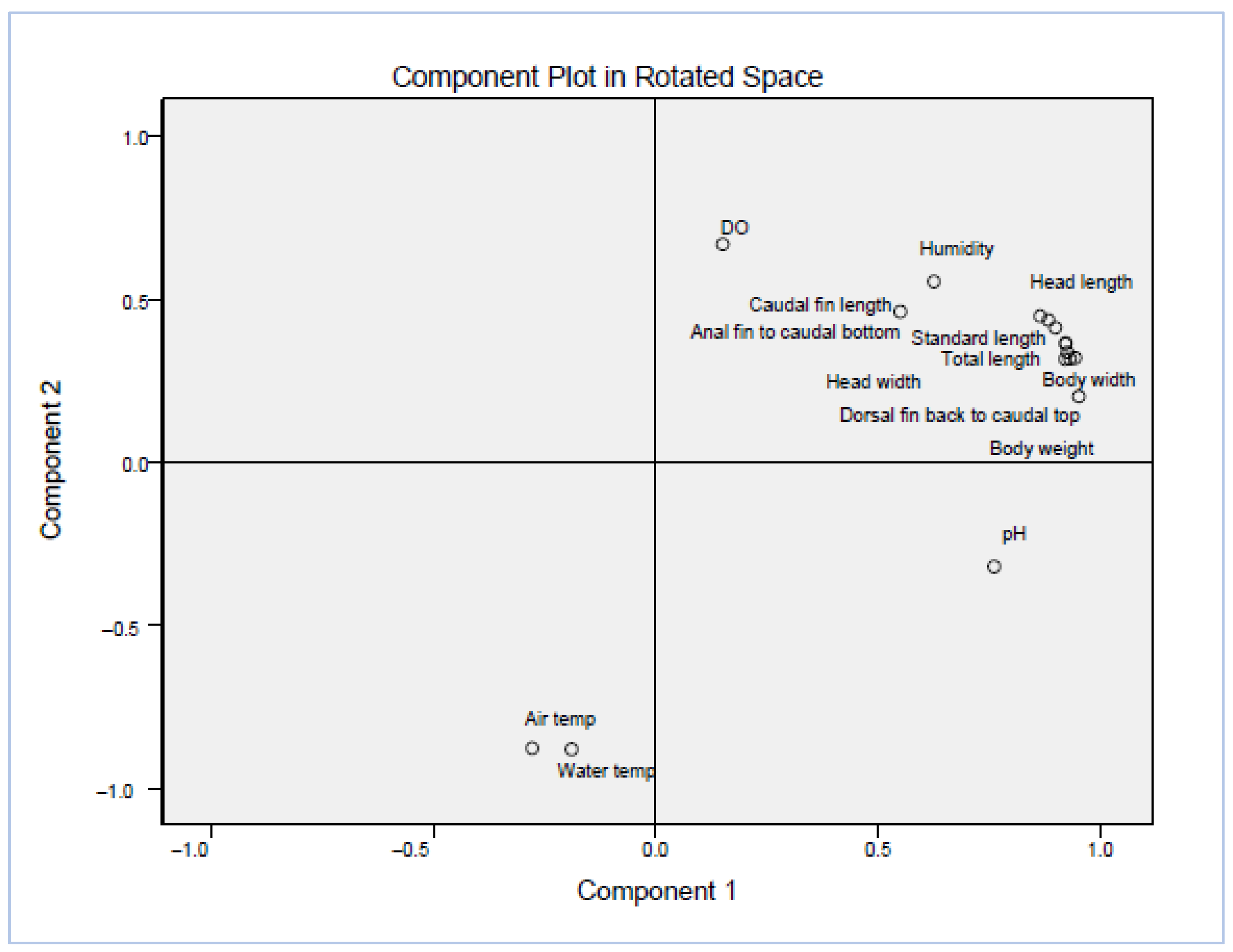
| Rotated Component Matrix a | ||
|---|---|---|
| Component | ||
| 1 | 2 | |
| Total length | 0.921 | 0.317 |
| Body weight | 0.951 | 0.203 |
| Standard length | 0.922 | 0.365 |
| Body width | 0.943 | 0.320 |
| Head length | 0.883 | 0.437 |
| Head width | 0.922 | 0.366 |
| Caudal fin length | 0.550 | 0.462 |
| Caudal fin width | 0.899 | 0.413 |
| Dorsal fin back to caudal top | 0.932 | 0.318 |
| Anal fin to caudal bottom | 0.865 | 0.449 |
| Eye area | 0.926 | 0.338 |
| Air temperature | −0.278 | −0.876 |
| Humidity | 0.626 | 0.555 |
| Water temperature | −0.189 | −0.879 |
| pH | 0.761 | −0.319 |
| DO | 0.151 | 0.669 |
| Principal Component | Initial Eigenvalues | ||
|---|---|---|---|
| Total | % of Variance | Cumulative % | |
| 1 | 11.898 | 74.360 | 74.360 |
| 2 | 1.815 | 11.346 | 85.706 |
| 3 | 0.783 | 4.894 | 90.600 |
| 4 | 0.655 | 4.094 | 94.694 |
| 5 | 0.420 | 2.624 | 97.318 |
| 6 | 0.207 | 1.296 | 98.614 |
| 7 | 0.092 | 0.573 | 99.187 |
| 8 | 0.058 | 0.360 | 99.546 |
| 9 | 0.032 | 0.199 | 99.745 |
| 10 | 0.017 | 0.109 | 99.854 |
| 11 | 0.011 | 0.066 | 99.921 |
| 12 | 0.006 | 0.037 | 99.958 |
| 13 | 0.003 | 0.019 | 99.977 |
| 14 | 0.003 | 0.017 | 99.994 |
| 15 | 0.001 | 0.004 | 99.998 |
| 16 | 0.000 | 0.002 | 100.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siddique, M.A.B.; Mahalder, B.; Haque, M.M.; Bashar, A.; Hasan, M.M.; Shohan, M.H.; Talukdar, M.M.N.; Biswas, J.C.; Ahammad, A.K.S. Assessment of Embryonic and Larval Development of Nile Tilapia under the Traditional and Re-Circulatory Thermostatic System in Relation to Climatic and Water Quality Variations. Aquac. J. 2023, 3, 70-89. https://doi.org/10.3390/aquacj3020008
Siddique MAB, Mahalder B, Haque MM, Bashar A, Hasan MM, Shohan MH, Talukdar MMN, Biswas JC, Ahammad AKS. Assessment of Embryonic and Larval Development of Nile Tilapia under the Traditional and Re-Circulatory Thermostatic System in Relation to Climatic and Water Quality Variations. Aquaculture Journal. 2023; 3(2):70-89. https://doi.org/10.3390/aquacj3020008
Chicago/Turabian StyleSiddique, Mohammad Abu Baker, Balaram Mahalder, Mohammad Mahfujul Haque, Abul Bashar, Md. Mahmudul Hasan, Mobin Hossain Shohan, Md. Mahamudun Naby Talukdar, Jatish Chandra Biswas, and A. K. Shakur Ahammad. 2023. "Assessment of Embryonic and Larval Development of Nile Tilapia under the Traditional and Re-Circulatory Thermostatic System in Relation to Climatic and Water Quality Variations" Aquaculture Journal 3, no. 2: 70-89. https://doi.org/10.3390/aquacj3020008