

Effects of Rearing Density, Substrate Height, and Feeding Frequency on Growth and Biomass Production of Hediste diversicolor

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Experimental Cultivation System Design and Experimental Conditions
2.3. Experimental Design and Statistical Procedures
2.3.1. Trial 1: Rearing Density and Substrate Height
2.3.2. Trial 2: Feeding Frequency
3. Results
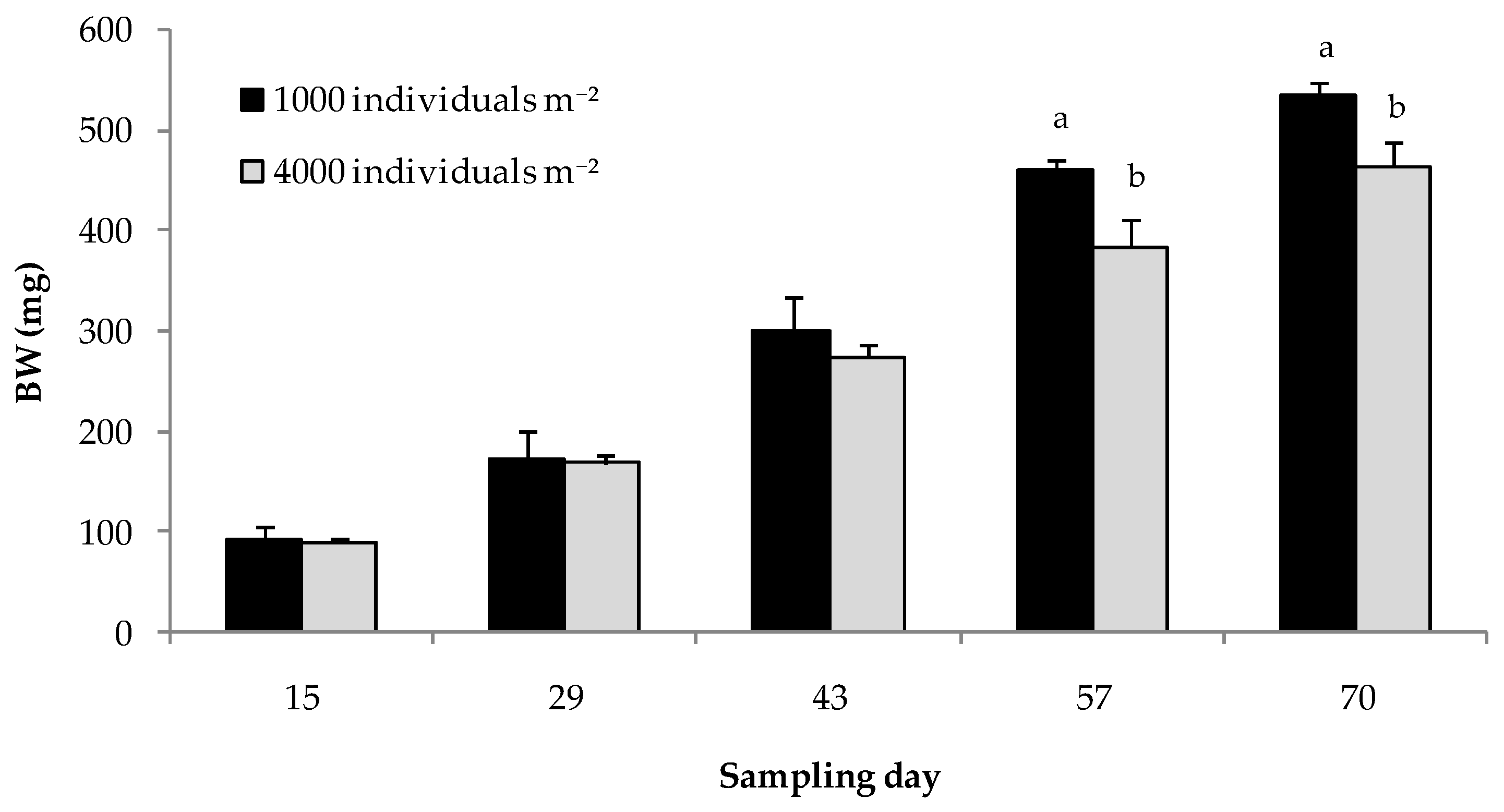
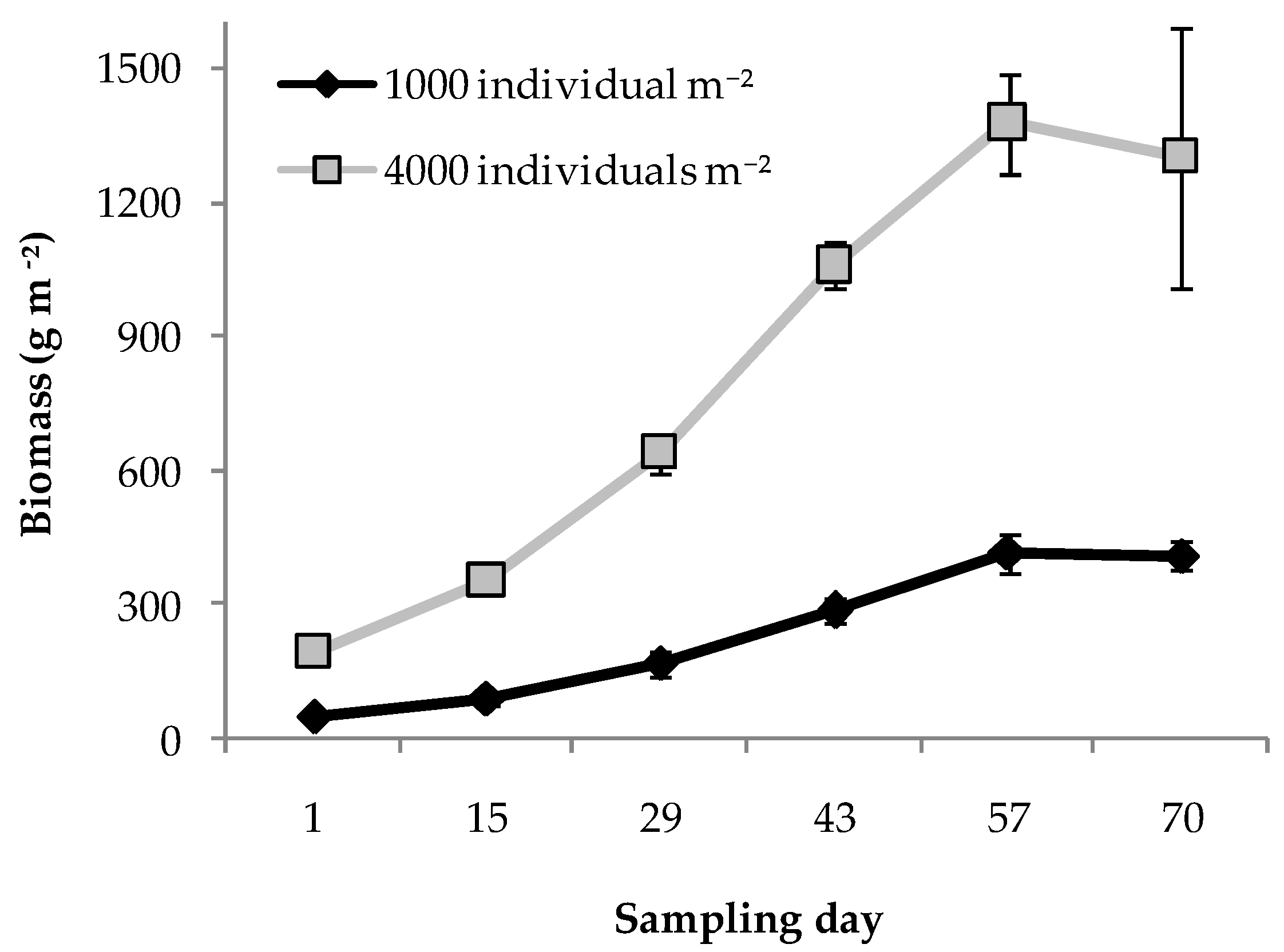
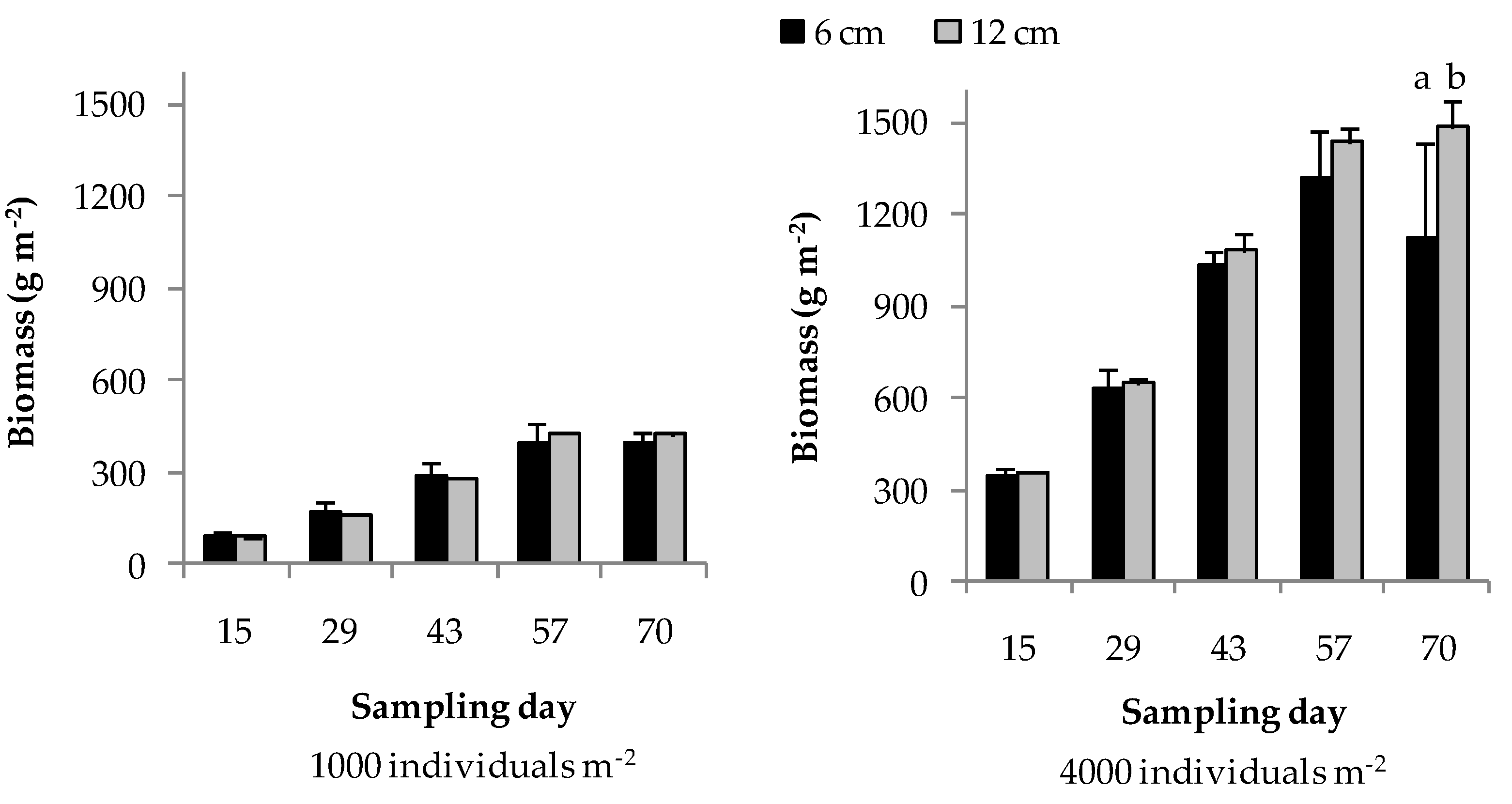
3.1. Trial 1: Rearing Density and Substrate Height
3.2. Trial 2: Feeding Frequency
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pombo, A.; Baptista, T.; Granada, L.; Ferreira, S.M.F.; Gonçalves, S.C.; Anjos, C.; Sá, E.; Chainho, P.; Cancela da Fonseca, L.; Fidalgo e Costa, P.; et al. Insight into aquaculture’s potential of marine annelid worms and ecological concerns: A review. Rev. Aquac. 2018, 12, 107–121. [Google Scholar] [CrossRef]
- Scaps, P. A review of the biology, ecology and potential use of the common ragworm Hediste diversicolor (O. F. Müller) (Annelida: Polychaeta). Hydrobiologia 2002, 470, 203–218. [Google Scholar] [CrossRef]
- Cole, V.J.; Chick, R.C.; Hutchings, P.A. A review of global fisheries for polychaete worms as a resource for recreational fishers: Diversity, sustainability and research needs. Rev. Fish Biol. Fish. 2018, 28, 543–565. [Google Scholar] [CrossRef]
- Batista, F.M.; Fidalgo e Costa, P.; Ramos, A.; Passos, A.M.; Pousao Ferreira, P.; Cancela da Fonseca, L. Production of the ragworm Nereis diversicolor (O. F. Müller, 1776), fed with a diet for gilthead seabream Sparus auratus L., 1758: Survival, growth, feed utilization and oogenesis. Bol. Inst. Esp. Oceanogr. 2003, 19, 447–451. [Google Scholar]
- Pajand, Z.O.; Haddadi Moghaddam, K.; Chubian, F.; Farzaneh, E.; Hosseinnia, E. Effects of dietary various oil sources on zootechnical performance, sexual maturation, whole-body proximate composition and fatty acid profile of Hediste diversicolor. Aquac. Res. 2022, 53, 3078–3089. [Google Scholar] [CrossRef]
- Kabeya, N.; Gür, I.; Oboh, A.; Evjemo, J.O.; Malzahn, A.M.; Hontoria, F.; Navarro, J.C.; Monroig, Ó. Unique fatty acid desaturase capacities uncovered in Hediste diversicolor illustrate the roles of aquatic invertebrates in trophic upgrading. Phil. Trans. R. Soc. B 2020, 375, 20190654. [Google Scholar] [CrossRef]
- Marques, B.; Lillebø, A.I.; Ricardo, F.; Nunes, C.; Coimbra, M.A.; Calado, R. Adding value to ragworms (Hediste diversicolor) through the bioremediation of a super-intensive marine fish farm. Aquac. Environ. Interact. 2018, 10, 79–88. [Google Scholar] [CrossRef]
- Mandario, M.A.E. Addressing gaps in the culture of pathogen–free polychaetes as feed in shrimp hatcheries. Fish People 2018, 16, 19–23. [Google Scholar]
- Nesto, N.; Simonini, R.; Prevedelli, D.; Da Ros, L. Effects of diet and density on growth, survival and gametogenesis of Hediste diversicolor (O. F. Müller, 1776) (Nereididae, Polychaeta). Aquaculture 2012, 362–363, 1–9. [Google Scholar] [CrossRef]
- Garcês, J.P.; Pousao-Ferreira, P. Intraspecific density effect on growth of Marphysa “sp.” Juveniles. Sci. Mar. 2021, 85, 137–144. [Google Scholar] [CrossRef]
- Esselink, P.; Zwarts, L. Seasonal trend in burrow depth and tidal variation in feeding activity of Nereis diversicolor. Mar. Ecol. Prog. Ser. 1989, 56, 243–254. [Google Scholar] [CrossRef]
- Davey, J.T. The architecture of the burrow of Nereis diversicolor and its quantification in relation to sediment-water exchange. J. Exp. Mar. Biol. Ecol. 1994, 179, 115–129. [Google Scholar] [CrossRef]
- Miron, G.; Desrosiers, G.; Retière, C.; Lambert, R. Évolution spatio-temporelle du réseau de galeries chez le polychète Nereis virens (Sars) en relation avec la densité. Can. J. Zool. 1991, 69, 39–42. [Google Scholar] [CrossRef]
- Scaps, P.; Brenot, S.; Retière, C.; Desrosiers, G. Space occupation by the polychaetous annelid Perinereis cultrifera: Influence of substratum heterogeneity and intraspecific interactions on burrow structure. J. Mar. Biol. Assoc. UK 1998, 78, 435–449. [Google Scholar] [CrossRef]
- Miron, G.; Desrosiers, G.; Retière, C.; Lambert, R. Dispersion and prospecting behavior of the polychaete Nereis virens (Sars) as a function of density. J. Exp. Mar. Biol. Ecol. 1991, 145, 65–77. [Google Scholar] [CrossRef]
- Fidalgo e Costa, P. Reproduction and growth in captivity of the polychaete Nereis diversicolor O. F. Müller, 1776, using two different kinds of sediment: Preliminary assays. Bol. Inst. Esp. Oceanogr. 1999, 15, 351–355. [Google Scholar]
- Wang, H.; Hagemann, A.; Reitan, K.I.; Handå, A.; Uhre, M.; Malzahn, A.M. Embryonic and larval development in the semelparous Nereid polychaete Hediste diversicolor (OF Müller, 1776) in Norway: Challenges and perspectives. Aquac. Res. 2020, 51, 4135–4151. [Google Scholar] [CrossRef]
- Fidalgo e Costa, P.; Narciso, L.; Cancela da Fonseca, L. Growth, survival and fatty acid profile of Nereis diversicolor (O. F. Müller, 1776) fed on six different diets. Bull. Mar. Sci. 2000, 67, 337–343. [Google Scholar]
- Santos, A.; Granada, L.; Baptista, T.; Anjos, C.; Simões, T.; Tecelão, C.; Fidalgo e Costa, P.; Costa, J.L.; Pombo, A. Effect of three diets on the growth and fatty acid profile of the common ragworm Hediste diversicolor (O. F. Müller, 1776). Aquaculture 2016, 465, 37–42. [Google Scholar] [CrossRef]
- Yousefi-Garakouei, M.; Kamali, A.; Soltani, M. Effects of rearing density on growth, fatty acid profile and bioremediation ability of polychaete Nereis diversicolor in an integrated aquaculture system with rainbow trout (Oncorhynchus mykiss). Aquac. Res. 2019, 50, 725–735. [Google Scholar] [CrossRef]
- Wang, H.; Seekamp, I.; Malzahn, A.; Hagemann, A.; Carvajal, A.K.; Slizyte, R.; Standal, I.B.; Handå, A.; Reitan, K.I. Growth and nutritional composition of the polychaete Hediste diversicolor (OF Múller, 1776) cultivated on waste from land based salmon smolt. Aquaculture 2019, 502, 232–241. [Google Scholar] [CrossRef]
- Jerónimo, D.; Lillebø, A.I.; Rey, F.; Ii, H.K.; Domingues, M.R.M.; Calado, R. Optimizing the Timeframe to Produce Polychaetes (Hediste diversicolor) Enriched with Essential Fatty Acids Under Different Combinations of Temperature and Salinity. Front. Mar. Sci. 2021, 8, 671545. [Google Scholar] [CrossRef]
- De Silva, S.; Turchini, G.; Francis, D. Nutrition. In Aquaculture: Farming Aquatic Animals and Plants, 2nd ed.; Lucas, J.S., Southgate, P.C., Eds.; Wiley-Blackwell: Oxford, UK, 2012; pp. 164–187. [Google Scholar]
- Xia, B.; Ren, Y.; Wang, J.; Sun, Y.; Zhang, Z. Effects of feeding frequency and density on growth, energy budget and physiological performance of sea cucumber Apostichopus japonicus (Selenka). Aquaculture 2017, 466, 26–32. [Google Scholar] [CrossRef]
- Manley, C.B.; Rakocinski, C.F.; Lee, P.G.; Blaylock, R.B. Feeding frequency mediates aggression and cannibalism in larval hatchery-reared spotted seatrout (Cynoscion nebulosus). Aquaculture 2015, 437, 155–160. [Google Scholar] [CrossRef]
- Satjarak, J.; Thongprajukaew, K.; Kaewtapee, C.; Suanyuk, N.; Klomklao, S.; Preedaphol, K. Optimal feeding frequency for bigfin reef squid (Sepioteuthis lessoniana). Aquac. Res. 2021, 52, 2740–2750. [Google Scholar] [CrossRef]
- Aguado-Giménez, F.; García-García, B.; Martín, I.E.; Rasines, I. Influence of Body Weight And Water Temperature on Growth in Ragworm Hediste diversicolor. Aquac. J. 2023, 3, 19–31. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar]
- Mistiaen, J.A.; Strand, I. Optimal feeding and harvest time for fish with weight-dependent prices. Mar. Resour. Econ. 1999, 13, 231–246. [Google Scholar] [CrossRef]
- Scaps, P.; Retière, C.; Desrosiers, G.; Miron, G. Effects de la ration alimentaire, de la densité intraspécifique et des relations entre individus sur la croissance des juvéniles de l’espèce Nereis diversicolor (Annelida: Polychaeta). Can. J. Zool. 1993, 71, 424–430. [Google Scholar] [CrossRef]
- Safarik, M.; Redden, A.M.; Schreider, M.J. Density-dependent growth of the polychaete Diopatra aciculata. Sci. Mar. 2006, 70, 337–341. [Google Scholar]
- Olive, P.J.W.; Garwood, P.R. Gametogenic cycle and population structure of Nereis (Hediste) diversicolor and Nereis (Nereis) pelagica from North-East England. J. Mar. Biol. Assoc. UK 1981, 61, 193–213. [Google Scholar] [CrossRef]
- Herwati, V.E.; Elfitasari, T.; Rismaningsih, N.; Riyadi, P.H.; Tarangkoon, W.; Radjasa, O.K.; Windarto, S. Analysis of growth and nutritional quality of sea worm (Nereis virens) as a mass cultured natural feed on different substrate media thicknesses. Biodiversitas 2021, 22, 3299–3305. [Google Scholar] [CrossRef]
- Gilbert, F.; Kristensen, E.; Aller, R.C.; Banta, G.T.; Archambault, F.; Belley, R.; Bellucci, L.G.; Calder, L.; Cuny, P.; de Montaudouin, X.; et al. Sediment reworking by the burrowing polychaete Hediste diversicolor modulated by environmental and biological factors across the temperate North Atlantic. A tribute to Gaston Desrosiers. J. Exp. Mar. Biol. Ecol. 2021, 541, 151588. [Google Scholar] [CrossRef]
- Kvaestad, B.; Solvang, T.; Hagemann, A.; Wang, H.; Reitan, K.I.; Handa, A.; Malzahn, A.M. Advances in polychaete cultivation technology—An indoor space efficient cultivation system. In Proceedings of the Aquaculture Europe, Berlin, Germany, 7–8 October 2019. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AGR | SGR | BW | Biomass | Survival | |
|---|---|---|---|---|---|
| Time | *** | *** | *** | *** | *** |
| Time × height | n.s. | n.s. | n.s. | * | n.s. |
| Time × density | n.s. | n.s. | *** | *** | n.s. |
| Time × height × density | n.s. | n.s. | n.s. | n.s. | n.s. |
| Density × height | n.s. | n.s. | n.s. | n.s. | n.s. |
| Density | ** | ** | ** | *** | n.s. |
| Height | n.s | n.s | n.s. | n.s. | n.s. |
| Time Period: | 1–15 | 15–29 | 29–43 | 43–57 | 57–70 | 1–70 | |
|---|---|---|---|---|---|---|---|
| AGR | 1000 | 3.19 ± 0.84 | 5.79 ± 1.15 | 9.04 ± 0.93 | 11.35 ± 3.46 a | 5.46 ± 3.00 | 7.07 ± 0.50 a |
| 4000 | 2.99 ± 0.21 | 5.26 ± 0.47 | 7.90 ± 0.99 | 7.87 ± 1.34 b | 5.63 ± 1.01 | 6.01 ± 0.37 b | |
| SGR | 1000 | 4.65 ± 0.90 | 4.45 ± 0.42 | 3.92 ± 0.42 | 3.03 ± 0.83 | 1.22 ± 0.72 | 3.47 ± 0.08 a |
| 4000 | 4.47 ± 0.27 | 4.27 ± 0.24 | 3.7 ± 0.45 | 2.40 ± 0.30 | 1.45 ± 0.29 | 3.28 ± 0.10 b |
| Substrate Height | Density | SR (%) |
|---|---|---|
| 6 cm | 1000 individuals m−2 | 77 ± 8 |
| 6 cm | 4000 individuals m−2 | 60 ± 14 |
| 12 cm | 1000 individuals m−2 | 80 ± 10 |
| 12 cm | 4000 individuals m−2 | 80 ± 0 |
| BW | SGR | AGR | Survival | CV | |
|---|---|---|---|---|---|
| Size | *** | *** | *** | ** | * |
| Feeding frequency | *** | *** | *** | n.s. | n.s. |
| Feeding frequency × size | *** | * | ** | n.s. | n.s. |
| D | 3T | 1T | ||
|---|---|---|---|---|
| Small (25–50 mg) | SGR | 7.4 ± 0.1 a | 6.4 ± 0.1 b | 5.3 ± 0.1 c |
| AGR | 4.7 ± 0.2 a | 3.9 ± 0.2 b | 2.9 ± 0.0 c | |
| BW | 103.0 ± 5.3 a | 93.3 ± 5.5 a | 77.7 ± 1.2 c | |
| Medium (100–150 mg) | SGR | 5.3 ± 0.4 a | 4.9 ± 0.1 a | 4.3 ± 0.5 a |
| AGR | 9.9 ± 0.8 a | 8.7 ± 0.4 a | 7.3 ± 1.3 a | |
| BW | 266.3 ± 9.5 a | 247.3 ± 10.1 a | 228.7 ± 20.2 a | |
| Large (250–350 mg) | SGR | 3.7 ± 0.5 a | 3.3 ± 0.3 a,b | 2.5 ± 0.5 b |
| AGR | 14.5 ± 2.5 a | 12.1 ± 1.5 a,b | 8.9 ± 1.8 b | |
| BW | 496.0 ± 31.5 a | 459.0 ± 18.5 a,b | 422.3 ± 23.1 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasines, I.; Martín, I.E.; Aguado-Giménez, F. Effects of Rearing Density, Substrate Height, and Feeding Frequency on Growth and Biomass Production of Hediste diversicolor. Aquac. J. 2023, 3, 121-132. https://doi.org/10.3390/aquacj3020011
Rasines I, Martín IE, Aguado-Giménez F. Effects of Rearing Density, Substrate Height, and Feeding Frequency on Growth and Biomass Production of Hediste diversicolor. Aquaculture Journal. 2023; 3(2):121-132. https://doi.org/10.3390/aquacj3020011
Chicago/Turabian StyleRasines, Inmaculada, Ignacio Eduardo Martín, and Felipe Aguado-Giménez. 2023. "Effects of Rearing Density, Substrate Height, and Feeding Frequency on Growth and Biomass Production of Hediste diversicolor" Aquaculture Journal 3, no. 2: 121-132. https://doi.org/10.3390/aquacj3020011