
Electrical Stunning Has the Potential to Delay Fillet Degradation Post-Harvest in Red Seabream (Pagrus major)
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Fish Sampling
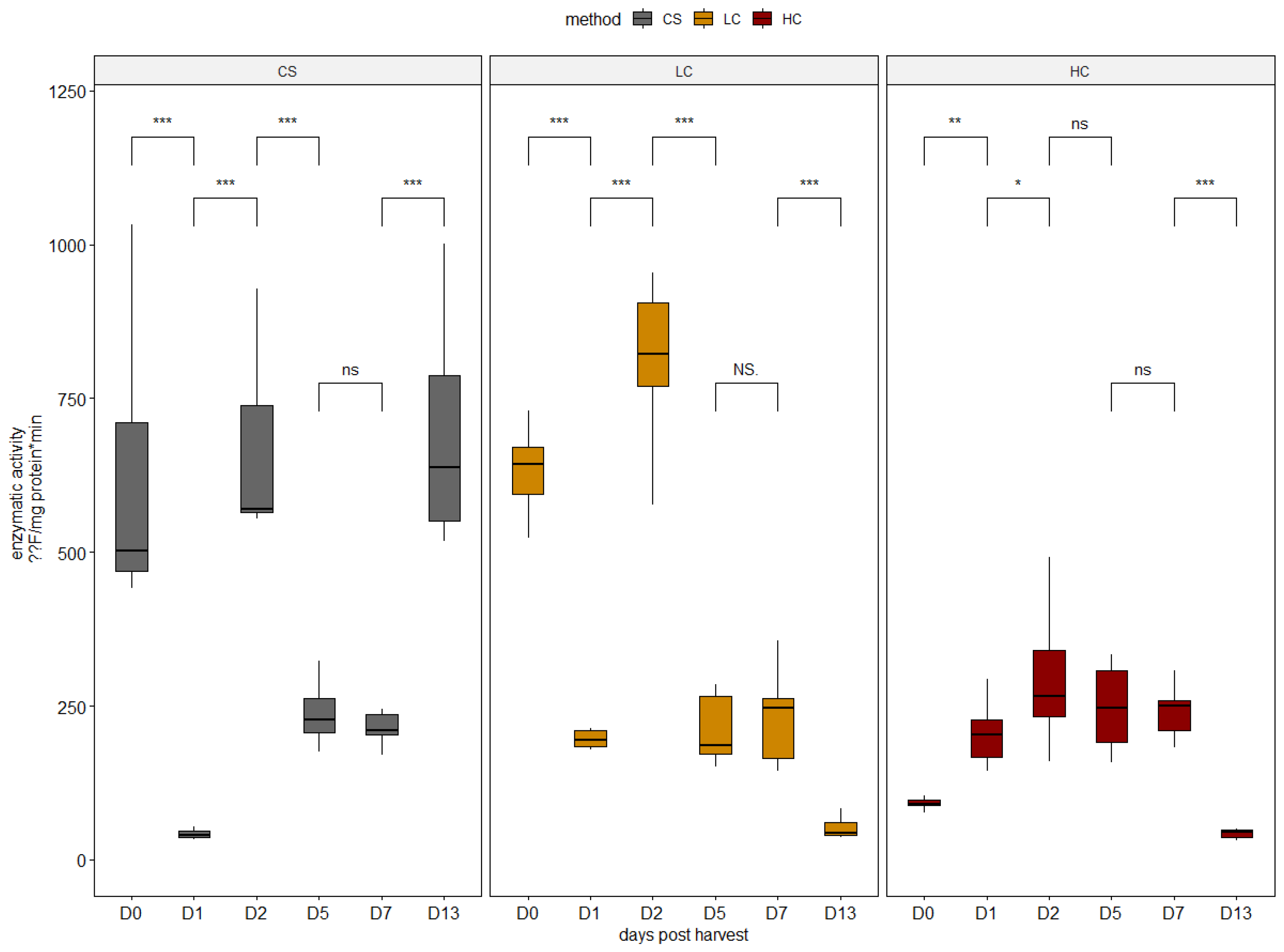
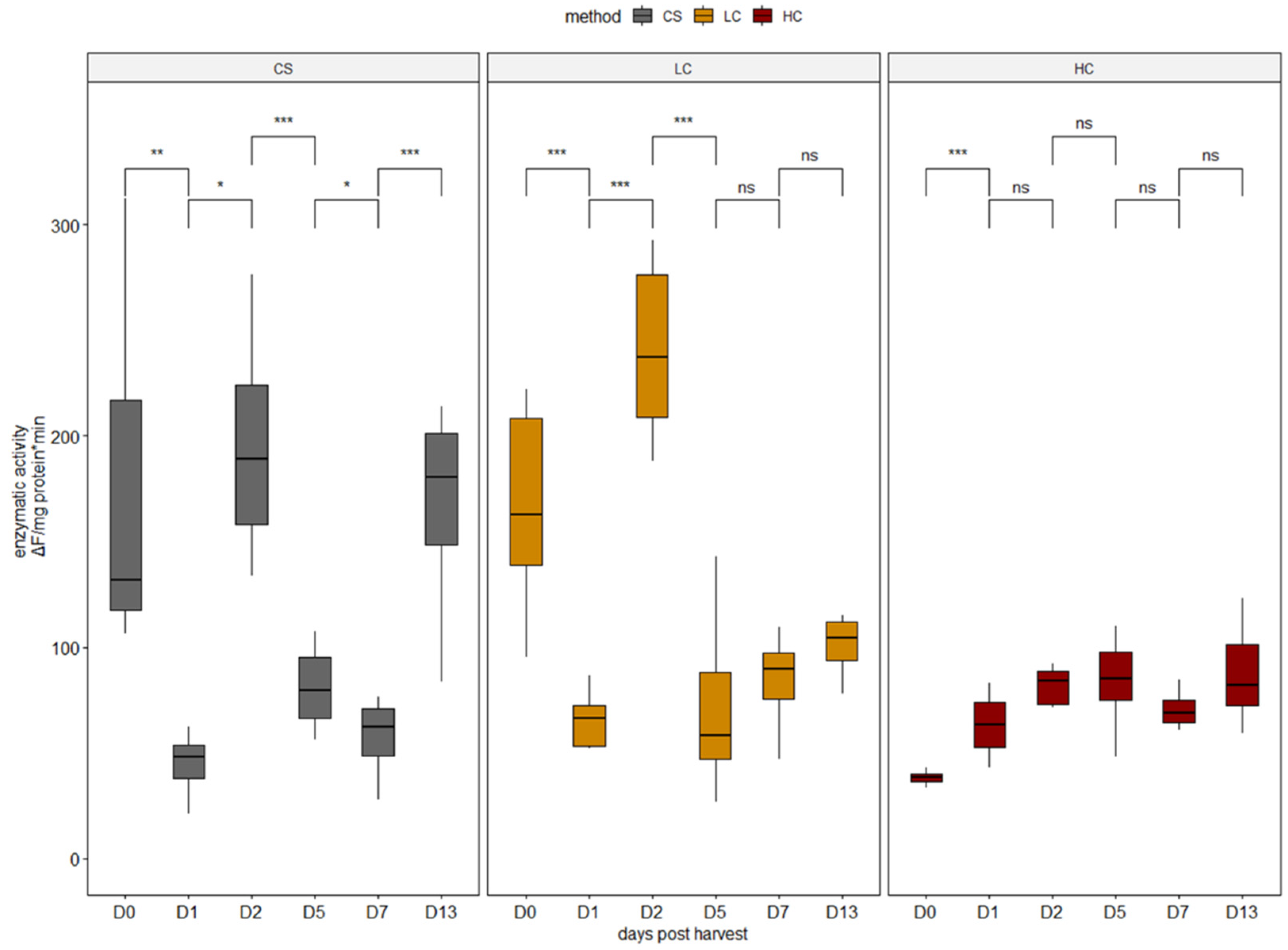
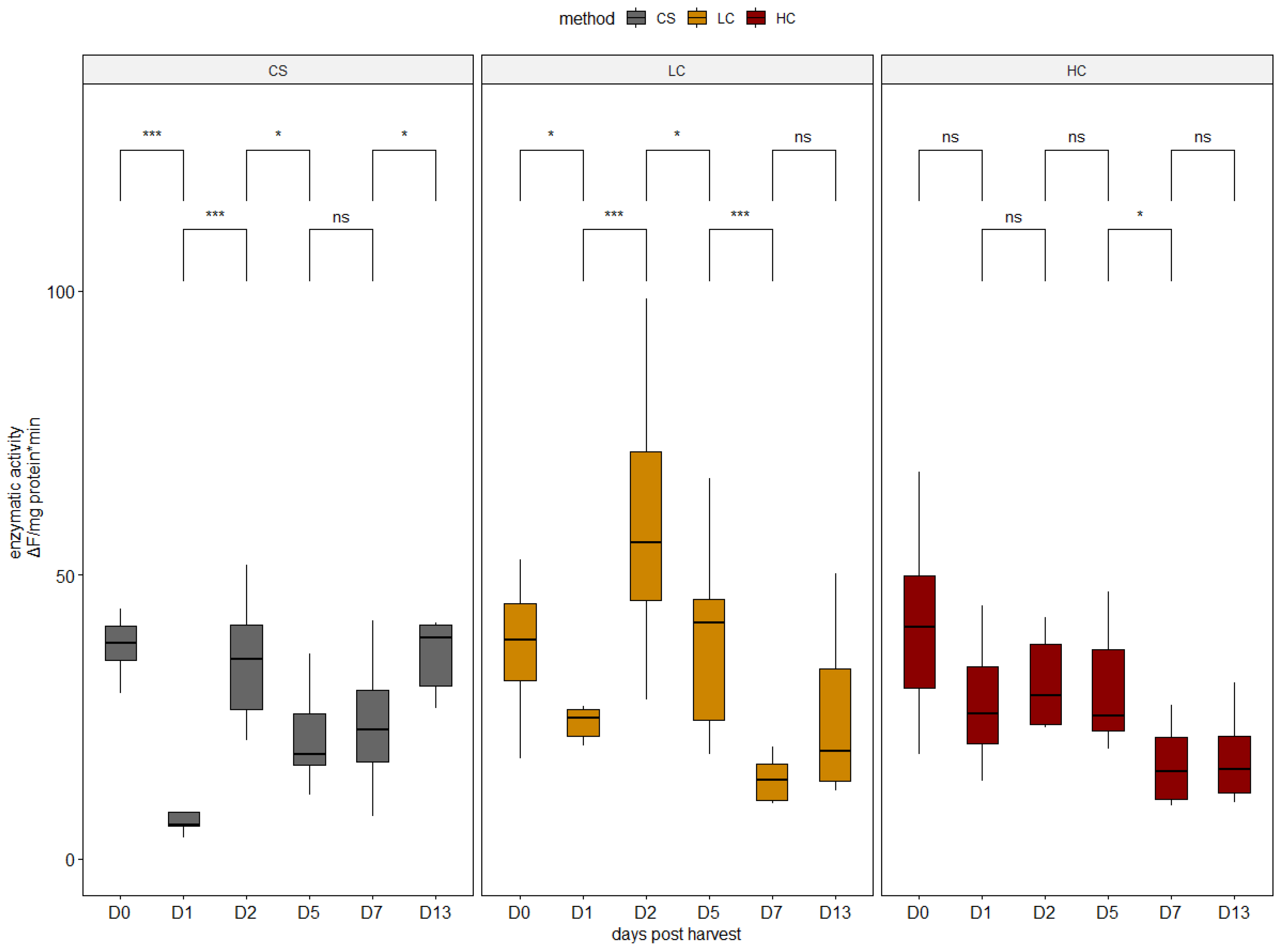
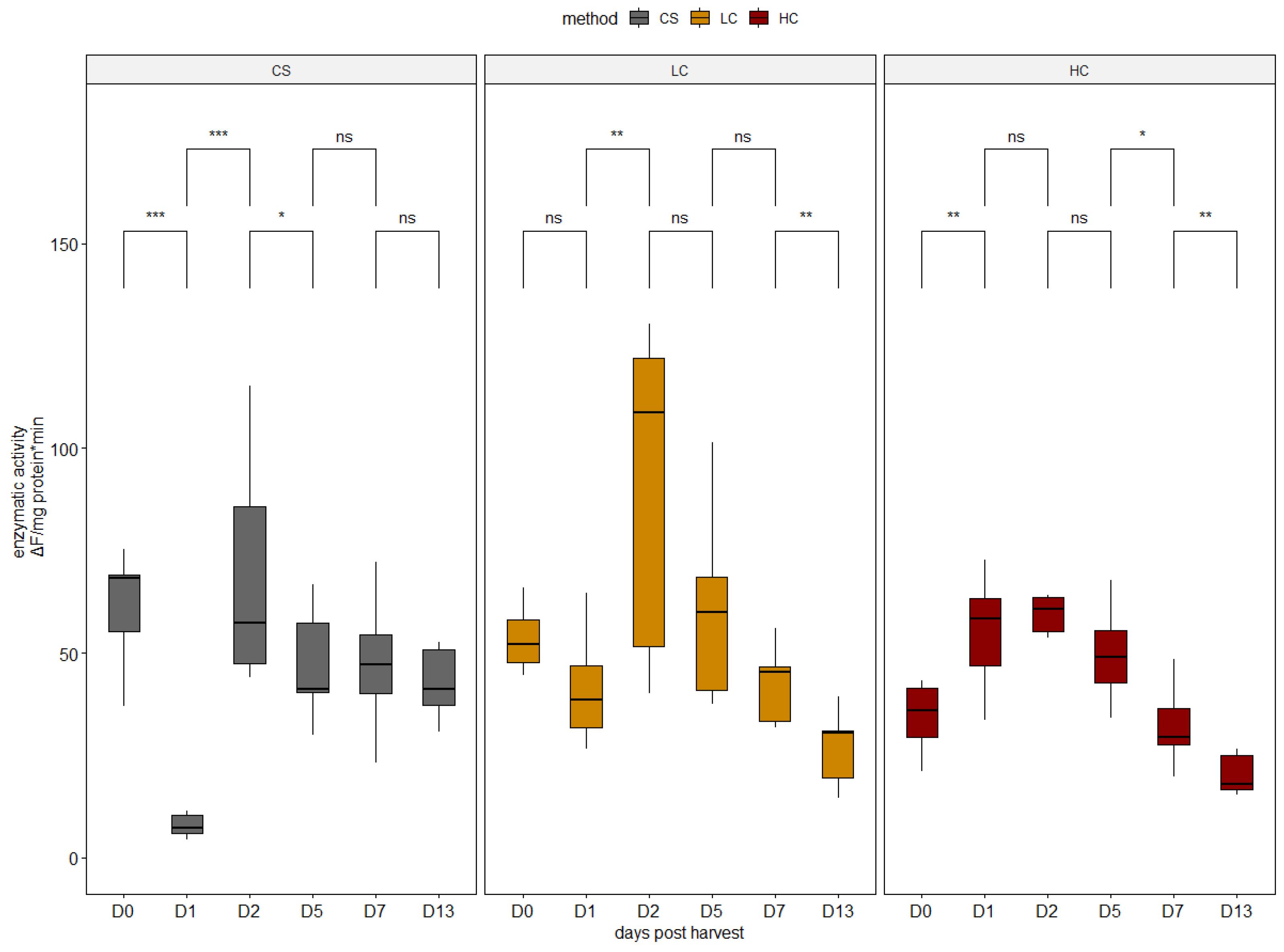
2.3. Proteolytic Enzyme Activities
2.4. Stereological Analysis
2.5. Statistical Analysis
3. Results and Discussion
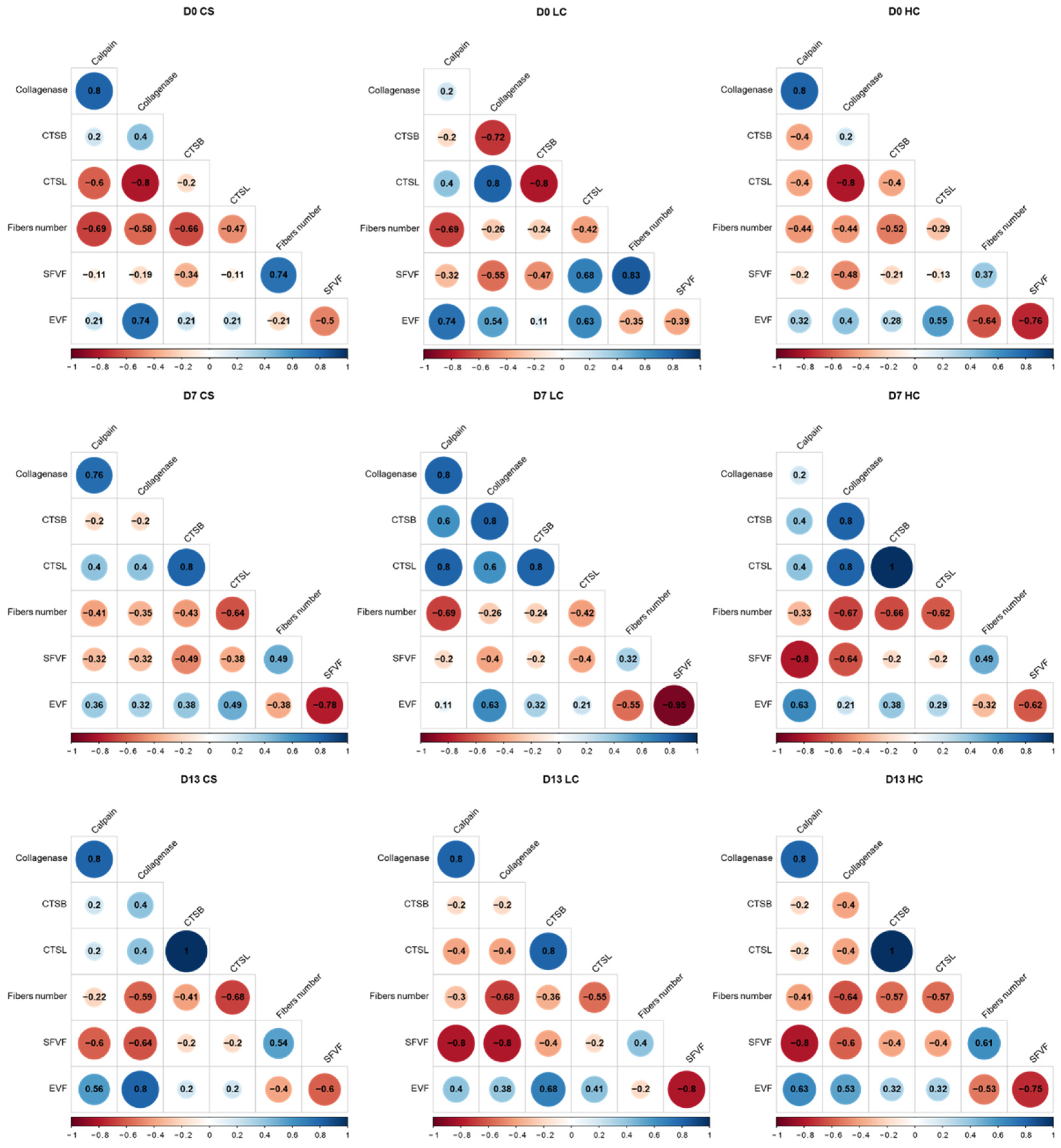
3.1. Enzymatic Activity Analysis
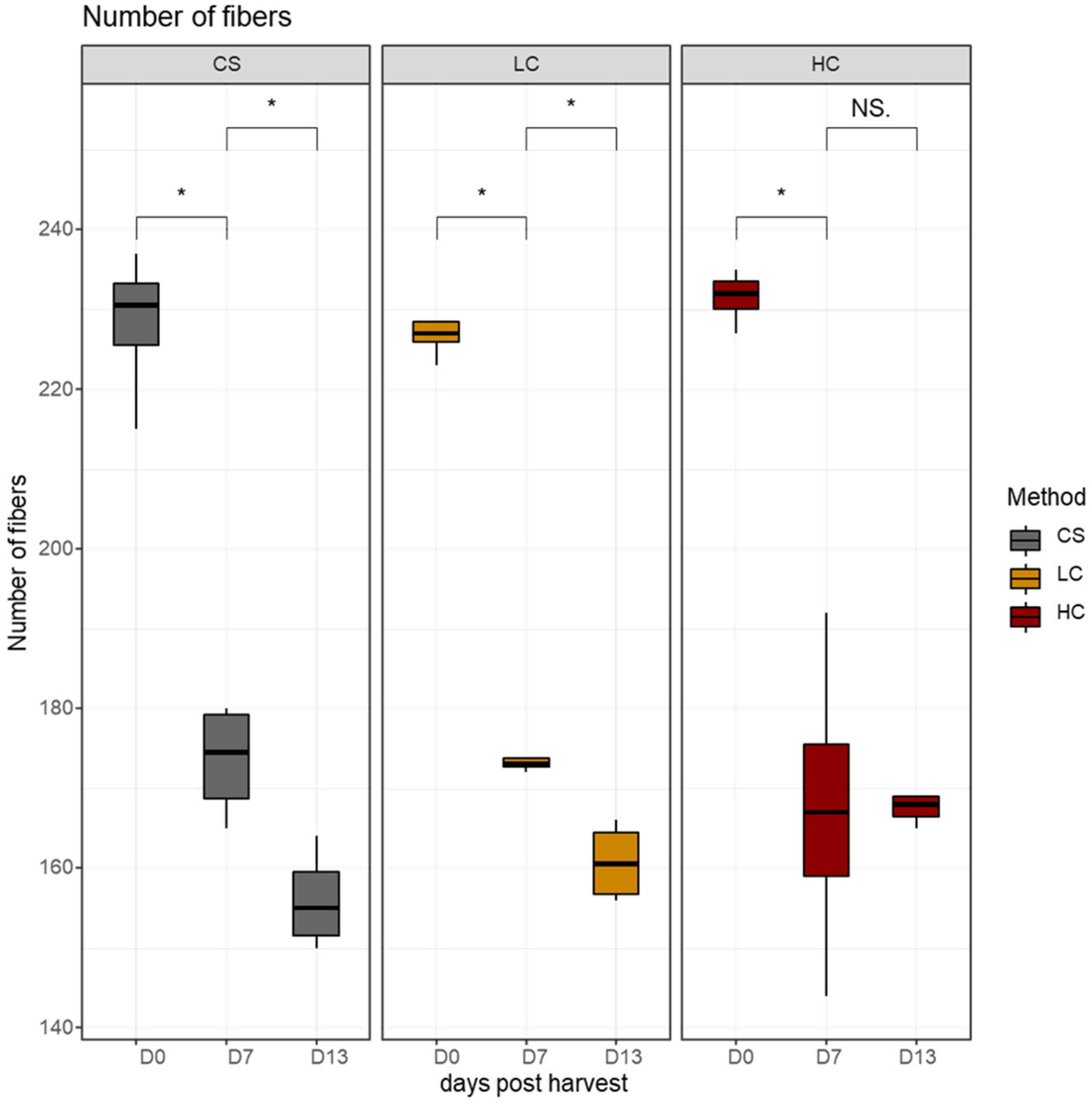
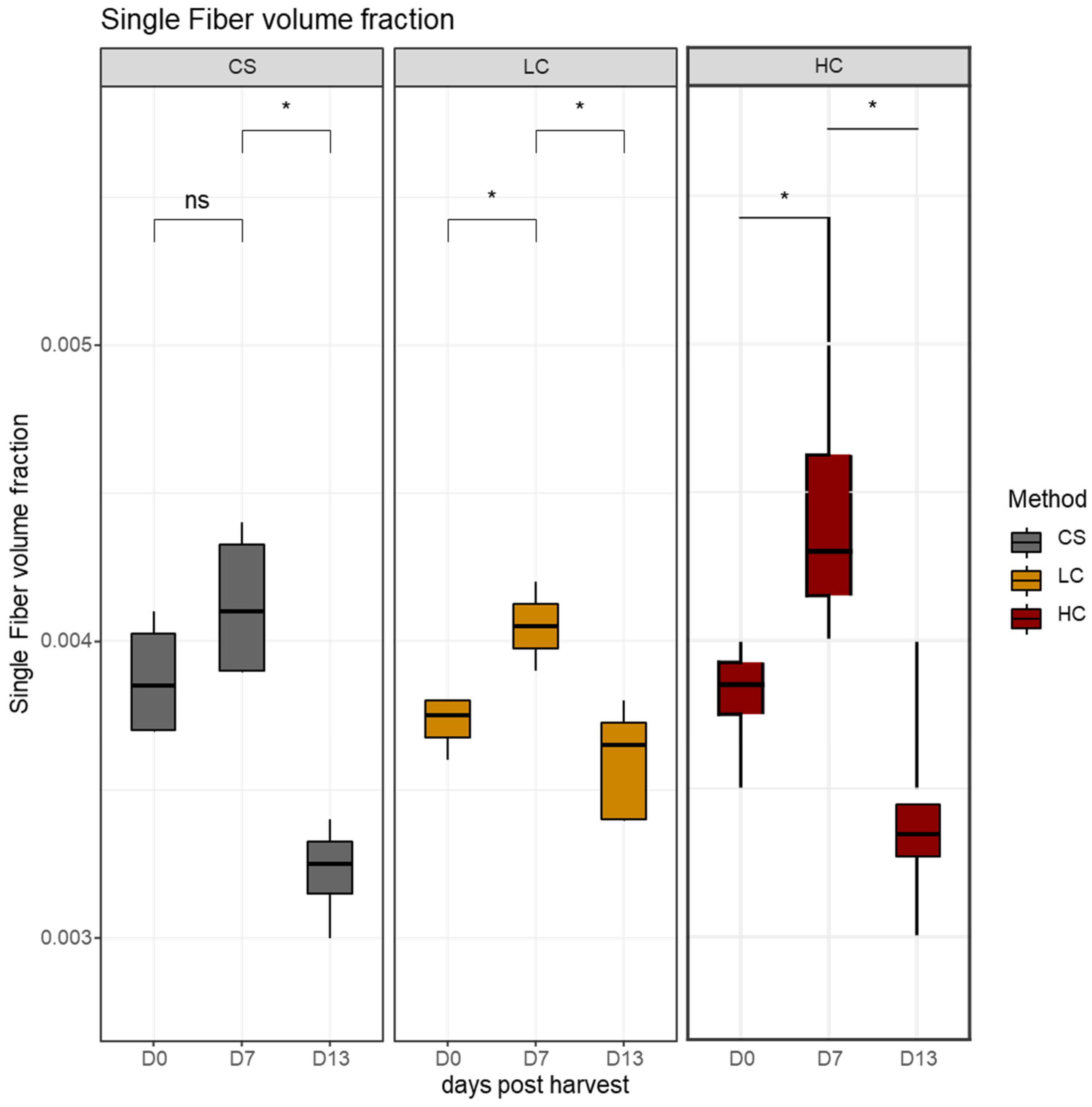
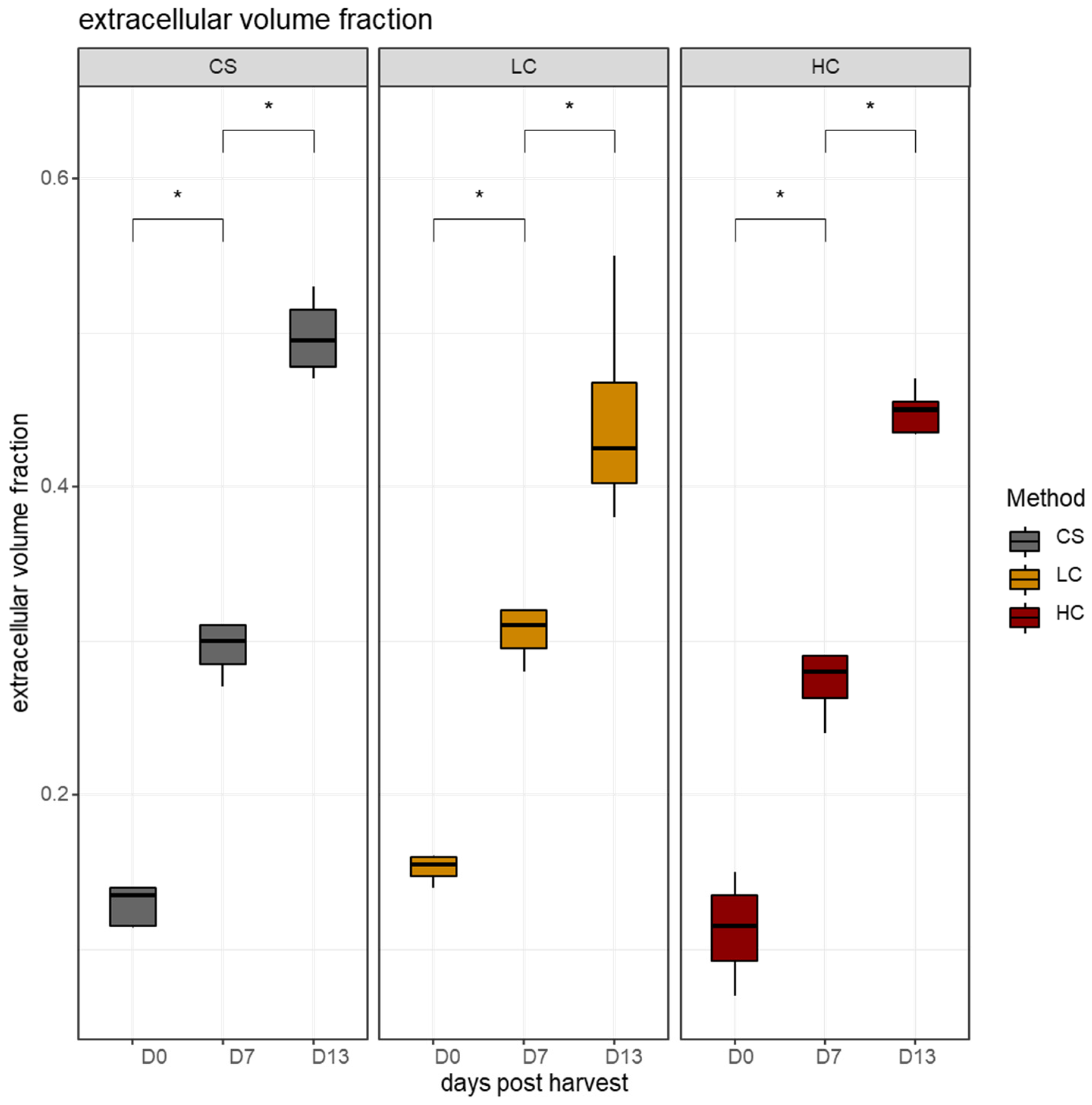
3.2. Stereological Analysis
4. Conclusions
Author Contributions
Funding

Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Tavares, J.; Martins, A.; Fidalgo, L.; Lima, V.; Amaral, R.; Pinto, C.; Silva, A.; Saraiva, J. Fresh Fish Degradation and Advances in Preservation Using Physical Emerging Technologies. Foods 2021, 10, 780. [Google Scholar] [CrossRef] [PubMed]
- LeDuc, A.; Zatylny-Gaudin, C.; Robert, M.; Corre, E.; Le Corguille, G.; Castel, H.; Lefevre-Scelles, A.; Fournier, V.; Gisbert, E.; Andree, K.B.; et al. Dietary aquaculture by-product hydrolysates: Impact on the transcriptomic response of the intestinal mucosa of European seabass (Dicentrarchus labrax) fed low fish meal diets. BMC Genom. 2018, 19, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shouchun, L.; Wen, F.; Saiyi, Z.; Changwei, M.; Pinglan, L.; Kang, Z.; Zhaohui, P.; Meijun, Z. Quality evaluation of tray-packed tilapia fillets stored at 0 °C based on sensory, microbiological, biochemical and physical attributes. Afr. J. Biotechnol. 2010, 9, 692–701. [Google Scholar] [CrossRef] [Green Version]
- Duarte, A.M.; Silva, F.; Pinto, F.R.; Barroso, S.; Gil, M.M. Quality Assessment of Chilled and Frozen Fish—Mini Review. Foods 2020, 9, 1739. [Google Scholar] [CrossRef] [PubMed]
- Delbarre-Ladrat, C.; Verrez-Bagnis, V.; Noël, J.; Fleurence, J. Relative contribution of calpain and cathepsins to protein degradation in muscle of sea bass (Dicentrarchus labrax L.). Food Chem. 2004, 88, 389–395. [Google Scholar] [CrossRef] [Green Version]
- Cheret, R.; Ladrat, C.D.; Lamballerieanton, M.; Verrez-Bagnis, V. Calpain and cathepsin activities in post mortem fish and meat muscles. Food Chem. 2007, 101, 1474–1479. [Google Scholar] [CrossRef] [Green Version]
- Digre, H.; Erikson, U.; Misimi, E.; Lambooij, B.; van de Vis, H. Electrical stunning of farmed Atlantic cod Gadus morhua L.: A comparison of an industrial and experimental method. Aquac. Res. 2009, 41, 1190–1202. [Google Scholar] [CrossRef]
- Fanouraki, E.; Mylonas, C.; Papandroulakis, N.; Pavlidis, M. Species specificity in the magnitude and duration of the acute stress response in Mediterranean marine fish in culture. Gen. Comp. Endocrinol. 2011, 173, 313–322. [Google Scholar] [CrossRef]
- Daskalova, A. Farmed Fish Welfare: Stress, Post-Mortem Muscle Metabolism, and Stress-Related Meat Quality Changes; Springer: Berlin, Germany, 2019; Volume 11, pp. 113–124. [Google Scholar]
- Berg, T.; Erikson, U.; Nordtvedt, T. Rigor Mortis Assessment of Atlantic Salmon (Salmo salar) and Effects of Stress. J. Food Sci. 1997, 62, 439–446. [Google Scholar] [CrossRef]
- Morzel, M.; Van De Vis, H. Effect of the slaughter method on the quality of raw and smoked eels (Anguilla anguilla L.). Aquac. Res. 2003, 34, 1–11. [Google Scholar] [CrossRef]
- Morzel, M.; Sohier, D.; Van De Vis, H. Evaluation of slaughtering methods for turbot with respect to animal welfare and flesh quality. J. Sci. Food Agric. 2003, 83, 19–28. [Google Scholar] [CrossRef]
- Papaharisis, L.; Tsironi, T.; Dimitroglou, A.; Taoukis, P.; Pavlidis, M. Stress assessment, quality indicators and shelf life of three aquaculture important marine fish, in relation to harvest practices, water temperature and slaughter method. Aquac. Res. 2019, 50, 2608–2620. [Google Scholar] [CrossRef]
- Zampacavallo, G.; Parisi, G.; Mecatti, M.; Lupi, P.; Giorgi, G.; Poli, B.M. Evaluation of different methods of stunning/killing sea bass (Dicentrarchus labrax) by tissue stress/quality indicators. J. Food Sci. Technol. 2015, 52, 2585–2597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skjervold, P.O.; Fjæra, S.O.; Østby, P.B.; Einen, O. Live-chilling and crowding stress before slaughter of Atlantic salmon (Salmo salar). Aquaculture 2001, 192, 265–280. [Google Scholar] [CrossRef]
- Rorvik, K.-A.; Skjervold, P.O.; Fjaera, S.O.; Mørkøre, T.; Steien, S.H.; Fjæra, S.O. Body temperature and seawater adaptation in farmed Atlantic salmon and rainbow trout during prolonged chilling. J. Fish Biol. 2001, 59, 330–337. [Google Scholar] [CrossRef]
- Van De Vis, H.; Kestin, S.; Robb, D.; Oehlenschläger, J.; Lambooij, B.; Münkner, W.; Kuhlmann, H.; Kloosterboer, K.; Tejada, M.; Huidobro, A.; et al. Is humane slaughter of fish possible for industry? Aquac. Res. 2003, 34, 211–220. [Google Scholar] [CrossRef]
- Lambooij, B.; Gerritzen, M.A.; Reimert, H.; Burggraaf, D.; André, G.; Van De Vis, H. Evaluation of electrical stunning of sea bass (Dicentrarchus labrax) in seawater and killing by chilling: Welfare aspects, product quality and possibilities for implementation. Aquac. Res. 2008, 39, 50–58. [Google Scholar] [CrossRef]
- Llonch, P.; Lambooij, E.; Reimert, H.; Van De Vis, J. Assessing effectiveness of electrical stunning and chilling in ice water of farmed yellowtail kingfish, common sole and pike-perch. Aquaculture 2012, 364–365, 143–149. [Google Scholar] [CrossRef]
- Algers, B.; Blokhuis, H.J.; Bøtner, A.; Broom, D.M.; Costa, P.; Domingo, M.; Greiner, M.; Hartung, J.; Koenen, F.; Müller-Graf, C.; et al. Species-specific welfare aspects of the main systems of stunning and killing of farmed Atlantic Salmon. EFSA J. 2009, 7, 1011. [Google Scholar] [CrossRef] [Green Version]
- Sattari, A.; Lambooij, E.; Sharifi, H.; Abbink, W.; Reimert, H.; Van De Vis, J. Industrial dry electro-stunning followed by chilling and decapitation as a slaughter method in Claresse® (Heteroclarias sp.) and African catfish (Clarias gariepinus). Aquaculture 2010, 302, 100–105. [Google Scholar] [CrossRef]
- Lines, J.; Kestin, S. Electrical stunning of fish: The relationship between the electric field strength and water conductivity. Aquaculture 2004, 241, 219–234. [Google Scholar] [CrossRef] [Green Version]
- Lines, J.; Spence, J. Humane harvesting and slaughter of farmed fish. OIE Rev. Sci. Tech. 2014, 33, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Federation of Greek Mariculture. Aquaculture in Greece 2020; Annual Report; FGM: Athens, Greece, 2020. [Google Scholar]
- Teixeira, B.; Fidalgo, L.; Mendes, R.; da Costa, G.; Cordeiro, C.; Marques, A.; Saraiva, J.; Nunes, M.L. Changes of Enzymes Activity and Protein Profiles Caused by High-Pressure Processing in Sea Bass (Dicentrarchus labrax) Fillets. J. Agric. Food Chem. 2013, 61, 2851–2860. [Google Scholar] [CrossRef] [PubMed]
- Barrett, A.J.; Kirschke, H. Cathepsin B, cathepsin H, and cathepsin L. Methods Enzymol. 1981, 80, 535–561. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ntzimani, A.; Angelakopoulos, R.; Semenoglou, I.; Dermesonlouoglou, E.; Tsironi, T.; Moutou, K.; Taoukis, P. Slurry ice as an alternative cooling medium for fish harvesting and transportation: Study of the effect on seabass flesh quality and shelf life. Aquac. Fish. 2021, in press. [Google Scholar] [CrossRef]
- Georgiou, S.; Alami-Durante, H.; Power, D.M.; Sarropoulou, E.; Mamuris, Z.; Moutou, K.A. Transient up- and down-regulation of expression of myosin light chain 2 and myostatin mRNA mark the changes from stratified hyperplasia to muscle fiber hypertrophy in larvae of gilthead sea bream (Sparus aurata L.). Cell Tissue Res. 2016, 363, 541–554. [Google Scholar] [CrossRef]
- Weibel, E.R.; Staubli, W.; Gnagi, H.R.; Hess, F.A. Correlated Morphometric and Biochemical Studies on the Liver Cell: I. Morphometric Model, Stereologic Methods, and Normal Morphometric Data for Rat Liver. J. Cell Biol. 1969, 42, 68–91. [Google Scholar] [CrossRef]
- Weibel, E.R. Morphometry of the human lung: The state of the art after two decades. Bull. Eur. Physiopathol. Respir. 1979, 15, 999–1013. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- RStudio Team. RStudio: Integrated Development Environment for R 2020; RStudio, PBC: Boston, MA, USA, 2020. [Google Scholar]
- Caballero, M.; Betancor, M.; Escrig, J.; Montero, D.; Monteros, A.E.D.L.; Castro, P.; Ginés, R.; Izquierdo, M. Post mortem changes produced in the muscle of sea bream (Sparus aurata) during ice storage. Aquaculture 2009, 291, 210–216. [Google Scholar] [CrossRef]
- Mendes, R. Technological processing of fresh gilthead seabream (Sparus aurata): A review of quality changes. Food Rev. Int. 2018, 35, 20–53. [Google Scholar] [CrossRef]
- Irianto, H.E. Enzymes in Fermented Fish. Adv. Food Nutr. Res. 2017, 80, 199–216. [Google Scholar] [CrossRef]
- Bao, Y.; Wang, K.; Yang, H.; Regenstein, J.M.; Ertbjerg, P.; Zhou, P. Protein degradation of black carp (Mylopharyngodon piceus) muscle during cold storage. Food Chem. 2020, 308, 125576. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Jiang, X.; Ge, L.; Zang, J.; Xia, W.; Jiang, Q. Inhibitory Effect of Edible Additives on Collagenase Activity and Softening of Chilled Grass Carp Fillets. J. Food Process. Preserv. 2017, 41, e12836. [Google Scholar] [CrossRef]
- Strateva, M.; Penchev, G. Histological, Physicochemical and Microbiological Changes in Fresh and Frozen/Thawed Fish. Trakia J. Sci. 2020, 18, 69–80. [Google Scholar] [CrossRef]
- Hu, R.; Zhang, M.; Liu, W.; Mujumdar, A.S.; Bai, B. Novel synergistic freezing methods and technologies for enhanced food product quality: A critical review. Compr. Rev. Food Sci. Food Saf. 2022, 21, 1979–2001. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, Y.; Jia, S.; Li, Y.; Li, Q.; Li, K.; Hong, H.; Luo, Y. Stunning stress-induced textural softening in silver carp (Hypophthalmichthys molitrix) fillets and underlying mechanisms. Food Chem. 2019, 295, 520–529. [Google Scholar] [CrossRef]
- Singh, A.; Benjakul, S. Proteolysis and Its Control Using Protease Inhibitors in Fish and Fish Products: A Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 496–509. [Google Scholar] [CrossRef] [Green Version]
- Ayala, M.D.; Abdel, I.; Santaella, M.; Martínez, C.; Periago, M.J.; Gil, F.; Blanco, A.; Albors, O.L. Muscle tissue structural changes and texture development in sea bream, Sparus aurata L., during post-mortem storage. LWT Food Sci. Technol. 2010, 43, 465–475. [Google Scholar] [CrossRef]
- Ge, L.; Xu, Y.; Xia, W.; Jiang, Q. Synergistic action of cathepsin B, L, D and calpain in disassembly and degradation of myofibrillar protein of grass carp. Food Res. Int. 2018, 109, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, T.; Shigehisa, T.; Taji, S.; Hayashi, R. Biochemical Effects of High Hydrostatic Pressure on the Lysosome and Proteases Involved in It. Biosci. Biotechnol. Biochem. 1992, 56, 1285–1288. [Google Scholar] [CrossRef] [Green Version]
- Sentandreu, M.; Coulis, G.; Ouali, A. Role of muscle endopeptidases and their inhibitors in meat tenderness. Trends Food Sci. Technol. 2002, 13, 400–421. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | LC | HC |
|---|---|---|
| Water Conductivity | 50 ms/cm | 50 ms/cm |
| Waveform | AC (sinusoidal voltage) | AC (sinusoidal voltage) |
| Amperage | 27 A | 48 A |
| Electric field | 1.5 V/cm | 1.8 V/cm |
| Frequency | 1000 Hz | 1000 Hz |
| Velocity | 1.6 m/s | 1.6 m/s |
| Exposure | 9 s | 9 s |
| Water pipe length | 14 m | 14 m |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Angelakopoulos, R.; Dimitroglou, A.; Papaharisis, L.; Moutou, K.A. Electrical Stunning Has the Potential to Delay Fillet Degradation Post-Harvest in Red Seabream (Pagrus major). Aquac. J. 2022, 2, 302-315. https://doi.org/10.3390/aquacj2040018
Angelakopoulos R, Dimitroglou A, Papaharisis L, Moutou KA. Electrical Stunning Has the Potential to Delay Fillet Degradation Post-Harvest in Red Seabream (Pagrus major). Aquaculture Journal. 2022; 2(4):302-315. https://doi.org/10.3390/aquacj2040018
Chicago/Turabian StyleAngelakopoulos, Rafael, Arkadios Dimitroglou, Leonidas Papaharisis, and Katerina A. Moutou. 2022. "Electrical Stunning Has the Potential to Delay Fillet Degradation Post-Harvest in Red Seabream (Pagrus major)" Aquaculture Journal 2, no. 4: 302-315. https://doi.org/10.3390/aquacj2040018