
The Effects of a Multi-Ingredient Supplement Containing Wasabia Japonica Extract, Theacrine, and Copper (I) Niacin Chelate on Peripheral Blood Mononuclear Cell DNA Methylation, Transcriptomics, and Sirtuin Activity

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Participant Characteristics from Our Prior Data Analysis
2.2. PBMC mRNA Expression Differences between Treatment Groups
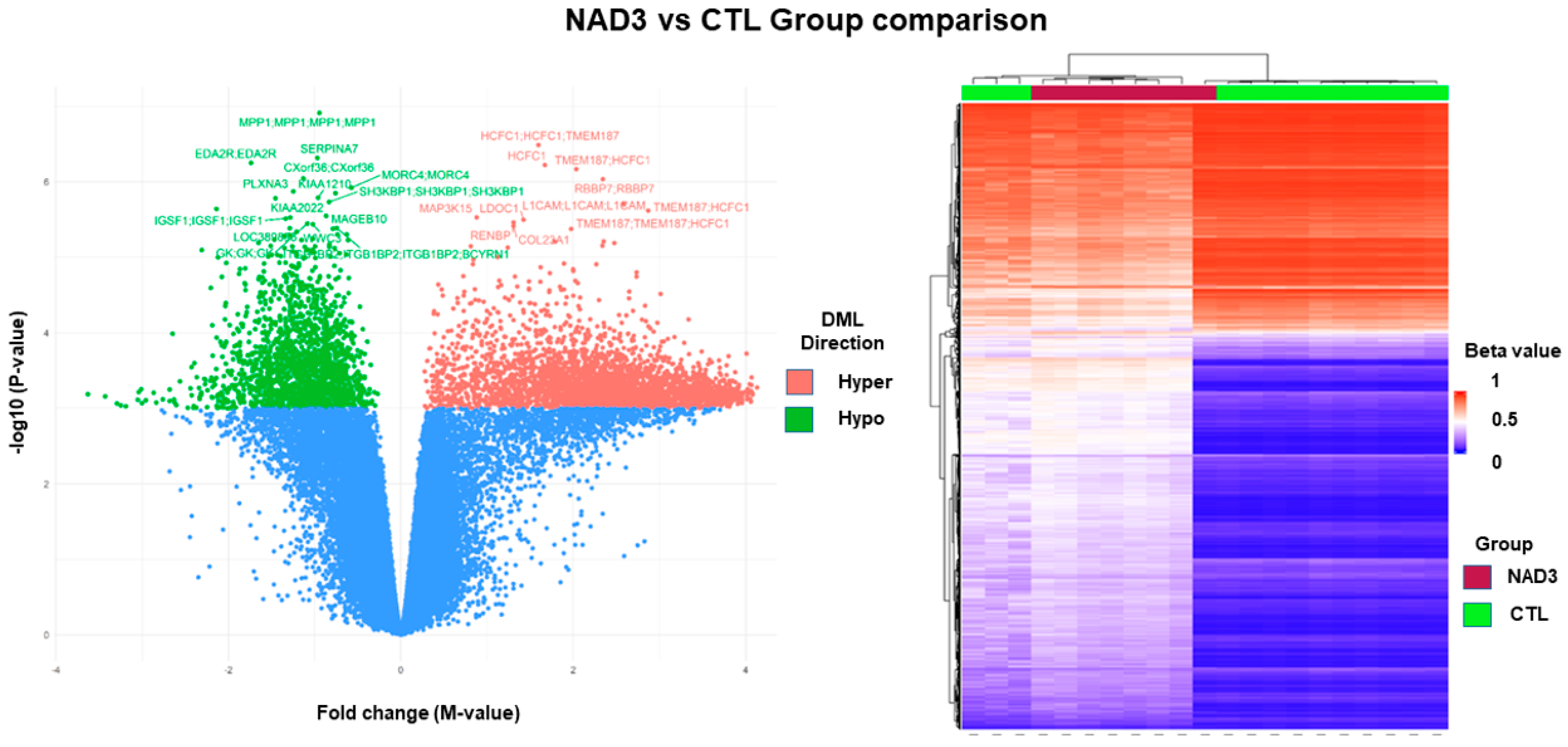
2.3. PBMC Methylome Differences between Groups
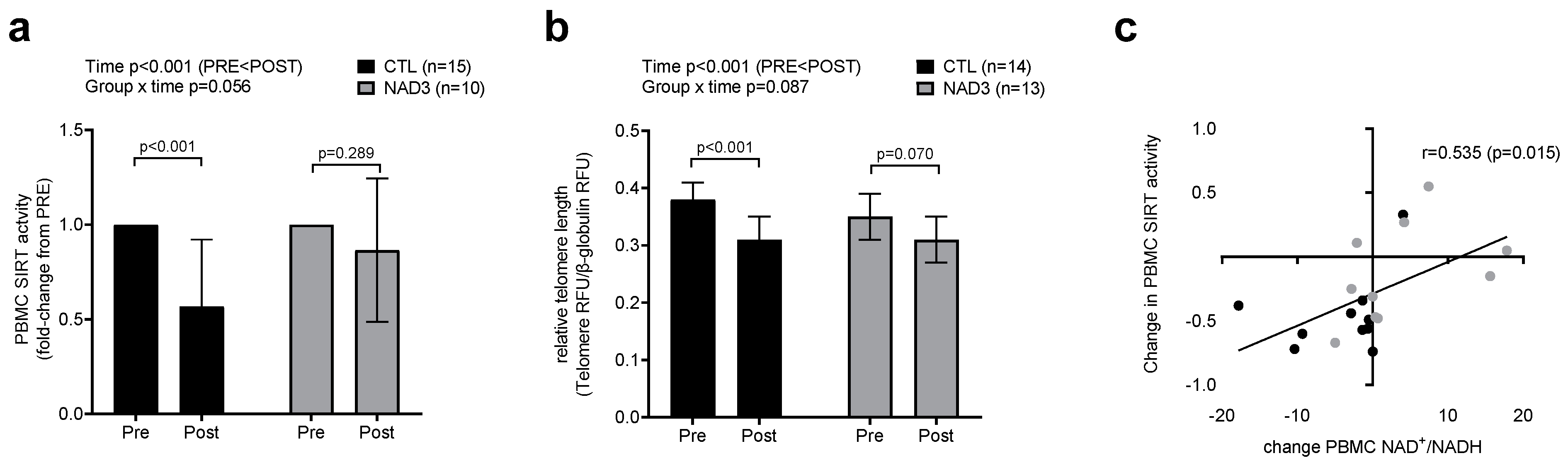
2.4. PBMC Telomere Length and SIRT Activity Differences between Treatment Groups
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Testing Sessions and Supplementation
4.3. PBMC Transcriptome Analyses
4.4. PBMC DNA Methylation Analysis
4.5. PBMC SIRT and Telomere Length Assays
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gualano, B.; Rawson, E.S.; Candow, D.G.; Chilibeck, P.D. Creatine supplementation in the aging population: Effects on skeletal muscle, bone and brain. Amino Acids 2016, 48, 1793–1805. [Google Scholar] [CrossRef]
- Troesch, B.; Eggersdorfer, M.; Laviano, A.; Rolland, Y.; Smith, A.D.; Warnke, I.; Weimann, A.; Calder, P.C. Expert opinion on benefits of long-chain omega-3 fatty acids (DHA and EPA) in aging and clinical nutrition. Nutrients 2020, 12, 2555. [Google Scholar] [CrossRef] [PubMed]
- Cercillieux, A.; Ciarlo, E.; Canto, C. Balancing NAD+ deficits with nicotinamide riboside: Therapeutic possibilities and limitations. Cell Mol. Life Sci. 2022, 79, 463. [Google Scholar] [CrossRef] [PubMed]
- Serino, A.; Salazar, G. Protective role of polyphenols against vascular inflammation, aging and cardiovascular disease. Nutrients 2018, 11, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, M.D.; Osburn, S.C.; Godwin, J.S.; Ruple, B.A.; La Monica, M.B.; Raub, B.; Sandrock, J.E.; Ziegenfuss, T.N.; Lopez, H.L. Enhance Trial: Effects of NAD3® on Hallmarks of Aging and Clinical Endpoints of Health in Middle Aged Adults: A Subset Analysis Focused on Blood Cell NAD+ Concentrations and Lipid Metabolism. Physiologia 2022, 2, 20–31. [Google Scholar]
- Taylor, L.; Mumford, P.; Roberts, M.; Hayward, S.; Mullins, J.; Urbina, S.; Wilborn, C. Safety of TeaCrine(R), a non-habituating, naturally-occurring purine alkaloid over eight weeks of continuous use. J. Int. Soc. Sports Nutr. 2016, 13, 2. [Google Scholar] [CrossRef] [Green Version]
- Mumford, P.W.; Osburn, S.C.; Fox, C.D.; Godwin, J.S.; Roberts, M.D. A Theacrine-Based Supplement Increases Cellular NAD+ Levels and Affects Biomarkers Related to Sirtuin Activity in C2C12 Muscle Cells In Vitro. Nutrients 2020, 12, 3727. [Google Scholar] [CrossRef]
- Sheng, Y.Y.; Xiang, J.; Wang, Z.S.; Jin, J.; Wang, Y.Q.; Li, Q.S.; Li, D.; Fang, Z.T.; Lu, J.L.; Ye, J.H.; et al. Theacrine From Camellia kucha and Its Health Beneficial Effects. Front. Nutr. 2020, 7, 596823. [Google Scholar] [CrossRef]
- Gao, M.; Zheng, J.; Zheng, C.; Huang, Z.; Huang, Q. Theacrine alleviates chronic inflammation by enhancing TGF-beta-mediated shifts via TGF-beta/SMAD pathway in Freund’s incomplete adjuvant-induced rats. Biochem. Biophys. Res. Commun. 2020, 522, 743–748. [Google Scholar] [CrossRef]
- Wang, G.E.; Li, Y.F.; Zhai, Y.J.; Gong, L.; Tian, J.Y.; Hong, M.; Yao, N.; Wu, Y.P.; Kurihara, H.; He, R.R. Theacrine protects against nonalcoholic fatty liver disease by regulating acylcarnitine metabolism. Metabolism 2018, 85, 227–239. [Google Scholar] [CrossRef]
- Li, W.X.; Li, Y.F.; Zhai, Y.J.; Chen, W.M.; Kurihara, H.; He, R.R. Theacrine, a purine alkaloid obtained from Camellia assamica var. kucha, attenuates restraint stress-provoked liver damage in mice. J. Agric. Food Chem. 2013, 61, 6328–6335. [Google Scholar] [CrossRef] [PubMed]
- Uto, T.; Hou, D.X.; Morinaga, O.; Shoyama, Y. Molecular Mechanisms Underlying Anti-Inflammatory Actions of 6-(Methylsulfinyl)hexyl Isothiocyanate Derived from Wasabi (Wasabia japonica). Adv. Pharmacol. Sci. 2012, 2012, 614046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greaney, A.J.; Maier, N.K.; Leppla, S.H.; Moayeri, M. Sulforaphane inhibits multiple inflammasomes through an Nrf2-independent mechanism. J. Leukoc. Biol. 2016, 99, 189–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caglayan, B.; Kilic, E.; Dalay, A.; Altunay, S.; Tuzcu, M.; Erten, F.; Orhan, C.; Gunal, M.Y.; Yulug, B.; Juturu, V.; et al. Allyl isothiocyanate attenuates oxidative stress and inflammation by modulating Nrf2/HO-1 and NF-kappaB pathways in traumatic brain injury in mice. Mol. Biol. Rep. 2019, 46, 241–250. [Google Scholar] [CrossRef]
- Gerhauser, C. Epigenetic impact of dietary isothiocyanates in cancer chemoprevention. Curr. Opin. Clin. Nutr. Metab. Care 2013, 16, 405–410. [Google Scholar] [CrossRef]
- Arredondo, M.; Munoz, P.; Mura, C.V.; Nunez, M.T. DMT1, a physiologically relevant apical Cu1+ transporter of intestinal cells. Am. J. Physiol. Cell Physiol. 2003, 284, C1525–C1530. [Google Scholar] [CrossRef]
- Thomas, P.D.; Campbell, M.J.; Kejariwal, A.; Mi, H.; Karlak, B.; Daverman, R.; Diemer, K.; Muruganujan, A.; Narechania, A. PANTHER: A library of protein families and subfamilies indexed by function. Genome Res. 2003, 13, 2129–2141. [Google Scholar] [CrossRef] [Green Version]
- Sexton, C.L.; Godwin, J.S.; McIntosh, M.C.; Ruple, B.A.; Osburn, S.C.; Hollingsworth, B.R.; Kontos, N.J.; Agostinelli, P.J.; Kavazis, A.N.; Ziegenfuss, T.N. Skeletal muscle DNA methylation and mRNA responses to a bout of higher versus lower load resistance exercise in previously trained men. Cells 2023, 12, 263. [Google Scholar] [CrossRef]
- Joglekar, M.V.; Satoor, S.N.; Wong, W.K.; Cheng, F.; Ma, R.C.; Hardikar, A.A. An optimised step-by-step protocol for measuring relative telomere length. Methods Protoc. 2020, 3, 27. [Google Scholar] [CrossRef] [Green Version]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Verdin, E. NAD+ in aging, metabolism, and neurodegeneration. Science 2015, 350, 1208–1213. [Google Scholar] [CrossRef]
- Goody, M.F.; Henry, C.A. A need for NAD+ in muscle development, homeostasis, and aging. Skelet. Muscle 2018, 8, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamb, D.A.; Moore, J.H.; Mesquita, P.H.C.; Smith, M.A.; Vann, C.G.; Osburn, S.C.; Fox, C.D.; Lopez, H.L.; Ziegenfuss, T.N.; Huggins, K.W.; et al. Resistance training increases muscle NAD+ and NADH concentrations as well as NAMPT protein levels and global sirtuin activity in middle-aged, overweight, untrained individuals. Aging 2020, 12, 9447–9460. [Google Scholar] [CrossRef] [PubMed]
- Sawarkar, R. Transcriptional lockdown during acute proteotoxic stress. Trends Biochem. Sci. 2022. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Dou, X.; Li, S.; Zhang, X.; Zeng, Y.; Song, Z. Nicotinamide ameliorates palmitate-induced ER stress in hepatocytes via cAMP/PKA/CREB pathway-dependent Sirt1 upregulation. Biochim. Biophys. Acta-Mol. Cell Res. 2015, 1853, 2929–2936. [Google Scholar] [CrossRef] [Green Version]
- Alexovic, M.; Lindner, J.R.; Bober, P.; Longuespee, R.; Sabo, J.; Davalieva, K. Human peripheral blood mononuclear cells: A review of recent proteomic applications. Proteomics 2022, 22, e2200026. [Google Scholar] [CrossRef]
- Farber, D.L.; Yudanin, N.A.; Restifo, N.P. Human memory T cells: Generation, compartmentalization and homeostasis. Nat. Rev. Immunol. 2014, 14, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Fulcher, D.A.; Basten, A. B cell life span: A review. Immunol. Cell Biol. 1997, 75, 446–455. [Google Scholar] [CrossRef]
- Patel, A.A.; Ginhoux, F.; Yona, S. Monocytes, macrophages, dendritic cells and neutrophils: An update on lifespan kinetics in health and disease. Immunology 2021, 163, 250–261. [Google Scholar] [CrossRef]
- Gu, J.; Xia, X.; Yan, P.; Liu, H.; Podust, V.N.; Reynolds, A.B.; Fanning, E. Cell cycle-dependent regulation of a human DNA helicase that localizes in DNA damage foci. Mol. Biol. Cell 2004, 15, 3320–3332. [Google Scholar] [CrossRef]
- Kleiveland, C.R. Peripheral blood mononuclear cells. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Springer: Berlin/Heidelberg, Germany, 2015; pp. 161–167. [Google Scholar]



{kind=link}
{kind=link}
| Gene | CTL PRE | CTL POST | NAD3 PRE | NAD3 POST | Delta-Fold Change | Interaction p-Value |
|---|---|---|---|---|---|---|
| Downregulated in NAD3 versus CTL from pre-to post-testing | ||||||
| ADAM22 | 5.66 | 6.09 | 6.73 * | 5.27 | −3.70 | 0.0063 |
| PTPRO | 4.97 | 5.34 | 6.25 * | 5.07 | −2.94 | 0.0089 |
| DCAF6 | 5.69 | 6.2 | 6.26 | 5.21 | −2.94 | 0.0094 |
| SRF | 4.18 | 4.66 | 5.13 * | 4.06 # | −2.93 | 0.0004 |
| LYRM2 | 4.89 | 5.05 | 6.08 * | 4.80 | −2.72 | 0.0011 |
| RRAS2 | 6.12 | 6.80 | 6.55 * | 5.84 # | −2.63 | 0.00001 |
| ZNF415 | 4.65 | 5.09 | 5.12 | 4.21 # | −2.55 | 0.002 |
| SNX1 | 4.91 | 5.42 | 5.62 * | 4.79 # | −2.53 | 0.0003 |
| THEGL | 6.61 | 7.09 | 7.23 | 6.39 # | −2.51 | 0.0086 |
| ECD | 5.93 | 6.62 | 6.26 | 5.64 # | −2.49 | 0.0086 |
| PPP6R1 | 4.62 | 4.89 | 5.29 * | 4.30 # | −2.40 | 0.0007 |
| FBXO16 | 6.08 | 6.62 | 6.80 * | 6.08 | −2.39 | 0.0017 |
| TNFRSF10B | 4.82 | 5.52 | 5.57 * | 5.02 | −2.38 | 0.0016 |
| EPT1 | 4.63 | 5.06 | 5.12 * | 4.31 | −2.36 | 0.0044 |
| GEMIN2 | 5.16 | 5.67 | 5.81 * | 5.12 # | −2.31 | 0.0004 |
| Upregulated in NAD3 versus CTL from pre- to post-testing | ||||||
| TNFSF13B | 6.97 | 5.23 | 5.10 * | 5.94 | 5.99 | 0.00006 |
| ST5 | 5.96 | 5.50 | 4.79 * | 6.25 # | 3.79 | 0.0022 |
| NDUFB4 | 6.53 | 5.54 | 5.72 | 6.57 | 3.59 | 0.0017 |
| ZZZ3 | 5.04 | 4.82 | 4.21 * | 5.66 # | 3.18 | 0.0009 |
| TRPC5 | 6.62 | 5.83 | 6.24 | 7.09 # | 3.10 | 0.0015 |
| OSBPL1A | 6.40 | 5.78 | 5.30 * | 6.31 | 3.09 | 0.00008 |
| STKLD1 | 7.00 | 6.19 | 6.20 * | 6.94 # | 2.92 | 0.0002 |
| POGZ | 6.99 | 5.86 | 5.96 * | 6.37 # | 2.92 | 0.0044 |
| CNTROB | 7.29 | 6.05 | 6.34 * | 6.65 | 2.90 | 0.0094 |
| MYO6 | 5.05 | 4.29 | 4.56 | 5.32 # | 2.87 | 0.0021 |
| POT1 | 6.26 | 5.71 | 5.48 | 6.40 # | 2.78 | 0.0006 |
| PPIE | 5.01 | 4.61 | 4.33 | 5.41 # | 2.78 | 0.0023 |
| ARMC2 | 4.27 | 3.88 | 4.00 | 5.01 # | 2.64 | 0.0067 |
| STXBP4 | 6.01 | 5.79 | 5.63 | 6.68 # | 2.41 | 0.0067 |
| CNIH3 | 5.32 | 5.06 | 5.21 | 6.21 # | 2.40 | 0.0083 |
| Biological Process GOTERM | Count (% of Pathway) | mRNAs | Significance of GOTERM |
|---|---|---|---|
| Upregulated in NAD3 versus CTL from pre- to post-testing | |||
| Helicase activity | 3 (4%) | DDX60, HFM1, SNRNP200 | 0.021 |
| Downregulated in NAD3 versus CTL from pre- to post-testing | |||
| Regulation of transcription from RNA PolII promoter | 10 (14.3%) | DCAF6, PRDM4, ECD, FOXOA1, GMEB1, IL17F, MOSPD1, NPAT, PHOX2B, SRF | 0.012 |
| Response to ER stress | 3 (4.3%) | DNAJB9, TNFRSF10B, HYOU1 | 0.028 |
| mRNA processing | 4 (5.7%) | ADAR, ECD, GEMIN2, PAN3 | 0.029 |
| Gene | CTL PRE | CTL POST | NAD3 PRE | NAD3 POST | Delta-Fold Change | Interaction p-Value |
|---|---|---|---|---|---|---|
| Downregulated in NAD3 versus CTL from pre- to post-testing | ||||||
| ZNF415 | 4.65 | 5.09 | 5.12 | 4.21 | −2.55 | 0.002 |
| THEGL | 6.61 | 7.09 | 7.23 | 6.39 | −2.51 | 0.0086 |
| ECD | 5.93 | 6.62 | 6.26 | 5.64 | −2.49 | 0.0086 |
| SPATA17 | 4.83 | 5.71 | 4.83 | 4.53 | −2.27 | 0.0001 |
| ZNF529-AS1 | 5.75 | 6.17 | 6.35 | 5.74 | −2.04 | 0.003 |
| ST5 | 6.2 | 6.61 | 6.43 | 5.84 | −2.01 | 0.0074 |
| OXR1 | 5.24 | 5.60 | 5.38 | 4.73 | −2.01 | 0.0013 |
| HINT1 | 6.08 | 6.49 | 6.52 | 5.96 | −1.96 | 0.0029 |
| ZNF233 | 4.86 | 5.59 | 5.18 | 4.94 | −1.96 | 0.009 |
| GLCCI1 | 4.47 | 4.98 | 4.58 | 4.12 | −1.95 | 0.0034 |
| DTD1 | 5.93 | 6.47 | 6.26 | 5.86 | −1.92 | 0.0027 |
| MOSPD1 | 4.65 | 4.76 | 5.06 | 4.28 | −1.87 | 0.0071 |
| DDX11 | 5.84 | 6.54 | 5.71 | 5.51 | −1.87 | 0.0042 |
| C14orf159 | 4.19 | 4.67 | 4.40 | 4.00 | −1.83 | 0.0028 |
| FOXA1 | 5.51 | 6.11 | 5.85 | 5.60 | −1.80 | 0.0025 |
| Upregulated in NAD3 versus CTL from pre- to post-testing | ||||||
| ST5 | 5.96 | 5.5 | 4.79 | 6.25 | 3.79 | 0.0022 |
| NDUFB4 | 6.53 | 5.54 | 5.72 | 6.57 | 3.59 | 0.0017 |
| TRPC5 | 6.62 | 5.83 | 6.24 | 7.09 | 3.10 | 0.0015 |
| MYO6 | 5.05 | 4.29 | 4.56 | 5.32 | 2.87 | 0.0021 |
| PPIE | 5.01 | 4.61 | 4.33 | 5.41 | 2.78 | 0.0023 |
| POT1 | 6.26 | 5.71 | 5.48 | 6.40 | 2.78 | 0.0006 |
| ARMC2 | 4.27 | 3.88 | 4.00 | 5.01 | 2.64 | 0.0067 |
| STXBP4 | 6.01 | 5.79 | 5.63 | 6.68 | 2.41 | 0.0067 |
| CNIH3 | 5.32 | 5.06 | 5.21 | 6.21 | 2.40 | 0.0083 |
| WRNIP1 | 4.94 | 4.48 | 4.32 | 5.11 | 2.37 | 0.0057 |
| SASS6 | 5.27 | 4.80 | 5.42 | 6.17 | 2.34 | 0.0082 |
| SLC26A6 | 6.02 | 5.25 | 5.50 | 5.91 | 2.29 | 0.0096 |
| ASB17 | 5.07 | 4.82 | 4.53 | 5.48 | 2.29 | 0.0047 |
| LNX1 | 4.24 | 4.24 | 4.10 | 5.17 | 2.11 | 0.0059 |
| C3orf52 | 5.04 | 4.82 | 4.67 | 5.47 | 2.03 | 0.0054 |
| Biological Process GOTERM | Count (% of Pathway) | mRNAs | Significance of GOTERM |
|---|---|---|---|
| Top upregulated biological processes in NAD3 versus CTL from pre- to post-testing | |||
| DNA conformation change * | 5 (4.7%) | POT1, HFM1, WRNIP1, DTD1, DDX11 | 1.4 × 10−5 |
| Only downregulated biological process in NAD3 versus CTL from pre- to post-testing | |||
| Cell communication | 8 (0.1%) | RAB12, STXBP4, ASB17, MYO6, CD24, HINT1, TEAD2, FOXA1 | 0.047 |
| Differentially Methylated Sites | CTL Group | NAD3 Group |
|---|---|---|
| Hypermethylated | 369 | 35 |
| Hypomethylated | 178 | 72 |
| Total differentially methylated sites | 547 | 107 |
| Classification | ID | Description | Genes |
|---|---|---|---|
| GO-related terms predicted to be upregulated based on genes hypomethylated at week 12 in the NAD3 versus CTL group | |||
| GO Molecular Function | GO:0003712 | transcription cofactor activity | BCOR,CITED1,FOXP3,HCFC1,KDM1A,MAGED1,MECP2,MED12,MED14,MSX2,MYOCD,NR0B1,NR2F2,PIR,POU3F2,PQBP1,RAP2C,RBBP8,RLIM,SMARCC1,SOX3,SSX4B,TADA2A,TAF1,TAF7L,TAF9B,TBL1X,TFDP1,UBE3A,UXT,VGLL1,YY1,ZCCHC12,ZCCHC18 |
| GO Molecular Function | GO:0017064 | fatty acid amide hydrolase activity | FAAH,FAAH2 |
| GO Cellular Component | GO:0000794 | condensed nuclear chromosome | ATRX,FAM9B,FAM9C,HSPA2,RGS12,SMC1A,SUV39H1,SYN1,TEX11 |
| GO Molecular Function | GO:0005275 | amine transmembrane transporter activity | SLC22A16,SLC32A1 |
| GO Molecular Function | GO:0052846 | inositol-1,5-bisdiphosphate-2,3,4,6-tetrakisphosphate 1-diphosphatase activity | NUDT10,NUDT11 |
| GO Molecular Function | GO:0016300 | tRNA (uracil) methyltransferase activity | KIAA1456,TRMT2B |
| GO-related terms predicted to be downregulated based on genes hypermethylated at week 12 in the NAD3 versus CTL group | |||
| GO Molecular Function | GO:0008484 | sulfuric ester hydrolase activity | ARSD,ARSE,ARSF,ARSH,ENSG00000241489,IDS,STS,SULF1 |
| GO Biological Process | GO:0002755 | MyD88-dependent toll-like receptor signaling pathway | BTK,IRAK1,TAB2,TAB3,TLR7,TLR8 |
| GO Biological Process | GO:0008215 | spermine metabolic process | SAT1,SATL1,SMS |
| GO Cellular Component | GO:0002189 | ribose phosphate diphosphokinase complex | PRPS1,PRPS2 |
| GO Molecular Function | GO:0048365 | Rac GTPase binding | ARHGAP4,CDKL5,FLNA,NOX1,OCRL,PAK3,WAS |
| GO Molecular Function | GO:0035586 | purinergic receptor activity | ADORA2A,GPR34,P2RX2,P2RY10,P2RY4 |
| GO Biological Process | GO:0016570 | histone modification | ATXN3L,BCOR,BRCC3,CUL4B,HCFC1,HDAC6,HDAC8,HUWE1,JADE3,KDM5C,KDM6A,MECP2,MORF4L2,MSL3,OGT,PADI1,PADI3,PHF8,RBBP7,SUV39H1,TAF1,TAF9B,TBL1X,UBE2A,USP51 |
| GO Molecular Function | GO:0035197 | siRNA binding | FMR1,MECP2,TLR7 |
| GO Molecular Function | GO:0061578 | Lys63-specific deubiquitinase activity | BRCC3,OTUD5,USP27X |
| GO Biological Process | GO:0071394 | cellular response to testosterone stimulus | AR,ELK1,MSN |
| GO Biological Process | GO:0009169 | purine ribonucleoside monophosphate catabolic process | HPRT1,NUDT10,NUDT11 |
| GO Cellular Component | GO:0005741 | mitochondrial outer membrane | ACSL4,ARMCX1,ARMCX2,ARMCX3,ARMCX6,DDX3X,FUNDC1,FUNDC2,GK,MAOA,MAOB,MLXIP,MTX2,SPATA19 |
| GO Molecular Function | GO:0008013 | beta-catenin binding | AJAP1,AMER1,AR,CDH5,FOXO4,HDAC6,MED12,SHROOM2,TBL1X |
| GO Biological Process | GO:0048488 | synaptic vesicle endocytosis | NLGN3,NLGN4X,OPHN1,SYP,SYT5 |
| GO Molecular Function | GO:0004145 | diamine N-acetyltransferase activity | SAT1,SATL1 |
| GO Biological Process | GO:0009128 | purine nucleoside monophosphate catabolic process | HPRT1,NUDT10,NUDT11 |
| GO Molecular Function | GO:0036459 | thiol-dependent ubiquitinyl hydrolase activity | ALG13,ATXN3L,BRCC3,OTUD5,OTUD6A,TAF9B,USP11,USP27X,USP51,USP9X,USP9Y |
| GO Biological Process | GO:0016569 | covalent chromatin modification | ATRX,ATXN3L,BCOR,BCORL1,BRCC3,CUL4B,HCFC1,HDAC6,HDAC8,HMGN5,HUWE1,JADE3,KDM5C,KDM6A,MECP2,MORF4L2,MSL3,OGT,PADI1,PADI3,PHF8,RBBP7,SMARCA1,SUV39H1,TAF1,TAF9B,TBL1X,TSPYL2,UBE2A,USP51 |
| GO Molecular Function | GO:0101005 | ubiquitinyl hydrolase activity | ALG13,ATXN3L,BRCC3,OTUD5,OTUD6A,TAF9B,USP11,USP27X,USP51,USP9X,USP9Y |
| GO Molecular Function | GO:0099095 | ligand-gated anion channel activity | GABRE,GLRA2,GLRA4 |
| Assay | CTL Group | NAD3 Group |
|---|---|---|
| Transcriptomics | N = 16 (51 ± 6 years old; 7 M/9 W) | N = 9 (53 ± 7 years old; 4 M/5 W) |
| Global DNA methylation | N = 13 (52 ± 6 years old; 7 M/6 W) | N = 8 (53 ± 8 years old; 3 M/5 W) |
| SIRT activity | N = 15 (52 ± 5 years old; 6 M/9 W) | N = 10 (51 ± 6 years old; 3 M/7 W) |
| Telomere length | N = 14 (51 ± 5 years old; 4 M/10 W) | N = 13 (50 ± 6 years old; 3 M/10 W) |
| NAD+: NADH * | N = 10 (51 ± 6 years old; 2 M/8 W) | N = 10 (52 ± 6 years old; 5 M/5 W) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roberts, M.D.; La Monica, M.B.; Raub, B.; Sandrock, J.E.; Ziegenfuss, T.N.; Smith, R.; Dwaraka, V.B.; Lopez, H.L. The Effects of a Multi-Ingredient Supplement Containing Wasabia Japonica Extract, Theacrine, and Copper (I) Niacin Chelate on Peripheral Blood Mononuclear Cell DNA Methylation, Transcriptomics, and Sirtuin Activity. Physiologia 2023, 3, 233-246. https://doi.org/10.3390/physiologia3020016
Roberts MD, La Monica MB, Raub B, Sandrock JE, Ziegenfuss TN, Smith R, Dwaraka VB, Lopez HL. The Effects of a Multi-Ingredient Supplement Containing Wasabia Japonica Extract, Theacrine, and Copper (I) Niacin Chelate on Peripheral Blood Mononuclear Cell DNA Methylation, Transcriptomics, and Sirtuin Activity. Physiologia. 2023; 3(2):233-246. https://doi.org/10.3390/physiologia3020016
Chicago/Turabian StyleRoberts, Michael D., Michael B. La Monica, Betsy Raub, Jennifer E. Sandrock, Tim N. Ziegenfuss, Ryan Smith, Varun B. Dwaraka, and Hector L. Lopez. 2023. "The Effects of a Multi-Ingredient Supplement Containing Wasabia Japonica Extract, Theacrine, and Copper (I) Niacin Chelate on Peripheral Blood Mononuclear Cell DNA Methylation, Transcriptomics, and Sirtuin Activity" Physiologia 3, no. 2: 233-246. https://doi.org/10.3390/physiologia3020016