
Curcumin Epigenetically Represses Histone Acetylation of Echinocandin B Producing Emericella rugulosa


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
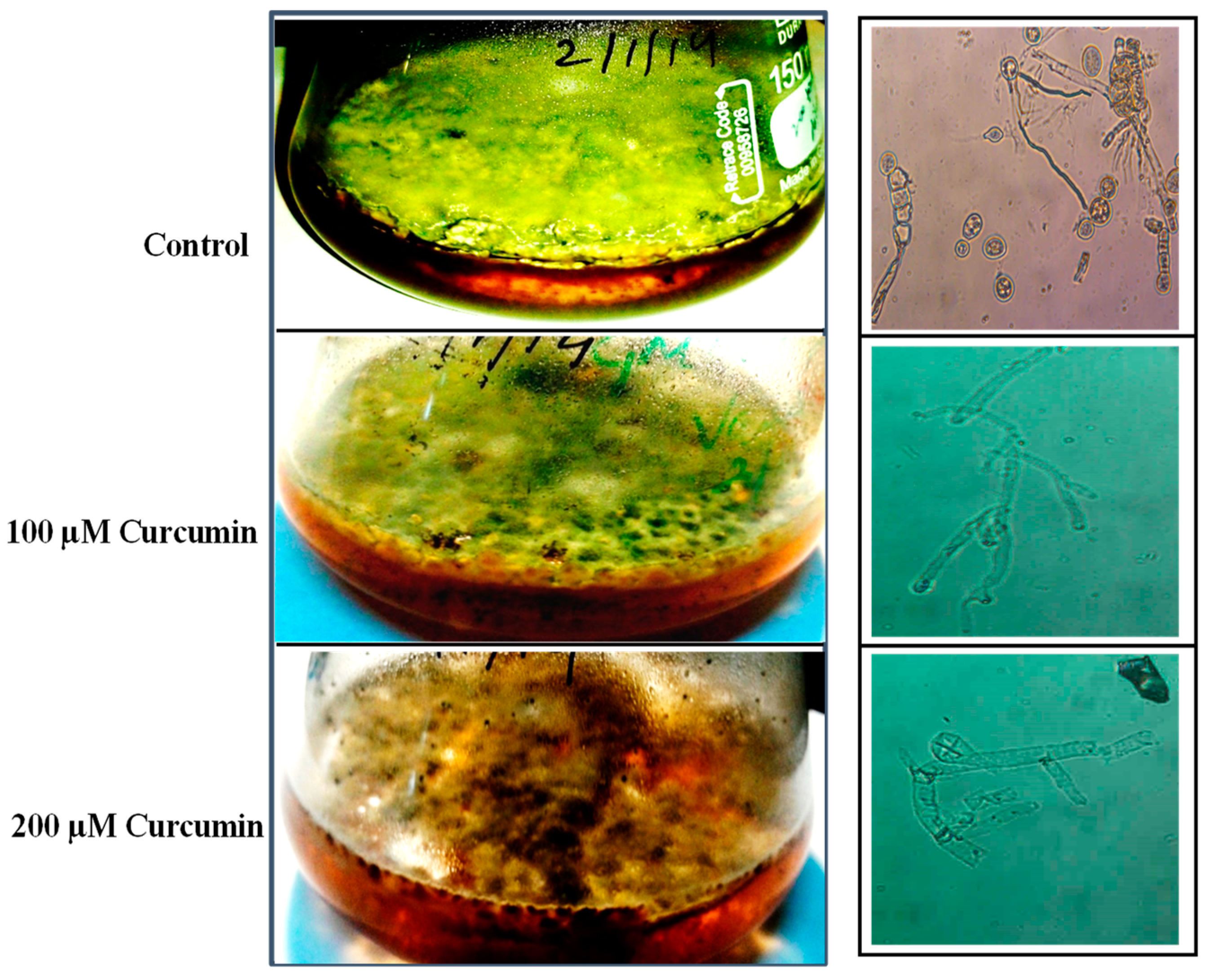
2.1. Effect of Curcumin on E. rugulosa Growth
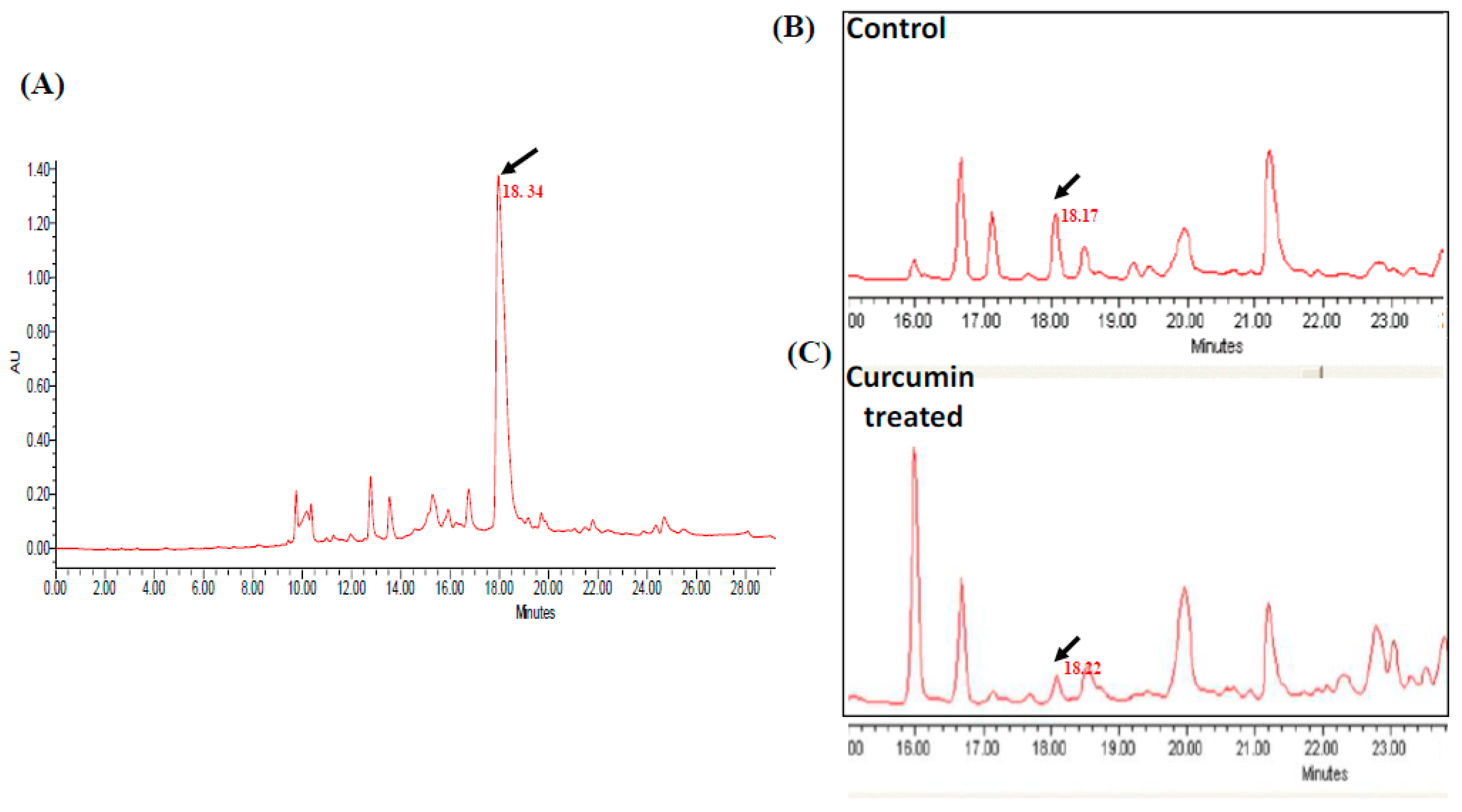
2.2. Effect of Curcumin on Echinocandin B Production
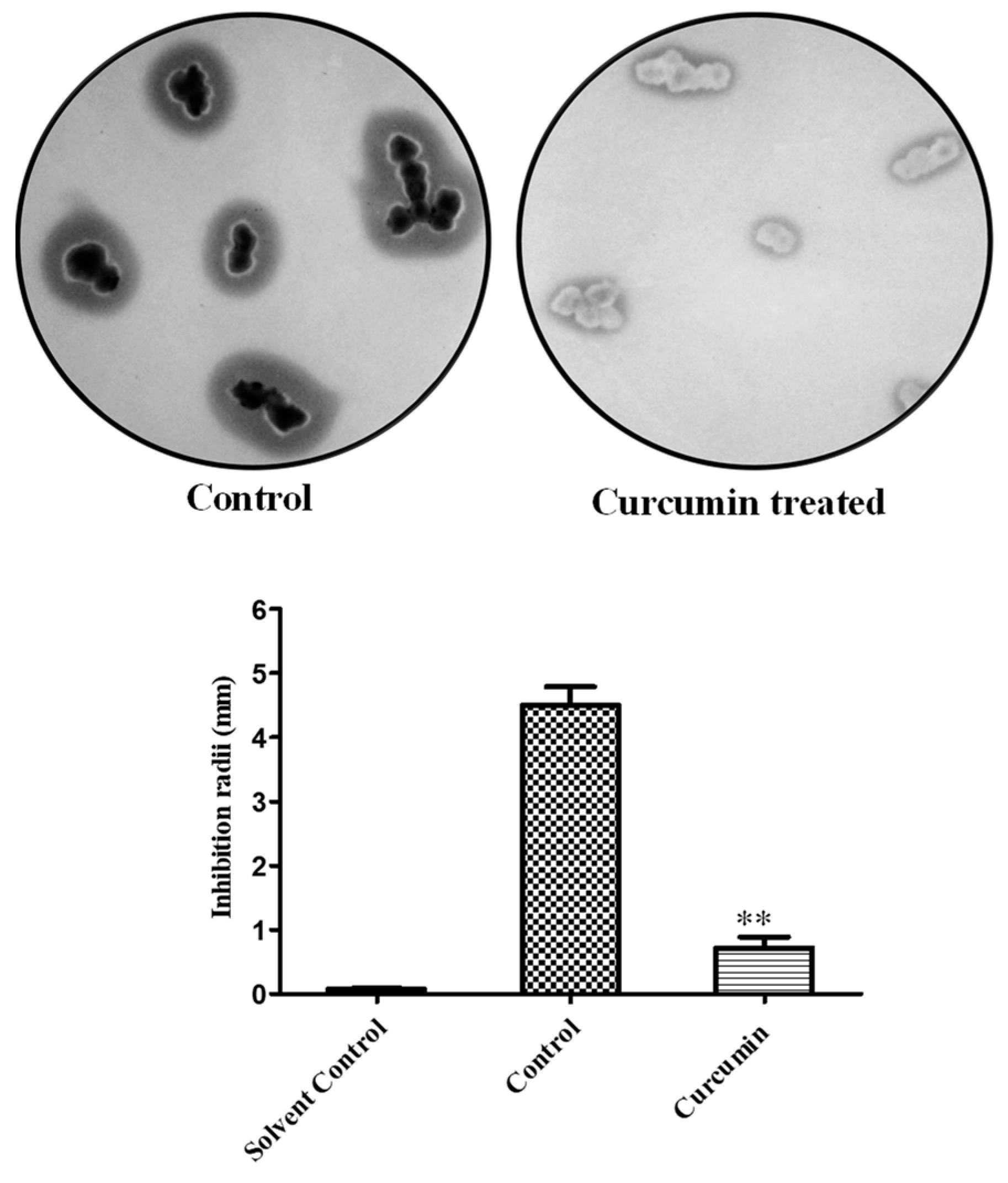
2.3. Cell Susceptibility Profiling of Curcumin-Treated Sample
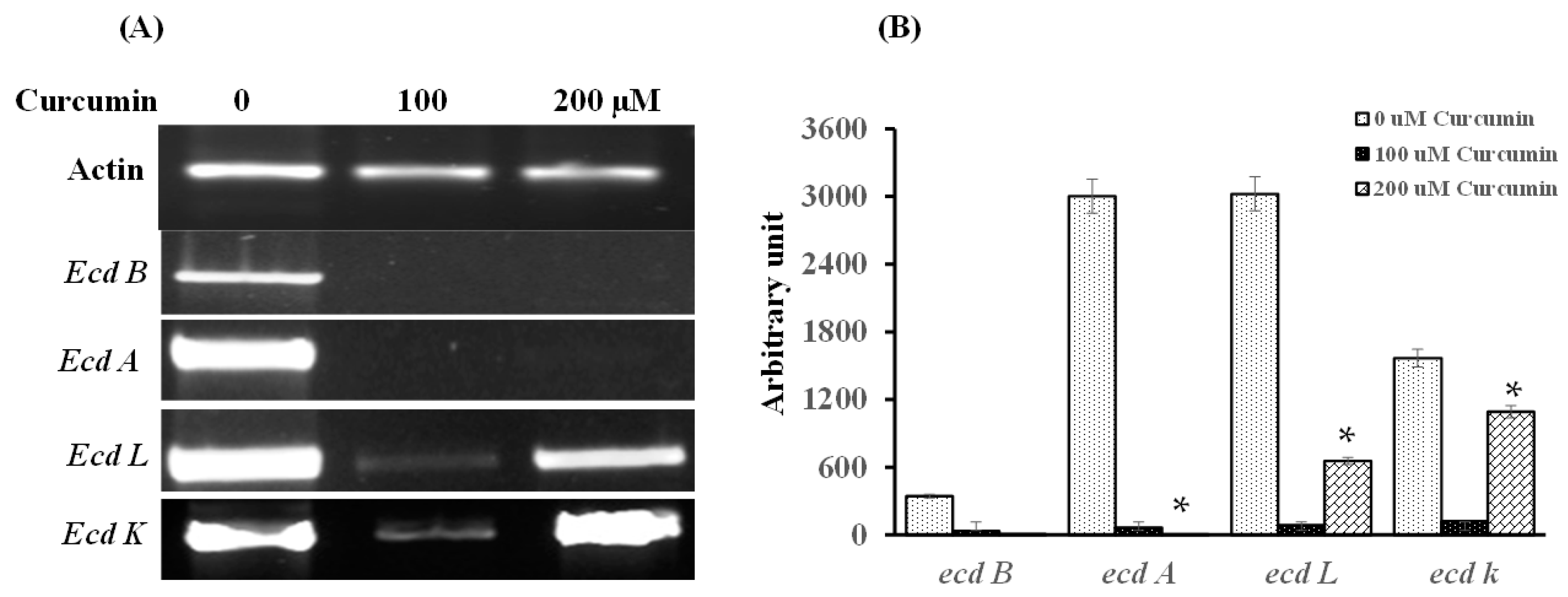
2.4. Curcumin Transcriptionally Represses Genes Present in the Echinocandin B Gene Cluster
2.5. Curcumin Transcriptionally Represses Genes Present in the Echinocandin B Gene Cluster
2.6. Interspecies Interactions Suppress the Inhibitory Effect of Curcumin on Echinocandin B Production
3. Discussion
4. Materials and Methods
4.1. Chemicals, Strains, and Media
4.2. Production of Echinocandin B
4.3. Measurement of Echinocandin B Production
4.4. Confrontation Assay
4.5. RNA Isolation
4.6. cDNA Synthesis
4.7. Transcription Gene Expression Profiling
4.8. E. rugulosa and C. albicans Interspecies Interactions
4.9. Crude Membrane Preparation
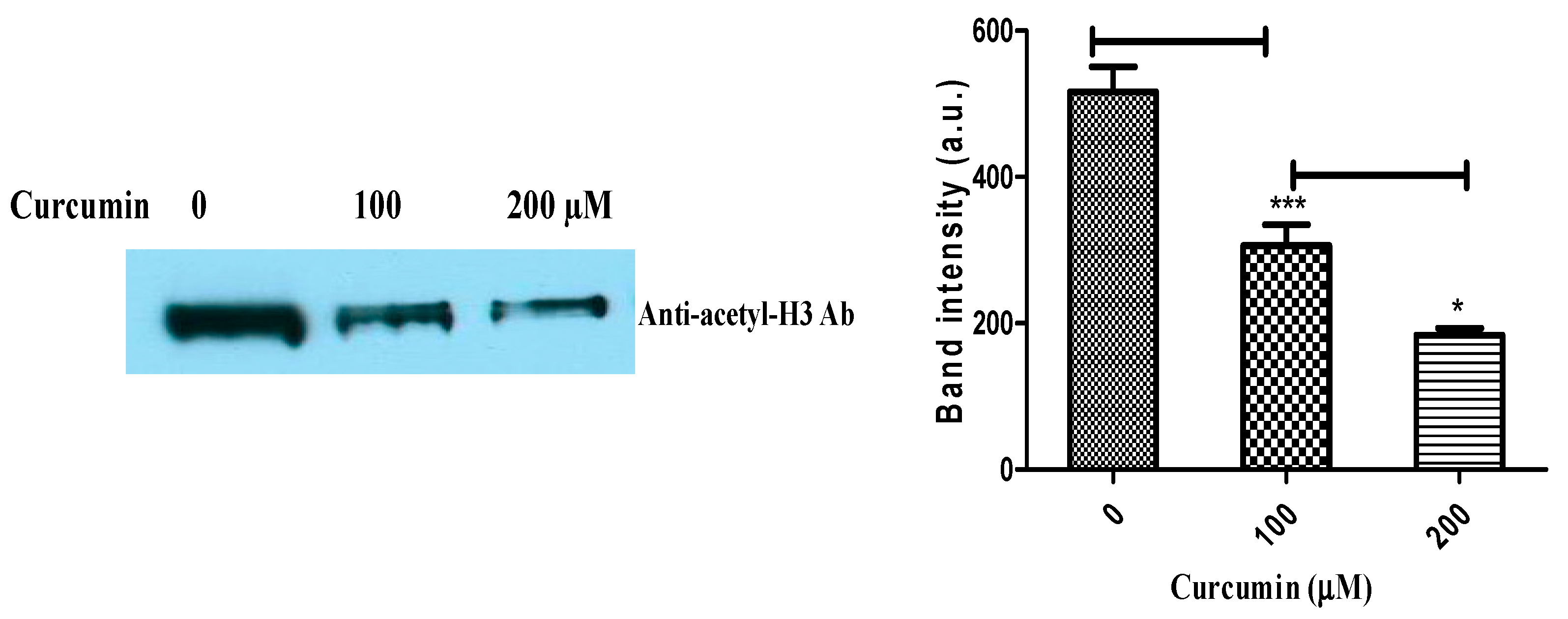
4.10. Immunodetection of Histone Acetylation
4.11. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Szymański, M.; Chmielewska, S.; Czyżewska, U.; Malinowska, M.; Tylicki, A. Echinocandins-structure, mechanism of action and use in antifungal therapy. J. Enzym. Inhib. Med. Chem. 2022, 37, 876–894. [Google Scholar] [CrossRef]
- Patil, A.; Majumdar, S. Echinocandins in antifungal pharmacotherapy. J. Pharm. Pharmacol. 2017, 69, 1635–1660. [Google Scholar] [CrossRef] [Green Version]
- Fisher, M.C.; Alastruey-Izquierdo, A.; Berman; Bicanic, T.; Bignell, E.M.; Bowyer, P.; Bromley, M.; Brüggemann, R.; Garber, G.; Cornely, O.A.; et al. Tackling the emerging threat of antifungal resistance to human health. Nat. Rev. Microbiol. 2022, 20, 557–571. [Google Scholar] [CrossRef] [PubMed]
- Cacho, R.A.; Jiang, W.; Chooi, Y.H.; Walsh, C.T.; Tang, Y. Identification and characterization of the echinocandin B biosynthetic gene cluster from Emericella rugulosa NRRL 11440. J. Am. Chem. Soc. 2012, 134, 16781–16790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Jaiswal, V.; Kumar, V.; Ray, A.; Kumar, A. Functional redundancy in Echinocandin B in-cluster transcription factor ecdB of Emericella rugulosa NRRL 11440. Biotechnol. Rep. 2018, 19, e00264. [Google Scholar] [CrossRef]
- Bera, K.; Rani, P.; Kishor, G.; Agarwal, S.; Kumar, A.; Singh, D.V. Structural elucidation of TMD0 Domain of EcdL: An MRP1 family of ABC transporter protein revealed by atomistic simulation. J. Biomol. Struct. Dyn. 2017, 36, 2938–2950. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Kumar, V.; Kumar, A. Functional characterization of host toxic EcdB transcription factor protein of echinocandin B biosynthetic gene cluster. Biotechnol. App. Biochem. 2019, 66, 626–633. [Google Scholar] [CrossRef]
- Hoffmeister, D.; Keller, N.P. Natural products of filamentous fungi: Enzymes, genes, and their regulation. Nat. Prod. Rep. 2007, 24, 393–416. [Google Scholar] [CrossRef]
- Yu, J.H.; Keller, N. Regulation of Secondary Metabolism in Filamentous Fungi. Ann. Rev. Phytopathol. 2005, 43, 437–458. [Google Scholar] [CrossRef]
- Collemare, J.; Seidl, M.F. Chromatin-dependent regulation of secondary metabolite biosynthesis in fungi: Is the picture complete? FEMS Microbiol. Rev. 2019, 43, 591–607. [Google Scholar] [CrossRef] [Green Version]
- Strauss, J.; Reyes-Dominguez, Y. Regulation of secondary metabolism by chromatin structure and epigenetic codes. Fungal Genet. Biol. 2011, 48, 62–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brakhage, A. Regulation of fungal secondary metabolism. Nat. Rev. Microbiol. 2013, 11, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Saavedra, F.; Marty-Lombardi, S.; Loyola, A. Characterization of Posttranslational Modifications on Histone Variants. Methods Mol. Biol. 2018, 1832, 21–49. [Google Scholar]
- Kim, J.; Lee, J.E.; Lee, J.S. Histone deacetylase-mediated morphological transition in Candida albicans. J. Microbiol. 2015, 53, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Wu, M.; Ma, X.; Huang, W.; Xu, Y. Function and mechanism of novel histone posttranslational modifications in health and disease. Biomed. Res. Int. 2021, 2021, 6635225. [Google Scholar] [CrossRef]
- Lopez, C.; Barnon, M.T.; Beacon, T.H.; Nardocci, G.; Davie, J.R. The key role of differential broad H3K4me3 and H3K4ac domains in breast cancer. Gene 2022, 826, 146463. [Google Scholar] [CrossRef]
- Jagganath, G. Turmeric, The Golden Spice: From Asia to Africa. Open Access J Arch Anthropol. 2020, 2, 1–6. [Google Scholar] [CrossRef]
- Joe, B.; Vijaykumar, M.; Lokesh, B.R. Biological Properties of Curcumin-Cellular and Molecular Mechanisms of Action. Crit. Rev. Food Sci. Nutr. 2004, 44, 97–111. [Google Scholar] [CrossRef]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A Review of Its Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef] [Green Version]
- Ojo, O.A.; Adeyemo, T.R.; Rotimi, D.; Batiha, G.E.; Mostafa-Hedeab, G.; Iyobhebhe, M.E.; Elebiyo, T.C.; Atunwa, B.; Adebola Busola Ojo, A.B.; Lima, C.M.G.; et al. Anticancer Properties of Curcumin Against Colorectal Cancer: A Review. Front. Oncol. 2022, 12, 881641. [Google Scholar] [CrossRef]
- Tanvir, E.M.; Hossen, S.; Hossain, F.; Afroz, R.; Gan, H.S.; Khalil, I.; Karim, N. Antioxidant Properties of Popular Turmeric (Curcuma longa) Varieties from Bangladesh. J. Food Qual. 2017, 2017, 8471785. [Google Scholar] [CrossRef] [Green Version]
- Jennings, M.R.; Parks, R.J. Curcumin as an Antiviral Agent. Viruses 2020, 12, 1242. [Google Scholar] [CrossRef] [PubMed]
- Hassan, F.; Saif-ur Rehman, M.; Khan, M.S.; Ali, M.A.; Javed, A.; Yang, A.C.N. Curcumin as an Alternative Epigenetic Modulator: Mechanism of Action and Potential Effects. Front. Genet. 2019, 10, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunagawa, Y.; Funamoto, M.; Shimizu, K.; Shimizu, S.; Sari, N.; Katanasaka, Y.; Miyazaki, Y.; Kakeya, H.; Hasegawa, K.; Morimoto, T. Curcumin, an Inhibitor of p300-HAT Activity, Suppresses the Development of Hypertension-Induced Left Ventricular Hypertrophy with Preserved Ejection Fraction in Dahl Rats. Nutrients 2021, 13, 2608. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Song, Z.; Wang, E.; Dong, L.; Bai, J.; Wang, D.; Zhu, J.; Zhang, C. Potential antifungal targets based on histones post-translational modifications against invasive aspergillosis. Front. Microbiol. 2022, 13, 980615. [Google Scholar] [CrossRef] [PubMed]
- Nutzmann, H.W.; Reyes-Dominguez, Y.; Scherlach, K.; Schroeckh, V.; Horn, F.; Gacek, A.; Schümann, J.; Hertweck, C.; Strauss, J.; Brakhage, A.A. Bacteria induced natural product formation in the fungus Aspergillus nidulans requires Saga/Ada-mediated histone acetylation. Proc. Natl. Acad. Sci. USA 2011, 108, 14282–14287. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Xu, C.; Sun, Q.; Xu, J.; Chai, Y.; Berg, G.; Cernava, T.; Ma, Z.; Chen, Y. Post-translational regulation of autophagy is involved in intra-microbiome suppression of fungal pathogens. Microbiome 2021, 9, 131. [Google Scholar] [CrossRef]
- Wang, J.; Cacho, R.A.; Chiou, G.; Garg, N.K.; Tang, Y.; Walsh, C.T. EcdGHK are Three Tailoring Iron Oxygenases for Amino Acid Building Blocks of the Echinocandin Scaffold. J. Am. Chem. Soc. 2013, 135, 4457–4466. [Google Scholar]
- Guzman-Chavez, F.; Salo, O.; Samol, M.; Ries, M.; Kuipers, J.; Bovenberg, R.A.L.; Vreeken, R.J.; Driessen, A.J.M. Deregulation of secondary metabolism in a histone deacetylase mutant of Penicillium chrysogenum. Microbiologyopen 2018, 7, e00598. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Rodríguez, E.Y.; Uresti-Rivera, E.E.; Patrón-Soberano, O.A.; Islas-Osuna, M.A.; Flores-Martínez, A.; Riego-Ruiz, L.; Rosales-Saavedra, M.T.; Casas-Flores, S. Histone acetyltransferase TGF-1 regulates Trichoderma atroviride secondary metabolism and mycoparasitism. PLoS ONE 2018, 13, e0193872. [Google Scholar] [CrossRef] [Green Version]
- Ramesha, K.P.; Mohana, N.C.; Nayaka, S.C.; Satish, S. Epigenetic Modifiers Revamp Secondary Metabolite Production in Endophytic Nigrospora sphaerica. Front. Microbiol. 2021, 21, 730355. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.; Sharma, V.K.; Singh, D.K.; Mishra, A.; Gond, S.K.; Verma, S.K.; Kumar, A.; Kharwar, R.N. Epigenetic Activation of Antibacterial Property of an Endophytic Streptomyces coelicolor Strain AZRA 37 and Identification of the Induced Protein Using MALDI TOF MS/MS. PLoS ONE 2016, 11, e0147876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Kumar, A.; Kaushal, M.; Kumar, P.; Mukhopadhyay, K.; Kumar, A. Fungal-derived xenobiotic exhibits antibacterial and antibiofilm activity against S. aureus. Drug Discov. Ther. 2018, 12, 214–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, A.; Kumar, A.; Singh, A.; Lynn, A.M.; Kapoor, K.; Prasad, R. A key structural domain of the Candida albicans Mdr1 protein. Biochem. J. 2012, 445, 313–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Shukla, S.; Mandal, A.; Shukla, S.; Ambudkar, S.; Prasad, R. Divergent Signature motifs of Nucleotide Binding Domains of ABC multidrug transporter, CaCdr1p of pathogenic Candida albicans, are functionally asymmetric and non-interchangeable. BBA Biomemb. 2010, 1798, 1757–1766. [Google Scholar] [CrossRef] [Green Version]
- Rai, V.; Gaur, M.; Kumar, A.; Shukla, S.; Komath, S.S.; Prasad, R. Novel catalytic mechanism for ATP hydrolysis employed by the N-terminal nucleotide-binding domain of Cdr1p, a multidrug ABC transporter of C. albicans. BBA Biomemb. 2008, 1778, 2143–2153. [Google Scholar] [CrossRef] [Green Version]
- Kumari, N.; Singh, S.; Kumari, V.; Kumar, S.; Kumar, V.; Kumar, A. Ouabain potentiates the antimicrobial activity of aminoglycosides against Staphylococcus aureus. BMC Complement. Altern. Med. 2019, 19, 119. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; Rosenzweig, R.; Asalla, A.; Nehra, S.; Prabhu, S.D.; Bansal, S.S. TNFR1 Contributes to Activation Induced Cell Death of Pathological Cd4+ T-Lymphocytes During Ischemic Heart Failure. JACC Basic Transl. Sci. 2022, 10, 1038–1049. [Google Scholar] [CrossRef]
- Irfan, M.; Kumar, P.; Kumar, V.; Datta, A. Fruit ripening specific expression of β-DN-acetylhexosaminidase (β-Hex) gene in tomato is transcriptionally regulated by Ethylene Response Factor SlERF. E4. Plant Sci. 2022, 323, 111380. [Google Scholar] [CrossRef]
- Kumar, V.; Irfan, M.; Ghosh, S.; Chakraborty, N.; Chakraborty, S.; Datta, A. Fruit ripening mutants reveal cell metabolism and redox state during ripening. Protoplasma 2016, 253, 581–594. [Google Scholar] [CrossRef]
- Irfan, M.; Ghosh, S.; Kumar, V.; Chakraborty, N.; Chakraborty, S.; Datta, A. Insights into transcriptional regulation of β-D-N-acetylhexosaminidase, an N-glycan-processing enzyme involved in ripening-associated fruit softening. J. Exp. Bot. 2014, 65, 5835–5848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.; Singh, U.K.; Meli, V.S.; Kumar, V.; Kumar, A.; Irfan, M.; Chakraborty, N.; Chakraborty, S.; Datta, A. Induction of senescence and identification of differentially expressed genes in tomato in response to monoterpene. PLoS ONE 2013, 8, e76029. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumari, V.; Kumar, V.; Kaushal, M.; Kumar, A. Curcumin Epigenetically Represses Histone Acetylation of Echinocandin B Producing Emericella rugulosa. Physiologia 2023, 3, 221-232. https://doi.org/10.3390/physiologia3020015
Kumari V, Kumar V, Kaushal M, Kumar A. Curcumin Epigenetically Represses Histone Acetylation of Echinocandin B Producing Emericella rugulosa. Physiologia. 2023; 3(2):221-232. https://doi.org/10.3390/physiologia3020015
Chicago/Turabian StyleKumari, Vandana, Vinay Kumar, Manisha Kaushal, and Antresh Kumar. 2023. "Curcumin Epigenetically Represses Histone Acetylation of Echinocandin B Producing Emericella rugulosa" Physiologia 3, no. 2: 221-232. https://doi.org/10.3390/physiologia3020015